
Polyamine Metabolism Is Involved in the Direct Regeneration of Shoots from Arabidopsis Lateral Root Primordia


 and
and 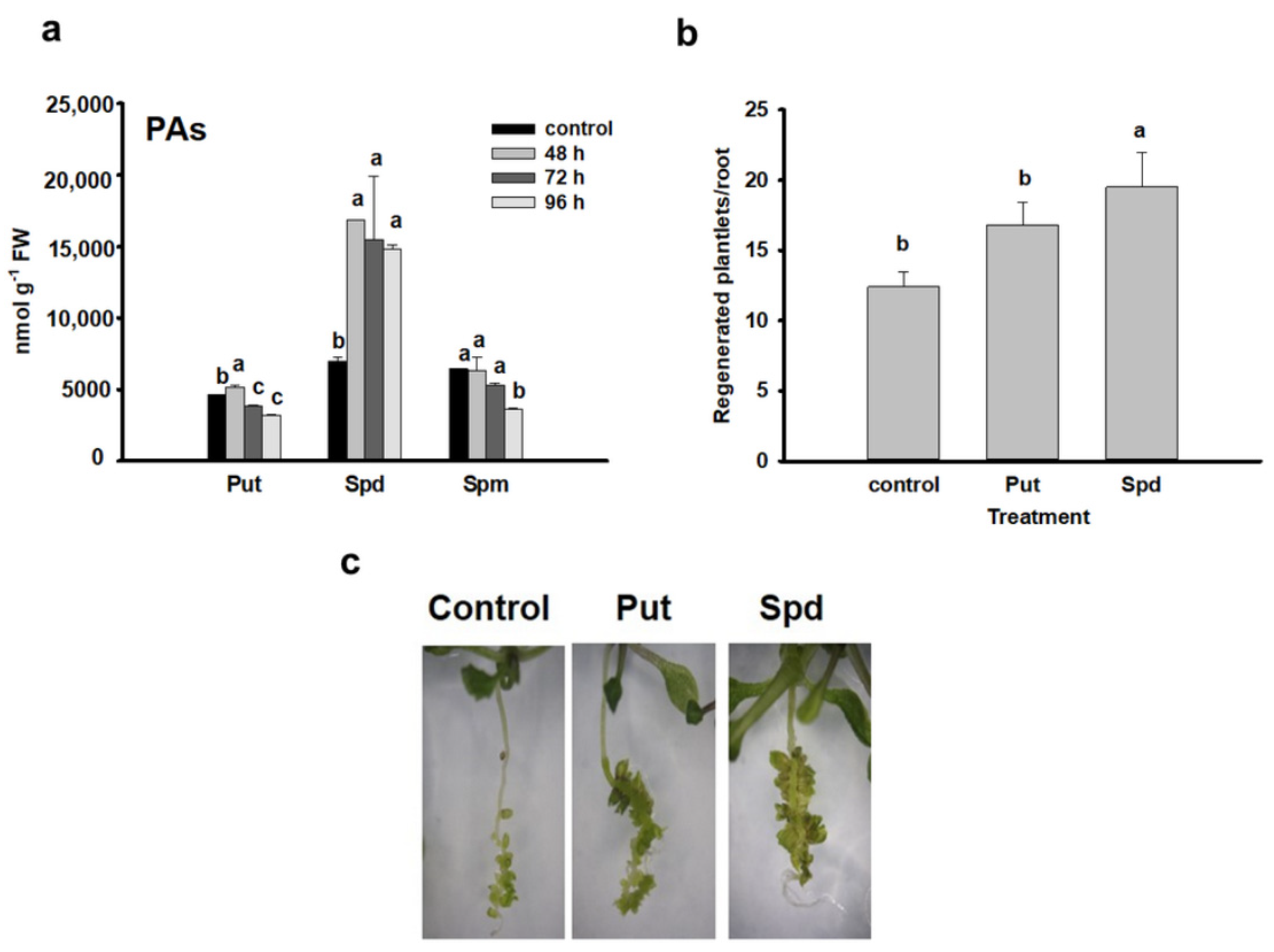{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials and Culture Conditions
2.2. Gene Expression Analysis
2.3. Light Microscopy
2.4. In Situ Detection of Reactive Oxygen Species
2.5. Free Polyamine Determination
2.6. Statistical Analysis and Data Representation
3. Results and Discussion
3.1. PA Accumulation Correlates with and Promotes the Formation of Shoot Meristems from Lateral Root Primordia
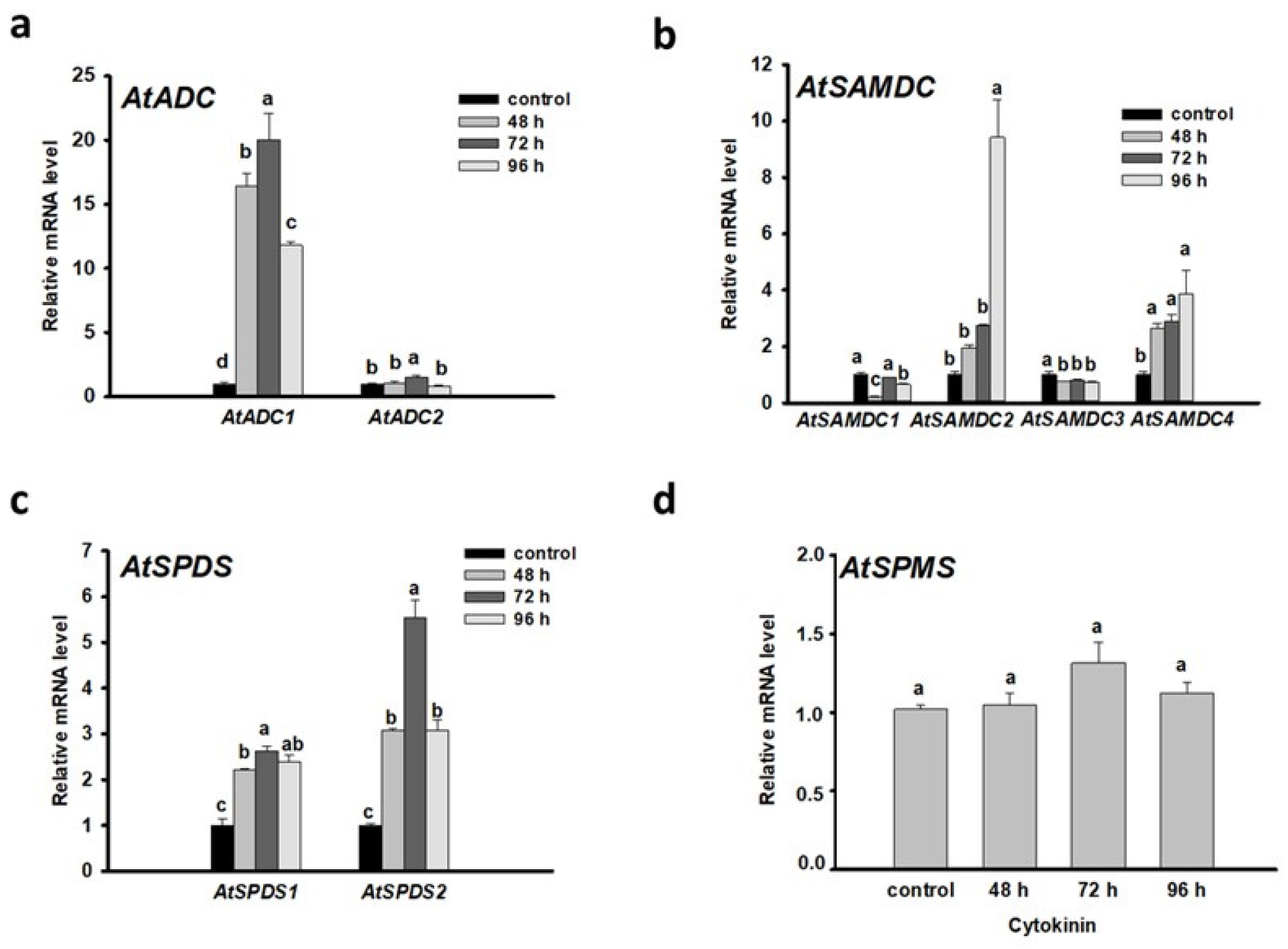
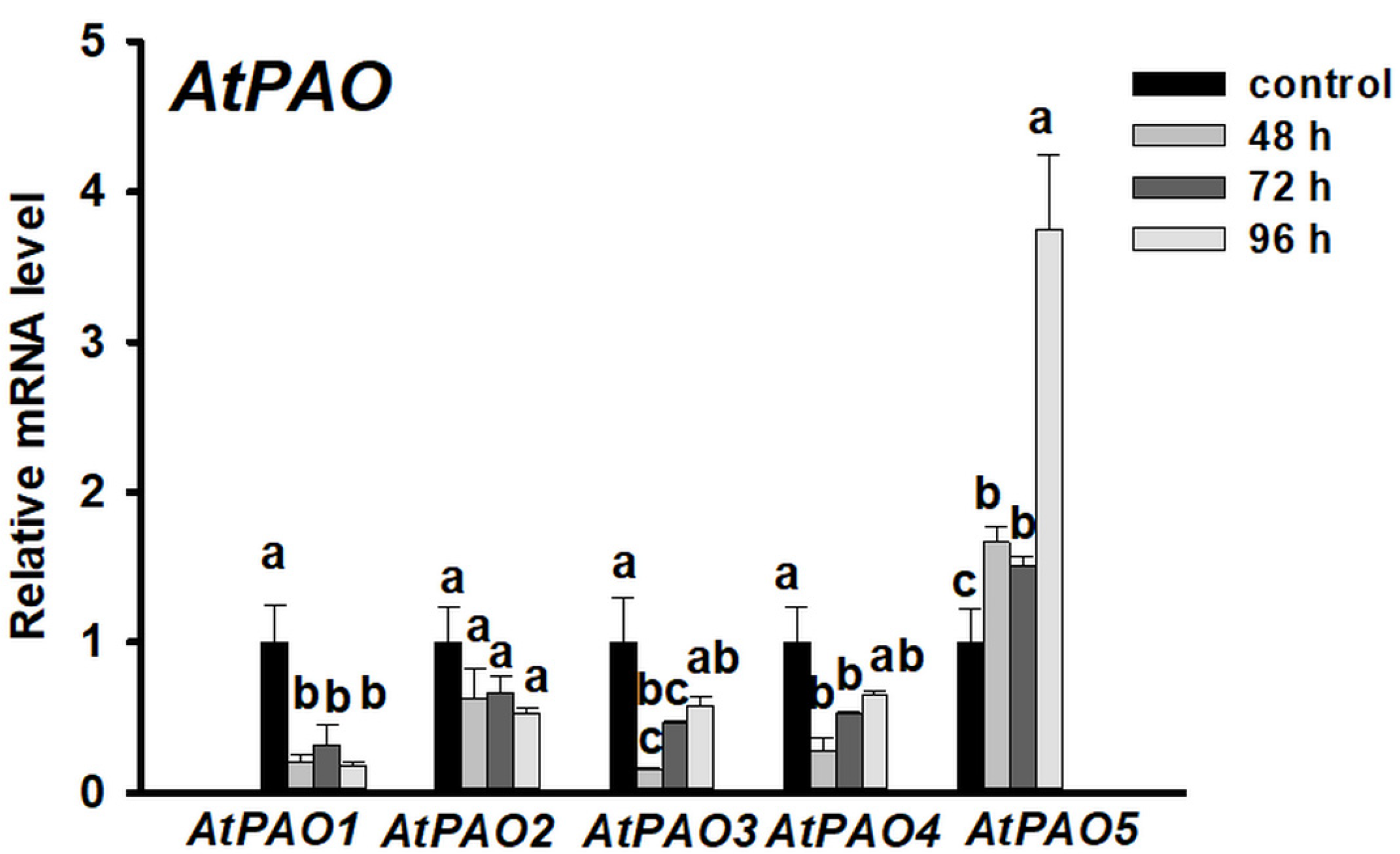
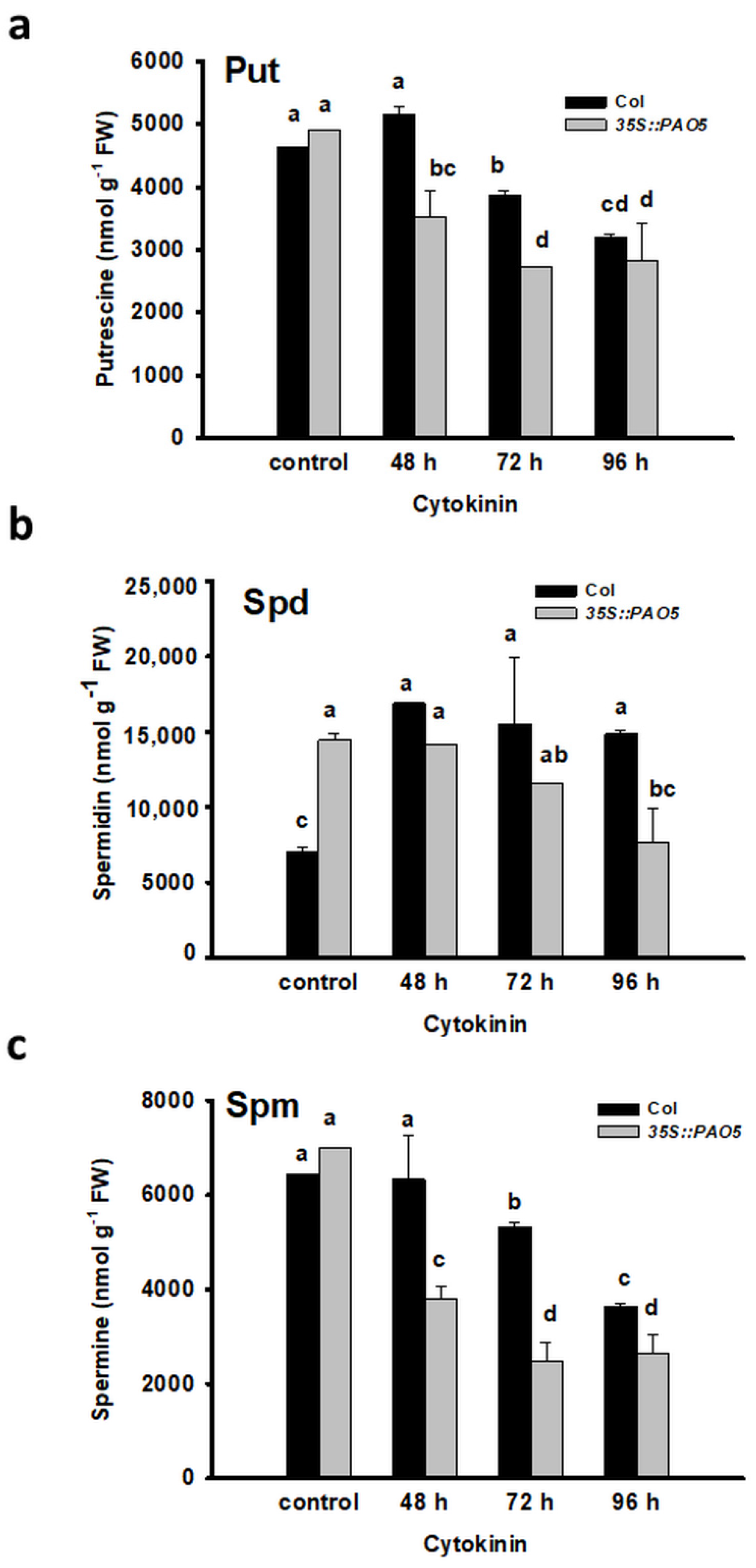
3.2. Put and Spd Synthesis Is Augmented during the Meristem Conversion Process
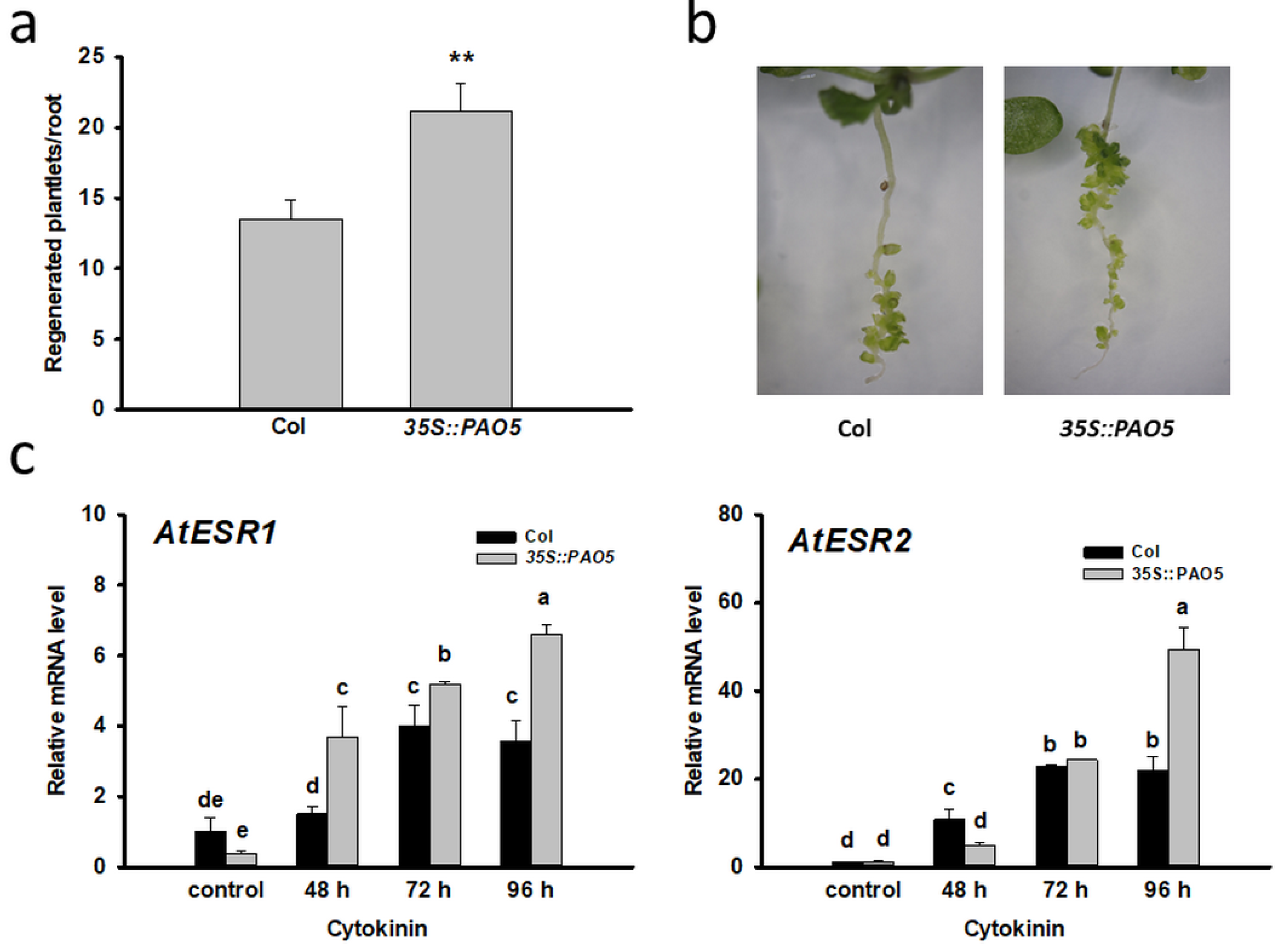
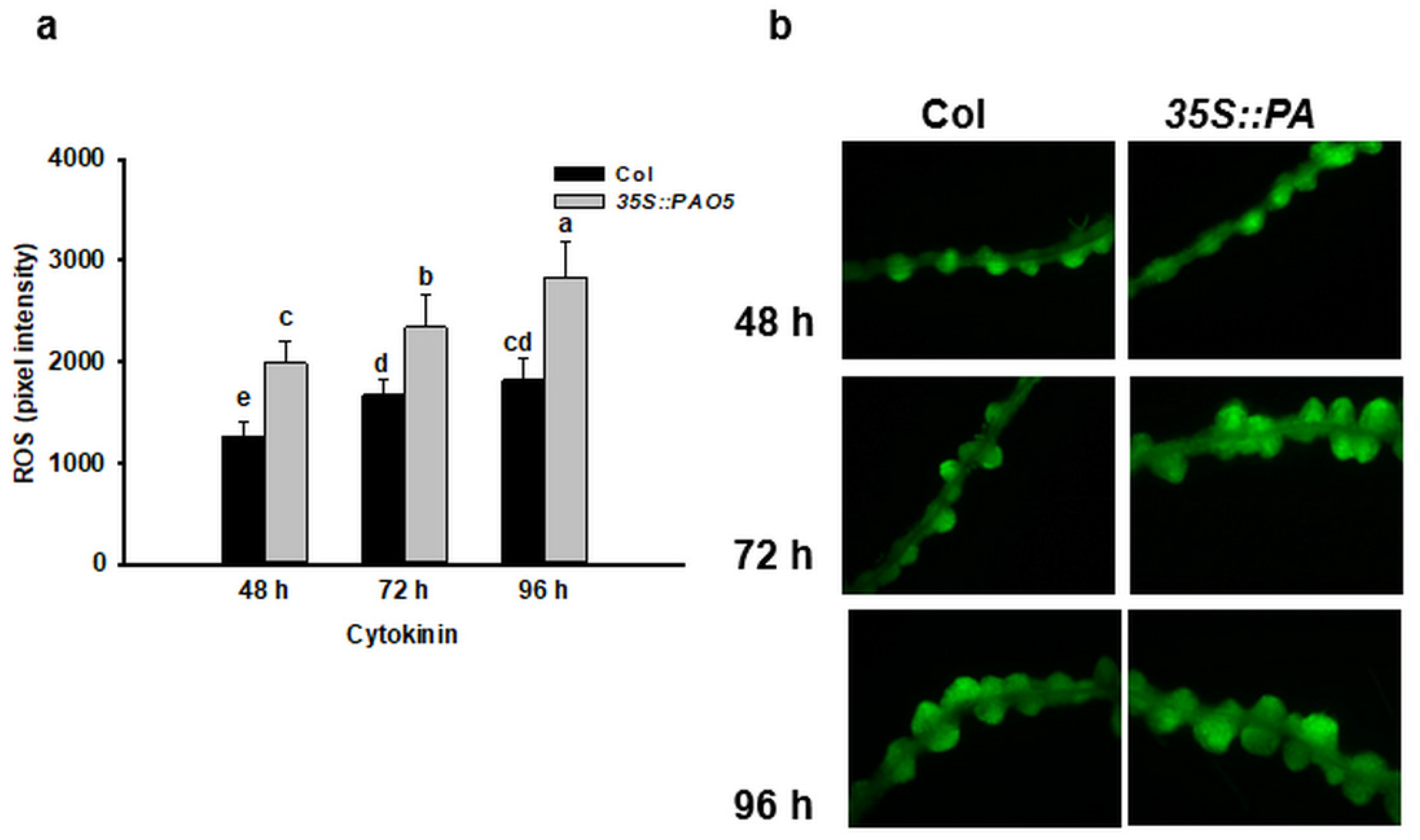
3.3. Ectopically Expressed AtPAO5 Enhances the Direct Conversion of LRPs to SMs
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ikeuchi, M.; Ogawa, Y.; Iwase, A.; Sugimoto, K. Plant Regeneration: Cellular Origins and Molecular Mechanisms. Development 2016, 143, 1442–1451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atta, R.; Laurens, L.; Boucheron-Dubuisson, E.; Guivarc’h, A.; Carnero, E.; Giraudat-Pautot, V.; Rech, P.; Chriqui, D. Pluripotency of Arabidopsis Xylem Pericycle Underlies Shoot Regeneration from Root and Hypocotyl Explants Grown in Vitro. Plant J. 2009, 57, 626–644. [Google Scholar] [CrossRef] [PubMed]
- Bernula, D.; Benkő, P.; Kaszler, N.; Domonkos, I.; Valkai, I.; Szőllősi, R.; Ferenc, G.; Ayaydin, F.; Fehér, A.; Gémes, K. Timely Removal of Exogenous Cytokinin and the Prevention of Auxin Transport from the Shoot to the Root Affect the Regeneration Potential of Arabidopsis Roots. Plant Cell Tissue Organ Cult. 2020, 140, 327–339. [Google Scholar] [CrossRef] [Green Version]
- Che, P.; Lall, S.; Howell, S.H. Developmental Steps in Acquiring Competence for Shoot Development in Arabidopsis Tissue Culture. Planta 2007, 226, 1183–1194. [Google Scholar] [CrossRef]
