
Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings




Department of Agricultural and Forest Sciences, Tuscia University, Via S. C. de Lellis, 01100 Viterbo, Italy
*
Author to whom correspondence should be addressed.
Plants 2023, 12(2), 412; https://doi.org/10.3390/plants12020412
Submission received: 21 December 2022
/
Revised: 3 January 2023
/
Accepted: 11 January 2023
/
Published: 16 January 2023
(This article belongs to the Special Issue Responses of Wheat to Abiotic Stress)
Abstract
:Soil salinity is a serious threat to food production now and in the near future. In this study, the root system of six durum wheat genotypes, including one highly salt-tolerant (J. Khetifa) used as a check genotype, was evaluated, by a high-throughput phenotyping system, under control and salt conditions at the seedling stage. Genotyping was performed using 11 SSR markers closely linked with genome regions associated with root traits. Based on phenotypic cluster analysis, genotypes were grouped differently under control and salt conditions. Under control conditions, genotypes were clustered mainly due to a root angle, while under salt stress, genotypes were grouped according to their capacity to maintain higher roots length, volume, and surface area, as J. Khetifa, Sebatel, and Azeghar. SSR analysis identified a total of 42 alleles, with an average of about three alleles per marker. Moreover, quite a high number of Private alleles in total, 18 were obtained. The UPGMA phenogram of the Nei (1972) genetic distance clusters for 11 SSR markers and all phenotypic data under control conditions discriminate genotypes almost into the same groups. The study revealed as the combination of high-throughput systems for phenotyping with SSR markers for genotyping it’s a useful tool to provide important data for the selection of suitable parental lines for salt-tolerance breeding. Nevertheless, the narrow root angle, which is an important trait in drought tolerance, is not a good indicator of salt tolerance. Instated for salt tolerance is more important the amount of roots.
1. Introduction
According to the FAO 2021 global map of salt-affected soils [1], around 400 million hectares of topsoil and more than 800 million hectares of subsoil are affected by salt. Moreover, FAO reported that soil salinity per year reduces production potential by taking up to 1.5 million ha of farmland out of production and more than 30 million USD as an annual loss in agricultural productivity [1]. Unfortunately, at such rates, 50% of total cultivated land is expected to be salinized by 2050 [2] as a consequence of the phenomenon of “desertification,” and land degradation due to climate change and human action by poor land management [3,4,5]. Projections up to 2099 assume an extreme increase in drought and the subsequent soil salt accumulation in Eurasia, Africa, and Australia [6]. Soil salinization may occur through a natural process (primary salinization) or by human management (secondary salinization), through irrigation with salty water, excessive use of fertilizers, and intensive agriculture practices [7,8,9].
Salt, mainly NaCl, in the soil slows down plant development due to two stresses. At first, the plant experiences a physiological drought (osmotic stress), which reduces leaves and root growth due to the inhibition of cell expansion, cell wall synthesis, and stomata conductance [10,11]. Later the plant response is related to the toxic effect of salt, where plants accumulate toxic Na+ and Cl− in old leaves over a long period. Due to the harmful effects of salt on germination, water uptake, transpiration, photosynthesis, enzyme activities, and metabolism of protein and lipids, a significant reduction in yield of up to 40% in agricultural lands has been reported [12,13]. Since the three most widely consumed kinds of cereal (maize, rice, and wheat) are susceptible to saline soil conditions [14], it is essential to improve studies about mechanisms to induce salt stress tolerance to secure our future on food storage.
Plants could tolerate salt stress by several strategies: (i) salt exclusion, retaining the Na+ in woody stems or roots; (ii) regulating toxic ions balance through the maintenance of a high K+/Na+ ratio with a mechanism of translocation of K+ over Na+, to reduce the Na+ concentration in the shoot cytoplasm; and (iii) by salt compartmentalization in vacuoles, protecting the cytoplasm from the ion toxicity [15,16]. The root system under an excessive salt concentration in the soil due to toxic ions accumulation suffers from inhibition of nutrition supply and elements adsorption [17]. Salinity, as drought, hurts the whole root system due to reduced cell activity in the root meristem [18]; however, growth rate reduction was determined more in the main root than in lateral roots [19]. As a response to stress, plants can remodel root system architecture (RSA) to adapt to adverse conditions [20]. However, the direction of root growth as a response to external stimuli differs in salt and drought conditions. Under drought conditions, roots use positive hydrotropism, growing towards the water, but in the presence of salt, roots remodeling due to negative halotropism directs the growth away from salinity. Moreover, it was reported that the roots of Arabidopsis exhibited reduced gravitropism under salt conditions to grow against the gravity vector [21]. In addition, the identified gene in cereal crops responsible for gravitropism versus anti-gravitropism regulatory mechanisms also controls the root angle [22]. The narrow root angle was determined as a very important feature in selecting genotypes for drought tolerance since genotypes with a narrow root angle can grow deeper compared with varieties with a wider root angle [23,24,25]. However, root system remodeling strategies to tolerate drought and salt stress differentiate in terms of tropism type, and it does not mean that a narrow root system able to adapt to drought is also capable of growing in saline soil.
In traditional breeding programs, phenotyping the above-ground plant parts was the main aspect of determining resistance to abiotic stress. However, the development of several high-throughput methods and techniques for root phenotyping in recent years has finally made it possible to determine which root systems and remodeling strategies are associated with the most successful plant development under stress conditions [26,27,28,29]. Phenotypic root traits could be used to determine genetic diversity; however, they are often influenced by environmental conditions. According to Soriano et al. [30], the stable quantitative trait loci (QTL) and closely linked molecular markers can improve durum wheat genetically through marker-assisted selection, cloning, and pyramiding in breeding programs to create new cultivars with desired quantitative traits. Molecular markers such as simple sequence repeat (SSR) and its based association mapping analysis may allow us to determine the chromosomal regions involved in root architecture features [30,31,32,33]. Moreover, SSRs are chromosome-specific and highly effective molecular tools due to their large level of polymorphism, distributed over all of the genomes [34,35]. Simple sequence repeat (SSR) molecular markers closely linked with genome regions associated with root architecture features could be helpful in selecting genotypes for salt tolerance breeding programs. Moreover, the identification of closely linked candidate genes can give a better understanding of the genetic response to specific stimuli.
In this study, six durum wheat genotypes, including one high salt-tolerant J. Kethifa as check genotype, were used to evaluate differences in shoot and root traits under control and salt conditions at the seedling stage and to determine their involvement in salt tolerance. The root system was evaluated using a high-throughput phenotyping system. The genotyping was performed using 11 SSR markers closely linked with genome regions associated with root traits, and in silico, candidate gene expression analysis was performed. The study aimed to evaluate genetic diversity among genotypes based on phenotypic and genotypic variations to identify candidate parental genotypes for salt-tolerance breeding.
2. Results
2.1. Salt Effect on Durum Wheat Shoots Traits
Highly statistically significant differences were detected between genotypes and both treatments (salt vs. control) for seedling length (PH), the number of leaves (NL), and the number of tillers (NT) (Table 1). Moreover, significant genotype by treatment interaction highlighted that the different genotypes used different strategies in terms of shoot development under salt stress. For example, the genotypes Azeghar and Sebatel had not shown a significant salt effect on any shoot traits and maintained one tall main stem with 5–7 leaves. In the genotypes, Cham1 and Pelsodur seedling length and the number of leaves were not significantly affected by salt; however, there was a significant negative reduction in the number of tillers. The salt-tolerant genotype J. Khetifa as a check plant, under salt stress, reduced seedling length and significantly increased the number of leaves (Table 1).
2.2. Salt Effect on the Root System
The analysis of variance revealed the presence of significant differences between genotypes and treatments for all root traits except average root diameter (AG) (Table 2). Significant interaction effects were detected for root angle (RA) and the number of tips (TI).
Azeghar, which maintained a wide root angle under both conditions, had a significant positive effect of salt only on root volume and surface area (Table 3). While Cham1, with wide and not affected by salt root angle as Azeghar, had a significant positive effect on root length (RL) that increased significantly by more than 80%, and the number of tips (TI) by more than double, and negative effect on root mean diameter (Table 3). Sebatel, Vulci, and Pelsodur showed a significant effect of salt only on root angle, which expanded in the presence of salt in the soil; this was particularly evident for Pelsodur, where the differences were around 40%. The highest positive significant effect on the root system under salt stress was determined for the salt-tolerant genotype J. Khetifa. Features such as root length (RL), root volume (RV), root surface area (SA), and the number of tips, forks, and crossings doubled under salt stress in seedlings of J. Khetifa. Under control conditions, Azeghar had a higher root length (RL) than J. Khetifa and Cham1, while under salt stress J. Khetifa maintained the longest roots, Sebatel and Azeghar had medium root length, and Vulci, Pelsodur, and Cham1 had the shortest. Under control conditions, Azeghar, Sebatel, and Cham1 had a wide root angle (RA), while J. Khetifa, Pelsodur, and Vulci had a narrow RA. However, RA widened significantly only for Pelsodur, Sebatel, and Vulci under stress. Under salt conditions, the mean root diameter (AG) significantly decreased only for Cham1 by about 24%.
2.3. Correlation among Traits
The correlation matrix among traits (Figure 1) shows that the root angle under both conditions (RAC and RAS) had a strong significant (p < 0.05) negative correlation with shoot traits such as the number of leaves and tillers under control (NLC, NTC) and salt (NLS and NTS) conditions. It could be suggested that genotypes with steeper root angles tend to produce more tillers and leaves.
Moreover, root angle under both conditions (RAC and RAS) had a highly significant positive correlation with root volume and surface area under control conditions. An interesting association was found between the root length (RL) and root traits that show the formation of new roots, such as the number of tips (TI), forks (FR), and crossings (CR). Root length under control (RLC) had a significant (p < 0.05) positive correlation with these traits (TIC, FRC, and CRC) only under control conditions and RLS with the same traits only under salt conditions. Moreover, root length under salt conditions (RLS) had a strong positive correlation with root volume (RVS) and root surface area (SAS) under salt conditions. In general, a significant positive correlation was found between root volume (RV) and surface area (SA) under both conditions, while both of them had a positive correlation with root angle (RAC) only under control conditions.
