
Assessment of Maternal Effects and Genetic Variability in Resistance to Verticillium dahliae in Olive Progenies

 , ,
, ,
Abstract
:1. Introduction
2. Results
2.1. Seed Germination Rate
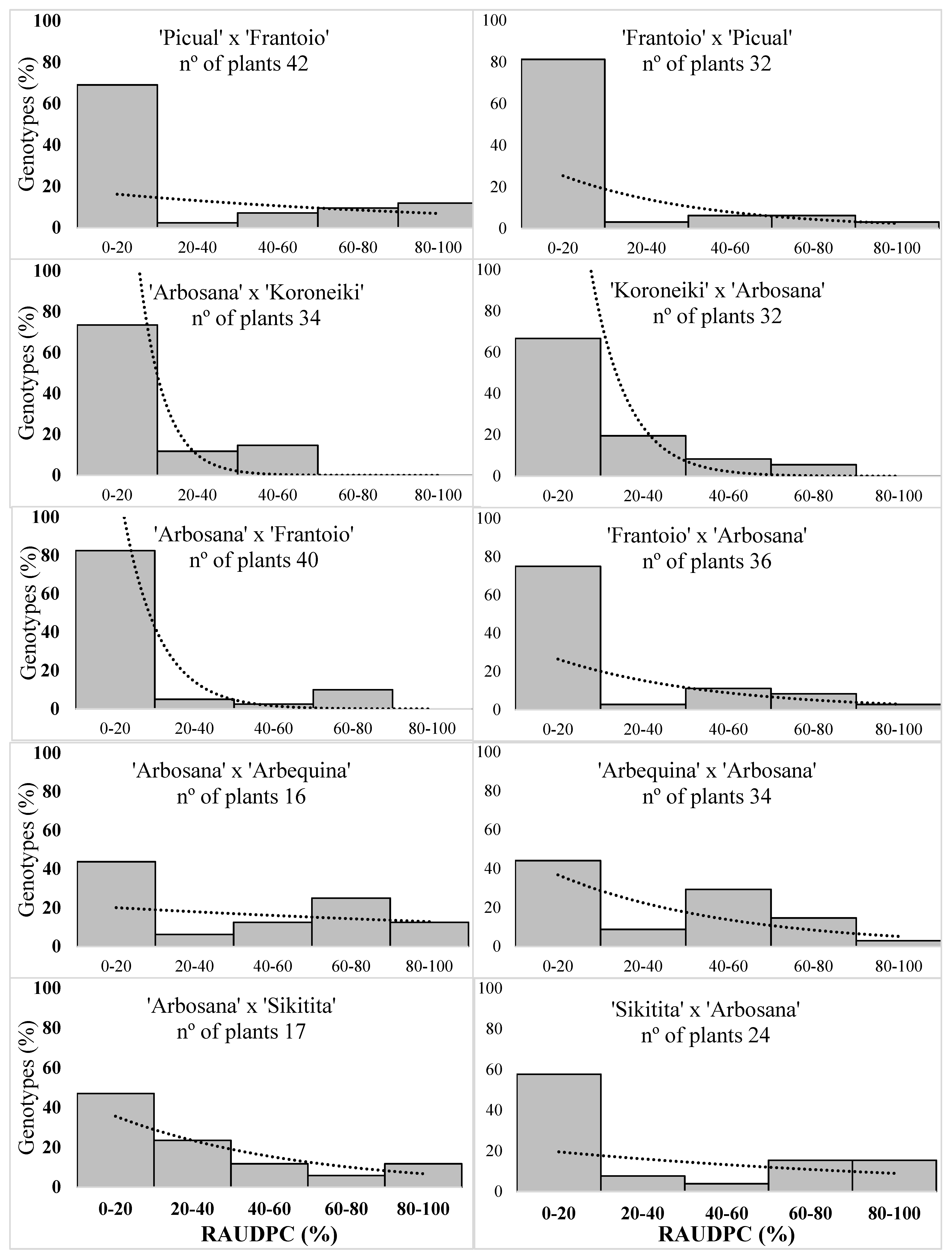
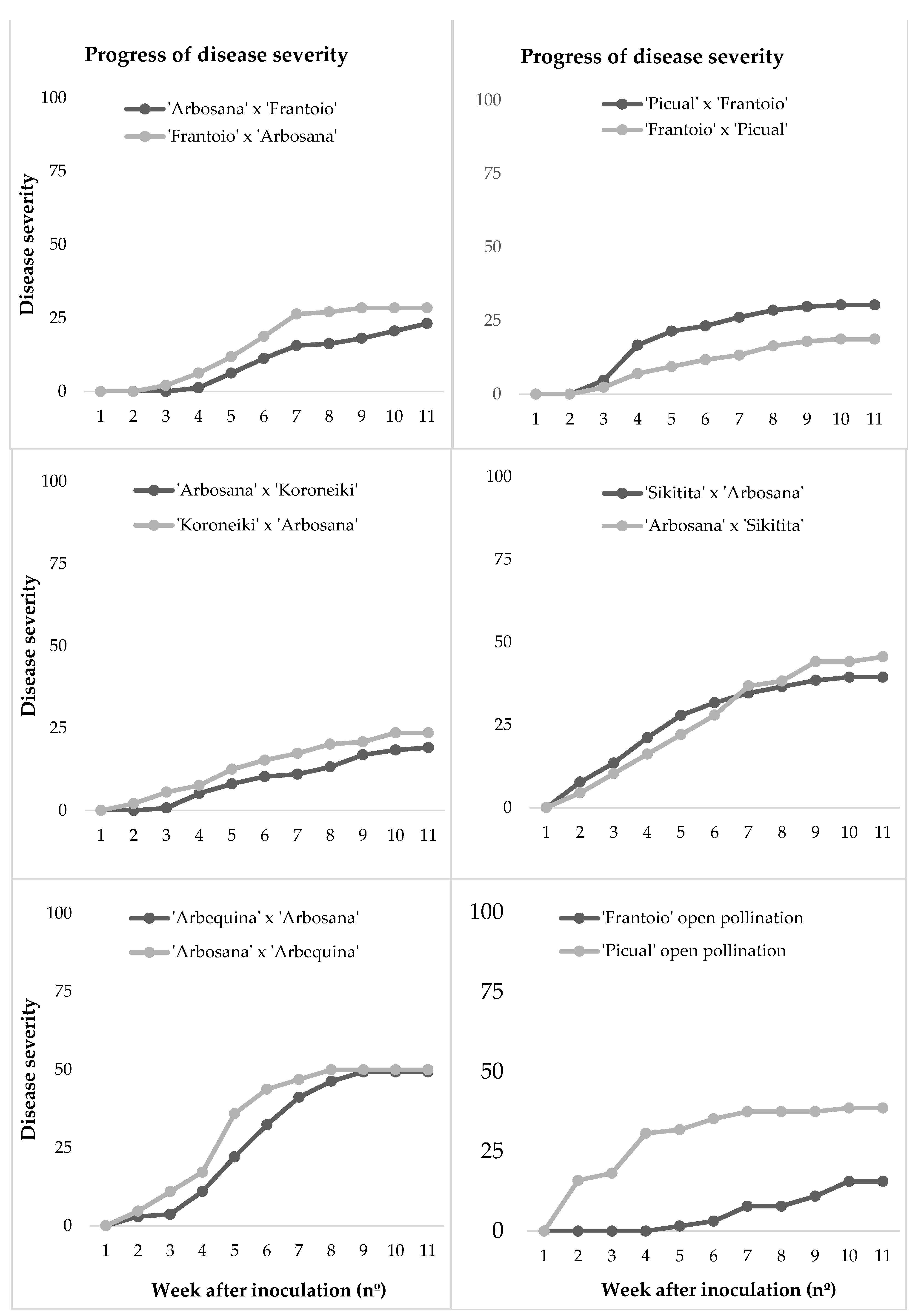
2.2. Symptom and Disease Parameters
2.3. Seedling Growth
2.4. Germination Rate and Resistance Level among Progenies
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Fungal Material and Plant Inoculation
4.3. Fungus Isolation
4.4. Experimental Design
4.5. Disease Evaluation
4.6. Plant Growth Evaluation
4.7. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barranco, D.; Fernandez-Escobar, R.; Rallo, L. Capítulo 3: Variedades y patrones. In El Cultivo del Olivo; Ediciones Mundi-Prensa: Madrid, Spain, 2017; pp. 65–95. [Google Scholar]
- FAO. FAOSTAT, Production Statistics. Rome, Italy. Available online: www.fao.org/faostat (accessed on 8 October 2020).
