Generation of a hTERT-Immortalized Human Sertoli Cell Model to Study Transporter Dynamics at the Blood-Testis Barrier


Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Cell Culture
2.3. Generation of Stable hTERT-Transduced Human Sertoli Cell Lines
2.4. RT-qPCR Analysis
2.5. Western Blotting
2.6. Transepithelial Electrical Resistance
2.7. Uridine Transport Assays
2.8. Affymetrix GeneChip Microarray Analysis
2.9. Statistical Analysis
3. Results
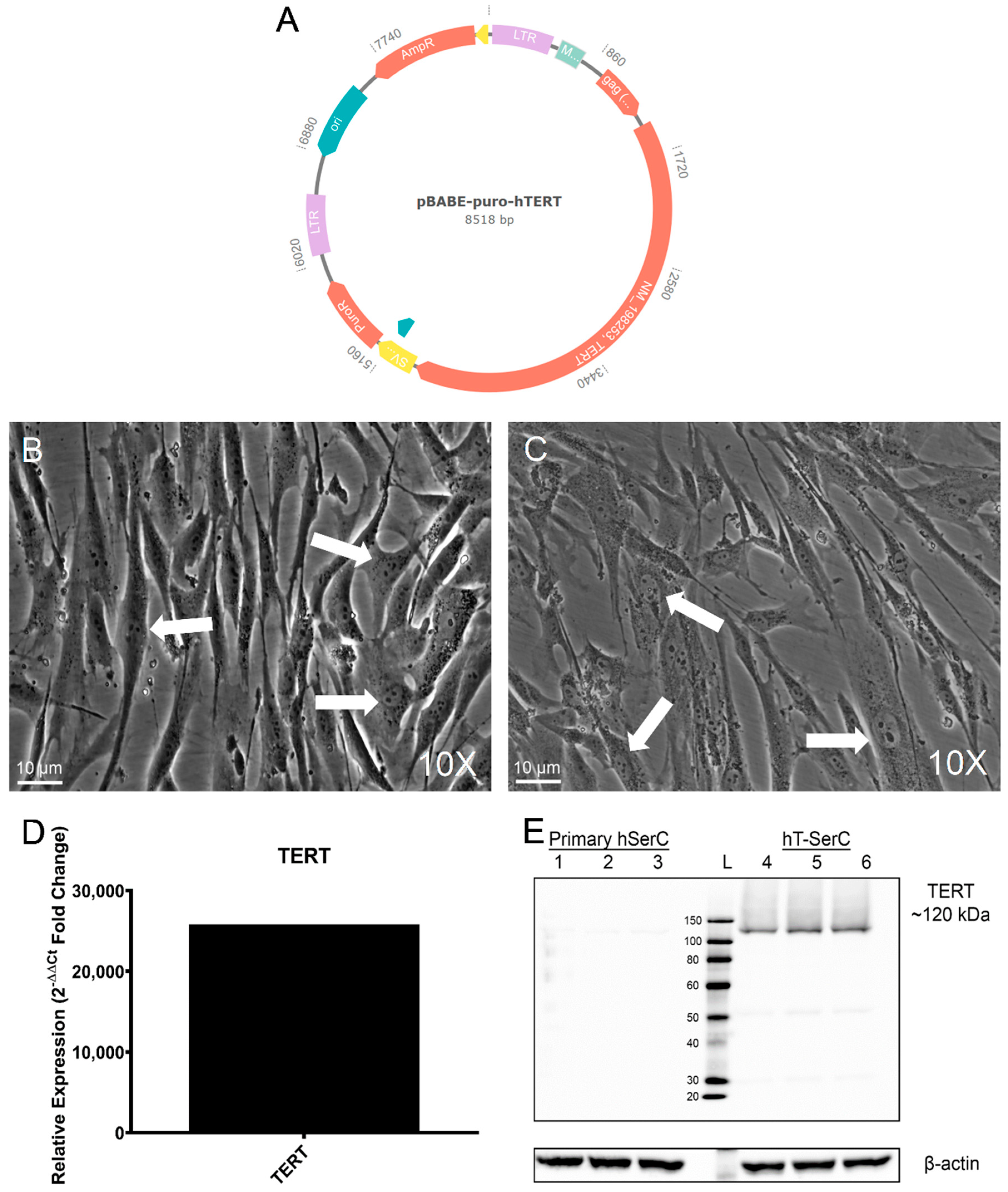
3.1. Immortalization of Human Sertoli Cells with Human Telomerase Reverse Transcriptase
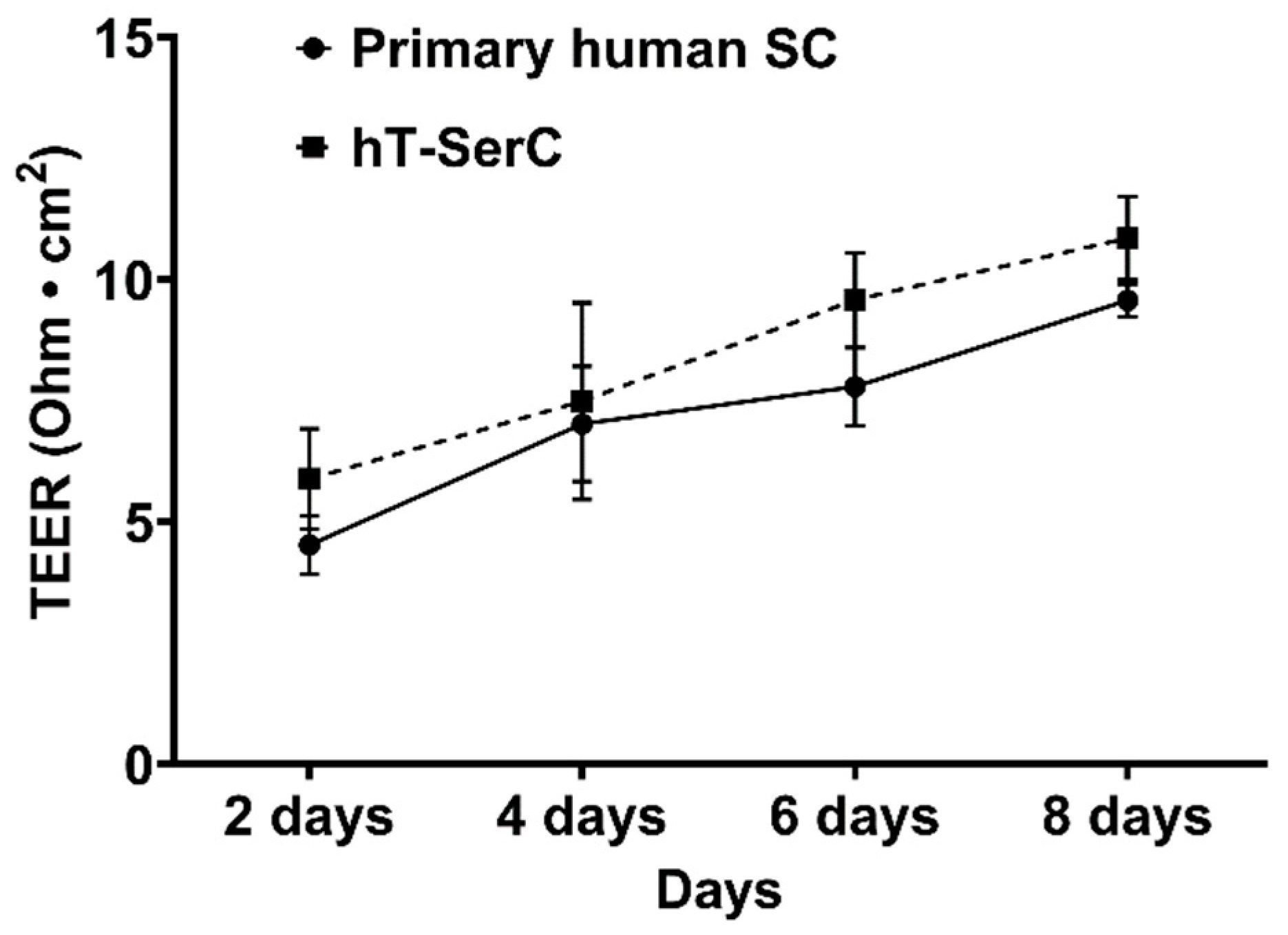
3.2. Transepithelial Electrical Resistance
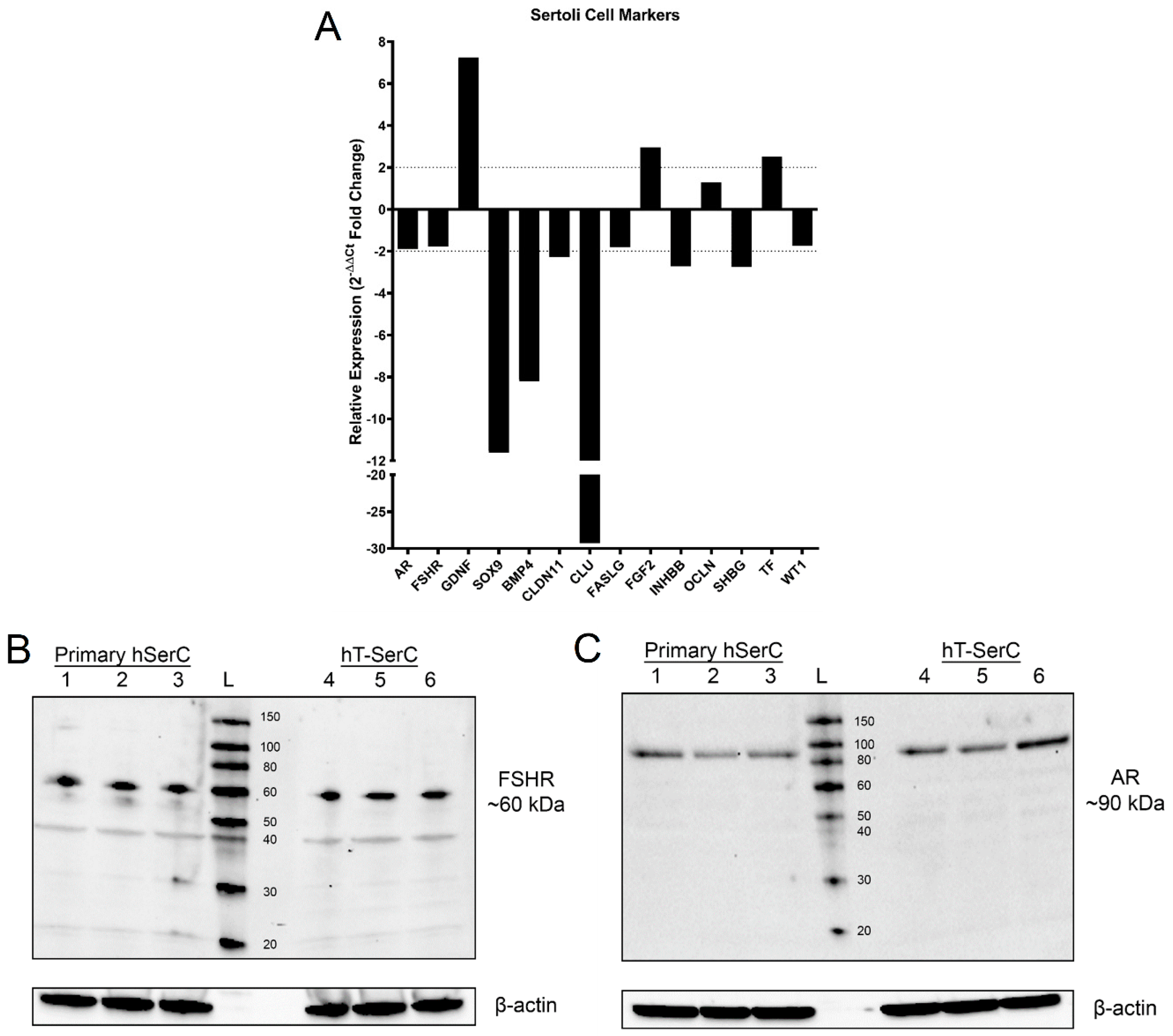
3.3. Gene Expression Profiles for Typical Sertoli Cell Markers
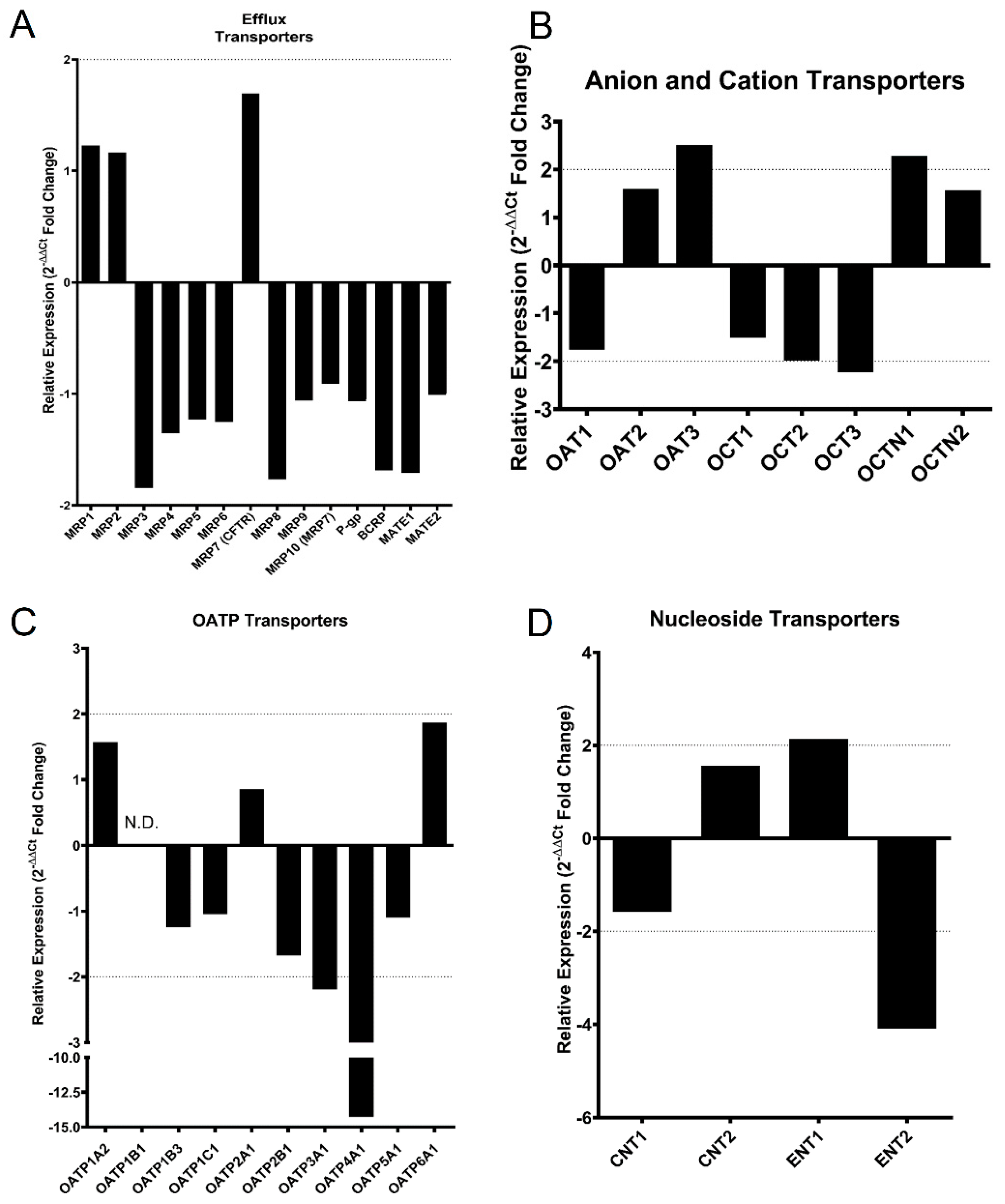
3.4. mRNA Expression of Common Xenobiotic Transporters
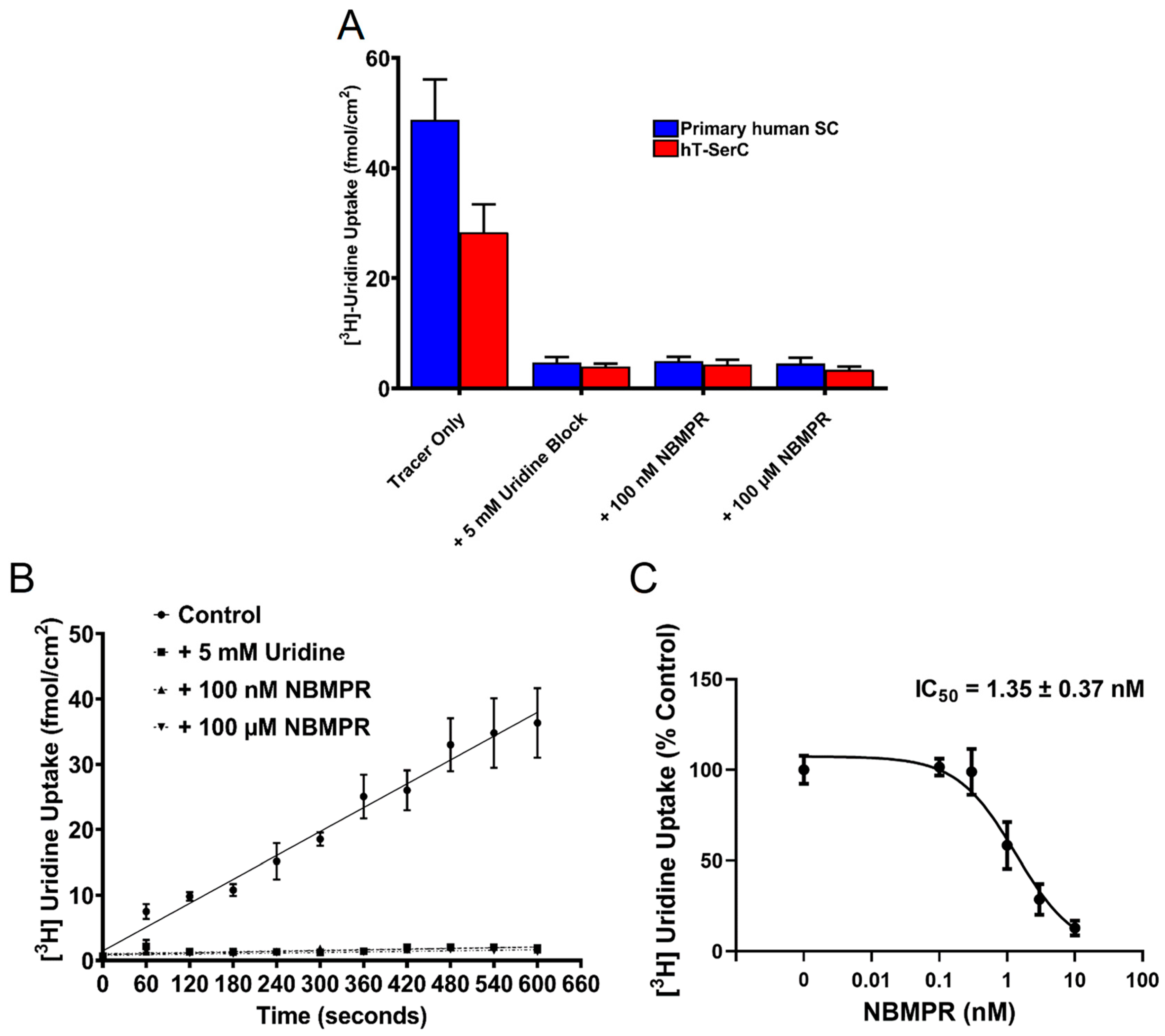
3.5. Inhibition of [3H] Uridine Uptake by NBMPR
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Griswold, M.D. The central role of Sertoli cells in spermatogenesis. Semin. Cell Dev. Biol. 1998, 9, 411–416. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griswold, M.D. 50 years of spermatogenesis: Sertoli cells and their interactions with germ cells. Biol. Reprod. 2018, 99, 87–100. [Google Scholar] [CrossRef] [PubMed]
- Bockers, T.M.; Nieschlag, E.; Kreutz, M.R.; Bergmann, M. Localization of follicle-stimulating hormone (FSH) immunoreactivity and hormone receptor mRNA in testicular tissue of infertile men. Cell Tissue Res. 1994, 278, 595–600. [Google Scholar] [CrossRef] [PubMed]
- Heckert, L.L.; Griswold, M.D. Expression of Follicle-Stimulating Hormone Receptor mRNA in Rat Testes and Sertoli Cells. Mol. Endocrinol. 1991, 5, 670–677. [Google Scholar] [CrossRef] [Green Version]