2.4. Principal Components Analysis (PCA)
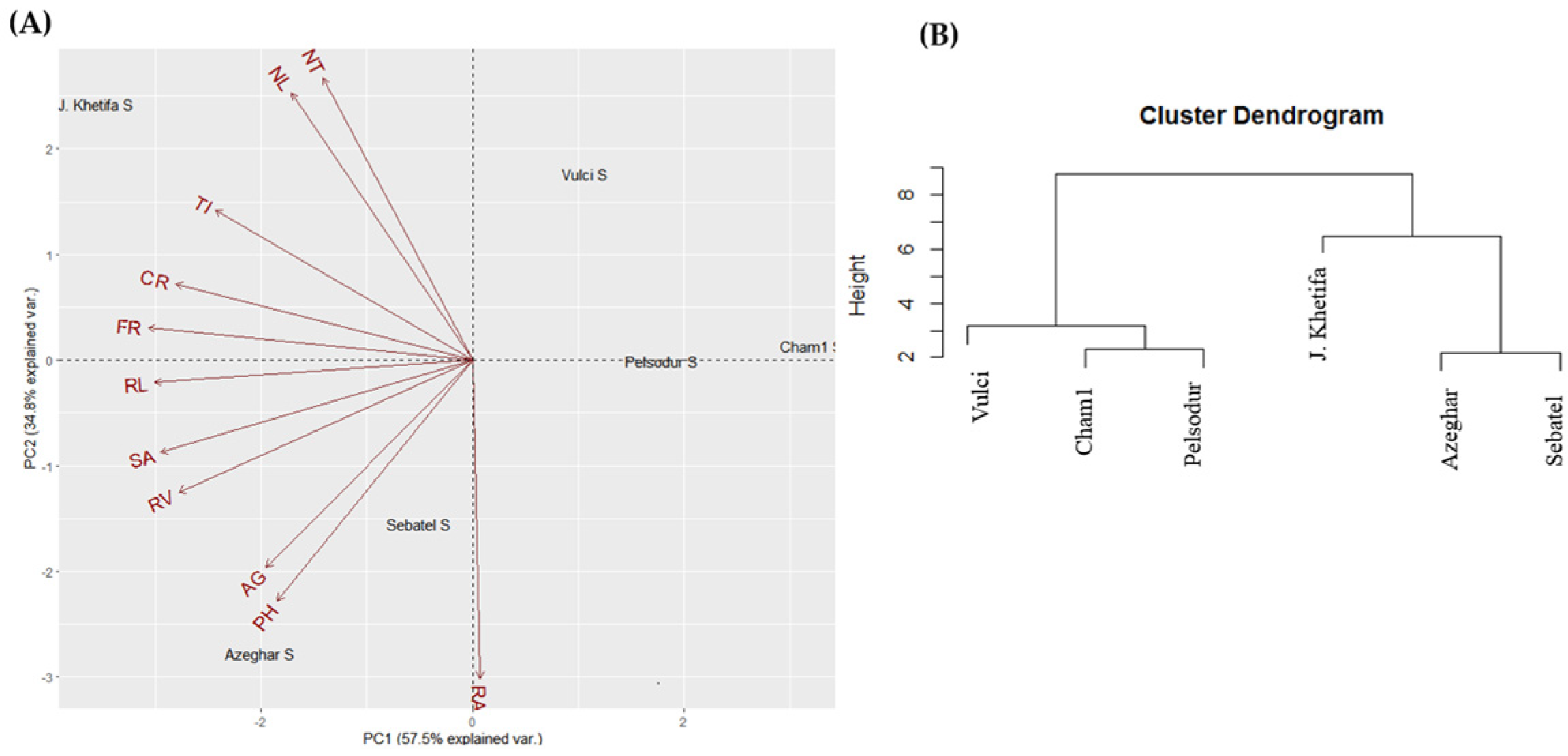
The top two principal components of PCA account for 82.6% of the total phenotypic variation under control conditions (Figure 2A). PC1 explained 49.1% and was mainly associated with root traits such as -RL, -SA, -TI, -FR, -CR (Table S1). At the same time, PC2 explained 33.5% of the total variation and was strongly associated with shoot trait and root angle (Figure 2A). Genotype J. Khetifa was grouped with Vulci and Pelsodur on the positive side, while Azeghar, Sebatel, and Cham1 were on the negative side of PC2. However, J. Khetifa with Cham1 was assigned on the PC1 positive side, Azeghar and Sebatel on the negative, and Vulci with Pelsodur were close to zero value. Three clusters of genotypes were identified based on hierarchical classification under control conditions (Figure 2B). One cluster grouped J. Khetifa, Vulci, and Pelsodur, the second group Sebatel and Azeghar; while Cham1 separated from the rest genotypes quite far as a third group. Under salt conditions, the top two principal components of PCA account for 92.3% of the total variation of root and shoot traits (Figure 3A). PC1 explained 57.5% and divided genotypes Vulci, Pelsodur, and Cham1 on the positive side of PC1, while J. Khetifa, Azeghar, and Sebatel were on the negative side.
PC2 explained 34.8% and was associated with shoot trait and root angle the same as under control conditions (Figure 3A). J. Khetifa with Vulci was assigned on the PC2 positive side, Azeghar and Sebatel on the negative, and Cham1 with Pelsodur were close to zero value. According to hierarchical analysis, all genotypes were distinguished into two major clusters, where Sebatel and Azeghar were assigned together with the salt-tolerant J. Khetifa to one cluster, and Vulci, Pelsodur, Cham1 were grouped to the second one (Figure 3B).
2.5. Genetic Diversity Based on SSR Markers
Eleven co-dominant SSR markers associated with specific root morphological traits were used to determine the genetic characteristics of the six genotypes. The number of alleles detected for each marker was very variable, ranging from 2 for wms205 to 5 for cfa2257, with an average of about three alleles per marker (Table 4). Some markers, i.e., gwm234 and wmc727, identified two loci labeled with a and b after the SSR name.
The observed heterozygosity was zero for most of the markers. Nevertheless, despite working with a self-pollinated species, the observed heterozygosity was not always equal to zero. Some markers detected high observed heterozygosity (i.e., wmc727a, wms205, and gwm573.2). Regarding the heterozygosity of wms205, it should be noted that all six genotypes have the same allelic situation leaving a suspect of a non-perfect allelic situation among the two detected bands common for all the genotypes. The gene diversity, computed as expected heterozygosity (He), and the polymorphism information content (PIC) provides information on a marker’s ability to determine polymorphism. In the present study, the gene diversity is quite high, ranging from 0.278 (gwm234a and wmc727b, which are second loci detected by an SSR marker) to 0.778 (cfa2086) (Table 4). The PIC values had a similar but not equal ranking among markers compared with the gene diversity parameter. PIC ranged from 0.24 for gwm234a and wmc727b to 0.74 for cfa2086 (Table 4). The Shannon Information Index (Table 4) indicates richness, and the evenness is always quite high ranging from 0.45 to 1.56, with an average of 1.01. The polymorphism among the different genotypes was not dramatically different; it ranged from about 13% in Pelsodur to about 27% in Azeghar and Sebatel, with an average of 21% (data not shown). Interestingly the number of Private alleles, i.e., alleles present only for a single genotype, is quite high (Table 5). All the genotypes have at least one allele present only in those genotypes. This goes from Vulci with one private allele to Azeghar and Pelsodur with four private alleles. Not surprisingly, cfa2086, with the higher PIC, also has a higher number of private alleles detected.
The UPGMA phenogram of the Nei (1972) [36] genetic distance cluster for 11 SSR markers (Figure 4) and all phenotypic data under control conditions (Figure 2B) discriminate the tested genotypes almost into similar clusters. Genotypes with wide root angles, such as Azeghar and Sebatel, were grouped morphologically and in terms of genetic distance. Another cluster, according to genetic distance, consisted of genotypes with narrow root angles, such as J. Khetifa, Pelsodur, and Vulci (Figure 4). Cham1, mainly in terms of phenotypic, was far from other genotypes, while genetically is far from the other genotypes but closer to Azeghar and Sebatel.
2.6. More Detailed Analysis of Genomic Regions Linked to Selected SSRs Markers and In Silico Candidate Gene Expression Analysis
The SSRs markers used in this study are distributed in seven chromosomes and overlap with 32 root-related QTLs (Table S2). For example, wms5, located on chromosome 3A in the durum wheat genome, overlapped with seven root-related QTLs for the number of root tips, primary root diameter, primary root length, lateral root diameter, total lateral root surface and volume, and total root number. Markers gwm427 and gwm459 located on chromosome 6A overlapped in total with eleven QTLs associated with root traits such as total root length, root angle, average root length, primary root length, root surface area, root tips, and total lateral root surface and volume, primary root volume. Markers gwm499 and gwm234, located on chromosome 5B, overlapped with three QTLs related to total root length, total lateral root length, and primary root volume. Gwm636 and cfa2086 markers on chromosome 2A overlapped with QTLs related to lateral root number per primary root, total root number, and root growth angle. Markers wmc727 and wms205 markers located on chromosome 5A overlapped with QTLs related to average root length and primary root length. Cfa2257 marker located on chromosome 7A overlapped with Meta QTL associated with total root number and primary root length. Marker, gwm573.2, located on chromosome 7B, overlapped with root traits related to Meta QTL.
Candidate genes for investigating and estimating gene expression levels were identified according to SSRs location on the durum wheat ‘Svevo’ genome (Table S3). A total of 57 gene models were detected and analyzed at http://www.wheat-expression.com/ (accessed on 13 January 2023), using their RNAseq to obtain the expression of genes involved in root response to abiotic stress at the seedling stage (Table S4).