- López-Escudero, F.J.; Mercado-Blanco, J. Verticillium wilt of olive: A case study to implement an integrated strategy to control a soil-borne pathogen. Plant Soil 2010, 344, 1–50. [Google Scholar] [CrossRef] [Green Version]
- Wilhelm, S.; Taylor, J.B. Control of Verticillium wilt of olive through natural recovery and resistance. Phytopathology 1965, 55, 310–316. [Google Scholar]
- Colella, C.; Miacola, C.; Amenduni, M.; D’Amico, M.; Bubici, G.N.; Cirulli, M. Sources of verticillium wilt resistance in wild olive germplasm from the Mediterranean region. Plant Pathol. 2008, 57, 533–539. [Google Scholar] [CrossRef]
- Lopez-Escudero, F.J.; Blanco-Lopez, M.A. Relationship between the inoculum density of Verticillium dahliae and the pro-gress of Verticillium wilt of olive. Plant Dis. 2007, 91, 1372–1378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Ruiz, G.M.; Trapero, C.; Varo-Suarez, A.; Trapero, A.; Javier López-Escudero, F.J. Identifying resistance to Verti-cillium wilt in local Spanish olive cultivars. Phytopathol. Mediterr. 2015, 54, 453–460. [Google Scholar] [CrossRef]
- Trapero, C.; Serrano, N.; Arquero, O.; Del Río, C.; López-Escudero, F.J. Field Resistance to Verticillium Wilt in Selected Olive Cultivars Grown in Two Naturally Infested Soils. Plant Dis. 2013, 97, 668–674. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trapero, C.; Díez, C.M.; Rallo, L.; Barranco, D.; López-Escudero, F.J. Effective inoculation methods to screen for resistance to Verticillium wilt in olive. Sci. Hortic. 2013, 162, 252–259. [Google Scholar] [CrossRef]
- Arias-Calderón, R.; León, L.; Bejarano-Alcázar, J.; Belaj, A.; de la Rosa, R.; Rodríguez-Jurado, D. Resistance to Verticillium wilt in olive progenies from open-pollination. Sci. Hortic. 2015, 185, 34–42. [Google Scholar] [CrossRef]
- Trapero, C.; Rallo, L.; Lopez-Escudero, F.J.; Barranco, D.; Diez, C.M. Variability and selection of verticillium wilt resistant genotypes in cultivated olive and in the Olea genus. Plant Pathol. 2015, 64, 890–900. [Google Scholar] [CrossRef]
- Wolf, J.B.; Wade, M.J. What are maternal effects (and what are they not)? Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1107–1115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, K.H.; Lai, Y.C.; Chang, K.Y.; Chen, Y.F.; Hwang, S.Y.; Lo, H.F. Improving breeding efficiency for quality and yield of sweetpotato. Bot. Stud. 2007, 48, 283–292. [Google Scholar]
- Rukundo, P.; Shimelis, H.; Laing, M.; Gahakwa, D. Combining Ability, Maternal Effects, and Heritability of Drought Tolerance, Yield and Yield Components in Sweetpotato. Front. Plant Sci. 2017, 7, 1981. [Google Scholar] [CrossRef] [Green Version]