- Da Silva, S.M.; Hacker, A.; Harley, V.; Goodfellow, P.; Swain, A.; Lovell-Badge, R. Sox9 expression during gonadal development implies a conserved role for the gene in testis differentiation in mammals and birds. Nat. Genet. 1996, 14, 62–68. [Google Scholar] [CrossRef]
- Pelletier, R.-M. The blood-testis barrier: The junctional permeability, the proteins and the lipids. Prog. Histochem. Cytochem. 2011, 46, 49–127. [Google Scholar] [CrossRef]
- Stammler, A.; Lüftner, B.U.; Kliesch, S.; Weidner, W.; Bergmann, M.; Middendorff, R.; Konrad, L. Highly Conserved Testicular Localization of Claudin-11 in Normal and Impaired Spermatogenesis. PLoS ONE 2016, 11, e0160349. [Google Scholar] [CrossRef] [Green Version]
- Cheng, C.Y.; Mruk, D.D. The Blood-Testis Barrier and Its Implications for Male Contraception. Pharmacol. Rev. 2011, 64, 16–64. [Google Scholar] [CrossRef] [Green Version]
- Dym, M.; Fawcett, D.W. The Blood-Testis Barrier in the Rat and the Physiological Compartmentation of the Seminiferous Epithelium. Biol. Reprod. 1970, 3, 308–326. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.R.; Cherrington, N.J. Transepithelial Transport Across the Blood-Testis Barrier. Reproduction 2018, 156, R187–R194. [Google Scholar] [CrossRef]
- Cheng, C.Y.; Mruk, D.; Silvestrini, B.; Bonanomi, M.; Wong, C.-H.; Siu, M.K.; Lee, N.P.; Lui, W.-Y.; Mo, M.-Y. AF-2364 [1-(2,4-dichlorobenzyl)-1H-indazole-3-carbohydrazide] is a potential male contraceptive: A review of recent data. Contraception 2005, 72, 251–261. [Google Scholar] [CrossRef] [PubMed]
- Grima, J.; Silvestrini, B.; Cheng, C.Y. Reversible inhibition of spermatogenesis in rats using a new male contraceptive, 1-(2,4-dichlorobenzyl)-indazole-3-carbohydrazide. Biol. Reprod. 2001, 64, 1500–1508. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, R.; Maeda, T.; Akaike, T.; Tamai, I. Nucleoside Transport at the Blood-Testis Barrier Studied with Primary-Cultured Sertoli Cells. J. Pharmacol. Exp. Ther. 2004, 312, 601–608. [Google Scholar] [CrossRef] [Green Version]