In the durum wheat genome, (RefSeq v1.0 Chr 2A region), the mapped marker gwm636 was close to nine genes models associated with growth and tolerance to abiotic and biotic stresses, such as cell differentiation protein RCD1, sterol 3-beta-glucosyltransferase, cytochrome P450, Sec24-like transport protein, AAR2 family protein, leucine-rich repeat receptor-like protein kinases. All genes were up-regulated in seedlings’ roots under abiotic stress (Table S4). The marker wms5 was close to seven genes models associated with response to stress or plant development under stress conditions, including thioredoxin-like protein, homeobox leucine-zipper protein, citrate-binding protein, DUF1677 domain-containing protein, AGAMOUS-like MADS-box protein, kinase superfamily protein, and core-2/I-branching beta-1, 6-N-acetylglucosaminyltransferase family protein. Five of these were up-regulated in seedlings’ roots under abiotic stress. Wmc727 on chromosome 5A was linked to 10 genes models mostly involved in the regulation of secondary metabolites related to antioxidant activity, metabolic pathways, transcription, and signaling, such as membrane protein insertase YidC, O-methyltransferase family protein, nuclease S1, short-chain dehydrogenase/reductase family protein, root phototropism protein, Chalcone, and Curcuminoid synthases, F-box-like protein, and Palmitoyl protein thioesterase containing protein. Eight from 10 genes linked to marker gwm499 on chromosome 5B were close to genes associated with response to abiotic stress, such as receptor kinase, basic helix-loop-helix transcription factor, and ADP-ribosylation factor GTPase-activating protein involved in the root growth direction, and was most expressed gene near gwm499 marker. The marker gwm427 on chromosome 6A was linked to seven genes models related to plant’s adaptation and tolerance to abiotic stress, such as actin-related protein 2/3 complex subunit, plastid movement impaired 1-related 1 G protein, cytochrome P450, inosine-5’-monophosphate dehydrogenase, CBS domain-containing protein. All genes linked to marker gwm427 were expressed under abiotic stress in seedling roots. The marker gwm573.2 was close to 21 gene models associated with plant growth and development under abiotic stress such as trehalose 6-phosphate phosphatase, a protein of the DNA-directed RNA polymerase the ABC transporter, phosphatidylinositol: ceramide inositol phosphotransferase, shikimate kinase, G-protein, ATP-dependent zinc metalloprotease FtsH1, Aldo/keto reductase family oxidoreductase, Sec14p-like phosphatidylinositol transfer family protein, Pentetratricopeptide repeat proteins, a bHLH transcription factor, the regulator of chromosome condensation (RCC1) family with FYVE zinc finger domain-containing protein, HVA22-like protein, TCP transcription factor, Photosystem (PS) II CP43 reaction center protein, BTB/POZ domain-containing protein, Sphingoid base hydroxylase 2, and two F-box-like protein. Almost all of them were up-regulated in wheat seedlings’ roots under abiotic stress conditions (Table S4).
3. Discussion
The root system remodeling strategies under adverse conditions is a great opportunity for adaptation and optimal development of the plant, as the first organ that senses and responds to abiotic stress, and then through efficient uptake of water and nutrients. Latest studies demonstrated that root system features, especially root angle in crop plants, play a main role in adapting to adverse conditions [20,22,37].
According to Pariyar et al. [38], together with root traits, leaf-related traits should be included for more efficient selection in breeding programs for whole plant establishment. At the same time, the ideotype of the root system plays a key role in ensuring sufficient leaf area, which in turn is responsible for efficient photosynthesis, the crucial process for plant development. Genotypes that produce new leaves faster than the die of old ones and maintain a high number of leaves or shoot biomass under stress conditions could be considered tolerant. Moreover, previous studies have shown that salt-tolerant genotypes of durum wheat accumulated higher shoot biomass and maintained a bigger leaf area under salt stress compared with susceptible varieties [39,40,41]. In our study, some shoot traits were associated with root angle. For example, the salt-tolerant genotype, J. Khetifa, under salt stress, maintained a narrower root angle and the highest number of leaves. Moreover, Vulci, genetically close to J. Khetifa, was characterized by a narrow root angle and a large number of leaves under salt conditions. However, in our previous drought study [25], Pelsodur, under control and drought conditions, maintained a narrow root angle, which was associated with good shoot development and drought tolerance. While in this study, under salt stress, the Pelsodur root angle changed significantly from narrow to wide, and this caused a significant negative effect on his shoot development, especially tillers that decreased by 60%. Moreover, in genotypes with wider root angles, such as Sebatel, Cham1, and Azeghar, the number of leaves was not significantly affected by salinity. It is well known that a narrow root angle is one of the main traits for drought tolerance, but it was demonstrated that a shallower root growth angle could enhance rice yields in saline environments [20].
Phenotypic data from our pot experiment obtained using the WinRHIZO software (version 4.0b) highlighted a few trends in plant response to salt stress at the seedling stage. Genotypes under control conditions were distinguished regarding root angle into two groups, genotypes Vulci and Pelsodur with high salt-tolerant J. Khetifa had a narrower root angle than genotypes Sebatel, Azeghar, and Cham1. It was reported that root angle depends on plant root ability to control gravitropic versus anti-gravitropic mechanisms, which is the main factor in avoiding salty soil [20,21,22]. In this regard, root tips could be a good indicator for selection as they are involved in root gravity perception and regulation. In addition, the salt-tolerant genotype J. Khetifa under salt stress doubled the number of root tips and had a significantly higher number compared with the other genotypes. However, under salt conditions, the salt-tolerant J. Khetifa, with a narrow root angle, was grouped with genotypes Azeghar and Sebatel, which had a wider root angle. These results suggest that a narrow root angle is not a good indicator of salt stress tolerance.
In general, the check genotype J. Khetifa under salt stress at the seedling stage improved the number of leaves, which is a good salt tolerance indicator in terms of accumulation of toxic ions and efficient photosynthesis. Moreover, J. Khetifa improved almost all root traits under salt conditions, except the root angle, which stayed narrow, and the mean root diameter. The other two genotypes, Azeghar and Sebatel, from the same cluster of J. Khetifa under salt stress, had wide root angles, and salt did not affect shoot traits significantly. Moreover, genotype Azeghar under salt increased root volume and surface area, and the roots of Sebatel were not affected significantly. Based on present results, it can be concluded that a narrow root angle is not a good indicator of salt tolerance, moreover, exist more root system adaptation strategies that depend on the inner capacity of genotypes to maintain an efficient amount of roots.
Dendrograms based on the cluster analysis of the genetic similarity coefficients and phenotypic data under control conditions grouped genotypes mainly due to root angle. In this regard, the SSR marker wms5 can be quite informative in selecting genotypes for narrow-angle root system ideotype. It was reported that the wms5 marker was significantly associated with root angle [42], and it was located on chromosome 3A in the durum wheat genome. In our study, genotypes grouped in the same clusters according to root angle, such as J. Khetifa, Vulci, and Pelsodur, had narrow root angles (wms5 band of 190 bp), and Sebatel and Azeghar (wms5 band of 180 bp), Cham1 (wms5 band of 185 bp) had wide root angles. Moreover, in the durum wheat genome, the location of wms5 was close to gene models representing proteins involved in plant development and response to stress. Such as thioredoxin, which is a small ubiquitous protein that reduces protein disulfides, protects cells from oxidative damage, and protects plants from the oxidative deactivation of photosynthesis-related enzymes [43]. Moreover, homeobox leucine-zipper transcription factors are responsible for plant growth adaptation in water deficit conditions [44,45]. Core-2/I-branching beta-1,6-N-acetylglucosaminyltransferase family protein is involved in cell elongation during the seedling growth stage and senescence of the leaf process [46]. Citrate-binding protein family is involved in plant development and growth, also in response to stress related to the accumulation of toxic ions in the vacuoles [47]. DUF1677 family proteins are DUF domain-containing proteins that play a vital role in plant stress responses, including salinity [48,49]. Kinases superfamily proteins are well known as signal transmitters in various stresses [2]. Members of the MADS-box gene family are involved in many processes of plant development and morphogenesis, but it was also reported to modulate root architecture [50,51].
Our results revealed that the root system under the salt of genotypes Azeghar and Sebatel were close to the one of salt-tolerant J. Khetifa, regardless of root angle differences, making them good candidates as parent lines in breeding programs. In this respect, some rare alleles of SSR markers could help to select salt-tolerant candidates in the segregant populations. For example, the salt-tolerant J. Khetifa and Sebatel had rare alleles near marker cfa2086, 277 bp, and 223 bp, respectively. Moreover, salt-tolerant J. Khetifa had rare alleles by SSR marker gwm234 (214 bp) and near marker gwm499 (154 bp). Moreover, genotype Sebatel had a rare allele also near marker gwm499. In the durum wheat genome, the mapped marker gwm499 was close to gene models for kinases superfamily protein, which mostly regulates transcription factors. Moreover, it was near to basic helix-loop-helix transcription factors, which are involved in a range of abiotic stress responses and adaptive responses to drought [52]. ADP-ribosylation factor GTPase-activating protein was shown that is crucial for determining directional root hair growth in Arabidopsis [53].
Genotypes Azeghar and Sebatel had rare alleles near marker gwm427, 201 bp, and 237 bp, respectively. The marker gwm427 position was found to overlap with previously reported durum wheat major QTL for root growth angle (qSRA-6A), where candidate gene analysis showed loci related to gravitropism, polar growth, and hormonal signaling [54]. In the durum wheat genome (RefSeq v1.0 Chr 6A region), the mapped marker gwm427 was close to genes models for actin-related protein and plastid movement-impaired related protein. Marker gwm427’s location in the durum wheat genome was close to two related genes such as (i) the actin-related protein and (ii) the plastid movement impaired 1-related 1 G protein that regulates organelle movement [55]. In general, the movement and positioning of the organelles play an important role in the adaptation of plants to environmental stress. Moreover, actin-related protein regulates developmental processes such as cell division and development and cell polarity [56]. Marker gwm427 location in the durum wheat genome was close to other five gene annotations (Table S4); (iii) cytochrome P450 from the oxidoreductases enzymes class has been reported to be involved in plant defense against various abiotic and biotic stresses [57]. (iv) Inosine-5’-monophosphate dehydrogenase catalyzes the biosynthesis of the guanine nucleotide, which is important for DNA and RNA synthesis, signal transduction, energy transfer, glycoprotein synthesis, and cellular proliferation [58]. (v) Proline-tRNA ligase is an enzyme involved in proline metabolism and KEGG and aminoacyl-tRNA biosynthesis pathways (BRENDA:EC 6.1.1.15). L-Proline in a stressful environment is crucial for plants due to its protective role as an osmoprotectant and stabilizes cellular structures and enzymes, and scavenging reactive oxygen species [59]. (vi) Potassium-transporting ATPase could help plant cells to maintain a high concentration of potassium and a low concentration of sodium inside in case of high sodium concentrations in the soil. (vii) Gene containing cystathionine β-synthase (CBS) domain was found to be upregulated under high salinity, and overexpressing this gene showed higher abiotic stress tolerance in transgenic plants [60].