- Gonzalo, M.; Vyn, T.; Holland, J.; McIntyre, L.M. Mapping reciprocal effects and interactions with plant density stress in Zea mays L. Heredity 2007, 99, 14–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maris, M. Analysis of an incomplete diallel cross among three ssp. tuberosum varieties and seven long-day adapted ssp. andigena clones of the potato (Solanum tuberosum L.). Euphytica 1989, 41, 163–182. [Google Scholar] [CrossRef]
- A Roach, D.; Wulff, R.D. Maternal Effects in Plants. Annu. Rev. Ecol. Syst. 1987, 18, 209–235. [Google Scholar] [CrossRef]
- Donohue, K. Completing the cycle: Maternal effects as the missing link in plant life histories. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 1059–1074. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lawson, H.; Cheverud, J.M.; Wolf, J. Genomic imprinting and parent-of-origin effects on complex traits. Nat. Rev. Genet. 2013, 14, 609–617. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, J.A.; Zilberman, D. Evolution and function of genomic imprinting in plants. Genes Dev. 2015, 29, 2517-31. [Google Scholar] [CrossRef]
- Jahnke, S.; Scholten, S. Epigenetic Resetting of a Gene Imprinted in Plant Embryos. Curr. Biol. 2009, 19, 1677–1681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kermicle, J.L. Dependence of the r-mottled aleurone phenotype in maize on mode of sexual transmission. Genetics 1970, 66, 69–85. [Google Scholar] [CrossRef]
- Piskurewicz, U.; Iwasaki, M.; Susaki, D.; Megies, C.; Kinoshita, T.; Lopez-Molina, L. Dormancy-specific imprinting underlies maternal inheritance of seed dormancy in Arabidopsis thaliana. Plant Biol. 2016, 5, e19573. [Google Scholar] [CrossRef]
- Pring, D.R.; Lonsdale, D.M. Cytoplasmic male sterility and maternal inheritance of disease susceptibility in maize. Annu. Rev. Phytopathol. 1989, 27, 483–502. [Google Scholar] [CrossRef]
- Sims, E.L.; Triplett, J.K. Costs and benefits of plant responses to disease: Resistance and tolerance. Evolution 1995, 48, 1973–1985. [Google Scholar] [CrossRef]
- Habarurema, I.; Asea, G.; Lamo, J.; Gibson, P.; Edema, R.; Séré, Y.; Onasanya, R.O. Genetic analysis of resistance to rice bacterial blight in Uganda. Afr. Crop Sci. J. 2012, 20, 105–112. [Google Scholar]
- Vivas, M.; Zas, R.; Sampedro, L.; Solla, A. Environmental Maternal Effects Mediate the Resistance of Maritime Pine to Biotic Stress. PLoS ONE 2013, 8, e70148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beyene, A.T.; Derera, J.; Sibiya, J.; Fikre, A. Gene action determining grain yield and chocolate spot (Botrytis fabae) resistance in faba bean. Euphytica 2015, 207, 293–304. [Google Scholar] [CrossRef]
- Welz, H.G.; Geiger, H.H. Genes for resistance to northern corn leaf blight in diverse maize populations. Plant Breed. 2000, 119, 1–14. [Google Scholar] [CrossRef]
- Rius, F.J.; Lacarte, J.M. La Revolución del Olivar: El Cultivo en Seto, 2nd ed.; Editorial Paraninfo: Madrid, Spain, 2015; ISBN 13: 9780646938646. [Google Scholar]
