
Consideration of Commercially Available Hepatocytes as Cell Sources for Liver-Microphysiological Systems by Comparing Liver Characteristics
Abstract
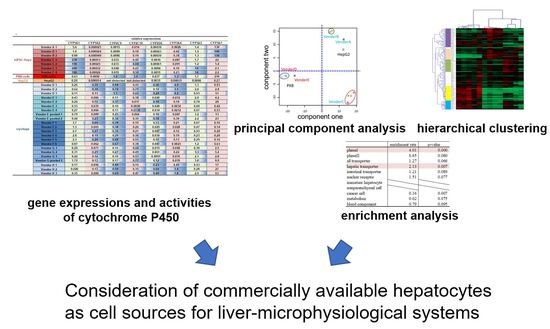
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Cell Culture
2.2. RNA Isolation
2.3. Measurement of Gene Expression by a TaqMan Real-Time PCR
2.4. Measurement of Metabolic Activity of CYP Enzymes with LC-MS/MS
2.5. Gene Expression Analysis Using the Focused Microarray Assay
2.6. Data Analysis
3. Results
3.1. Comparing the CYP Gene Expression and Activities of Cell Source Candidates for Liver-MPS and CryoHeps
3.2. Performance Evaluation of the Focused Microaasay
3.3. Characteristic Comparison between Cells Based on the Expression Data from the Focused Microarray Assay
3.4. Gene Classification by Expression Pattern and Biological Interpretation by Enrichment Analysis
3.5. Gene Expression Analysis of Intestinal Markers
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Watkins, P.B. Drug safety sciences and the bottleneck in drug development. Clin. Pharmacol. Ther. 2011, 89, 788–790. [Google Scholar] [CrossRef] [PubMed]
- Günther, K.; Sonntag, F.; Moritzer, E.; Hirsch, A.; Klotzbach, U.; Lasagni, A.F. Universal Micromachining Platform and Basic Technologies for the Manufacture and Marking of Microphysiological Systems. Micromachines 2017, 8, 246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sances, S.; Ho, R.; Vatine, G.; West, D.; Laperle, A.; Meyer, A.; Godoy, M.; Kay, P.S.; Mandefro, B.; Hatata, S.; et al. Human iPSC-Derived Endothelial Cells and Microengineered Organ-Chip Enhance Neuronal Development. Stem Cell Rep. 2018, 10, 1222–1236. [Google Scholar] [CrossRef] [Green Version]
- Jang, K.J.; Otieno, M.A.; Ronxhi, J.; Lim, H.K.; Ewart, L.; Kodella, K.R.; Petropolis, D.; Kulkarni, G.; Rubins, J.E.; Conegliano, D.; et al. Reproducing human and cross-species drug toxicities using a Liver-Chip. Sci. Transl. Med. 2019, 11, eaax5516. [Google Scholar] [CrossRef] [PubMed]
- Kasendra, M.; Luc, R.; Yin, J.; Manatakis, D.V.; Kulkarni, G.; Lucchesi, C.; Sliz, J.; Apostolou, A.; Sunuwar, L.; Obrigewitch, J.; et al. Duodenum Intestine-Chip for preclinical drug assessment in a human relevant model. Elife 2020, 9, e50135. [Google Scholar] [CrossRef] [PubMed]
- Nawroth, J.C.; Lucchesi, C.; Cheng, D.; Shukla, A.; Ngyuen, J.; Shroff, T.; Varone, A.; Karalis, K.; Lee, H.H.; Alves, S.; et al. Microengineered Airway Lung Chip Models Key Features of Viral-induced Exacerbation of Asthma. Am. J. Respir. Cell Mol. Biol. 2020, 63, 591–600. [Google Scholar] [CrossRef] [PubMed]
- Tao, T.P.; Brandmair, K.; Gerlach, S.; Przibilla, J.; Géniès, C.; Jacques-Jamin, C.; Schepky, A.; Marx, U.; Hewitt, N.J.; Maschmeyer, I.; et al. Demonstration of the first-pass metabolism in the skin of the hair dye, 4-amino-2-hydroxytoluene, using the Chip2 skin-liver microphysiological model. J. Appl. Toxicol. 2021, 41, 1553–1567. [Google Scholar] [CrossRef]
- Shabestani Monfared, G.; Ertl, P.; Rothbauer, M. Microfluidic and Lab-on-a-Chip Systems for Cutaneous Wound Healing Studies. Pharmaceutics 2021, 13, 793. [Google Scholar] [CrossRef]