- Chatfield, S.P.; Capron, R.; Severino, A.; Penttila, P.-A.; Alfred, S.; Nahal, H.; Provart, N.J. Incipient Stem Cell Niche Conversion in Tissue Culture: Using a Systems Approach to Probe Early Events in WUSCHEL-Dependent Conversion of Lateral Root Primordia into Shoot Meristems. Plant J. 2013, 73, 798–813. [Google Scholar] [CrossRef]
- Kareem, A.; Radhakrishnan, D.; Wang, X.; Bagavathiappan, S.; Trivedi, Z.B.; Sugimoto, K.; Xu, J.; Mähönen, A.P.; Prasad, K. Protocol: A Method to Study the Direct Reprogramming of Lateral Root Primordia to Fertile Shoots. Plant Methods 2016, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosspopoff, O.; Chelysheva, L.; Saffar, J.; Lecorgne, L.; Gey, D.; Caillieux, E.; Colot, V.; Roudier, F.; Hilson, P.; Berthomé, R.; et al. Direct Conversion of Root Primordium into Shoot Meristem Relies on Timing of Stem Cell Niche Development. Development 2017, 144, 1187–1200. [Google Scholar] [CrossRef] [Green Version]
- Alabdallah, O.; Ahou, A.; Mancuso, N.; Pompili, V.; Macone, A.; Pashkoulov, D.; Stano, P.; Cona, A.; Angelini, R.; Tavladoraki, P. The Arabidopsis Polyamine Oxidase/Dehydrogenase 5 Interferes with Cytokinin and Auxin Signaling Pathways to Control Xylem Differentiation. J. Exp. Bot. 2017, 68, 997–1012. [Google Scholar] [CrossRef]
- Kim, D.W.; Watanabe, K.; Murayama, C.; Izawa, S.; Niitsu, M.; Michael, A.J.; Berberich, T.; Kusano, T. Polyamine Oxidase5 Regulates Arabidopsis Growth through Thermospermine Oxidase Activity. Plant Physiol. 2014, 165, 1575–1590. [Google Scholar] [CrossRef] [Green Version]
- Thiruvengadam, M.; Ill-Min, C.; Se-Chul, C. Influence of Polyamines on in Vitro Organogenesis in Bitter Melon (Momordica Charantia, L.). J. Med. Plants Res. 2012, 6, 3579–3585. [Google Scholar] [CrossRef]
- Smith, T.A. Polyamines. Annu. Rev. Plant Physiol. 1985, 36, 117–143. [Google Scholar] [CrossRef]
- Chen, D.; Shao, Q.; Yin, L.; Younis, A.; Zheng, B. Polyamine Function in Plants: Metabolism, Regulation on Development, and Roles in Abiotic Stress Responses. Front. Plant Sci. 2019, 9. [Google Scholar] [CrossRef] [PubMed]
