Antimicrobial Biophotonic Treatment of Ampicillin-Resistant Pseudomonas aeruginosa with Hypericin and Ampicillin Cotreatment Followed by Orange Light

 ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. LED-Based Orange Light Source
2.3. Bacterial Cultures and Growth Conditions
2.4. APDT of Bacteria and Fungi
2.4.1. APDT of S. aureus and MRSA
2.4.2. APDT of C. albicans KCTC 7965
2.4.3. APDT of P. aeruginosa PAO1
2.5. Determination of Hypericin Intake in P. aeruginosa and S. aureus by Measuring the Fluorescence Intensity of Hypericin
2.6. C. elegans Strain and Maintenance
2.7. Toxicity Test of Orange Light on C. elegans
2.8. Evaluation of APDT in C. elegans by Measuring Growth Rate
2.9. Confirmation of APDT in C. elegans by Measuring Intestinal Permeability
2.10. Statistics
3. Results and Discussion
3.1. APDT Effectively Acts on the Gram-Positive Bacteria S. aureus and MRSA and the Fungus C. albicans
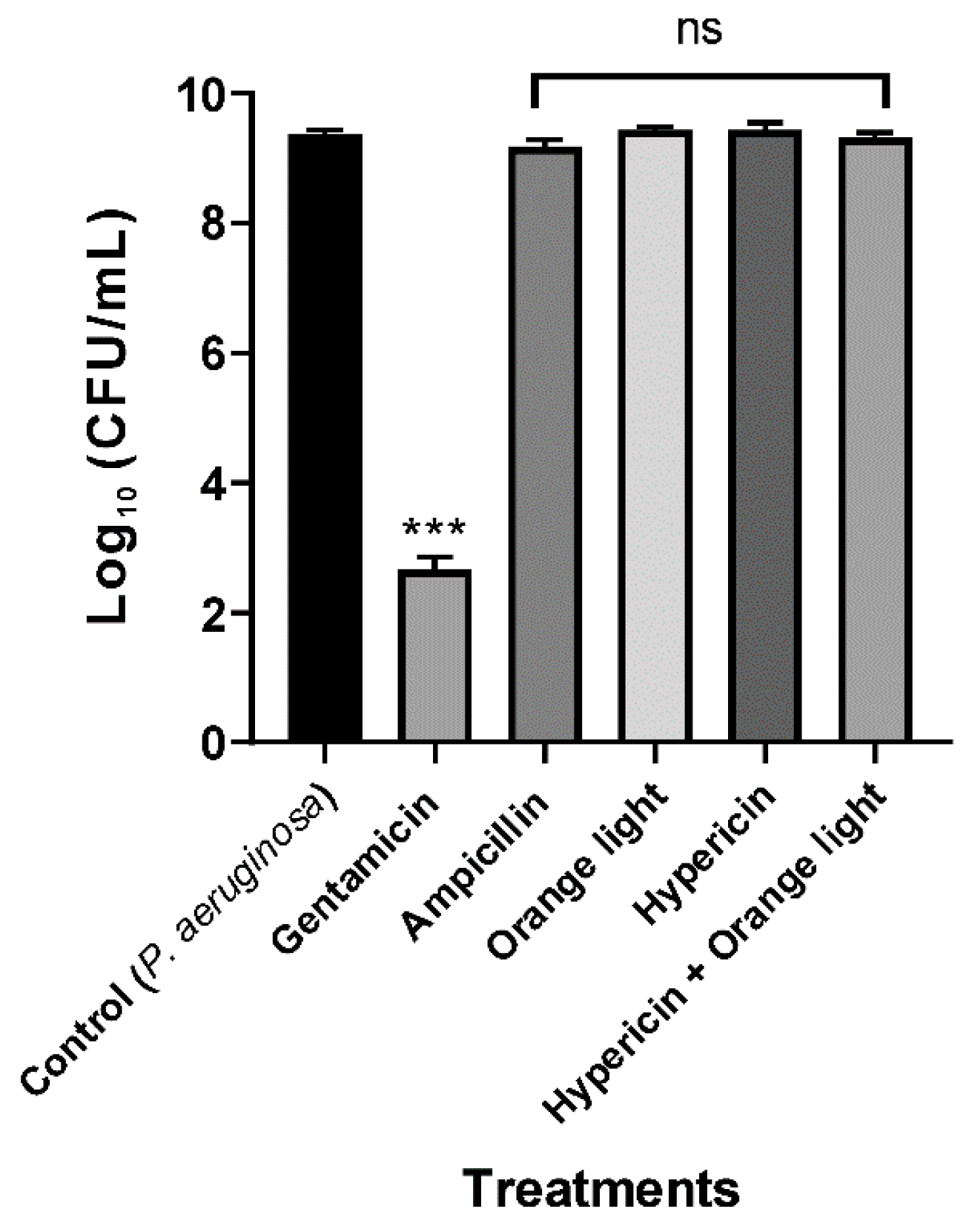
3.2. Conventional APDT with Hypericin and Orange Light Did Not Kill P. aeruginosa PAO1
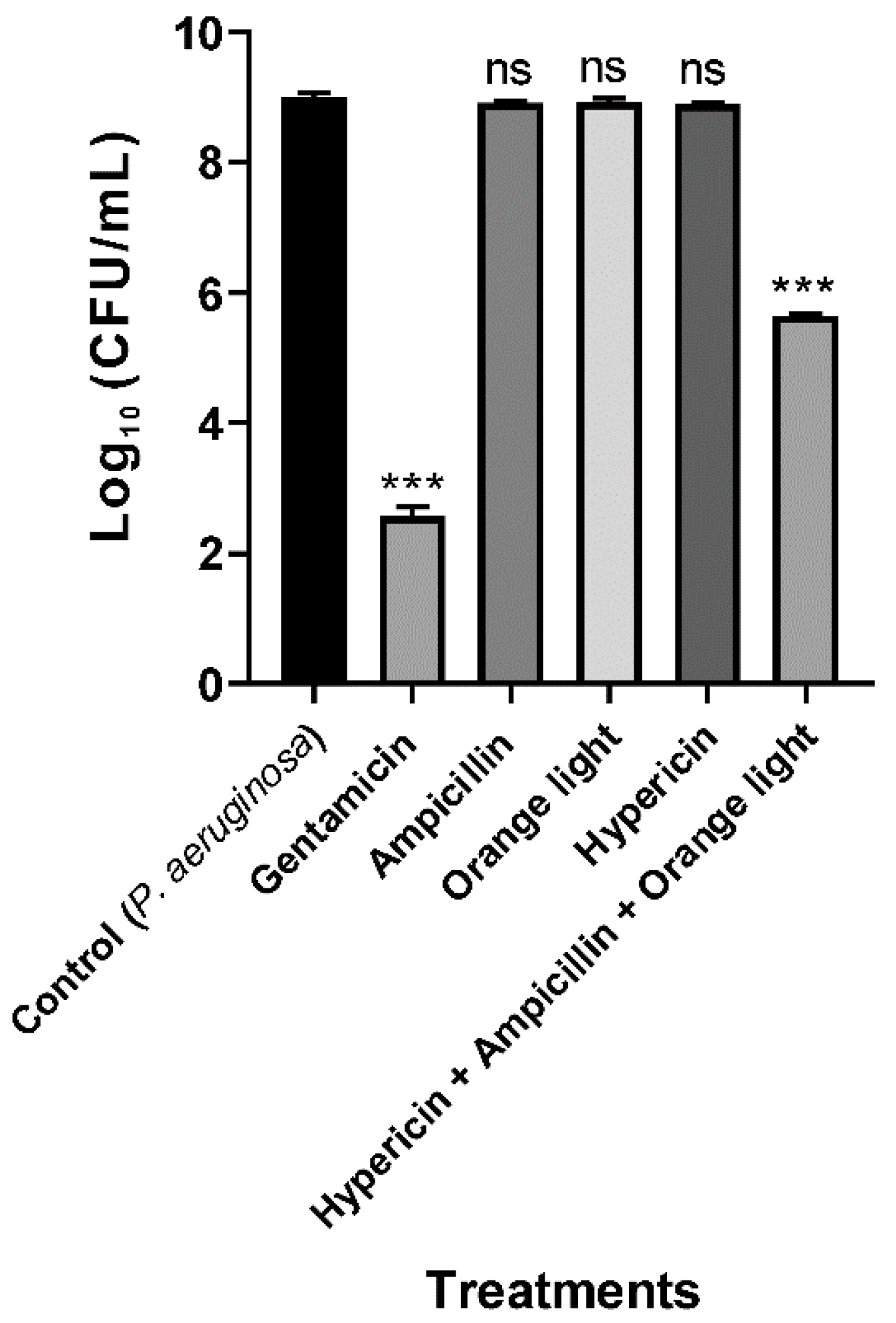
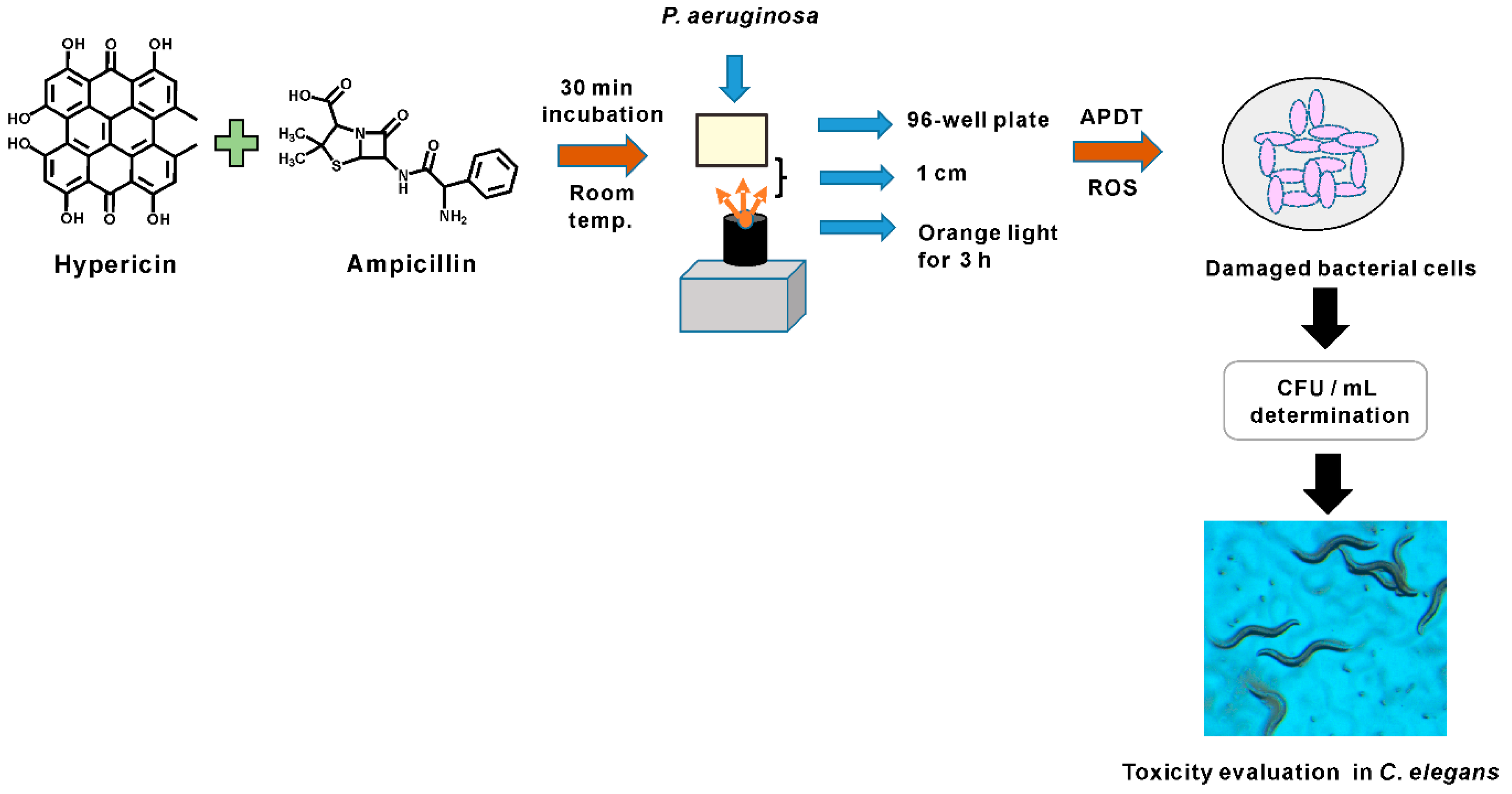
3.3. Hypericin and Ampicillin Cotreatment Followed by Orange Light Significantly Inhibited P. aeruginosa PAO1
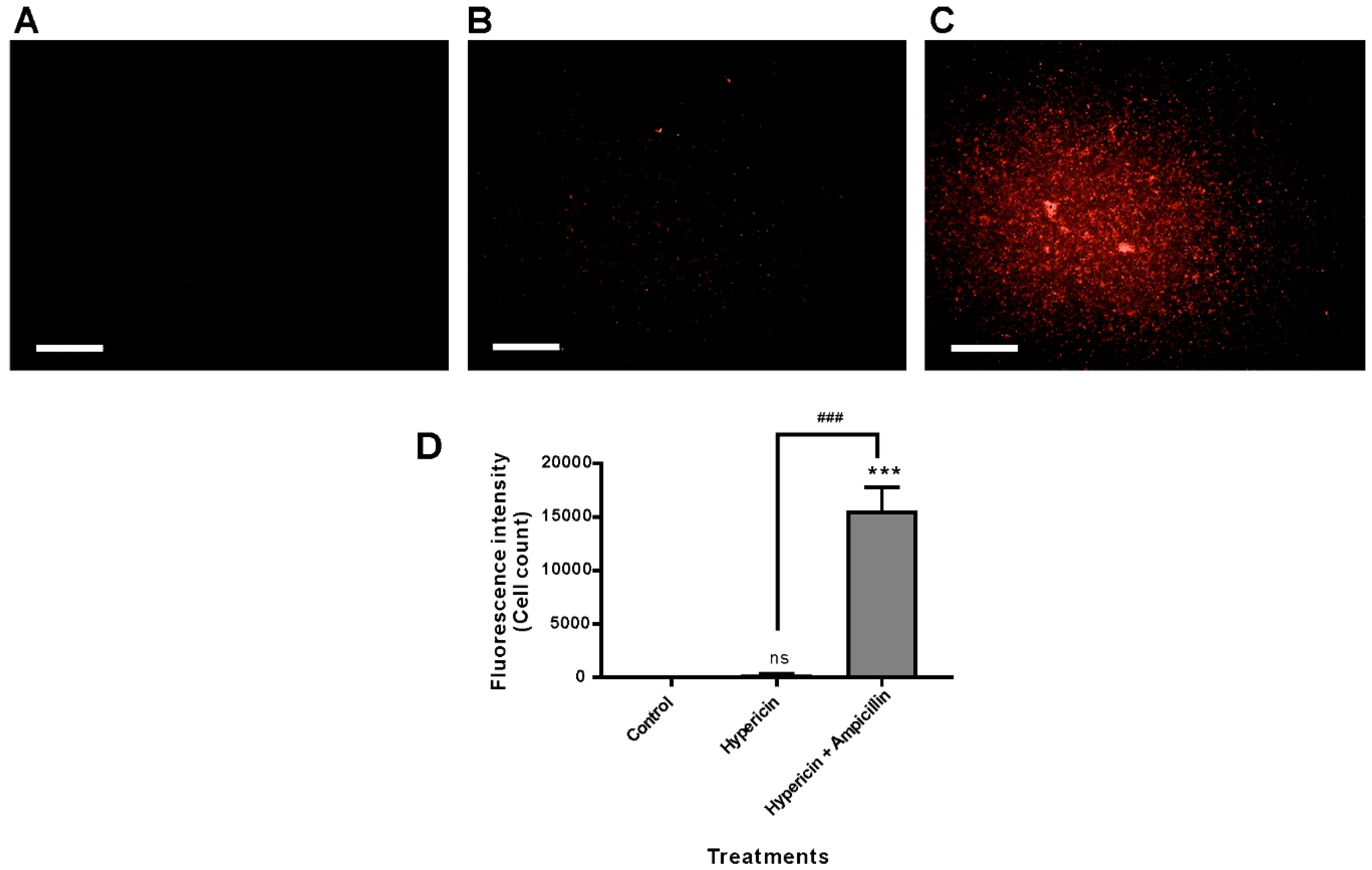
3.4. Ampicillin Facilitated Hypericin Uptake in the Gram-Negative Bacterium P. aeruginosa PAO1
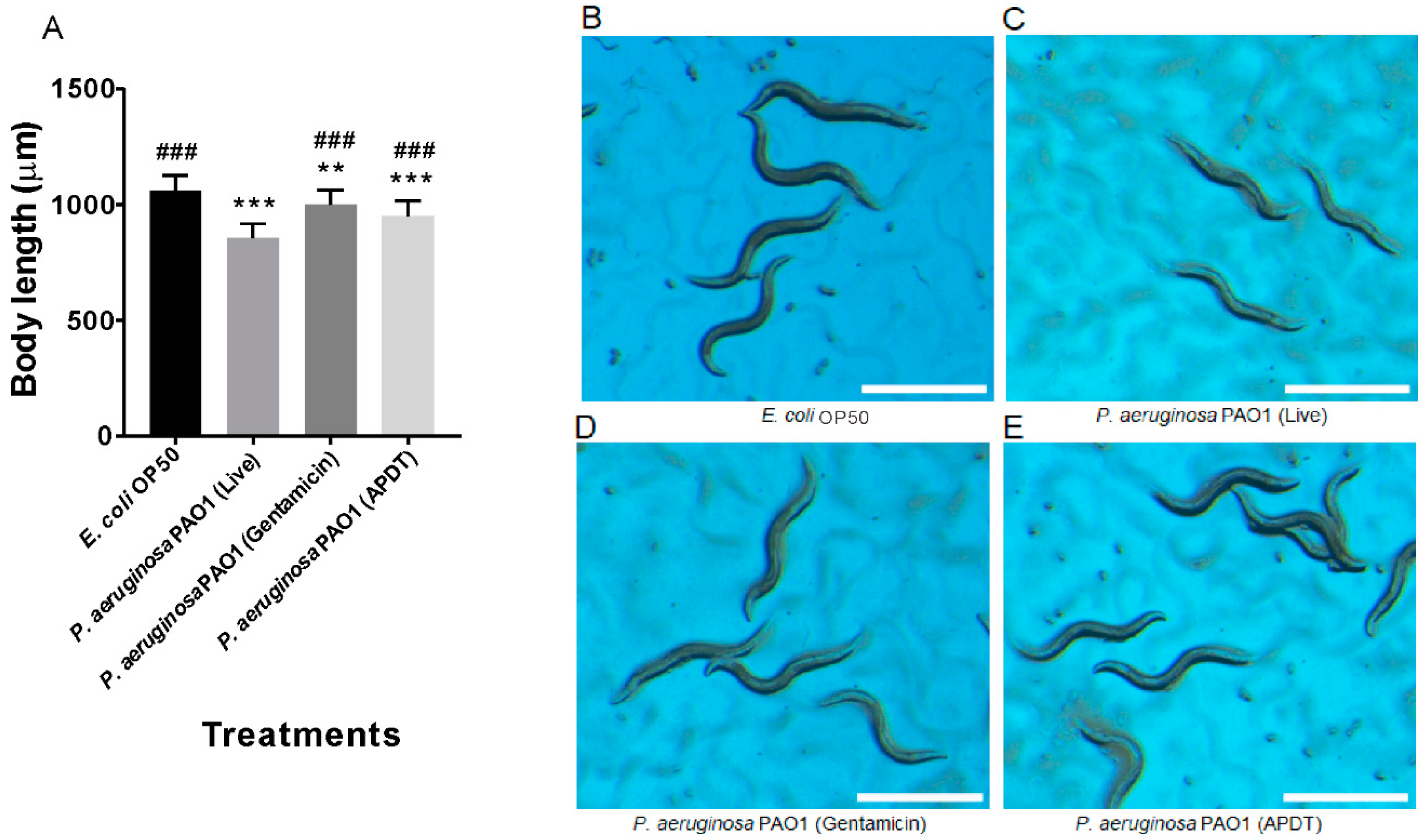
3.5. APDT with Ampicillin and Hypericin Was Evaluated in the C. elegans Model by Measuring Worm Growth
3.6. APDT Rescued Intestinal Permeability Dysfunction in C. elegans