- Ishida, S. Research and Development of Microphysiological Systems in Japan Supported by the AMED-MPS Project. Front. Toxicol. 2021, 3, 657765. [Google Scholar] [CrossRef]
- Zhou, Z.; Zhu, J.; Jiang, M.; Sang, L.; Hao, K.; He, H. The Combination of Cell Cultured Technology and In Silico Model to Inform the Drug Development. Pharmaceutics 2021, 13, 704. [Google Scholar] [CrossRef]
- Kim, Y.S.; Asif, A.; Chethikkattuveli Salih, A.R.; Lee, J.W.; Hyun, K.N.; Choi, K.H. Gravity-Based Flow Efficient Perfusion Culture System for Spheroids Mimicking Liver Inflammation. Biomedicines 2021, 9, 1369. [Google Scholar] [CrossRef] [PubMed]
- Shinohara, M.; Arakawa, H.; Oda, Y.; Shiraki, N.; Sugiura, S.; Nishiuchi, T.; Satoh, T.; Iino, K.; Leo, S.; Kato, Y.; et al. Coculture with hiPS-derived intestinal cells enhanced human hepatocyte functions in a pneumatic-pressure-driven two-organ microphysiological system. Sci. Rep. 2021, 11, 5437. [Google Scholar] [CrossRef] [PubMed]
- Rubiano, A.; Indapurkar, A.; Yokosawa, R.; Miedzik, A.; Rosenzweig, B.; Arefin, A.; Moulin, C.M.; Dame, K.; Hartman, N.; Volpe, D.A.; et al. Characterizing the reproducibility in using a liver microphysiological system for assaying drug toxicity, metabolism, and accumulation. Lin. Transl. Sci. 2021, 14, 1049–1061. [Google Scholar] [CrossRef] [PubMed]
- Koenig, L.; Ramme, A.P.; Faust, D.; Mayer, M.; Flötke, T.; Gerhartl, A.; Brachner, A.; Neuhaus, W.; Appelt-Menzel, A.; Metzger, M.; et al. A Human Stem Cell-Derived Brain-Liver Chip for Assessing Blood-Brain-Barrier Permeation of Pharmaceutical Drugs. Cells 2022, 11, 3295. [Google Scholar] [CrossRef]
- Specioso, G.; Bovard, D.; Zanetti, F.; Maranzano, F.; Merg, C.; Sandoz, A.; Titz, B.; Dalcanale, F.; Hoeng, J.; Renggli, K.; et al. Apical Medium Flow Influences the Morphology and Physiology of Human Proximal Tubular Cells in a Microphysiological System. Bioengineering 2022, 9, 516. [Google Scholar] [CrossRef]
- Baudy, A.R.; Otieno, M.A.; Hewitt, P.; Gan, J.; Roth, A.; Keller, D.; Sura, R.; Van Vleet, T.R.; Proctor, W.R. Liver microphysiological systems development guidelines for safety risk assessment in the pharmaceutical industry. Lab. Chip. 2020, 20, 215–225. [Google Scholar] [CrossRef]
- Cayo, M.A.; Cai, J.; DeLaForest, A.; Noto, F.K.; Nagaoka, M.; Clark, B.S.; Collery, R.F.; Si-Tayeb, K.; Duncan, S.A. JD induced pluripotent stem cell-derived hepatocytes faithfully recapitulate the pathophysiology of familial hypercholesterolemia. Hepatology 2012, 56, 2163–2171. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.F.; Tseng, C.Y.; Wang, H.W.; Kuo, H.C.; Yang, V.W.; Lee, O.K. Rapid generation of mature hepatocyte-like cells from human induced pluripotent stem cells by an efficient three-step protocol. Hepatology 2012, 55, 1193–1203. [Google Scholar] [CrossRef] [Green Version]