- Roustan, J.-P.; Chraibi, K.M.; Latché, A.; Fallot, J. Relationship between Ethylene and Polyamine Synthesis in Plant Regeneration. In Cellular and Molecular Aspects of the Plant Hormone Ethylene, Proceedings of the International Symposium on Cellular and Molecular Aspects of Biosynthesis and Action of the Plant Hormone Ethylene, Agen, France, 31 August–4 September 1992; Pech, J.C., Latché, A., Balagué, C., Eds.; Current Plant Science and Biotechnology in Agriculture; Springer: Dordrecht, The Netherlands, 1993; pp. 365–366. [Google Scholar]
- Tiburcio, A.F.; Alcázar, R. Potential Applications of Polyamines in Agriculture and Plant Biotechnology. In Polyamines: Methods and Protocols; Alcázar, R., Tiburcio, A.F., Eds.; Methods in Molecular Biology; Springer: New York, NY, USA, 2018; pp. 489–508. [Google Scholar]
- Kakkar, R.K.; Nagar, P.K.; Ahuja, P.S.; Rai, V.K. Polyamines and Plant Morphogenesis. Biol. Plant. 2000, 43, 1–11. [Google Scholar] [CrossRef]
- Pua, E.-C. Regulation of Plant Morphogenesis In Vitro: Role of Ethylene and Polyamines. In Biotechnology and Sustainable Agriculture 2006 and Beyond; Xu, Z., Li, J., Xue, Y., Yang, W., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 89–95. [Google Scholar]
- Urano, K.; Hobo, T.; Shinozaki, K. Arabidopsis ADC Genes Involved in Polyamine Biosynthesis Are Essential for Seed Development. FEBS Lett. 2005, 579, 1557–1564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ge, C.; Cui, X.; Wang, Y.; Hu, Y.; Fu, Z.; Zhang, D.; Cheng, Z.; Li, J. BUD2, Encoding an S-Adenosylmethionine Decarboxylase, Is Required for Arabidopsis Growth and Development. Cell Res. 2006, 16, 446–456. [Google Scholar] [CrossRef] [PubMed]
- Cui, X.; Ge, C.; Wang, R.; Wang, H.; Chen, W.; Fu, Z.; Jiang, X.; Li, J.; Wang, Y. The BUD2 Mutation Affects Plant Architecture through Altering Cytokinin and Auxin Responses in Arabidopsis. Cell Res. 2010, 20, 576–586. [Google Scholar] [CrossRef] [Green Version]
- Kakehi, J.; Kuwashiro, Y.; Niitsu, M.; Takahashi, T. Thermospermine Is Required for Stem Elongation in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1342–1349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miyamoto, M.; Shimao, S.; Tong, W.; Motose, H.; Takahashi, T. Effect of Thermospermine on the Growth and Expression of Polyamine-Related Genes in Rice Seedlings. Plants 2019, 8, 269. [Google Scholar] [CrossRef] [Green Version]