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Huang, L.; Xuan, Y.; Koide, Y.; Zhiyentayev, T.; Tanaka, M.; Hamblin, M.R. Type I and Type II mechanisms of antimicrobial photodynamic therapy: An in vitro study on gram-negative and gram-positive bacteria. Lasers Surg. Med. 2012, 44, 490–499. [Google Scholar] [CrossRef] [PubMed]
- Harbarth, S.; Balkhy, H.H.; Goossens, H.; Jarlier, V.; Kluytmans, J.; Laxminarayan, R.; Saam, M.; Van Belkum, A.; Pittet, D. Antimicrobial resistance: One world, one fight! Antimicrob. Resist. Infect. Control 2015, 4, 49. [Google Scholar] [CrossRef]
- Santajit, S.; Indrawattana, N. Mechanisms of Antimicrobial Resistance in ESKAPE Pathogens. Biomed. Res. Int. 2016, 2016, 2475067. [Google Scholar] [CrossRef] [PubMed]
- Engelhardt, V.; Krammer, B.; Plaetzer, K. Antibacterial photodynamic therapy using water-soluble formulations of hypericin or mTHPC is effective in inactivation of Staphylococcus aureus. Photochem. Photobiol. Sci. 2010, 9, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Kiesslich, T.; Krammer, B.; Plaetzer, K. Cellular mechanisms and prospective applications of hypericin in photodynamic therapy. Curr. Med. Chem. 2006, 13, 2189–2204. [Google Scholar] [CrossRef]
- Kwiatkowski, S.; Knap, B.; Przystupski, D.; Saczko, J.; Kedzierska, E.; Knap-Czop, K.; Kotlinska, J.; Michel, O.; Kotowski, K.; Kulbacka, J. Photodynamic therapy—Mechanisms, photosensitizers and combinations. Biomed. Pharmacother. 2018, 106, 1098–1107. [Google Scholar] [CrossRef] [PubMed]
- Vatansever, F.; De Melo, W.C.; Avci, P.; Vecchio, D.; Sadasivam, M.; Gupta, A.; Chandran, R.; Karimi, M.; Parizotto, N.A.; Yin, R.; et al. Antimicrobial strategies centered around reactive oxygen species—Bactericidal antibiotics, photodynamic therapy, and beyond. FEMS Microbiol. Rev. 2013, 37, 955–989. [Google Scholar] [CrossRef]