- Choi, S.M.; Kim, Y.; Him, J.S.; Park, J.T.; Wang, R.H.; Leach, S.D.; Liu, J.O.; Deng, C.; Ye, Z.; Jang, Y.Y. Efficient drug screening and gene correction for treating liver disease using patient-specific stem cells. Hepatology 2013, 57, 2458–2468. [Google Scholar] [CrossRef] [Green Version]
- Fattahi, F.; Asgari, S.; Pournasr, B.; Seifinejad, A.; Totonchi, M.; Taei, A.; Aghdami, N.; Salekdeh, G.H.; Baharvand, H. Disease-corrected hepatocyte-like cells from familial hypercholesterolemia-induced pluripotent stem cells. Mol. Biotechnology 2013, 54, 863–873. [Google Scholar] [CrossRef]
- Ghodsizadeh, A.; Taei, A.; Totonchi, M.; Seifinejad, A.; Gourabi, H.; Pournasr, B.; Aghdami, N.; Malekzadeh, R.; Almadani, N.; Salekdeh, G.H.; et al. Generation of liver disease-specific induced pluripotent stem cells along with efficient differentiation to functional hepatocyte-like cells. Stem Cell Rev. 2021, 17, 1516–1517. [Google Scholar] [CrossRef] [PubMed]
- Hannan, N.R.; Segeritz, C.P.; Touboul, T.; Vallier, L. Production of hepatocyte-like cells from human pluripotent stem cells. Nat. Protoc. 2013, 8, 430–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jozefczuk, J.; Prigione, A.; Chavez, L.; Adjaye, J. Comparative analysis of human embryonic stem cell and induced pluripotent stem cell-derived hepatocyte-like cells reveals current drawbacks and possible strategies for improved differentiation. Stem Cells Dev. 2011, 20, 1259–1275. [Google Scholar] [CrossRef] [Green Version]
- Ma, X.; Duan, Y.; Tschudy-Seney, B.; Roll, G.; Behbahan, I.S.; Ahuja, T.P.; Tolstikov, V.; Wang, C.; McGee, J.; Khoobyari, S.; et al. Highly efficient differentiation of functional hepatocytes from human induced pluripotent stem cells. Stem Cells Transl. Med. 2013, 2, 409–419. [Google Scholar] [CrossRef]
- Sgodda, M.; Mobus, S.; Hoepfner, J.; Sharma, A.D.; Schambach, A.; Greber, B.; Ott, M.; Cantz, T. Improved hepatic differentiation strategies for human induced pluripotent stem cells. Curr. Mol. Med. 2013, 13, 842–855. [Google Scholar] [CrossRef] [PubMed]
- Si-Tayeb, K.; Noto, F.; KNagaoka, M.; Li, J.; Battle, M.A.; Duris, C.; North, P.E.; Dalton, S.; Duncan, S.A. Highly efficient generation of human hepatocyte-like cells from induced pluripotent stem cells. Hepatology 2010, 51, 297–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Cai, J.; Liu, Y.; Zhao, D.; Yong, J.; Duo, S.; Song, X.; Guo, Y.; Zhao, Y.; Qin, H.; et al. Efficient generation of hepatocyte-like cells from human induced pluripotent stem cells. Cell Res. 2009, 19, 1233–1242. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, G.J.; Hay, D.C.; Park, I.H.; Fletcher, J.; Hannoun, Z.; Payne, C.M.; Dalgetty, D.; Black, J.R.; Ross, J.A.; Samuel, K.; et al. Generation of functional human hepatic endoderm from human induced pluripotent stem cells. Hepatology 2010, 51, 329–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takayama, K.; Inamura, M.; Kawabata, K.; Katayama, K.; Higuchi, M.; Tashiro, K.; Nonaka, A.; Sakurai, F.; Hayakawa, T.; Furue, M.K.; et al. Efficient generation of functional hepatocytes from human embryonic stem cells and induced pluripotent stem cells by HNF4alpha transduction. Mol. Ther. 2012, 20, 127–137. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, T.; Takayama, K.; Kondoh, M.; Sakurai, F.; Tani, H.; Sakamoto, N.; Matsuura, Y.; Mizuguchi, H.; Yagi, K. Use of human hepatocyte-like cells derived from induced pluripotent stem cells as a model for hepatocytes in hepatitis C virus infection. Biochem. Biophys. Res. Commun. 2011, 416, 119–124. [Google Scholar] [CrossRef]