- Klein, D.M.; Evans, K.K.; Hardwick, R.N.; Dantzler, W.H.; Wright, S.H.; Cherrington, N.J. Basolateral uptake of nucleosides by Sertoli cells is mediated primarily by equilibrative nucleoside transporter. J. Pharmacol. Exp. Ther. 2013, 346, 121–129. [Google Scholar] [CrossRef] [Green Version]
- Tash, J.S.; Attardi, B.; Hild, S.A.; Chakrasali, R.; Jakkaraj, S.R.; Georg, G.I. A Novel Potent Indazole Carboxylic Acid Derivative Blocks Spermatogenesis and Is Contraceptive in Rats after a Single Oral Dose. Biol. Reprod. 2008, 78, 1127–1138. [Google Scholar] [CrossRef] [Green Version]
- Tash, J.S.; Chakrasali, R.; Jakkaraj, S.R.; Hughes, J.; Smith, S.K.; Hornbaker, K.; Heckert, L.L.; Ozturk, S.B.; Hadden, M.K.; Kinzy, T.G.; et al. Gamendazole, an Orally Active Indazole Carboxylic Acid Male Contraceptive Agent, Targets HSP90AB1 (HSP90BETA) and EEF1A1 (eEF1A), and Stimulates Il1a Transcription in Rat Sertoli Cells. Biol. Reprod. 2008, 78, 1139–1152. [Google Scholar] [CrossRef] [Green Version]
- Elsheikh, M.G.; Hosny, M.B.; ElShenoufy, A.; Elghamrawi, H.; Fayad, A.; Abdelrahman, S. Combination of vitamin E and clomiphene citrate in treating patients with idiopathic oligoasthenozoospermia: A prospective, randomized trial. Andrology 2015, 3, 864–867. [Google Scholar] [CrossRef]
- Kobayashi, D.; Goto, A.; Maeda, T.; Nezu, J.-I.; Tsuji, A.; Tamai, I. OCTN2-mediated transport of carnitine in isolated Sertoli cells. Reproduction 2005, 129, 729–736. [Google Scholar] [CrossRef] [Green Version]
- Maeda, T.; Goto, A.; Kobayashi, D.; Tamai, I. Transport of Organic Cations across the Blood−Testis Barrier. Mol. Pharm. 2007, 4, 600–607. [Google Scholar] [CrossRef]
- Agarwal, A.; Said, T.M. Carnitines and male infertility. Reprod. Biomed. Online 2004, 8, 376–384. [Google Scholar] [CrossRef]