- Greeson, J.M.; Sanford, B.; Monti, D.A. St. John’s wort (Hypericum perforatum): A review of the current pharmacological, toxicological, and clinical literature. Psychopharmacology 2001, 153, 402–414. [Google Scholar] [CrossRef]
- Barnes, J.; Anderson, L.A.; Phillipson, J.D. St John’s wort (Hypericum perforatum L.): A review of its chemistry, pharmacology and clinical properties. J. Pharm. Pharmacol. 2001, 53, 583–600. [Google Scholar] [CrossRef]
- García, I.; Ballesta, S.; Gilaberte, Y.; Rezusta, A.; Pascual, Á. Antimicrobial photodynamic activity of hypericin against methicillin-susceptible and resistant Staphylococcus aureus biofilms. Future Microbiol. 2015, 10, 347–356. [Google Scholar] [CrossRef]
- Lavie, G.; Mazur, Y.; Lavie, D.; Prince, A.; Pascual, D.; Liebes, L.; Levin, B.; Meruelo, D. Hypericin as an inactivator of infectious viruses in blood components. Transfusion 1995, 35, 392–400. [Google Scholar] [CrossRef] [PubMed]
- Stevenson, N.R.; Lenard, J. Antiretroviral activities of hypericin and rose bengal: Photodynamic effects on Friend leukemia virus infection of mice. Antivir. Res. 1993, 21, 119–127. [Google Scholar] [CrossRef]
- Zhang, J.N.; Zhang, F.; Tang, Q.J.; Xu, C.S.; Meng, X.H. Effect of photodynamic inactivation of Escherichia coli by hypericin. World J. Microbiol. Biotechnol. 2018, 34, 100. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, J.J.; Rosenson, J. Chemistry of St. John’s Wort: Hypericin and hyperforin. J. Chem. Educ. 2004, 81, 1450. [Google Scholar] [CrossRef]
- Yow, C.M.; Tang, H.M.; Chu, E.S.; Huang, Z. Hypericin-mediated photodynamic antimicrobial effect on clinically isolated pathogens. J. Photochem. Photobiol. 2012, 88, 626–632. [Google Scholar] [CrossRef]