- Yu, Y.; Liu, H.; Ikeda, Y.; Amiot, B.P.; Rinaldo, P.; Duncan, S.A.; Nyberg, S.L. Hepatocyte-like cells differentiated from human induced pluripotent stem cells: Relevance to cellular therapies. Stem Cell Res. 2012, 9, 196–207. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umeda, K.; Suzuki, K.; Yamazoe, T.; Shiraki, N.; Higuchi, Y.; Tokieda, K.; Kume, K.; Mitani, K.; Kume, S. Albumin gene targeting in human embryonic stem cells and induced pluripotent stem cells with helper-dependent adenoviral vector to monitor hepatic differentiation. Stem Cell Res. 2013, 10, 179–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakai, Y.; Iwao, T.; Susukida, T.; Nukaga, T.; Takemura, A.; Sekine, S.; Ito, K.; Matsunaga, T. In vitro bile acid-dependent hepatocyte toxicity assay system using human induced pluripotent stem cell-derived hepatocytes: Current status and disadvantages to overcome. Drug Metab. Pharm. 2019, 34, 264–271. [Google Scholar] [CrossRef]
- Kondo, Y.; Iwao, T.; Nakamura, K.; Sasaki, T.; Takahashi, S.; Kamada, N.; Matsubara, T.; Gonzalez, F.J.; Akutsu, H.; Miyagawa, Y.; et al. An efficient method for differentiation of human induced pluripotent stem cells into hepatocyte-like cells retaining drug metabolizing activity. Drug Metab. Pharm. 2014, 29, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Katoh, M.; Matsui, T.; Nakajima, M.; Tateno, C.; Kataoka, M.; Soeno, Y.; Horie, T.; Iwasaki, K.; Yoshizato, K.; Yokoi, T. Expression of human cytochromes P450 in chimeric mice with humanized liver. Drug Metab. Dispos. 2004, 32, 1402–1410. [Google Scholar] [CrossRef]
- Yamasaki, C.; Ishida, Y.; Yanagi, A.; Yoshizane, Y.; Kojima, Y.; Ogawa, Y.; Kageyama, Y.; Iwasaki, Y.; Ishida, S.; Chayama, K.; et al. Culture density contributes to hepatic functions of fresh human hepatocytes isolated from chimeric mice with humanized livers: Novel, long-term, functional two-dimensional in vitro tool for developing new drugs. PLoS ONE 2020, 15, e0237809. [Google Scholar] [CrossRef]
- Katoh, M.; Matsui, T.; Okumura, H.; Nakajima, M.; Nishimura, M.; Naito, S.; Tateno, C.; Yoshizato, K.; Yokoi, T. Expression of human phase II enzymes in chimeric mice with humanized liver. Drug Metab. Dispos. 2005, 33, 1333–1340. [Google Scholar] [CrossRef] [Green Version]
- Okumura, H.; Katoh, M.; Sawada, T.; Nakajima, M.; Soeno, Y.; Yabuuchi, H.; Ikeda, T.; Tateno, C.; Yoshizato, K.; Yokoi, T. Humanization of excretory pathway in chimeric mice with humanized liver. Toxicol. Sci. 2007, 97, 533–538. [Google Scholar] [CrossRef]