- Griffith, D.A.; Jarvis, S.M. Nucleoside and nucleobase transport systems of mammalian cells. Biochim. et Biophys. Acta (BBA) Rev. Biomembr. 1996, 1286, 153–181. [Google Scholar] [CrossRef]
- Griffiths, M.; Yao, Y.M.S.; Abidi, F.; Phillips, S.E.; Cass, E.C.; Young, D.J.; Baldwin, A.S. Molecular cloning and characterization of a nitrobenzylthioinosine-insensitive (ei) equilibrative nucleoside transporter from human placenta. Biochem. J. 1997, 328, 739–743. [Google Scholar] [CrossRef]
- Yao, S.Y.M.; Ng, A.M.L.; Muzyka, W.R.; Griffiths, M.; Cass, C.E.; Baldwin, S.A.; Young, J.D. Molecular Cloning and Functional Characterization of Nitrobenzylthioinosine (NBMPR)-sensitive (es) and NBMPR-insensitive (ei) Equilibrative Nucleoside Transporter Proteins (rENT1 and rENT2) from Rat Tissues. J. Biol. Chem. 1997, 272, 28423–28430. [Google Scholar] [CrossRef] [Green Version]
- Orth, J.M.; Gunsalus, G.L.; Lamperti, A.A. Evidence from Sertoli Cell-Depleted Rats Indicates That Spermatid Number in Adults Depends on Numbers of Sertoli Cells Produced During Perinatal Development. Endocrinology 1988, 122, 787–794. [Google Scholar] [CrossRef]
- Petersen, C.; Söder, O. The Sertoli Cell—A Hormonal Target and ‘Super’ Nurse for Germ Cells That Determines Testicular Size. Horm. Res. Paediatr. 2006, 66, 153–161. [Google Scholar] [CrossRef]
- Russell, L.D.; Bartke, A.; Goh, J.C. Postnatal development of the sertoli cell barrier, tubular lumen, and cytoskeleton of sertoli and myoid cells in the rat, and their relationship to tubular fluid secretion and flow. Am. J. Anat. 1989, 184, 179–189. [Google Scholar] [CrossRef]
- Sharpe, R.M.; McKinnell, C.; Kivlin, C.; Fisher, J.S. Proliferation and functional maturation of Sertoli cells, and their relevance to disorders of testis function in adulthood. Reproduction 2003, 125, 769–784. [Google Scholar] [CrossRef]