Genotype Azeghar had three more rare alleles, two near marker cfa2257 (104 bp and 142 bp) and one near gwm573.2 (181 bp) (Table S4). The marker gwm573.2 was close to gene models translated to trehalose 6-phosphate phosphatase, which regulates the activity of protein kinase, and this has a large impact on cell division, development, and function [61]. The protein of the DNA-directed RNA polymerase belongs to complex molecular machines for the essential function of the synthesis of RNA from DNA templates, otherwise transcription [62]. Abscisic acid is a substrate of the ABC transporter that is involved in the active transport of various molecules across biological membranes, such as heavy metals, lipids, glucosinolates, and phytohormones [63] allowing to coordinate of physiological and developmental processes at the whole-plant level and being involved in response to several induced by biotic stresses. Other related genes (Table S4) are (i) Phosphatidylinositol: ceramide inositol phosphotransferase, which is essential for sphingolipid biosynthesis and plays an important role in dehydration stress tolerance [64]. (ii) Shikimate kinase catalyzes the reaction of the shikimate pathway that redirects carbon from the central metabolism pool to a wide range of secondary metabolites responsible for plant development, growth, and stress responses [65]. (iii) G-proteins are universal signal transducers involved in many cellular responses of plants [66]. (iv) ATP-dependent zinc metalloprotease FtsH 1 is involved in the formation of thylakoid membranes during early chloroplast development and in the proteolysis of damaged photosystem II (PSII) protein complex repair cycle, which is essential to avoid photoinhibition due to the accumulation of photodamaged PSII [67]. (v) Aldo/keto reductase family oxidoreductase is involved in many plant metabolites reactions, such as the biosynthesis of osmolytes, reactive aldehyde detoxification, secondary metabolism, and membrane transport [68]. (vi) Sec14p-like phosphatidylinositol transfer family protein is a plasma membrane-associated protein that has a crucial role in rooting patterning due to auxin signaling and PIN distribution in Arabidopsis [69]. (vii) Pentatricopeptide repeat proteins regulate chloroplast gene expression and RNA metabolism in higher plants [70]. (viii) Transcription factor bHLH involved in response and tolerance to various plant stresses, including soil salinity [71]. (ix) The regulator of chromosome condensation (RCC1) family with FYVE zinc finger domain-containing protein is essential for regulating auxin flows toward the direction of gravity and root branch angle in Arabidopsis [72]. (x) Transcriptomic analysis in roots revealed that the ABA-related gene encoding HVA22-like proteins was up-regulated, and considering that ABA biosynthesis is correlated with auxin transport to the root tips, their co-expression can be beneficial for the development and morphology of the roots [73]. (xi) TCP transcription factor may activate OsNHX1 gene expression, which response to salt and PEG-induced drought stress in rice, and may be associated with abiotic stress tolerance [74]. (xii) Photosystem (PS) II CP43 reaction center protein is the main antenna of the PS II pigment-protein complex in the thylakoid membrane and plays an essential role in the photosynthetic responses to stresses in higher plants [75]. (xiii) BTB/POZ domain-containing proteins are associated with plant growth and development, also involved in plant responses and defense against biotic and abiotic stresses [76]. (xiv) The enzyme Sphingoid base hydroxylase II is essential for growth and viability in Arabidopsis [77]. The marker gwm573.2 was also close to (xv) two F-box-like protein genes models, which were described earlier as genes involved in salt tolerance, and rooting abilities.
4. Materials and Methods
4.1. Plant Material
For phenotyping and genotyping was used six Triticum turgidum L. subsp. durum (Desf.) Husn. accessions, including one high salt-tolerant genotype Jennah Khetifa as check [78,79]. Other genotypes characterized by different root angles, Cham1, Azeghar 2-1 (56) (Azeghar), and Sebatel2 (45) (Sebatel) with wide root angle, and two genotypes Mv-Pelsodur (Pelsodur) and Vulci with narrow root angle [25].
4.2. Pot Experiment Condition and Design
A pot experiment was performed in a greenhouse at Tuscia University’s experimental farm (Viterbo, Italy) from January to February 2021. A Randomized Complete Block Design (RCBD) with three replications, six genotypes, and two salinity levels was used. The six genotypes were sown with one seed per pot placed with the embryo facing down on 20 January 2021. Each pot was 17 cm (diameter) by 16 cm (high) and filled with 2.5 L of sand (Figure S1). The temperature in the greenhouse was constantly maintained at around 25 °C during the day and around 15 °C during the night to have optimum growing conditions. Before starting the treatments, all pots (control and salinity treatment) were irrigated, using a dropping pipe, three times per week (80 mL/per pot) with a water nutrition solution composed of dihydrogen phosphate potassium (0.13 g/L), potassium sulfate (0.04 g/L), nitric acid (0.286 mL/L), calcium nitrate (0.432 g/L), potassium nitrate (0.436 g/L), magnesium nitrate (0.244 g/L) and Micron (0.23 g/L) [25]. Two weeks after sowing, when all seedlings reached two fully expanded leaves stage, plants under salinity treatment were started treated with a salt solution (250 mM NaCl) 3 times per week (80 mL/per pot) for 14 days, while the ones under control treatment were irrigated with the same solution but without NaCl. After two weeks of salt treatment, pots were watered with a water nutrition solution without salt for 14 days. After two weeks (no NaCl), for a week, the salt solution was applied again, and after that, the plants were removed for analysis (Figure 5).
4.3. Roots Phenotyping
Plants for analysis were collected after 7 weeks after sowing in total. Roots were carefully washed to remove the sand, using a soft spray watering head, and analyzed using Win-RHIZO Pro software v2009 (version 4.0b; Regent Instruments, Montreal, QC, Canada). The morphological traits of the roots were recorded for the whole root system (Figure 6A), such as total root length (RL), the sum of all roots lengths, cm, surface area (SA), total root surface area, cm2, root volume (RV), total root volume, cm3, number of tips (TI), forks (FR), and crossings (CR). The root angle (RA, degree on the vertical °) between the two extreme sides of the roots with the center set in the middle of the crown was measured using the software ImageJ, which could be freely download at https://imagej.nih.gov/ij/download.html; accessed on 13 January 2023 (Figure 6B). Morphological traits for shoots, such as plant height, number of leaves, and number of tillers, were also recorded.
4.4. DNA Extraction, PCR Amplifications, and Gel Electrophoresis
For genotypic diversity analysis, 11 microsatellites associated or linked to genomic regions related to root traits from previously reported studies (Table S2) were chosen. The genomic DNA was extracted from fresh leaves of each genotype using the extraction kit PureLink Plant Total DNA Purification kit (Invitrogen; Thermo Fisher Scientific, Waltham, Massachusetts USA). The PCR amplifications were carried out using 0.125 μL of GoTaq G2 DNA Polymerase (Promega, Madison, Wisconsin, USA), 5 μL of 5X Colorless GoTaq® Reaction Buffer, 0.5 μL of 10 µM dNTP mix, 2 μL of 10 µM/μL forward and reverse primer, 100 ng of gDNA, and sterile water for a total reaction amount of 25 μL. The amplifications were performed in Swift Maxi Thermal Cyclers (Esco Technologies, St. Louis, Missouri, USA) with an initial denaturation of 5 min at 94 °C followed by 35 cycles with 15 s at 94 °C and 20 s at different temperatures, according to the annealing temperature of each marker (see Table 6), and 20 s at 72 °C. As a final step, it was performed an extension of 10 min at 72 °C. The electrophoresis was carried out through the capillary electrophoresis device QIAxcel Advanced Instrument (Qiagen, Hilden, Germania) using the QIAxcel DNA High-Resolution Kit, and allele analyses were performed with QIAxcel ScreenGel Software (Qiagen, Hilden, Germania). Each allele was named for its band size in the base pair (Table S5).
Gene models were identified in the NCBI database (https://www.ncbi.nlm.nih.gov/data-hub/genome/GCA_900231445.1/, accessed on 13 January 2023) based on the positions of SSR markers in the durum wheat reference sequence (https://wheat.pw.usda.gov/GG3/jbrowse_Durum_Svevo, accessed on 13 January 2023). Identification of upregulated gene models under abiotic stress at the seedling stage was carried out using the RNAseq data at http://www.wheat-expression.com/ (accessed on 13 January 2023), using gene models from the NCBI database.
4.5. Statistical Analysis
The statistical analyses of phenotypic data were performed using R Studio (Version R-4.1.0, R Foundation for Statistical Computing, Vienna, Austria). Two-way analysis of variance (ANOVA) was conducted at a significance level of 5% using the aov() function. At the same time, one-way ANOVA was used to test the variance component of each trait under each treatment, with genotype as a factor. Fisher’s least significant difference (LSD) test was used for means comparisons. Pearson correlation coefficients were calculated using the corrplot function R package [80]. Principal component analysis was performed using the prcomp() function, and then a biplot was generated with the ggbiplot function R package [80]. The genetic distance [36] matrix was utilized to construct, using GDA software [81]. The Mantel test to compare genetic distance and Euclidean matrices was computed by GenAlEx, an Excel macro realized by Peakall and Smouse [82].
5. Conclusions
The identification of molecular markers linked to root system traits is very important for marker-assisted selection in breeding programs. The SSR markers linked to genetic regions associated with root traits in this study were effective in discriminating the genotypes and can be used to screen candidate lines for salt tolerance. Nevertheless, the narrow root angle is not a good indicator of salt tolerance since there are more root system adaptation strategies that depend on the inner capacity of genotypes to maintain an efficient amount of roots. Moreover, present results of phenotypic response to salt stress using Win-RHIZO Pro software together with molecular analysis can provide useful highlights to increase the efficiency of plant breeding programs.
Supplementary Materials
The following supporting information can be downloaded at: https://www.mdpi.com/article/10.3390/plants12020412/s1, Table S1: Principal Components Analysis of traits detected under salt and control conditions; Table S2: Markers’ locations onto the durum wheat reference genome associated with root-related QTL; Table S3: Genes linked with genes from NCBI database; Table S4: Expressed genes involved in abiotic stress in the roots of seedlings. Table S5: PCR results and alleles distribution among the 11 SSRs. Figure S1: Water and nutrient distribution in the pots filled with sand.
Author Contributions
Conceptualization, I.U., L.B., and M.A.P.; Data curation, I.U. and L.B.; Formal analysis, I.U., L.B. and M.A.P.; Investigation, I.U. and L.B.; Methodology, M.A.P.; Project administration, M.A.P.; Resources, M.A.P.; Software, I.U. and L.B.; Supervision, M.A.P.; Validation, I.U., L.B., and M.A.P.; Visualization, I.U.; Writing—original draft, I.U.; Writing—review & editing, M.A.P. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the European Union’s Horizon 2020 research and innovation program under grant agreement No 771367 (ECOBREED project) and by the European Union Next-GenerationEU (Piano Nazionale di Ripresa e Resilienza (PNRR)—missione 4 componente 2, investimento 1.4—D.D. 1032 17/06/2022, CN00000022). This manuscript reflects only the authors’ views and opinions; neither the European Union nor the European Commission can be considered responsible for them.