- Rallo, L.; Barranco, D.; Díez, C.M.; Rallo, P.; Suárez, M.P.; Trapero, C.; Pliego-Alfaro, F. Strategies for Olive (Olea europaea L.) Breeding: Cultivated Genetic Resources and Crossbreeding. In Advances in Plant Breeding Strategies: Fruit; Springer: Cham, Switzerland, 2018. [Google Scholar] [CrossRef]
- Serrano, A.; Jurado, M.D.R.; Román, B.; Bejarano-Alcázar, J.; de la Rosa, R.; León, L. Verticillium Wilt Evaluation of Olive Breeding Selections Under Semi-Controlled Conditions. Plant Dis. 2021. [Google Scholar] [CrossRef]
- Liu, L.; Chen, C.-X.; Zhu, Y.-F.; Xue, L.; Liu, Q.-W.; Qi, K.-J.; Zhang, S.-L.; Wu, J. Maternal inheritance has impact on organic acid content in progeny of pear (Pyrus spp.) fruit. Euphytica 2016, 209, 305–321. [Google Scholar] [CrossRef]
- Palasciano, M.; Camposeo, S.; Ferrara, G.; Godini, A. Pollen production by popular olive cultivars. Acta Hortic. 2008, 489–492. [Google Scholar] [CrossRef]
- Díez, C.M.; Moral, J.; Cabello, D.; Morello, P.; Rallo, L.; Barranco, D. Cultivar and Tree Density As Key Factors in the Long-Term Performance of Super High-Density Olive Orchards. Front. Plant Sci. 2016, 7, 1226. [Google Scholar] [CrossRef] [Green Version]
- Alcalá, A.; Barranco, D. Prediction of Flowering Time in Olive for the Cordoba Olive Collection. HortScience 1992, 27, 1205–1207. [Google Scholar] [CrossRef] [Green Version]
- Kirkbride, R.C.; Lu, J.; Zhang, C.; Mosher, R.A.; Baulcombe, D.C.; Chen, Z.J. Maternal small RNAs mediate spatial-temporal regulation of gene expression, imprinting, and seed development in Arabidopsis. Proc. Natl. Acad. Sci. USA 2019, 116, 2761–2766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-Escudero, F.J.; Blanco-López, M.; Rincón, C.D.R.; Reig, J.M.C. Response of Olive Cultivars to Stem Puncture Inoculation with a Defoliating Pathotype of Verticillium dahliae. HortScience 2007, 42, 294–298. [Google Scholar] [CrossRef] [Green Version]
- Martos-Moreno, C.; López-Escudero, F.J.; Blanco-López, M.A. Resistance of Olive Cultivars to the Defoliating Pathotype of Verticillium dahliae. HortScience 2006, 41, 1313–1316. [Google Scholar] [CrossRef] [Green Version]
- Trujillo, I.; Ojeda, M.A.; Urdiroz, N.M.; Potter, D.; Barranco, D.; Rallo, L.; Diez, C.M. Identification of the Worldwide Olive Germplasm Bank of Córdoba (Spain) using SSR and morphological markers. Tree Genet. Genomes 2013, 10, 141–155. [Google Scholar] [CrossRef]
- Santos-Antunes, F.; Leon, L.; de la Rosa, R.; Alvarado, J.; Mohedo, A.; Trujillo, I.; Rallo, L. The length of the juvenile pe-riod in olive as influenced by vigor of the seedlings and the precocity of the parents. Hortscience 2005, 40, 1213–1215. [Google Scholar] [CrossRef] [Green Version]