- Chui, K.; Trivedi, A.; Cheng, C.Y.; Cherbavaz, D.B.; Dazin, P.F.; Huynh, A.L.T.; Mitchell, J.B.; Rabinovich, G.A.; Noble-Haeusslein, L.J.; John, C.M. Characterization and Functionality of Proliferative Human Sertoli Cells. Cell Transplant. 2011, 20, 619–635. [Google Scholar] [CrossRef] [Green Version]
- Gaur, M.; Ramathal, C.; Pera, R.A.R.; Turek, P.J.; John, C.M. Isolation of human testicular cells and co-culture with embryonic stem cells. Reproduction 2018, 155, 151–164. [Google Scholar] [CrossRef]
- Guo, Y.; Hai, Y.; Yao, C.; Chen, Z.; Hou, J.; Li, Z.; He, Z. Long-term culture and significant expansion of human Sertoli cells whilst maintaining stable global phenotype and AKT and SMAD1/5 activation. Cell Commun. Signal. 2015, 13, 20. [Google Scholar] [CrossRef] [Green Version]
- Wen, L.; Yuan, Q.; Sun, M.; Niu, M.; Wang, H.; Fu, H.; Zhou, F.; Yao, C.; Wang, X.; Li, Z.; et al. Generation and characteristics of human Sertoli cell line immortalized by overexpression of human telomerase. Oncotarget 2017, 8, 16553–16570. [Google Scholar] [CrossRef] [Green Version]
- Hayflick, L. The limited in vitro lifetime of human diploid cell strains. Exp. Cell Res. 1965, 37, 614–636. [Google Scholar] [CrossRef]
- Olovnikov, A.M. Telomeres, telomerase, and aging: Origin of the theory. Exp. Gerontol. 1996, 31, 443–448. [Google Scholar] [CrossRef]
- Counter, C.M.; Hahn, W.C.; Wei, W.; Caddle, S.D.; Beijersbergen, R.L.; Lansdorp, P.M.; Sedivy, J.M.; Weinberg, R.A. Dissociation among in vitro telomerase activity, telomere maintenance, and cellular immortalization. Proc. Natl. Acad. Sci. USA 1998, 95, 14723–14728. [Google Scholar] [CrossRef] [Green Version]
- Stanford University Nolan Lab. Available online: http://web.stanford.edu/group/nolan/_OldWebsite/retroviral_systems/phx.html (accessed on 8 August 2020).