Informed Consent Statement
Not applicable.
Data Availability Statement
https://doi.org/10.5281/zenodo.7534377 (accessed on 20 December 2022).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
AG: root diameter; CR: total number of crossings; FR: total number of forks; NL: number of leaves; NT: number of tillers; PH: seedling length; RA: root angle; RL: sum of all root lengths; RV: total root volume; SA: total root surface area; TI: total number of tips.
References
- Food and Agriculture Organization of the United Nations. FAO 2022 Global Map of Salt-Affected Soils. Fao Soils Portal. Available online: https://www.fao.org/soils-portal/data-hub/soil-maps-and-databases/global-map-of-salt-affected-soils/en/ (accessed on 10 August 2022).
- Urbanavičiūtė, I.; Bonfiglioli, L.; Pagnotta, M.A. One hundred candidate genes and their roles in drought and salt tolerance in wheat. Int. J. Mol. Sci. 2021, 22, 6378. [Google Scholar] [CrossRef] [PubMed]
- D’Odorico, P.; Bhattachan, A.; Davis, K.F.; Ravi, S.; Runyan, C.W. Global desertification: Drivers and feedbacks. Adv. Water Resour. 2013, 51, 326–344. [Google Scholar] [CrossRef]
- Daneshmand, H.; Alaghmand, S.; Camporese, M.; Talei, A.; Yeh, P.J.F.; Daly, E. Long-Term Impacts of Partial Afforestation on Water and Salt Dynamics of an Intermittent Catchment under Climate Change. Water 2020, 12, 1067. [Google Scholar] [CrossRef] [Green Version]
- El-Ramady, H.; Faizy, S.; Amer, M.M.; Elsakhawy, T.A.; Omara, A.E.-D.; Eid, Y.; Brevik, E. Management of Salt-Affected Soils: A Photographic Mini-Review. Environ. Biodivers. Soil Secur. 2022, 6, 61–79. [Google Scholar] [CrossRef]
- Corwin, D.L. Climate change impacts on soil salinity in agricultural areas. Eur. J. Soil Sci. 2021, 72, 842–862. [Google Scholar] [CrossRef]
- Daliakopoulos, I.N.; Tsanis, I.K.; Koutroulis, A.; Kourgialas, N.N.; Varouchakis, A.E.; Karatzas, G.P.; Ritsema, C.J. The threat of soil salinity: A European scale review. Sci. Total Environ. 2016, 573, 727–739. [Google Scholar] [CrossRef]
- Evelin, H.; Devi, T.S.; Gupta, S.; Kapoor, R. Mitigation of salinity stress in plants by arbuscular mycorrhizal symbiosis: Current understanding and new challenges. Front. Plant Sci. 2019, 10, 470. [Google Scholar] [CrossRef] [Green Version]
- Tedeschi, A. Irrigated Agriculture on Saline Soils: A Perspective. Agronomy 2020, 10, 1630. [Google Scholar] [CrossRef]
- MUNNS, R. Physiological processes limiting plant growth in saline soils: Some dogmas and hypotheses. Plant. Cell Environ. 1993, 16, 15–24. [Google Scholar] [CrossRef]
- Wang, J.; Zhu, J.; Zhang, Y.; Fan, F.; Li, W.; Wang, F.; Zhong, W.; Wang, C.; Yang, J. Comparative transcriptome analysis reveals molecular response to salinity stress of salt-tolerant and sensitive genotypes of indica rice at seedling stage. Sci. Rep. 2018, 8, 2085. [Google Scholar] [CrossRef]
- Qadir, M.; Quillérou, E.; Nangia, V.; Murtaza, G.; Singh, M.; Thomas, R.J.; Drechsel, P.; Noble, A.D. Economics of salt-induced land degradation and restoration. Nat. Resour. Forum 2014, 38, 282–295. [Google Scholar] [CrossRef]
- Safdarian, M.; Askari, H.; Shariati, J.V.; Nematzadeh, G. Transcriptional responses of wheat roots inoculated with Arthrobacter nitroguajacolicus to salt stress. Sci. Rep. 2019, 9, 1792. [Google Scholar] [CrossRef]
- Majeed, A.; Muhammad, Z.; Islam, S.; Ahmad, H. Salinity imposed stress on principal cereal crops and employing seed priming as a sustainable management approach. Acta Ecol. Sin. 2019, 39, 280–283. [Google Scholar] [CrossRef]
- Munns, R.; James, R.A.; Läuchli, A. Approaches to increasing the salt tolerance of wheat and other cereals. J. Exp. Bot. 2006, 57, 1025–1043. [Google Scholar] [CrossRef] [Green Version]
- Mansour, E.; Moustafa, E.S.A.; Desoky, E.S.M.; Ali, M.M.A.; Yasin, M.A.T.; Attia, A.; Alsuhaibani, N.; Tahir, M.U.; El-Hendawy, S. Multidimensional Evaluation for Detecting Salt Tolerance of Bread Wheat Genotypes Under Actual Saline Field Growing Conditions. Plants 2020, 9, 1324. [Google Scholar] [CrossRef] [PubMed]
- Attia, H.; Arnaud, N.; Karray, N.; Lachaâl, M. Long-term effects of mild salt stress on growth, ion accumulation and superoxide dismutase expression of Arabidopsis rosette leaves. Physiol. Plant. 2008, 132, 293–305. [Google Scholar] [CrossRef]
- West, G.; Inzé, D.; Beemster, G.T.S. Cell Cycle Modulation in the Response of the Primary Root of Arabidopsis to Salt Stress. Plant Physiol. 2004, 135, 1050–1058. [Google Scholar] [CrossRef]
- Julkowska, M.M.; Koevoets, I.T.; Mol, S.; Hoefsloot, H.; Feron, R.; Tester, M.A.; Keurentjes, J.J.B.; Korte, A.; Haring, M.A.; De Boer, G.J.; et al. Genetic components of root architecture remodeling in response to salt stress. Plant Cell 2017, 29, 3198–3213. [Google Scholar] [CrossRef] [Green Version]
- Kitomi, Y.; Hanzawa, E.; Kuya, N.; Inoue, H.; Hara, N.; Kawai, S.; Kanno, N.; Endo, M.; Sugimoto, K.; Yamazaki, T.; et al. Root angle modifications by the DRO1 homolog improve rice yields in saline paddy fields. Proc. Natl. Acad. Sci. USA 2020, 117, 21242–21250. [Google Scholar] [CrossRef] [PubMed]
- Sun, F.; Zhang, W.; Hu, H.; Li, B.; Wang, Y.; Zhao, Y.; Li, K.; Liu, M.; Li, X. Salt modulates gravity signaling pathway to regulate growth direction of primary roots in arabidopsis. Plant Physiol. 2007, 146, 178–188. [Google Scholar] [CrossRef]
- Fusi, R.; Rosignoli, S.; Lou, H.; Sangiorgi, G.; Bovina, R.; Pattem, J.K.; Borkar, A.N.; Lombardi, M.; Forestan, C.; Milner, S.G.; et al. Root angle is controlled by EGT1 in cereal crops employing an antigravitropic mechanism. Proc. Natl. Acad. Sci. USA 2022, 119, e2201350119. [Google Scholar] [CrossRef]
- El Hassouni, K.; Alahmad, S.; Belkadi, B.; Filali-Maltouf, A.; Hickey, L.T.; Bassi, F.M. Root system architecture and its association with yield under different water regimes in Durum wheat. Crop Sci. 2018, 58, 2331–2346. [Google Scholar] [CrossRef] [Green Version]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.S.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Urbanaviči, I.; Bonfiglioli, L.; Pagnotta, M.A. Diversity in Root Architecture of Durum Wheat at Stem Elongation under Drought Stress. Agronomy 2022, 12, 1329. [Google Scholar] [CrossRef]
- Watt, M.; Moosavi, S.; Cunningham, S.C.; Kirkegaard, J.A.; Rebetzke, G.J.; Richards, R.A. A rapid, controlled-environment seedling root screen for wheat correlates well with rooting depths at vegetative, but not reproductive, stages at two field sites. Ann. Bot. 2013, 112, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Gioia, T.; Galinski, A.; Lenz, H.; Müller, C.; Lentz, J.; Heinz, K.; Briese, C.; Putz, A.; Fiorani, F.; Watt, M.; et al. GrowScreen-PaGe, a non-invasive, high-throughput phenotyping system based on germination paper to quantify crop phenotypic diversity and plasticity of root traits under varying nutrient supply. Funct. Plant Biol. 2016, 44, 76–93. [Google Scholar] [CrossRef]
- Nagel, K.A.; Putz, A.; Gilmer, F.; Heinz, K.; Fischbach, A.; Pfeifer, J.; Faget, M.; Blossfeld, S.; Ernst, M.; Dimaki, C.; et al. GROWSCREEN-Rhizo is a novel phenotyping robot enabling simultaneous measurements of root and shoot growth for plants grown in soil-filled rhizotrons. Funct. Plant Biol. 2012, 39, 891–904. [Google Scholar] [CrossRef] [Green Version]
- Canales, F.J.; Nagel, K.A.; Müller, C.; Rispail, N.; Prats, E. Deciphering Root Architectural Traits Involved to Cope with Water Deficit in Oat. Front. Plant Sci. 2019, 10, 1558. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soriano, J.M.; Alvaro, F. Discovering consensus genomic regions in wheat for root-related traits by QTL meta-analysis. Sci. Rep. 2019, 9, 10537. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanguineti, M.C.; Li, S.; MacCaferri, M.; Corneti, S.; Rotondo, F.; Chiari, T.; Tuberosa, R. Genetic dissection of seminal root architecture in elite durum wheat germplasm. Ann. Appl. Biol. 2007, 151, 291–305. [Google Scholar] [CrossRef]
- Canè, M.A.; Maccaferri, M.; Nazemi, G.; Salvi, S.; Francia, R.; Colalongo, C.; Tuberosa, R. Association mapping for root architectural traits in durum wheat seedlings as related to agronomic performance. Mol. Breed. 2014, 34, 1629–1645. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iannucci, A.; Marone, D.; Russo, M.A.; De Vita, P.; Miullo, V.; Ferragonio, P.; Blanco, A.; Gadaleta, A.; Mastrangelo, A.M. Mapping QTL for Root and Shoot Morphological Traits in a Durum Wheat × T. dicoccum Segregating Population at Seedling Stage. Int. J. Genom. 2017, 2017, 6876393. [Google Scholar] [CrossRef] [Green Version]