- Takano, A.; Kakehi, J.-I.; Takahashi, T. Thermospermine Is Not a Minor Polyamine in the Plant Kingdom. Plant Cell Physiol. 2012, 53, 606–616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alcázar Hernández, R.; Bitrian, M.; Zarza, X.; Fernández Tiburcio, A. Polyamine metabolism and signaling in plant abiotic stress protection. In Recent Advances in Pharmaceutical Sciences; Transworld Research Network: Kerala, India, 2012; Volume II, pp. 29–47. [Google Scholar]
- Clay, N.K.; Nelson, T. Arabidopsis Thickvein Mutation Affects Vein Thickness and Organ Vascularization, and Resides in a Provascular Cell-Specific Spermine Synthase Involved in Vein Definition and in Polar Auxin Transport. Plant Physiol. 2005, 138, 767–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshimoto, K.; Takamura, H.; Kadota, I.; Motose, H.; Takahashi, T. Chemical Control of Xylem Differentiation by Thermospermine, Xylemin, and Auxin. Sci. Rep. 2016, 6, 21487. [Google Scholar] [CrossRef] [Green Version]
- Wang, W.; Paschalidis, K.; Feng, J.-C.; Song, J.; Liu, J.-H. Polyamine Catabolism in Plants: A Universal Process With Diverse Functions. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.-H.; Wang, F.-L.; Cheng, X.-Q.; Zhu, Q.-H.; Sun, Y.-Q.; Zhu, H.-G.; Sun, J. Polyamine and Its Metabolite H2O2 Play a Key Role in the Conversion of Embryogenic Callus into Somatic Embryos in Upland Cotton (Gossypium Hirsutum, L.). Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [Green Version]
- Angelini, R.; Cona, A.; Federico, R.; Fincato, P.; Tavladoraki, P.; Tisi, A. Plant Amine Oxidases “on the Move”: An Update. Plant Physiol. Biochem. 2010, 48, 560–564. [Google Scholar] [CrossRef] [PubMed]
- Pottosin, I.; Velarde-Buendía, A.M.; Bose, J.; Zepeda-Jazo, I.; Shabala, S.; Dobrovinskaya, O. Cross-Talk between Reactive Oxygen Species and Polyamines in Regulation of Ion Transport across the Plasma Membrane: Implications for Plant Adaptive Responses. J. Exp. Bot. 2014, 65, 1271–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fincato, P.; Moschou, P.N.; Spedaletti, V.; Tavazza, R.; Angelini, R.; Federico, R.; Roubelakis-Angelakis, K.A.; Tavladoraki, P. Functional Diversity inside the Arabidopsis Polyamine Oxidase Gene Family. J. Exp. Bot. 2011, 62, 1155–1168. [Google Scholar] [CrossRef]