- Jiang, Y.; Leung, A.W.; Wang, X.; Zhang, H.; Xu, C. Inactivation of Staphylococcus aureus by photodynamic action of hypocrellin B. Photodiagn. Photodyn. Ther. 2013, 10, 600–606. [Google Scholar] [CrossRef]
- Guedes, R.C.; Eriksson, L.A. Theoretical study of hypericin. J. Photochem. Photobiol. 2005, 172, 293–299. [Google Scholar] [CrossRef]
- Kashef, N.; Borghei, Y.S.; Djavid, G.E. Photodynamic effect of hypericin on the microorganisms and primary human fibroblasts. Photodiagn. Photodyn. Ther. 2013, 10, 150–155. [Google Scholar] [CrossRef]
- Sperandio, F.F.; Huang, Y.Y.; Hamblin, M.R. Antimicrobial photodynamic therapy to kill Gram-negative bacteria. Recent Pat. Anti Infect. Drug Discov. 2013, 8, 108–120. [Google Scholar] [CrossRef]
- Lopez-Chicon, P.; Paz-Cristobal, M.P.; Rezusta, A.; Aspiroz, C.; Royo-Canas, M.; Andres-Ciriano, E.; Gilaberte, Y.; Agut, M.; Nonell, S. On the mechanism of Candida spp. photoinactivation by hypericin. Photochem. Photobiol. Sci. 2012, 11, 1099–1107. [Google Scholar] [CrossRef]
- Gad, G.F.; El-Domany, R.A.; Ashour, H.M. Antimicrobial susceptibility profile of Pseudomonas aeruginosa isolates in Egypt. J. Urol. 2008, 180, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Silby, M.W.; Winstanley, C.; Godfrey, S.A.; Levy, S.B.; Jackson, R.W. Pseudomonas genomes: Diverse and adaptable. FEMS Microbiol. Rev. 2011, 35, 652–680. [Google Scholar] [CrossRef] [PubMed]
- De Kievit, T. Quorum sensing in Pseudomonas aeruginosa biofilms. Environ. Microbiol. 2009, 11, 279–288. [Google Scholar] [CrossRef] [PubMed]
- Sadikot, R.T.; Blackwell, T.S.; Christman, J.W.; Prince, A.S. Pathogen–host interactions in Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 2005, 171, 1209–1223. [Google Scholar] [CrossRef] [PubMed]