- Swift, S.; Lorens, J.; Achacoso, P.; Nolan, G.P. Rapid Production of Retroviruses for Efficient Gene Delivery to Mammalian Cells Using 293TCell–Based Systems. Curr. Protoc. Immunol. 1999, 31, 10–17. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Miller, S.R.; Hau, R.K.; Jilek, J.L.; Morales, M.N.; Wright, S.H.; Cherrington, N.J. Nucleoside Reverse Transcriptase Inhibitor Interaction with Human Equilibrative Nucleoside Transporters 1 and 2. Drug Metab. Dispos. 2020, 48, 603–612. [Google Scholar] [CrossRef]
- Sandoval, P.J.; Zorn, K.M.; Clark, A.M.; Ekins, S.; Wright, S.H. Assessment of Substrate-Dependent Ligand Interactions at the Organic Cation Transporter OCT2 Using Six Model Substrates. Mol. Pharmacol. 2018, 94, 1057–1068. [Google Scholar] [CrossRef] [Green Version]
- Severance, A.C.; Sandoval, P.J.; Wright, S.H. Correlation between Apparent Substrate Affinity and OCT2 Transport Turnover. J. Pharmacol. Exp. Ther. 2017, 362, 405–412. [Google Scholar] [CrossRef]
- Paniagua, R.; Nistal, M.; Amat, P. Ultrastructural observations on nucleoli and related structures during human spermatogenesis. Brain Struct. Funct. 1986, 174, 301–306. [Google Scholar] [CrossRef]
- Kraaij, R.; Verhoef-Post, M.; A Grootegoed, J.; Themmen, A.P. Alternative splicing of follicle-stimulating hormone receptor pre-mRNA: Cloning and characterization of two alternatively spliced mRNA transcripts. J. Endocrinol. 1998, 158, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Mattiske, D.; Pask, A.J.; Shaw, J.; Shaw, G. Structure and expression of the follicle-stimulating hormone receptor gene in a marsupial, Macropus eugenii. Mol. Reprod. Dev. 2002, 63, 24–31. [Google Scholar] [CrossRef]
- James, K.; Bhartiya, D.; Ganguly, R.; Kaushik, A.; Gala, K.; Singh, P.; Metkari, S.M. Gonadotropin and steroid hormones regulate pluripotent very small embryonic-like stem cells in adult mouse uterine endometrium. J. Ovarian Res. 2018, 11, 83. [Google Scholar] [CrossRef] [Green Version]
- Bart, J.; Hollema, H.; Groen, H.; De Vries, E.G.; Hendrikse, N.; Sleijfer, D.; Wegman, T.; Vaalburg, W.; Van Der Graaf, W.T.A. The distribution of drug-efflux pumps, P-gp, BCRP, MRP1 and MRP2, in the normal blood–testis barrier and in primary testicular tumours. Eur. J. Cancer 2004, 40, 2064–2070. [Google Scholar] [CrossRef]
- Klein, D.M.; Wright, S.H.; Cherrington, N.J. Localization of Multidrug Resistance-Associated Proteins along the Blood-Testis Barrier in Rat, Macaque, and Human Testis. Drug Metab. Dispos. 2013, 42, 89–93. [Google Scholar] [CrossRef] [Green Version]
- Koraichi, F.; Inoubli, L.; Lakhdari, N.; Meunier, L.; Vega, A.; Mauduit, C.; Benahmed, M.; Prouillac, C.; Lecoeur, S. Neonatal exposure to zearalenone induces long term modulation of ABC transporter expression in testis. Toxicology 2013, 310, 29–38. [Google Scholar] [CrossRef]
- Kort, A.; Durmus, S.; Sparidans, R.W.; Wagenaar, E.; Beijnen, J.H.; Schinkel, A.H. Brain and Testis Accumulation of Regorafenib is Restricted by Breast Cancer Resistance Protein (BCRP/ABCG2) and P-glycoprotein (P-GP/ABCB1). Pharm. Res. 2015, 32, 2205–2216. [Google Scholar] [CrossRef] [Green Version]
- Melaine, N.; Liénard, M.-O.; Dorval, I.; Le Goascogne, C.; Lejeune, H.; Jégou, B. Multidrug Resistance Genes and P-Glycoprotein in the Testis of the Rat, Mouse, Guinea Pig, and Human. Biol. Reprod. 2002, 67, 1699–1707. [Google Scholar] [CrossRef] [Green Version]
- Robillard, K.R.; Hoque, T.; Bendayan, R. Expression of ATP-Binding Cassette Membrane Transporters in Rodent and Human Sertoli Cells: Relevance to the Permeability of Antiretroviral Therapy at the Blood-Testis Barrier. J. Pharmacol. Exp. Ther. 2011, 340, 96–108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, L.; Cheng, C.Y.; Mruk, D.D. Drug transporter, P-glycoprotein (MDR1), is an integrated component of the mammalian blood–testis barrier. Int. J. Biochem. Cell Biol. 2009, 41, 2578–2587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wijnholds, J.; Scheffer, G.L.; Van Der Valk, M.; Van Der Valk, P.; Beijnen, J.H.; Scheper, R.J.; Borst, P. Multidrug Resistance Protein 1 Protects the Oropharyngeal Mucosal Layer and the Testicular Tubules against Drug-induced Damage. J. Exp. Med. 1998, 188, 797–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiss, A.; Farah, K.; Kim, J.; Garriock, R.J.; Drysdale, T.A.; Hammond, J.R. Molecular cloning and functional characterization of inhibitor-sensitive (mENT1) and inhibitor-resistant (mENT2) equilibrative nucleoside transporters from mouse brain. Biochem. J. 2000, 352, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Plagemann, P.G.; Wohlhueter, R.M. Nucleoside transport in cultured mammalian cells multiple forms with different sensitivity to inhibition by nitrobenzylthioinosine or hypoxanthine. Biochim. et Biophys. Acta (BBA) Biomembr. 1984, 773, 39–52. [Google Scholar] [CrossRef]