- Ahou, A.; Martignago, D.; Alabdallah, O.; Tavazza, R.; Stano, P.; Macone, A.; Pivato, M.; Masi, A.; Rambla, J.L.; Vera-Sirera, F.; et al. A Plant Spermine Oxidase/Dehydrogenase Regulated by the Proteasome and Polyamines. J. Exp. Bot. 2014, 65, 1585–1603. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kamada-Nobusada, T.; Hayashi, M.; Fukazawa, M.; Sakakibara, H.; Nishimura, M. A Putative Peroxisomal Polyamine Oxidase, AtPAO4, Is Involved in Polyamine Catabolism in Arabidopsis thaliana. Plant Cell Physiol. 2008, 49, 1272–1282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moschou, P.N.; Delis, I.D.; Paschalidis, K.A.; Roubelakis-Angelakis, K.A. Transgenic Tobacco Plants Overexpressing Polyamine Oxidase Are Not Able to Cope with Oxidative Burst Generated by Abiotic Factors. Physiol. Plant 2008, 133, 140–156. [Google Scholar] [CrossRef]
- Ono, Y.; Kim, D.W.; Watanabe, K.; Sasaki, A.; Niitsu, M.; Berberich, T.; Kusano, T.; Takahashi, Y. Constitutively and Highly Expressed Oryza Sativa Polyamine Oxidases Localize in Peroxisomes and Catalyze Polyamine Back Conversion. Amino Acids 2012, 42, 867–876. [Google Scholar] [CrossRef]
- Tavladoraki, P.; Cona, A.; Angelini, R. Copper-Containing Amine Oxidases and FAD-Dependent Polyamine Oxidases Are Key Players in Plant Tissue Differentiation and Organ Development. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for General Users and for Biologist Programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Winter, D.; Vinegar, B.; Nahal, H.; Ammar, R.; Wilson, G.V.; Provart, N.J. An “Electronic Fluorescent Pictograph” Browser for Exploring and Analyzing Large-Scale Biological Data Sets. PLoS ONE 2007, 2, e718. [Google Scholar] [CrossRef] [PubMed]
- Benkő, P.; Jee, S.; Kaszler, N.; Fehér, A.; Gémes, K. Polyamines Treatment during Pollen Germination and Pollen Tube Elongation in Tobacco Modulate Reactive Oxygen Species and Nitric Oxide Homeostasis. J. Plant Physiol. 2020, 244, 153085. [Google Scholar] [CrossRef]