- Gellatly, S.L.; Hancock, R.E. Pseudomonas aeruginosa: New insights into pathogenesis and host defenses. Pathog. Dis. 2013, 67, 159–173. [Google Scholar] [CrossRef] [PubMed]
- Hauser, A.R. The type III secretion system of Pseudomonas aeruginosa: Infection by injection. Nat. Rev. Microbiol. 2009, 7, 654–665. [Google Scholar] [CrossRef]
- Epand, R.M.; Walker, C.; Epand, R.F.; Magarvey, N.A. Molecular mechanisms of membrane targeting antibiotics. Biochim. Biophys. Acta 2016, 1858, 980–987. [Google Scholar] [CrossRef]
- Kapoor, G.; Saigal, S.; Elongavan, A. Action and resistance mechanisms of antibiotics: A guide for clinicians. J. Anaesthesiol. Clin. Pharmacol. 2017, 33, 300–305. [Google Scholar] [CrossRef]
- Hoogkamp-Korstanje, J.A.; Westerdaal, N.A. In vitro Susceptibility of Pseudomonas to Four β-Lactamantibiotics (Ampicillin, Cephalothin, Carbenicillin, Piperacillin), to Four Aminoglycosides (Kanamycin, Amikacin, Gentamicin, Tobramycin) and to Colimycin. Chemotherapy 1979, 25, 48–53. [Google Scholar] [CrossRef]
- Livermore, D.M. Multiple mechanisms of antimicrobial resistance in Pseudomonas aeruginosa: Our worst nightmare? Clin. Infect. Dis. 2002, 34, 634–640. [Google Scholar] [CrossRef]
- Perez-Laguna, V.; Perez-Artiaga, L.; Lampaya-Perez, V.; Garcia-Luque, I.; Ballesta, S.; Nonell, S.; Paz-Cristobal, M.P.; Gilaberte, Y.; Rezusta, A. Bactericidal Effect of Photodynamic Therapy, Alone or in Combination with Mupirocin or Linezolid, on Staphylococcus aureus. Front. Microbiol. 