- Maccaferri, M.; Stefanelli, S.; Rotondo, F.; Tuberosa, R.; Sanguineti, M.C. Relationships among durum wheat accessions. I. Comparative analysis of SSR, AFLP, and phenotypic data. Genome 2007, 50, 373–384. [Google Scholar] [CrossRef] [PubMed]
- Mondini, L.; Noorani, A.; Pagnotta, M.A. Assessing Plant Genetic Diversity by Molecular Tools. Diversity 2009, 1, 19–35. [Google Scholar] [CrossRef] [Green Version]
- Nei, M. Genetic Distance between Populations. Am. Nat. 1972, 106, 283–292. [Google Scholar] [CrossRef]
- Schneider, H.M.; Lor, V.S.N.; Hanlon, M.T.; Perkins, A.; Kaeppler, S.M.; Borkar, A.N.; Bhosale, R.; Zhang, X.; Rodriguez, J.; Bucksch, A.; et al. Root angle in maize influences nitrogen capture and is regulated by calcineurin B-like protein (CBL)-interacting serine/threonine-protein kinase 15 (ZmCIPK15). Plant. Cell Environ. 2022, 45, 837–853. [Google Scholar] [CrossRef] [PubMed]
- Pariyar, S.R.; Nagel, K.A.; Lentz, J.; Galinski, A.; Wilhelm, J.; Putz, A.; Adels, S.; Heinz, K.; Frohberg, C.; Watt, M. Variation in Root System Architecture among the Founder Parents of Two 8-way MAGIC Wheat Populations for Selection in Breeding. Agronomy 2021, 11, 2452. [Google Scholar] [CrossRef]
- Houshmand, S.; Arzani, A.; Maibody, S.A.M.; Feizi, M. Evaluation of salt-tolerant genotypes of durum wheat derived from in vitro and field experiments. F. Crop. Res. 2005, 91, 345–354. [Google Scholar] [CrossRef]
- Katerji, N.; Van Hoorn, J.W.; Hamdy, A.; Mastrorilli, M.; Oweis, T. Salt tolerance analysis of chickpea, faba bean and durum wheat varieties: I. Chickpea and faba bean. Agric. Water Manag. 2005, 72, 177–194. [Google Scholar] [CrossRef]
- Chamekh, Z.; Ayed, S.; Sahli, A.; Ayadi, S.; Hammemi, Z.; Jallouli, S.; Trifa, Y.; Amara, H.; Othmani, A.; Rezgui, M.; et al. Effect of salt stress on the flag leaf area and yield components in twenty five durum wheat genotypes (Triticum turgidum ssp. durum). J. New Sci. 2014, 6, 15–23. [Google Scholar]
- Ojeda, V.; Pérez-Ruiz, J.M.; González, M.; Nájera, V.A.; Sahrawy, M.; Serrato, A.J.; Geigenberger, P.; Cejudo, F.J. NADPH Thioredoxin Reductase C and Thioredoxins Act Concertedly in Seedling Development. Plant Physiol. 2017, 174, 1436–1448. [Google Scholar] [CrossRef]
- Li, S.; Chen, N.; Li, F.; Mei, F.; Wang, Z.; Cheng, X.; Kang, Z.; Mao, H. Characterization of wheat homeodomain-leucine zipper family genes and functional analysis of TaHDZ5-6A in drought tolerance in transgenic Arabidopsis. BMC Plant Biol. 2020, 20, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Harris, J.C.; Sornaraj, P.; Taylor, M.; Bazanova, N.; Baumann, U.; Lovell, B.; Langridge, P.; Lopato, S.; Hrmova, M. Molecular interactions of the γ-clade homeodomain-leucine zipper class I transcription factors during the wheat response to water deficit. Plant Mol. Biol. 2016, 90, 435–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ke, S.; Liu, S.; Luan, X.; Xie, X.M.; Hsieh, T.F.; Zhang, X.Q. Mutation in a putative glycosyltransferase-like gene causes programmed cell death and early leaf senescence in rice. Rice 2019, 12, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, F.; Cui, Y.; Zhang, J.; Du, X.; Hu, Y.; Yan, Y.; Mostofa, G.; Bajguz, A.; Yang, J.; Duan, W.; et al. Genome-Wide Identification and Characterisation of Wheat MATE Genes Reveals Their Roles in Aluminium Tolerance. Int. J. Mol. Sci. 2022, 23, 4418. [Google Scholar] [CrossRef]
- Nabi, R.B.S.; Tayade, R.; Imran, Q.M.; Hussain, A.; Shahid, M.; Yun, B.W. Functional Insight of Nitric-Oxide Induced DUF Genes in Arabidopsis thaliana. Front. Plant Sci. 2020, 11, 1041. [Google Scholar] [CrossRef] [PubMed]
- He, X.; Hou, X.; Shen, Y.; Huang, Z. TaSRG, a wheat transcription factor, significantly affects salt tolerance in transgenic rice and Arabidopsis. FEBS Lett. 2011, 585, 1231–1237. [Google Scholar] [CrossRef] [Green Version]
- Voss-Fels, K.P.; Robinson, H.; Mudge, S.R.; Richard, C.; Newman, S.; Wittkop, B.; Stahl, A.; Friedt, W.; Frisch, M.; Gabur, I.; et al. VERNALIZATION1 Modulates Root System Architecture in Wheat and Barley. Mol. Plant 2018, 11, 226. [Google Scholar] [CrossRef] [Green Version]
- Alvarez-Buylla, E.R.; García-Ponce, B.; de la Sánchez, M.P.; Espinosa-Soto, C.; García-Gómez, M.L.; Piñeyro-Nelson, A.; Garay-Arroyo, A. MADS-box genes underground becoming mainstream: Plant root developmental mechanisms. New Phytol. 2019, 223, 1143–1158. [Google Scholar] [CrossRef] [Green Version]
- Castilhos, G.; Lazzarotto, F.; Spagnolo-Fonini, L.; Bodanese-Zanettini, M.H.; Margis-Pinheiro, M. Possible roles of basic helix-loop-helix transcription factors in adaptation to drought. Plant Sci. 2014, 223, 1–7. [Google Scholar] [CrossRef]
- Yoo, C.M.; Wen, J.; Motes, C.M.; Sparks, J.A.; Blancaflor, E.B. A Class I ADP-Ribosylation Factor GTPase-Activating Protein Is Critical for Maintaining Directional Root Hair Growth in Arabidopsis. Plant Physiol. 2008, 147, 1659–1674. [Google Scholar] [CrossRef]
- Alahmad, S.; El Hassouni, K.; Bassi, F.M.; Dinglasan, E.; Youssef, C.; Quarry, G.; Aksoy, A.; Mazzucotelli, E.; Juhász, A.; Able, J.A.; et al. A major root architecture QTL responding to water limitation in durum wheat. Front. Plant Sci. 2019, 10, 436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suetsugu, N.; Higa, T.; Kong, S.G.; Wada, M. PLASTID MOVEMENT IMPAIRED1 and PLASTID MOVEMENT IMPAIRED1-RELATED1 Mediate Photorelocation Movements of Both Chloroplasts and Nuclei. Plant Physiol. 2015, 169, 1155–1167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qi, T.; Wang, J.; Sun, Q.; Day, B.; Guo, J.; Ma, Q. TaARPC3, contributes to wheat resistance against the stripe rust fungus. Front. Plant Sci. 2017, 8, 1245. [Google Scholar] [CrossRef] [Green Version]
- Pandian, B.A.; Sathishraj, R.; Djanaguiraman, M.; Prasad, P.V.V.; Jugulam, M. Role of Cytochrome P450 Enzymes in Plant Stress Response. Antioxidants 2020, 9, 454. [Google Scholar] [CrossRef] [PubMed]
- Buey, R.M.; Ledesma-Amaro, R.; Balsera, M.; de Pereda, J.M.; Revuelta, J.L. Increased riboflavin production by manipulation of inosine 5′-monophosphate dehydrogenase in Ashbya gossypii. Appl. Microbiol. Biotechnol. 2015, 99, 9577–9589. [Google Scholar] [CrossRef]
- Meena, M.; Divyanshu, K.; Kumar, S.; Swapnil, P.; Zehra, A.; Shukla, V.; Yadav, M.; Upadhyay, R.S. Regulation of L-proline biosynthesis, signal transduction, transport, accumulation and its vital role in plants during variable environmental conditions. Heliyon 2019, 5, e02952. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singh, A.K.; Kumar, R.; Pareek, A.; Sopory, S.K.; Singla-Pareek, S.L. Overexpression of rice CBS domain containing protein improves salinity, oxidative, and heavy metal tolerance in transgenic tobacco. Mol. Biotechnol. 2012, 52, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Lawlor, D.W.; Paul, M.J. Source/sink interactions underpin crop yield: The case for trehalose 6-phosphate/SnRK1 in improvement of wheat. Front. Plant Sci. 2014, 5, 418. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ma, H. Step-wise and lineage-specific diversification of plant RNA polymerase genes and origin of the largest plant-specific subunits. New Phytol. 2015, 207, 1198–1212. [Google Scholar] [CrossRef]
- Krattinger, S.G.; Kang, J.; Bräunlich, S.; Boni, R.; Chauhan, H.; Selter, L.L.; Robinson, M.D.; Schmid, M.W.; Wiederhold, E.; Hensel, G.; et al. Abscisic acid is a substrate of the ABC transporter encoded by the durable wheat disease resistance gene Lr34. New Phytol. 2019, 223, 853–866. [Google Scholar] [CrossRef] [PubMed]
- Gietler, M.; Nykiel, M.; Orzechowski, S.; Fettke, J.; Zagdańska, B. Protein carbonylation linked to wheat seedling tolerance to water deficiency. Environ. Exp. Bot. 2017, 137, 84–95. [Google Scholar] [CrossRef]