- Gémes, K.; Poór, P.; Horváth, E.; Kolbert, Z.; Szopkó, D.; Szepesi, Á.; Tari, I. Cross-Talk between Salicylic Acid and NaCl-Generated Reactive Oxygen Species and Nitric Oxide in Tomato during Acclimation to High Salinity. Physiol. Plant. 2011, 142, 179–192. [Google Scholar] [CrossRef]
- Takács, Z.; Poór, P.; Tari, I. Comparison of Polyamine Metabolism in Tomato Plants Exposed to Different Concentrations of Salicylic Acid under Light or Dark Conditions. Plant Physiol. Biochem. 2016, 108, 266–278. [Google Scholar] [CrossRef]
- Minocha, S.C.; Minocha, R. Role of Polyamines in Somatic Embryogenesis. In Somatic Embryogenesis and Synthetic Seed I; Biotechnology in Agriculture and Forestry; Bajaj, Y.P.S., Ed.; Springer: Berlin/Heidelberg, Germany, 1995; pp. 53–70. [Google Scholar]
- Pua, E.-C. Morphogenesis in Cell and Tissue Cultures. In Morphogenesis in Plant Tissue Cultures; Soh, W.-Y., Bhojwani, S.S., Eds.; Springer: Dordrecht, The Netherlands, 1999; pp. 255–303. [Google Scholar]
- Pua, E.-C.; Teo, S.-H.; Loh, C.-S. Interactive Role of Ethylene and Polyamines on Shoot Regenerabillty of Chinese Kale (Brassica Alboglabra Bailey) in vitro. J. Plant Physiol. 1996, 149, 138–148. [Google Scholar] [CrossRef]
- Hu, W.-W.; Gong, H.; Pua, E.-C. Modulation of SAMDC Expression in Arabidopsis thaliana Alters in Vitro Shoot Organogenesis. Physiol. Plant. 2006, 128, 740–750. [Google Scholar] [CrossRef]
- Lim, T.S.; Chitra, T.R.; Han, P.; Pua, E.-C.; Yu, H. Cloning and Characterization of Arabidopsis and Brassica Juncea Flavin-Containing Amine Oxidases. J. Exp. Bot. 2006, 57, 4155–4169. [Google Scholar] [CrossRef] [Green Version]
- Rossi, F.R.; Marina, M.; Pieckenstain, F.L. Role of Arginine Decarboxylase (ADC) in Arabidopsis thaliana Defence against the Pathogenic Bacterium Pseudomonas Viridiflava. Plant Biol. 2015, 17, 831–839. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, T.; Tong, W. Regulation and Diversity of Polyamine Biosynthesis in Plants. In Polyamines: A Universal Molecular Nexus for Growth, Survival, and Specialized Metabolism; Kusano, T., Suzuki, H., Eds.; Springer: Tokyo, Japan, 2015; pp. 27–44. [Google Scholar]