2017, 8, 1002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ewbank, J.J.; Zugasti, O.C. elegans: Model host and tool for antimicrobial drug discovery. Dis. Models Mech. 2011, 4, 300–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, F.; Jain, S.; Oloketuyi, S.F. Bacteria and bacterial products: Foe and friends to Caenorhabditis elegans. Microbiol. Res. 2018, 215, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Sifri, C.D.; Begun, J.; Ausubel, F.M. The worm has turned–microbial virulence modeled in Caenorhabditis elegans. Trends Microbiol. 2005, 13, 119–127. [Google Scholar] [CrossRef]
- Balla, K.M.; Troemel, E.R. Caenorhabditis elegans as a model for intracellular pathogen infection. Cell Microbiol. 2013, 15, 1313–1322. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.Y.; Le, T.A.N.; Lee, S.Y.; Song, D.G.; Hong, S.C.; Cha, K.H.; Lee, J.W.; Pan, C.H.; Kang, K. 3,3′-Diindolylmethane Improves Intestinal Permeability Dysfunction in Cultured Human Intestinal Cells and the Model Animal Caenorhabditis elegans. J. Agric. Food Chem. 2019, 67, 9277–9285. [Google Scholar] [CrossRef]
- Tan, M.W.; Rahme, L.G.; Sternberg, J.A.; Tompkins, R.G.; Ausubel, F.M. Pseudomonas aeruginosa killing of Caenorhabditis elegans used to identify P. aeruginosa virulence factors. Proc. Natl. Acad. Sci. USA 1999, 96, 2408–2413. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Shang, K.; Wu, X.; Song, S.; Li, Z.; Pei, Z.; Pei, Y. Highly efficient green synthesis and photodynamic therapeutic study of hypericin and its derivatives. RSC Adv. 2018, 8, 21786–21792. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.B.; Cha, K.H.; Kim, S.N.; Altantsetseg, S.; Shatar, S.; Sarangerel, O.; Nho, C.W. The antimicrobial activity of essential oil from Dracocephalum foetidum against pathogenic microorganisms. J. Microbiol. 2007, 45, 53–57. [Google Scholar]
- Rubin, J.E.; Ball, K.R.; Chirino-Trejo, M. Antimicrobial susceptibility of Staphylococcus aureus and Staphylococcus pseudintermedius isolated from various animals. Can. Vet. J. 2011, 52, 153–157. [Google Scholar]