- Díaz, A.; De La Rosa, R.; Rallo, P.; Muñoz-Diez, C.; Trujillo, I.; Barranco, D.; Martín, A.; Belaj, A.; Morillo, P.R. Selections of an Olive Breeding Program Identified by Microsatellite Markers. Crop. Sci. 2007, 47, 2317–2322. [Google Scholar] [CrossRef]
- Blanco-López, M.A.; Jimenez-Diaz, R.M.; Caballero, J.M. Symptomatology, incidence and distribution of Verticillium wilt of olive tree in Andalucía. Phytopathol. Mediterr. 1984, 23, 1–8. [Google Scholar]
- Valverde, P.; Trapero, C.; Arquero, O.; Serrano, N.; Barranco, D.; Díez, C.M.; López-Escudero, F.J. Highly infested soils undermine the use of resistant olive rootstocks as a control method of verticillium wilt. Plant Pathol. 2020, 70, 144–153. [Google Scholar] [CrossRef]
- Campbell, C.L.; Madden, L.V. Introduction to Plant Disease Epidemilogy; Willy: Hoboken, NJ, USA, 1990. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Crosses | Seeds | Germination 1 |
|---|---|---|
 | (n°) | (%) |
| ‘Arbosana’ × ‘Koroneiki’ | 104 | 54.8 bcd |
| ‘Koroneiki’ × ‘Arbosana’ | 104 | 56.7 bcd |
| ‘Arbosana’ × ‘Frantoio’ | 104 | 68.3 cd |
| ‘Frantoio’ × ‘Arbosana’ | 104 | 56.7 bcd |
| ‘Picual’ × ‘Frantoio’ | 104 | 76.0 de |
| ‘Frantoio’ × ‘Picual’ | 104 | 62.5 bcd |
| ‘Arbosana’ × ‘Sikitita’ | 104 | 29.8 a |
| ‘Sikitita’ × ‘Arbosana’ | 104 | 42.3 ab |
| ‘Arbosana’ × ‘Arbequina’ | 104 | 29.8 a |
| ‘Arbequina’ × ‘Arbosana’ | 104 | 50.0 abc |
| ‘Frantoio’ open pollination | 52 | 55.8 bcd |
| ‘Picual’ open pollination | 52 | 48.6 ab |
| Mean 2016 | 1144 | 52.6 |
| ‘Picual’ × ‘Frantoio’ | 160 | 56.3 b |
| ‘Frantoio’ × ‘Picual’ | 152 | 36.2 ab |
| ‘Arbosana’ × ‘Sikitita’ | 206 | 39.3 ab |
| ‘Sikitita’ × ‘Arbosana’ | 156 | 23.7 a |
| ‘Frantoio’ open pollination | 81 | 72.8 bc |
| ‘Picual’ open pollination | 93 | 80.6 c |
| Mean 2018 | 848 | 51.5 |
| Total | 1992 | 52.2 |
| Crosses 1 | Seedlings | Disease Incidence 2 | Final Severity 3 | RAUDPC 3 | Mortality 2 | Growth 3 (cm) | |
|---|---|---|---|---|---|---|---|
 | (n) | (%) | (%) | (%) | (%) | Inoculated | Control |
| ‘Arbosana’ × ‘Koroneiki’ | 34 | 32.4 | 19.1 c | 10.4 d | 2.9 | 10.1 cd | 20.6 a |
| ‘Koroneiki’ × ‘Arbosana’ | 36 | 41.7 | 23.6 bc | 14.4 cd | 5.5 | 9.8 d | 18.0 a |
| ‘Arbosana’ × ‘Frantoio’ | 40 | 35.0 | 23.1 bc | 11.2 d | 12.5 | 11.4 bcd | 18.4 a |
| ‘Frantoio’ × ‘Arbosana’ | 36 | 47.2 | 28.5 abc | 18.2 bcd | 19.4 | 12.5 abcd | 19.0 a |
| ‘Picual’ × ‘Frantoio’ | 42 | 35.7 | 30.3 abc | 21.8 abcd | 26.2 | 12.4 abcd | 16.1 a |
| ‘Frantoio’ × ‘Picual’ | 32 | 21.9 | 18.7 c | 11.8 d | 12.5 | 13.9 abc | 19.5 a |
| ‘Arbosana’ × ‘Sikitita’ | 18 | 58.8 | 44.1 ab | 30.1 abc | 35.3 | 16.0 ab | 18.0 a |
| ‘Sikitita’ × ‘Arbosana’ | 26 | 50.0 | 39.4 abc | 29.8 ab | 34.6 | 15.8 ab | 19.1 a |
| ‘Arbosana’ x’ Arbequina’ | 16 | 62.5 | 49.3 a | 38.1 a | 43.8 | 15.3 ab | 16.4 a |