- Urano, K.; Yoshiba, Y.; Nanjo, T.; Ito, T.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Arabidopsis Stress-Inducible Gene for Arginine Decarboxylase AtADC2 Is Required for Accumulation of Putrescine in Salt Tolerance. Biochem. Biophys. Res. Commun. 2004, 313, 369–375. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Rangel, D.; Chávez-Martínez, A.I.; Rodríguez-Hernández, A.A.; Maruri-López, I.; Urano, K.; Shinozaki, K.; Jiménez-Bremont, J.F. Simultaneous Silencing of Two Arginine Decarboxylase Genes Alters Development in Arabidopsis. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Majumdar, R.; Shao, L.; Turlapati, S.A.; Minocha, S.C. Polyamines in the Life of Arabidopsis: Profiling the Expression of S-Adenosylmethionine Decarboxylase (SAMDC) Gene Family during Its Life Cycle. BMC Plant Biol. 2017, 17, 264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, X.-L.; Li, X.-J.; Li, Y.; Huang, L.-Z. The Effect of AOA on Ethylene and Polyamine Metabolism during Early Phases of Somatic Embryogenesis in Medicago Sativa. Physiol. Plant. 2001, 113, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Imai, A.; Matsuyama, T.; Hanzawa, Y.; Akiyama, T.; Tamaoki, M.; Saji, H.; Shirano, Y.; Kato, T.; Hayashi, H.; Shibata, D.; et al. Spermidine Synthase Genes Are Essential for Survival of Arabidopsis. Plant Physiol. 2004, 135, 1565–1573. [Google Scholar] [CrossRef] [Green Version]
- Banno, H.; Ikeda, Y.; Niu, Q.-W.; Chua, N.-H. Overexpression of Arabidopsis ESR1 Induces Initiation of Shoot Regeneration. Plant Cell 2001, 13, 2609–2618. [Google Scholar] [CrossRef]
- Matsuo, N.; Makino, M.; Banno, H. Arabidopsis ENHANCER OF SHOOT REGENERATION (ESR)1 and ESR2 Regulate in Vitro Shoot Regeneration and Their Expressions Are Differentially Regulated. Plant Sci. 2011, 181, 39–46. [Google Scholar] [CrossRef]
- Wimalasekera, R.; Schaarschmidt, F.; Angelini, R.; Cona, A.; Tavladoraki, P.; Scherer, G.F.E. POLYAMINE OXIDASE2 of Arabidopsis Contributes to ABA Mediated Plant Developmental Processes. Plant Physiol. Biochem. 2015, 96, 231–240. [Google Scholar] [CrossRef]