- Lee, S.Y.; Kang, K. Measuring the Effect of Chemicals on the Growth and Reproduction of Caenorhabditis elegans. J. Vis. Exp. 2017, e56437. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.Y.; Kim, J.Y.; Jung, Y.J.; Kang, K. Toxicological evaluation of the topoisomerase inhibitor, etoposide, in the model animal Caenorhabditis elegans and 3T3-L1 normal murine cells. Environ. Toxicol. 2017, 32, 1836–1843. [Google Scholar] [CrossRef] [PubMed]
- Rezusta, A.; López-Chicón, P.; Paz-Cristobal, M.P.; Alemany-Ribes, M.; Royo-Díez, D.; Agut, M.; Semino, C.; Nonell, S.; Revillo, M.J.; Aspiroz, C. In vitro fungicidal photodynamic effect of hypericin on Candida species. J. Photochem. Photobiol. 2012, 88, 613–619. [Google Scholar] [CrossRef] [PubMed]
- De Paula, L.F.; Santos, R.O.; Menezes, H.D.; De Britto, J.R.; Vieira, J.B.; Gontijo, P.P.; De Oliveira, C.A. A Comparative Study of Irradiation Systems for Photoinactivation of Microorganisms. J. Braz. Chem. Soc. 2010, 21, 694–700. [Google Scholar] [CrossRef]
- Boyce, J.M.; Pittet, D. Guideline for Hand Hygiene in Health-Care Settings. Recommendations of the Healthcare Infection Control Practices Advisory Committee and the HIPAC/SHEA/APIC/IDSA Hand Hygiene Task Force. Am. J. Infect. Control 2002, 30, S1–S46. [Google Scholar] [CrossRef]
- Kim, H.; Kim, S.W.; Seok, K.H.; Hwang, C.W.; Ahn, J.C.; Jin, J.O.; Kang, H.W. Hypericin-assisted photodynamic therapy against anaplastic thyroid cancer. Photodiagn. Photodyn. Ther. 2018, 24, 15–21. [Google Scholar] [CrossRef]