- Fucile, G.; Falconer, S.; Christendat, D. Evolutionary Diversification of Plant Shikimate Kinase Gene Duplicates. PLoS Genet. 2008, 4, e1000292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trusov, Y.; Botella, J.R. Plant G-proteins come of age: Breaking the bond with animal models. Front. Chem. 2016, 4, 24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, Y.; Sakamoto, W. FtsH protease in the thylakoid membrane: Physiological functions and the regulation of protease activity. Front. Plant Sci. 2018, 9, 855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sengupta, D.; Naik, D.; Reddy, A.R. Plant aldo-keto reductases (AKRs) as multi-tasking soldiers involved in diverse plant metabolic processes and stress defense: A structure-function update. J. Plant Physiol. 2015, 179, 40–55. [Google Scholar] [CrossRef] [PubMed]
- Tejos, R.; Rodriguez-Furlán, C.; Adamowski, M.; Sauer, M.; Norambuena, L.; Friml, J. PATELLINS are regulators of auxin-mediated PIN1 relocation and plant development in Arabidopsis thaliana. J. Cell Sci. 2018, 131, jcs204198. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Xiao, J.; Li, Y.; Su, B.; Xu, H.; Shan, X.; Song, C.; Xie, J.; Li, R. PDM3, a pentatricopeptide repeat-containing protein, affects chloroplast development. J. Exp. Bot. 2017, 68, 5615–5627. [Google Scholar] [CrossRef]
- Gahlaut, V.; Jaiswal, V.; Kumar, A.; Gupta, P.K. Transcription factors involved in drought tolerance and their possible role in developing drought tolerant cultivars with emphasis on wheat (Triticum aestivum L.). Theor. Appl. Genet. 2016, 129, 2019–2042. [Google Scholar] [CrossRef]
- Furutani, M.; Hirano, Y.; Nishimura, T.; Nakamura, M.; Taniguchi, M.; Suzuki, K.; Oshida, R.; Kondo, C.; Sun, S.; Kato, K.; et al. Polar recruitment of RLD by LAZY1-like protein during gravity signaling in root branch angle control. Nat. Commun. 2020, 11, 1–3. [Google Scholar] [CrossRef] [Green Version]
- Grzesiak, M.T.; Hordyńska, N.; Maksymowicz, A.; Grzesiak, S.; Szechyńska-Hebda, M. Variation among Spring Wheat (Triticum aestivum L.) Genotypes in Response to the Drought Stress. II—Root System Structure. Plants 2019, 8, 584. [Google Scholar] [CrossRef] [PubMed]
- Almeida, D.M.; Gregorio, G.B.; Oliveira, M.M.; Saibo, N.J.M. Five novel transcription factors as potential regulators of OsNHX1 gene expression in a salt tolerant rice genotype. Plant Mol. Biol. 2016, 93, 61–77. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.J.; Yuan, S.; Zhang, N.H.; Lei, T.; Duan, H.G.; Liang, H.G.; Lin, H.H. Effect of water stress on photosystem 2 in two wheat cultivars. Biol. Plant. 2006, 50, 597–602. [Google Scholar] [CrossRef]
- Nigro, D.; Blanco, A.; Piarulli, L.; Signorile, M.A.; Colasuonno, P.; Blanco, E.; Simeone, R. Fine Mapping and Candidate Gene Analysis of Pm36, a Wild Emmer-Derived Powdery Mildew Resistance Locus in Durum Wheat. Int. J. Mol. Sci. 2022, 23, 13659. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.; Markham, J.E.; Dietrich, C.R.; Jaworski, J.G.; Cahoon, E.B. Sphingolipid Long-Chain Base Hydroxylation Is Important for Growth and Regulation of Sphingolipid Content and Composition in Arabidopsis. Plant Cell 2008, 20, 1862–1878. [Google Scholar] [CrossRef] [Green Version]
- Mondini, L.; Nachit, M.; Porceddu, E.; Pagnotta, M.A. Identification of SNP mutations in DREB1, HKT1, and WRKY1 genes involved in drought and salt stress tolerance in durum wheat (Triticum turgidum L. var durum). Omi. A J. Integr. Biol. 2012, 16, 178–187. [Google Scholar] [CrossRef]
- Mondini, L.; Nachit, M.M.; Pagnotta, M.A. Allelic variants in durum wheat (Triticum turgidum L. var. durum) DREB genes conferring tolerance to abiotic stresses. Mol. Genet. Genom. 2015, 290, 531–544. [Google Scholar] [CrossRef]
- GitHub. Taiyun/Corrplot: A visual Exploratory Tool on Correlation Matrix. Available online: https://github.com/taiyun/corrplot (accessed on 23 December 2021).
- Vincent, Q.V. GitHub—Vqv/Ggbiplot: A Biplot Based on ggplot2. Available online: https://github.com/vqv/ggbiplot (accessed on 23 December 2021).
- Lewis, P.O. Genetic Data Analysis: Computer Program for the Analysis of Allelic Data. Version 1.0 (d16c). Available online: https://plewis.github.io/software/ (accessed on 20 December 2022).
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
Figure 1.
Correlation matrix among all the recorded traits in pots experiment under C-control and S-salt conditions. Positive (in green) and negative (in red) indicate significant (p < 0.05) correlations. For abbreviations, please see the abbreviation list at the end of the paper.
Figure 1.
Correlation matrix among all the recorded traits in pots experiment under C-control and S-salt conditions. Positive (in green) and negative (in red) indicate significant (p < 0.05) correlations. For abbreviations, please see the abbreviation list at the end of the paper.

Figure 2.
Biplot of the principal component analysis (A) and hierarchical analysis (B) of the shoot and root traits under control conditions. For abbreviations, please see the abbreviation list at the end of the paper.
Figure 2.
Biplot of the principal component analysis (A) and hierarchical analysis (B) of the shoot and root traits under control conditions. For abbreviations, please see the abbreviation list at the end of the paper.

Figure 3.
Biplot of the principal component analysis (A) and hierarchical analysis (B) of the shoot and root traits under salt conditions. For abbreviations, please see the abbreviation list at the end of the paper.
Figure 3.
Biplot of the principal component analysis (A) and hierarchical analysis (B) of the shoot and root traits under salt conditions. For abbreviations, please see the abbreviation list at the end of the paper.

Figure 4.
UPGMA phenogram of the Nei genetic distance among six durum wheat genotypes based on 11 SSRs markers.
Figure 4.
UPGMA phenogram of the Nei genetic distance among six durum wheat genotypes based on 11 SSRs markers.

Figure 5.
Experimental model, indicating when salt stress was applied by adding NaCl to the water nutrition solution.
Figure 5.
Experimental model, indicating when salt stress was applied by adding NaCl to the water nutrition solution.

Figure 6.
The measurements of morphological root traits (A) using Win-RHIZO system and root angle (B) using ImageJ (°).
Figure 6.
The measurements of morphological root traits (A) using Win-RHIZO system and root angle (B) using ImageJ (°).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Salt versus control effect on shoot traits of six durum wheat genotypes.
| Genotype | Seedling Length, cm | Number of Leaves | Number of Tillers | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Control | Salt | Diff. (%) | Control | Salt | Diff. (%) | Control | Salt | Diff. (%) | |
| Azeghar | 34.6 ± 3.2 b | 40.5 ± 1.6 a | 16.96 ns | 5.0 ± 0.0 c | 6.7 ± 0.6 c | 33.3 ns | 1.0 ± 0.0 c | 1.0 ± 0.0 b | 0.0 ns |
| Cham1 | 37.1 ± 0.6 ab | 31.3 ± 0.8 bc | −15.71 ns | 6.0 ± 1.7 c | 6.3 ± 1.2 c | 5.6 ns | 2.3 ± 0.6 b | 1.0 ± 0.0 b | −57.1 ** |
| J. Khetifa | 42.8 ± 1.1 a | 33.1 ± 2.6 b | −22.74 * | 9.3 ± 0.6 a | 14.0 ±0.0 a | 50.0 *** | 3.0 ± 0.0 ab | 3.3 ± 0.6 a | 11.1 ns |
| Pelsodur | 34.0 ± 1.5 b | 29.2 ± 2.2 c | −14.18 ns | 8.0 ± 0.0 ab | 7.3 ± 1.5 c | −8.3 ns | 3.3 ± 0.6 a | 1.3 ± 0.6 b | −60.0 *** |
| Sebatel | 36.4 ± 2.8 b | 34.8 ± 0.8 b | −4.40 ns | 5.7 ± 0.6 c | 7.3 ± 0.6 c | 29.4 ns | 1.0 ± 0.0 c | 1.0 ± 0.0 b | 0.0 ns |
| Vulci | 35.2 ± 7.0 b | 27.6 ± 2.4 c | −21.76 ns | 6.7 ± 1.5 bc | 11.3 ± 1.2 b | 70.0 *** | 2.3 ± 0.6 b | 2.7 ± 0.6 a | 14.3 ns |
| Genotype | ** | *** | *** | ||||||
| Treatment | *** | *** | ** | ||||||
| G × T | ** | *** | *** | ||||||
Values are means ± standard deviations (n = 3). Means with the same letter in the column of each treatment are not significantly different between genotypes (p < 0.05) (LSD test). Diff.% reports the differences between Control (column) and Salt (column). It is computed as [Diff.% = ((salt/control) − 1) × 100], and its statistic is indicated as ns—Not significant; *, **, and *** indicate significance at p < 0.05, p < 0.01, and p < 0.001 levels, respectively.
Table 2.
Analysis of variance (ANOVA) for root traits.
| ANOVA | Root Volume | Root Length | Tips | Root Surface Area |
| Genotype | * | * | * | ** |
| Treatments | *** | *** | *** | *** |
| G × T | ns | ns | * | ns |
| ANOVA | Root Angle | Crossings | Forks | Average Root Diameter |
| Genotype | *** | * | ** | ns |
| Treatments | *** | * | ** | ns |
| G × T | *** | ns | ns | ns |
A description is added, i.e., ns—Not significant; *, **, and *** indicate significance at p < 0.05, p < 0.01, and p < 0.001 levels, respectively.
Table 3.