- Tateno, C.; Yoshizane, Y.; Saito, N.; Kataoka, M.; Utoh, R.; Yamasaki, C.; Tachibana, A.; Soeno, Y.; Asahina, K.; Hino, H.; et al. Near completely humanized liver in mice shows human-type metabolic responses to drugs. Am. J. Pathol. 2004, 165, 901–912. [Google Scholar] [CrossRef] [Green Version]
- Kakuni, M.; Yamasaki, C.; Tachibana, A.; Yoshizane, Y.; Ishida, Y.; Tateno, C. Chimeric mice with humanized livers: A unique tool for in vivo and in vitro enzyme induction studies. Int. J. Mol. Sci. 2013, 15, 58–74. [Google Scholar] [CrossRef]
- Liang, H.; Zhang, L.; Wang, H.; Tang, J.; Yang, J.; Wu, C.; Chen, S. Inhibitory Effect of Gardenoside on Free Fatty Acid-Induced Steatosis in HepG2 Hepatocytes. Int. J. Mol. Sci. 2015, 16, 27749–27756. [Google Scholar] [CrossRef] [PubMed]
- Nagarajan, S.R.; Paul-Heng, M.; Krycer, J.R.; Fazakerley, D.J.; Sharland, A.F.; Hoy, A.J. Lipid and glucose metabolism in hepatocyte cell lines and primary mouse hepatocytes: A comprehensive resource for in vitro studies of hepatic metabolism. Am. J. Physiol. Endocrinol. Metab. 2019, 316, E578–E589. [Google Scholar] [CrossRef] [PubMed]
- Marroquin, L.D.; Hynes, J.; Dykens, J.A.; Jamieson, J.D.; Will, Y. Circumventing the Crabtree effect: Replacing media glucose with galactose increases susceptibility of HepG2 cells to mitochondrial toxicants. Toxicol. Sci. 2007, 97, 539–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westerink, W.M.; Schoonen, W.G. Cytochrome P450 enzyme levels in HepG2 cells and cryopreserved primary human hepatocytes and their induction in HepG2 cells. Toxicol. Vitro 2007, 21, 1581–1591. [Google Scholar] [CrossRef] [PubMed]
- Okuzaki, D.; Fukushima, T.; Tougan, T.; Ishii, T.; Kobayashi, S.; Yoshizaki, K.; Akita, T.; Nojima, H. Genopal™: A novel hollow fibre array for focused microarray analysis. DNA Res. 2010, 17, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Niwa, M.; Nagai, K.; Oike, H.; Kobori, M. Evaluation of the skin irritation using a DNA microarray on a reconstructed human epidermal model. Biol. Pharm. Bull. 2009, 32, 203–208. [Google Scholar] [CrossRef] [Green Version]
- Chong, J.; Soufan, O.; Li, C.; Caraus, I.; Li, S.; Bourque, G.; Wishart, D.S.; Xia, J. MetaboAnalyst 4.0: Towards more transparent and integrative metabolomics analysis. Nucleic Acids Res. 2018, 46, W486–W494. [Google Scholar] [CrossRef] [Green Version]
- Saeed, A.I.; Sharov, V.; White, J.; Li, J.; Liang, W.; Bhagabati, N.; Braisted, J.; Klapa, M.; Currier, T.; Thiagarajan, M.; et al. TM4: A free, open-source system for microarray data management and analysis. Biotechniques 2003, 34, 374–378. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.; Kabeya, T.; Ogawa, I.; Anno, S.; Hayashi, H.; Kanaki, T.; Hashita, T.; Iwao, T.; Matsunaga, T. Gellan Gum Promotes the Differentiation of Enterocytes from Human Induced Pluripotent Stem Cells. Pharmaceutics 2020, 12, 951. [Google Scholar] [CrossRef]
- Yamasaki, C.; Kataoka, M.; Kato, Y.; Kakuni, M.; Usuda, S.; Ohzone, Y.; Matsuda, S.; Adachi, Y.; Ninomiya, S.; Itamoto, T.; et al. In Vitro Evaluation of Cytochrome P450 and Glucuronidation Activities in Hepatocytes Isolated from Liver-Humanized Mice. Drug Metab. Pharm. 2010, 25, 539–550. [Google Scholar] [CrossRef]