| ‘Arbequina’ × ‘Arbosana’ | 34 | 61.8 | 50.0 a | 29.1 ab | 35.3 | 12.0 abcd | 16.6 a |
| ‘Frantoio’ free pollination | 16 | 25.0 | 15.62 c | 12.3 d | 6.3 | 16.2 a | 19.3 a |
| ‘Picual’ free pollination | 22 | 45.5 | 37.5 abc | 30.1 ab | 22.7 | 14.0 abc | 21.5 a |
| Mean 2016 | 352 | 43.1 | 31.7 | 21.4 | 21.4 | 13.28 | 18.5 |
| ‘Arbosana’ × ‘Sikitita’ | 36 | 66.7 | 57.5 a | 33.3 a | 44.4 | 13.4 a | 16.9 a |
| ‘Sikitita’ × ‘Arbosana’ | 16 | 56.3 | 52.5 ab | 34.4 a | 50 | 16.5 a | 16.6 a |
| ‘Picual’ × ‘Frantoio’ | 36 | 44.4 | 30.0 bc | 13.5 b | 16.7 | 12.3 a | 19.0 a |
| ‘Frantoio’ × ‘Picual’ | 20 | 57.9 | 50.0 ab | 20.3 ab | 31.6 | 16.3 a | 19.8 a |
| ‘Frantoio’ open pollination | 20 | 21.1 | 17.5 c | 6.9 b | 5.3 | 14.4 a | 16.4 a |
| ‘Picual’ free pollination | 32 | 41.9 | 30.0 bc | 14.4 b | 12.9 | 16.2 a | 16.2 a |
| Mean 2018 | 160 | 48.0 | 39.6 | 20.5 | 26.8 | 14.85 | 17.5 |
| Total | 512 | 44.8 | 34.3 | 21.1 | 23.2 | 13.8 | 18.1 |
| Crosses | Incidence 1 | Final Severity 2 | RAUDPC 2 | Mortality 1 | Germination 1 |
|---|---|---|---|---|---|
 | (%) | (%) | (%) | (%) | (%) |
| ‘Frantoio’ open pollination | 23.1 d | 17.5 bcd | 6.3 b | 7.7 bc | 55.8 abc |
| ‘Arbosana’ × ‘Koroneiki’ | 37.1 bcd | 22.5 cd | 12.8 b | 4.3 c | 49.0 bc |
| ‘Koroneiki’ × ‘Arbosana’ | |||||
| ‘Arbosana’ × ‘Frantoio’ | 40.8 bcd | 25.0 cd | 14.5 b | 15.8 bc | 59.6 ab |
| ‘Frantoio’ × ‘Arbosana’ | |||||
| ‘Picual’ × ‘Frantoio’ | 29.8 cd | 25.0 cd | 17.5 b | 20.3 ab | 69.2 a |
| ‘Frantoio’ × ‘Picual’ | |||||
| ‘Sikitita’ × ‘Arbosana’ | 53.5 ab | 42.5 ab | 29.4 a | 34.9 a | 62.5 ab |
| ‘Arbosana’ × ‘Sikitita’ | |||||
| ‘Picual’ open pollination | 43.4 abc | 37.5 abc | 32.3 a | 21.7 ab | 48.6 bc |
| ‘Arbequina’ × Arbosana | 62.0 a | 50.0 a | 33.1 a | 38.0 a | 36.1 c |
| ‘Arbosana’ × ‘Arbequina’ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valverde Caballero, P.; Trapero Ramírez, C.; Barranco Navero, D.; López-Escudero, F.J.; Gordon Bermúdez-Coronel, A.; Díez, C.M. Assessment of Maternal Effects and Genetic Variability in Resistance to Verticillium dahliae in Olive Progenies. Plants 2021, 10, 1534. https://doi.org/10.3390/plants10081534
Valverde Caballero P, Trapero Ramírez C, Barranco Navero D, López-Escudero FJ, Gordon Bermúdez-Coronel A, Díez CM. Assessment of Maternal Effects and Genetic Variability in Resistance to Verticillium dahliae in Olive Progenies. Plants. 2021; 10(8):1534. https://doi.org/10.3390/plants10081534
Chicago/Turabian StyleValverde Caballero, Pedro, Carlos Trapero Ramírez, Diego Barranco Navero, Francisco J. López-Escudero, Ana Gordon Bermúdez-Coronel, and Concepción Muñoz Díez. 2021. "Assessment of Maternal Effects and Genetic Variability in Resistance to Verticillium dahliae in Olive Progenies" Plants 10, no. 8: 1534. https://doi.org/10.3390/plants10081534