Salt versus control effect on root traits of six durum wheat genotypes.
| Trait | Treat | Genotype | |||||
|---|---|---|---|---|---|---|---|
| Azeghar | Cham1 | J. Khetifa | Pelsodur | Sebatel | Vulci | ||
| Root volume (cm3) | control | 0.80 ± 0.28 a | 0.57 ± 0.15 a | 0.57 ± 0.04 a | 0.61 ± 0.19 a | 0.68 ± 0.18 a | 0.48 ± 0.18 a |
| salt | 1.28 ± 0.24 a | 0.57 ± 0.02 c | 1.07 ± 0.34 ab | 0.68 ± 0.06 bc | 0.99 ± 0.15 abc | 0.77 ± 0.29 bc | |
| Diff. (%) | 59.6 * | 0.0 ns | 88.8 ** | 12.4 ns | 45.1 ns | 61.5 ns | |
| Root Length (cm) | control | 684 ± 106 a | 333 ± 73 c | 475 ± 104 bc | 521 ± 80 abc | 578 ± 55 ab | 530 ± 120 ab |
| salt | 826 ± 52 ab | 603 ± 23 b | 890 ± 192 a | 590 ± 7 b | 737 ± 200 ab | 627 ± 155 b | |
| Diff. (%) | 20.6 ns | 81.2 * | 87.3 ** | 13.4 ns | 27.6 ns | 18.2 ns | |
| Number of tips | control | 1243 ± 287 a | 633 ± 94 a | 1188 ± 406 a | 1214 ± 225 a | 1326 ± 507 a | 1272 ± 353 a |
| salt | 1510 ± 119 b | 1362 ± 21 b | 2399 ± 143 a | 1401 ± 265 b | 1480 ± 352 b | 1311 ± 309 b | |
| Diff. (%) | 21.5 ns | 115.3 * | 101.9 *** | 15.4 ns | 11.6 ns | 3.0 ns | |
| Root surface area (cm2) | control | 81.4 ± 7.8 a | 48.5 ± 11.8 b | 57.7 ± 4.2 b | 62.8 ± 14.5 ab | 69.9 ± 11.6 ab | 56.4 ± 16.5 b |
| salt | 114.9 ± 14 a | 65.5 ± 0.0 c | 109 ± 28 ab | 71.1 ± 2.6 bc | 95.3 ± 20.3 abc | 77.2 ± 21.7 bc | |
| Diff. (%) | 41.2 * | 35.1 ns | 89.0 ** | 13.2 ns | 36.3 ns | 37.0 ns | |
| Root angle (°) | control | 110.1 ± 4.4 a | 109.4 ± 7.4 a | 90.1 ± 3.3 b | 79.7 ± 1.3 c | 106.1 ± 2.3 a | 84.9 ± 2.8 bc |
| salt | 117.0 ± 7.9 a | 104.5 ± 3.2 bc | 95.4 ± 4.3 d | 109.5 ± 6.6 ab | 115.3 ± 2.7 a | 96.7 ± 2.9 cd | |
| Diff. (%) | 6.2 ns | −4.5 ns | 6.0 ns | 37.3 *** | 8.7 * | 13.9 * | |
| Crossings | control | 956 ± 487 a | 421 ± 162 a | 942 ± 204 a | 887 ± 62 a | 857 ± 136 a | 859 ± 218 a |
| salt | 1056 ± 97 abc | 512 ± 315 c | 1543 ± 423 a | 981 ± 241 abc | 1122 ± 503 ab | 916 ± 266 bc | |
| Diff. (%) | 10.5 ns | 21.8 ns | 63.9 * | 10.6 ns | 30.9 ns | 6.6 ns | |
| Forks | control | 4990 ± 914 a | 2774 ± 542 b | 4651 ± 511 a | 4736 ± 531 a | 4587 ± 897 a | 4152 ± 931 ab |
| salt | 6606 ± 143 ab | 3547 ± 242 c | 8180 ± 1987 a | 4801 ± 183 bc | 6100 ± 2120 abc | 5111 ± 1201 bc | |
| 32.4 ns | 27.9 ns | 75.9 * | 1.4 ns | 33.0 ns | 23.1 ns | ||
| Mean diameter (mm) | control | 0.39 ± 0.1 ab | 0.46 ± 0.01 a | 0.39 ± 0.06 ab | 0.38 ± 0.03 ab | 0.38 ± 0.05 ab | 0.33 ± 0.3 b |
| salt | 0.44 ± 0.03 a | 0.35 ± 0.01 b | 0.39 ± 0.03 ab | 0.39 ± 0.02 ab | 0.42 ± 0.03 ab | 0.39 ± 0.06 ab | |
| Diff. (%) | 12.8 ns | −23.9 * | −1.7 ns | 1.3 ns | 8.7 ns | 17.0 ns | |
Values are means ± standard deviations (n = 3). Means with the same letter in each row are not significantly different between genotypes (p < 0.05) (LSD test). The differences between Control and Salt [Diff.% = ((salt/control) − 1)*100] and its statistic is indicated as ns—Not significant; *, **, and *** indicate significance at p < 0.05, p < 0.01, and p < 0.001 levels, respectively.
Table 4.
Genetic diversity in durum wheat based on 11 SSR markers identifying 13 loci.
| Locus | Na | I | Ho | He | uHe | PIC | Size, bp |
|---|---|---|---|---|---|---|---|
| cfa2086 | 5 | 1.561 | 0.000 | 0.778 | 0.848 | 0.744 | 223–299 |
| wms5 | 3 | 1.011 | 0.000 | 0.611 | 0.667 | 0.536 | 180–190 |
| gwm234a | 2 | 0.451 | 0.000 | 0.278 | 0.303 | 0.240 | 214–218 |
| gwm234b | 2 | 0.693 | 0.000 | 0.500 | 0.545 | 0.375 | 242–256 |
| wmc727a | 2 | 0.679 | 0.833 | 0.486 | 0.530 | 0.368 | 88–96 |
| wmc727b | 2 | 0.451 | 0.000 | 0.278 | 0.303 | 0.240 | null-231 |
| cfa2257 | 5 | 1.424 | 0.333 | 0.722 | 0.788 | 0.680 | 104–156 |
| wms205 | 2 | 0.693 | 1.000 | 0.500 | 0.545 | 0.375 | 157–170 |
| gwm427 | 4 | 1.330 | 0.000 | 0.722 | 0.788 | 0.672 | 201–256 |
| gwm573.2 | 5 | 1.424 | 1.000 | 0.722 | 0.788 | 0.680 | 181–240 |
| gwm636 | 3 | 1.011 | 0.000 | 0.611 | 0.667 | 0.536 | 110–131 |
| gwm459a | 3 | 1.011 | 0.000 | 0.611 | 0.667 | 0.536 | 131–153 |
| gwm499b | 4 | 1.330 | 0.000 | 0.722 | 0.788 | 0.672 | 147–197 |
| mean | 3.2 | 1.005 | 0.244 | 0.580 | 0.633 |
Na—No. of Alleles, I—Shannon’s Information Index = −1 * Sum (pi * Ln (pi)), Ho—Observed Heterozygosity, He—Expected Heterozygosity, uHe—Unbiased Expected Heterozygosity, PIC—polymorphism information content. Different locus of same marker are highlighted as “a” and “b”.
Table 5.
Private alleles (defined by a set of populations currently active).
| Locus | Allele | Frequency | Found in |
|---|---|---|---|
| cfa2086 | 223 | 1 | Sebatel |
| cfa2086 | 299 | 1 | Pelsodur |
| cfa2086 | 269 | 1 | Cham |
| cfa2086 | 277 | 1 | J. Khetifa |
| wms5 | 185 | 1 | Cham |
| gwm234a | 214 | 1 | J. Khetifa |
| wmc727a | 1 | 1 | Pelsodur |
| cfa2257 | 121 | 1 | Cham |
| cfa2257 | 142 | 0.5 | Azeghar |
| cfa2257 | 104 | 0.5 | Azeghar |
| gwm427 | 237 | 1 | Sebatel |
| gwm427 | 201 | 1 | Azeghar |
| gwm573.2 | 226 | 0.5 | Pelsodur |
| gwm573.2 | 181 | 0.5 | Azeghar |
| gwm636 | 131 | 1 | Pelsodur |
| gwm459a | 131 | 1 | Vulci |
| gwm499b | 147 | 1 | Sebatel |
| gwm499b | 154 | 1 | J. Khetifa |
Different locus of same marker are highlighted as “a” and “b”.
Table 6.
Primers used for PCR amplification.
| Marker | Primer Forward (5′-3′) | Primer Reverse (5′-3′) | Chr. | AT (°C) |
|---|---|---|---|---|
| cfa2086 | TCTACTTTCAGGGCACCTCG | TCTCTCCAAACCTCCCTGTAA | 2A | 56 |
| gwm573.2 | AAGAGATAACATGCAAGAAA | TTCAAATATGTGGGAACTAC | 7B | 45 |
| wmc727 | CATAATCAGGACAGCCGCAC | TAGTGGCCTGATGTATCTAGTTGG | 5A | 55 |
| wms205 | CGACCCGGTTCACTTCAG | AGTCGCCGTTGTATAGTGCC | 5A | 56 |
| wms5 | GCCAGCTACCTCGATACAACTC | AGAAAGGGCCAGGCTAGTAGT | 3A | 56 |
| gmw459 | ATGGAGTGGTCACACTTTGAA | AGCTTCTCTGACCAACTTCTCG | 6A | 54 |
| gwm234 | GAGTCCTGATGTGAAGCTGTTG | CTCATTGGGGTGTGTACGTG | 5B | 55 |
| gwm427 | AAACTTAGAACTGTAATTTCAGA | AGTGTGTTCATTTGACAGTT | 6A | 45 |
| gwm499 | ACTTGTATGCTCCATTGATTGG | GGGAGTGGAAACTGCATAA | 5B | 52 |
| gwm637 | AAAGAGGTCTGCCGCTAACA | TATACGGTTTTGTGAGGGGG | 4A | 55 |
| cfa2257 | GATACAATAGGTGCCTCCGC | CCATTATGTAAATGCTTCTGTTTGA | 7A | 49 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Urbanavičiūtė, I.; Bonfiglioli, L.; Pagnotta, M.A. Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings. Plants 2023, 12, 412. https://doi.org/10.3390/plants12020412
AMA Style
Urbanavičiūtė I, Bonfiglioli L, Pagnotta MA. Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings. Plants. 2023; 12(2):412. https://doi.org/10.3390/plants12020412
Chicago/Turabian StyleUrbanavičiūtė, Ieva, Luca Bonfiglioli, and Mario A. Pagnotta. 2023. "Phenotypic and Genotypic Diversity of Roots Response to Salt in Durum Wheat Seedlings" Plants 12, no. 2: 412. https://doi.org/10.3390/plants12020412
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.