- Kratochwil, N.A.; Meille, C.; Fowler, S.; Klammers, F.; Ekiciler, A.; Molitor, B.; Simon, S.; Walter, I.; McGinnis, C.; Walther, J.; et al. Metabolic Profiling of Human Long-Term Liver Models and Hepatic Clearance Predictions from In Vitro Data Using Nonlinear Mixed-Effects Modeling. AAPS J. 2017, 19, 534–550. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, T.; Takayama, K.; Hori, M.; Harada-Shiba, M.; Mizuguchi, H. Pharmaceutical Research for Inherited Metabolic Disorders of the Liver Using Human Induced Pluripotent Stem Cell and Genome Editing Technologies. Biol. Pharm. Bull. 2019, 42, 312–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imagawa, K.; Takayama, K.; Isoyama, S.; Tanikawa, K.; Shinkai, M.; Harada, K.; Tachibana, M.; Sakurai, F.; Noguchi, E.; Hirata, K.; et al. Generation of a bile salt export pump deficiency model using patient-specific induced pluripotent stem cell-derived hepatocyte-like cells. Sci. Rep. 2017, 7, 41806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horiuchi, S.; Kuroda, Y.; Oyafuso, R.; Komizu, Y.; Takaki, T.; Maeda, K.; Ishida, S. Construction of a culture protocol for functional bile canaliculi formation to apply human iPS cell-derived hepatocytes for cholestasis evaluation. Sci. Rep. 2022, 12, 15192. [Google Scholar] [CrossRef] [PubMed]
- Qiu, S.; Li, Y.; Imakura, Y.; Mima, S.; Hashita, T.; Iwao, T.; Matsunaga, T. An Efficient Method for the Differentiation of Human iPSC-Derived Endoderm toward Enterocytes and Hepatocytes. Cells 2021, 10, 812. [Google Scholar] [CrossRef]
- Lundquist, P.; Lööf, J.; Fagerholm, U.; Sjögren, I.; Johansson, J.; Briem, S.; Hoogstraate, J.; Afzelius, L.; Andersson, T.B. Prediction of in vivo rat biliary drug clearance from an in vitro hepatocyte efflux model. Drug Metab. Dispos. 2014, 42, 459–468. [Google Scholar] [CrossRef] [Green Version]
- Abe, K.; Bridges, A.S.; Brouwer, K.L.R. Use of sandwich-cultured human hepatocytes to predict biliary clearance of angiotensin II receptor blockers and HMG-CoA reductase inhibitors. Drug Metab. Dispos. 2009, 37, 447–452. [Google Scholar] [CrossRef]










Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horiuchi, S.; Kuroda, Y.; Komizu, Y.; Ishida, S. Consideration of Commercially Available Hepatocytes as Cell Sources for Liver-Microphysiological Systems by Comparing Liver Characteristics. Pharmaceutics 2023, 15, 55. https://doi.org/10.3390/pharmaceutics15010055
Horiuchi S, Kuroda Y, Komizu Y, Ishida S. Consideration of Commercially Available Hepatocytes as Cell Sources for Liver-Microphysiological Systems by Comparing Liver Characteristics. Pharmaceutics. 2023; 15(1):55. https://doi.org/10.3390/pharmaceutics15010055
Chicago/Turabian StyleHoriuchi, Shinichiro, Yukie Kuroda, Yuji Komizu, and Seiichi Ishida. 2023. "Consideration of Commercially Available Hepatocytes as Cell Sources for Liver-Microphysiological Systems by Comparing Liver Characteristics" Pharmaceutics 15, no. 1: 55. https://doi.org/10.3390/pharmaceutics15010055