- Xuan, W.; He, Y.; Huang, L.; Huang, Y.Y.; Bhayana, B.; Xi, L.; Gelfand, J.A.; Hamblin, M.R. Antimicrobial Photodynamic Inactivation Mediated by Tetracyclines in Vitro and in Vivo: Photochemical Mechanisms and Potentiation by Potassium Iodide. Sci. Rep. 2018, 8, 17130. [Google Scholar] [CrossRef] [Green Version]
- Agostinis, P.; Vantieghem, A.; Merlevede, W.; De Witte, P.A. Hypericin in cancer treatment: More light on the way. Int. J. Biochem. Cell Biol. 2002, 34, 221–241. [Google Scholar] [CrossRef]
- Bolze, F.; Jenni, S.; Sour, A.; Heitz, V. Molecular photosensitisers for two-photon photodynamic therapy. Chem. Commun. 2017, 53, 12857–12877. [Google Scholar] [CrossRef]
- Lee, H.H.; Choi, M.G.; Hasan, T. Application of photodynamic therapy in gastrointestinal disorders: An outdated or re-emerging technique? Korean J. Intern. Med. 2017, 32, 1–10. [Google Scholar] [CrossRef]










© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alam, S.T.; Le, T.A.N.; Park, J.-S.; Kwon, H.C.; Kang, K. Antimicrobial Biophotonic Treatment of Ampicillin-Resistant Pseudomonas aeruginosa with Hypericin and Ampicillin Cotreatment Followed by Orange Light. Pharmaceutics 2019, 11, 641. https://doi.org/10.3390/pharmaceutics11120641
Alam ST, Le TAN, Park J-S, Kwon HC, Kang K. Antimicrobial Biophotonic Treatment of Ampicillin-Resistant Pseudomonas aeruginosa with Hypericin and Ampicillin Cotreatment Followed by Orange Light. Pharmaceutics. 2019; 11(12):641. https://doi.org/10.3390/pharmaceutics11120641
Chicago/Turabian StyleAlam, Seemi Tasnim, Tram Anh Ngoc Le, Jin-Soo Park, Hak Cheol Kwon, and Kyungsu Kang. 2019. "Antimicrobial Biophotonic Treatment of Ampicillin-Resistant Pseudomonas aeruginosa with Hypericin and Ampicillin Cotreatment Followed by Orange Light" Pharmaceutics 11, no. 12: 641. https://doi.org/10.3390/pharmaceutics11120641