- Ferdousy, N.; Arif, T.; Sagor, G. Polyamine Oxidase 5 (PAO5) Mediated Antioxidant Response to Promote Salt Tolerance in Arabidopsis thaliana. J. Bangladesh Agric. Univ. 2020, 1. [Google Scholar] [CrossRef]
- Su, G.-X.; Zhang, W.-H.; Liu, Y.-L. Involvement of Hydrogen Peroxide Generated by Polyamine Oxidative Degradation in the Development of Lateral Roots in Soybean. J. Integr. Plant Biol. 2006, 48, 426–432. [Google Scholar] [CrossRef]
- Considine, M.J.; Foyer, C.H. Redox Regulation of Plant Development. Antioxid Redox Signal 2013, 21, 1305–1326. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schmidt, R.; Schippers, J.H.M. ROS-Mediated Redox Signaling during Cell Differentiation in Plants. Biochim. Biophys. Acta (BBA) Gen. Subj. 2015, 1850, 1497–1508. [Google Scholar] [CrossRef] [PubMed]
- Zeng, J.; Dong, Z.; Wu, H.; Tian, Z.; Zhao, Z. Redox Regulation of Plant Stem Cell Fate. EMBO J. 2017, 36, 2844–2855. [Google Scholar] [CrossRef]
- Shin, J.; Bae, S.; Seo, P.J. De Novo Shoot Organogenesis during Plant Regeneration. J. Exp. Bot. 2020, 71, 63–72. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, T.T.; Liu, H.; Shi, D.Y.; Wang, M.; Bie, X.M.; Li, X.G.; Zhang, X.S. Thioredoxin-Mediated ROS Homeostasis Explains Natural Variation in Plant Regeneration. Plant Physiol. 2018, 176, 2231–2250. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaszler, N.; Benkő, P.; Bernula, D.; Szepesi, Á.; Fehér, A.; Gémes, K. Polyamine Metabolism Is Involved in the Direct Regeneration of Shoots from Arabidopsis Lateral Root Primordia. Plants 2021, 10, 305. https://doi.org/10.3390/plants10020305
Kaszler N, Benkő P, Bernula D, Szepesi Á, Fehér A, Gémes K. Polyamine Metabolism Is Involved in the Direct Regeneration of Shoots from Arabidopsis Lateral Root Primordia. Plants. 2021; 10(2):305. https://doi.org/10.3390/plants10020305
Chicago/Turabian StyleKaszler, Nikolett, Péter Benkő, Dóra Bernula, Ágnes Szepesi, Attila Fehér, and Katalin Gémes. 2021. "Polyamine Metabolism Is Involved in the Direct Regeneration of Shoots from Arabidopsis Lateral Root Primordia" Plants 10, no. 2: 305. https://doi.org/10.3390/plants10020305