- Ward, J.L.; Sherali, A.; Mo, Z.-P.; Tse, C.-M. Kinetic and Pharmacological Properties of Cloned Human Equilibrative Nucleoside Transporters, ENT1 and ENT2, Stably Expressed in Nucleoside Transporter-deficient PK15 Cells. Ent2 exhibits a low affinity for guanosine and cytidine but a high affinity for inosine. J. Biol. Chem. 2000, 275, 8375–8381. [Google Scholar] [CrossRef] [Green Version]
- Kato, R.; Maeda, T.; Akaike, T.; Tamai, I. Characterization of Nucleobase Transport by Mouse Sertoli Cell Line TM4. Biol. Pharm. Bull. 2009, 32, 450–455. [Google Scholar] [CrossRef] [Green Version]
- Groves, C.E.; Evans, K.K.; Dantzler, W.H.; Wright, S.H. Peritubular organic cation transport in isolated rabbit proximal tubules. Am. J. Physiol. Physiol. 1994, 266, F450–F458. [Google Scholar] [CrossRef]
- Mruk, D.D.; Cheng, C.Y. An In Vitro System to Study Sertoli Cell Blood-Testis Barrier Dynamics. Cartil. Tissue Eng. 2011, 763, 237–252. [Google Scholar] [CrossRef] [Green Version]
- Selva, D.; Hirsch-Reinshagen, V.; Burgess, B.; Zhou, S.; Chan, J.; McIsaac, S.; Hayden, M.R.; Hammond, G.L.; Vogl, A.W.; Wellington, C. The ATP-binding cassette transporter 1 mediates lipid efflux from Sertoli cells and influences male fertility. J. Lipid Res. 2004, 45, 1040–1050. [Google Scholar] [CrossRef] [Green Version]
- Chu, X.; Bleasby, K.; Evers, R. Species differences in drug transporters and implications for translating preclinical findings to humans. Expert Opin. Drug Metab. Toxicol. 2012, 9, 237–252. [Google Scholar] [CrossRef]
- Srinivasan, B.; Kolli, A.R.; Esch, M.B.; Abaci, H.E.; Shuler, M.L.; Hickman, J.J. TEER Measurement Techniques for In Vitro Barrier Model Systems. J. Lab. Autom. 2015, 20, 107–126. [Google Scholar] [CrossRef] [Green Version]
- Prozialeck, W.C.; Lamar, P.C. Cadmium (Cd2+) disrupts E-cadherin-dependent cell-cell junctions in MDCK cells. Vitr. Cell. Dev. Biol. Anim. 1997, 33, 516–526. [Google Scholar] [CrossRef] [PubMed]
- Ishii, K.; Kanatsu-Shinohara, M.; Toyokuni, S.; Shinohara, T. FGF2 mediates mouse spermatogonial stem cell self-renewal via upregulation of Etv5 and Bcl6b through MAP2K1 activation. Development 2012, 139, 1734–1743. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meng, X.; Lindahl, M.; Hyvönen, M.E.; Parvinen, M.; De Rooij, D.G.; Hess, M.W.; Raatikainen-Ahokas, A.; Sainio, K.; Rauvala, H.; Lakso, M.; et al. Regulation of Cell Fate Decision of Undifferentiated Spermatogonia by GDNF. Science 2000, 287, 1489–1493. [Google Scholar] [CrossRef]
- Fröjdman, K.; Harley, V.R.; Pelliniemi, L.J. Sox9 protein in rat Sertoli cells is age and stage dependent. Histochem. Cell Biol. 2000, 113, 31–36. [Google Scholar] [CrossRef]
- Pellegrini, M.; Grimaldi, P.; Rossi, P.; Geremia, R.; Dolci, S. Developmental expression of BMP4/ALK3/SMAD5 signaling pathway in the mouse testis: A potential role of BMP4 in spermatogonia differentiation. J. Cell Sci. 2003, 116, 3363–3372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsushita, K.; Miyake, H.; Fujisawa, M. Clusterin produced by Sertoli cells inhibits heat stress-induced apoptosis in the rat testis. Andrologia 2016, 48, 11–19. [Google Scholar] [CrossRef]
- Augustine, L.M.; Markelewicz, R.J.; Boekelheide, K.; Cherrington, N.J. Xenobiotic and endobiotic transporter mRNA expression in the blood-testis barrier. Drug Metab. Dispos. 2005, 33, 182–189. [Google Scholar] [CrossRef] [Green Version]
- Klein, D. Nucleoside and HIV Drug Transport at the Blood-Testis Barrier. Ph.D. Thesis, The University of Arizona, Tucson, AZ, USA, 8 September 2015. [Google Scholar]
- Grañé-Boladeras, N.; Spring, C.M.; Hanna, W.J.B.; Pastor-Anglada, M.; Coe, I.R. Novel nuclear hENT2 isoforms regulate cell cycle progression via controlling nucleoside transport and nuclear reservoir. Cell. Mol. Life Sci. 2016, 73, 4559–4575. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Forward Primer | Reverse Primer |
|---|---|---|
| GAPDH | 5′-CGACCACTTTGTCAAGCTCA-3′ | 5′-CCCTGTTGCTGTAGCCAAAT-3′ |
| TERT | 5′-CGGTGTGCACCAACATTAC-3′ | 5′-GGGTTCTTCCAAACTTGCTG-3′ |
| AR | 5′-CCATAAGCCACTTGGATGCT-3′ | 5′-TGTCATGTCTGAGGCACTCC-3′ |
| FSHR | 5′-AAATGGAGCTTGCATTCTGG-3′ | 5′-TCCTTCCCAGATTCTCCTGA-‘3 |
| GDNF | 5′-TCTTCATGGTTCTGCCCTTC-3′ | 5′-GCTGGGGTTTGTCACTGTTT-3′ |
| SOX9 | 5′-AGACCTTTGGGCTGCCTTAT-3′ | 5′-TAGCCTCCCTCACTCCAAGA-3′ |
| BMP4 | 5′-ACTGGCTGACCACCTCACT-3′ | 5′-GTTCAGTGGGCACACAACAG-3′ |
| CLDN11 | 5′-TTCACGGTATTGCAGTGGTAA-3′ | 5′-GTTTCTGATTGCTGCCCATT-3′ |
| CLU | 5′-GCTGCAAATGGAAGCTTTTC-3′ | 5′-TTCTGGGCACCAAATGTTTT-3′ |
| FASLG | 5′-CCATGTGAAGAGGGAGAAGC-3′ | 5′-AAGACAGTCCCCCTTGAGGT-3′ |
| FGF2 | 5′-CCATCCTTTCTCCCTCGTTT-3′ | 5′-TTCCCTCCAATGTTTCATTCA-3′ |
| INHBB | 5′-TGAACGCACATGACATAGCA-3′ | 5′-ACGTGGCACTTGGACATCTA-3′ |
| OCLN | 5′-ATGCCTAGCTACCCCATCT-3′ | 5′-AATGCCAATCCTGCATTCTC-3′ |
| SHBG | 5′-CTCCCCTCCTTAACCTCTGG-3′ | 5′-AGAGGTTTCCTTCCCCTCAA-3′ |
| TF | 5′-AAGCCTGCACTTTCCGTAGA-3′ | 5′-AAGCCTGCACTTTCCGTAGA-3′ |
| WT1 | 5′-TACCTCCTTGCACAAATGGA-3′ | 5′-CCTGGACCATCCCCTATTTT-3′ |
| MRP1 | 5′-CTCGTTAGAGCCCAAAGTGG-3′ | 5′-ACAAAAGGATCCCCCAAAAC-3′ |
| MRP2 | 5′-CCGTATCAGGTTTGCCAGTT-3′ | 5′-TGGAGGTGATCCAGGAAAAG-3′ |
| MRP3 | 5′-GGCACTGCTGATTGAAGACA-3′ | 5′-TGTCACCTGCACCTTCTCTG-3′ |
| MRP4 | 5′-TCCTGATGATGGTGGCTGTA-3′ | 5′-ATGCAATTTCAGGGAGGTGA-3′ |
| MRP5 | 5′-AGTTCTGTTTGTTACCCACCAGTT-3′ | 5′-ACCCTTGTCTTGTGACTTCTTCTG-3′ |
| MRP6 | 5′-GCTCTATCCTCAGGAACTCGAAGAC-3′ | 5′-GCTTTCTCTGCATTCATAGCATTCT |
| MRP7 (CFTR) | 5′-CTACTCTCCTTCGCCACATTTTC-3′ | 5′-TGGCTCAGAGAGGCCTTCTC-3′ |
| MRP8 | 5′-CTCCTCAGGGATTTTCACCA-3′ | 5′-AGGACCAGGAACTGCTCTGA-3′ |
| MRP9 | 5′-CCAAGACTGACACCCTGGTT-3′ | 5′-GACGTGATCGCAGRRGAGA-3′ |
| MRP10 (MRP7) | 5′-AAAGGGCACCCTGGATTACT-3′ | 5′-AATGCAAGTGGGCTCCTATG-3′ |
| P-gp | 5′-GCCAAAGCCAAAATATCAGC-3′ | 5′-TTCCAATGTGTTCGGCATTA-3′ |
| BCRP | 5′-CCCTGACATTCTGTCACAACA-3′ | 5′-GGGACAGGTATGTGAAAAGCA-3′ |
| MATE1 | 5′-ATGCTGTTTCCCACCTCTTTG-3′ | 5′-CCGAGGCACGTTGTTTACTT-3′ |
| MATE2 | 5′-TGGGGCATATTTTTACCAATG-3′ | 5′-GAACTCGCCCATAGACACAAC-3′ |
| OAT1 | 5′-CAGCAACAAGAGCACCAGAA-3′ | 5′-TGGGTCACCATTTCCTCTTC-3′ |
| OAT2 | 5′-CCCAAGGGACAAAAAGAACA-3′ | 5′-ATGAGACCAGTGGGTTGGAG-3′ |
| OAT3 | 5′-TCCCAGAGGATCCCTCTACA-3′ | 5′-TGCCTGGCTAGGATCAGTCT-3′ |
| OCT1 | 5′-TGGAGGCATGGTGAAATACA-3′ | 5′-GTCACCCACTTCCGTGATCT-3′ |
| OCT2 | 5′-GATCCTGCCAAATTCTTCCA-3′ | 5′-TAGCCCACAGTTCCCCTATG-3′ |
| OCT3 | 5′-ATGGTTGCTGAACCCAAAAC-3′ | 5′-CCCAGATCGTTAAACCCAGA-3′ |
| OCTN1 | 5′-ACCCCATTTGGTGAAGTGAA-3′ | 5′-GGTTGGTTTGTAAAGCAAGGAC-3′ |
| OCTN2 | 5′-ATTTGCCCTTCAGAATGCAC-3′ | 5′-GCAGACAATTGCCAGAGTGA-3′ |
| CNT1 | 5′-AGGTCCTGCCCATCATTGTC-3′ | 5′-CAAGTAGGGCCGGATCAGTA-3′ |
| CNT2 | 5′-AATGGGTGTTTGCAGGAGTC-3′ | 5′-GAAGACCTAGGCCCGAAAAC-3′ |
| ENT1 | 5′-GCTGGGTCTGACCGTTGTAT-3′ | 5′-CTGTACAGGGTGCATGATGG-3′ |
| ENT2 | 5′-AGCCTGCATGTGTGTACTGC-3′ | 5′-ACCACGGACCAGTCACTTTC-3′ |
| OATP1A2 | 5′-GCATCAAGGGCAGATGATTT-3′ | 5′-GGCTGGGAAGTCAAGAGATG-3′ |
| OATP1B1 | 5′-GCTGGGGCAGATAGTGAAAC-3′ | 5′-GGACCAGGAACTCCTCAAAA-3′ |
| OATP1B3 | 5′-GAAAAGGTTGTTTAAAGGAATCTGG-3′ | 5′-CGAAATCATCAATGTAAGAAAGCC-3′ |
| OATP1C1 | 5′-ACTCCCATTCAGCCTTTGGG-3′ | 5′-CAGAAAGGCACAGCTGCAAG-3′ |
| OATP2A1 | 5′-TTCCAAAGCCACCTCATTTC-3′ | 5′-GGTTAGTTGCAGGGCATCAT-3′ |
| OATP2B1 | 5′-GGCTTTGAGACTTTCCCACA-3′ | 5′-CTGGGAAACAAGAGGGATGA-3′ |
| OATP3A1 | 5′-CAGGCCATGCTCTCCGAAA-3′ | 5′-CTGCTGCTCCAGGTACTTCC-3′ |
| OATP4A1 | 5′-CTGCCAGCCAGAACACTACA-3′ | 5′-AGAAGGAGGGGCTTTCTCTG-3′ |
| OATP5A1 | 5′-TCATGCTCCCCTACGGTACAG-3′ | 5′-GCTCACCTTTGTTTGGAGTGTTAG-3′ |
| OATP6A1 | 5′-GGAGCCAGGATGAAGTCTCAA-3′ | 5′-GAACCTTATCAAGGCCTCTGGAAG-3′ |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hau, R.K.; Miller, S.R.; Wright, S.H.; Cherrington, N.J. Generation of a hTERT-Immortalized Human Sertoli Cell Model to Study Transporter Dynamics at the Blood-Testis Barrier. Pharmaceutics 2020, 12, 1005. https://doi.org/10.3390/pharmaceutics12111005
Hau RK, Miller SR, Wright SH, Cherrington NJ. Generation of a hTERT-Immortalized Human Sertoli Cell Model to Study Transporter Dynamics at the Blood-Testis Barrier. Pharmaceutics. 2020; 12(11):1005. https://doi.org/10.3390/pharmaceutics12111005
Chicago/Turabian StyleHau, Raymond K., Siennah R. Miller, Stephen H. Wright, and Nathan J. Cherrington. 2020. "Generation of a hTERT-Immortalized Human Sertoli Cell Model to Study Transporter Dynamics at the Blood-Testis Barrier" Pharmaceutics 12, no. 11: 1005. https://doi.org/10.3390/pharmaceutics12111005