
ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets

Department of Laboratory Medicine, Division of Clinical Pharmacology, Karolinska Institute and Karolinska University Hospital, C1:68, 141 86 Stockholm, Sweden
Pharmaceuticals 2023, 16(2), 162; https://doi.org/10.3390/ph16020162
Submission received: 27 December 2022
/
Revised: 15 January 2023
/
Accepted: 18 January 2023
/
Published: 22 January 2023
(This article belongs to the Special Issue Calcium-Permeable Ion Channels as Targets in Pharmacotherapy)
Abstract
:The changes in intracellular free calcium (Ca2+) levels are one of the most widely regulators of cell function; therefore, calcium as a universal intracellular mediator is involved in very important human diseases and disorders. In many cells, Ca2+ inflow is mediated by store-operated calcium channels, and it is recognized that the store-operated calcium entry (SOCE) is mediated by the two partners: the pore-forming proteins Orai (Orai1-3) and the calcium store sensor, stromal interaction molecule (STIM1-2). Importantly, the Orai/STIM channels are involved in crucial cell signalling processes such as growth factors, neurotransmitters, and cytokines via interaction with protein tyrosine kinase coupled receptors and G protein-coupled receptors. Therefore, in recent years, the issue of Orai/STIM channels as a drug target in human diseases has received considerable attention. This review summarizes and highlights our current knowledge of the Orai/STIM channels in human diseases and disorders, including immunodeficiency, myopathy, tubular aggregate, Stormorken syndrome, York platelet syndrome, cardiovascular and metabolic disorders, and cancers, as well as suggesting these channels as drug targets for pharmacological therapeutic intervention. Moreover, this work will also focus on the pharmacological modulators of Orai/STIM channel complexes. Together, our thoughtful of the biology and physiology of the Orai/STIM channels have grown remarkably during the past three decades, and the next important milestone in the field of store-operated calcium entry will be to identify potent and selective small molecules as a therapeutic agent with the purpose to target human diseases and disorders for patient benefit.

1. Introduction
The hint of SOCE that describes how Ca2+ inflow across the plasma membrane is triggered by the depletion of endoplasmic reticulum (ER) Ca2+ reserves was first introduced by [1] about three decades ago. Today, the concept of SOCE is well-established and universal in various mammalian cell types. This unique mechanism of Ca2+ entry controls the basal calcium, refills intracellular Ca2+ stores, and performs an extensive variety of specific activities. Therefore, SOCE is involved in very important physiological and pathological processes [2]. These processes comprise gene expression, proliferation, secretion, and metabolism, as well as cell growth. Furthermore, many studies have reported that SOCE is involved in various human diseases, such as allergies, cancer, some types of immunodeficiency and autoimmunity disorders, inflammatory bowel disease, and—recently by our lab—diabetic nephropathy [2,3].
1.1. Membrane Topology Structure of the ORAI/STIM Channel Complexes
Ion channels are homomeric or heteromeric protein subunit assemblies to form ion conduction pathways through cell membranes, which are otherwise very hydrophobic. Moreover, channel dimers, trimers, tetramers, pentamers, and hexamers have all been depicted [4]. Following the discovery of genes encoding Orai family members, scientists began using biochemical, functional, and imaging approaches to conclude the stoichiometric composition of the Orai channel pore. Tandem dimeric, trimeric, or tetrameric Orai1 constructs with or without pore-blocking dominant-negative mutations were co-expressed with STIM1 to investigate Orai channel stoichiometry [5]. In the past, the Orai channel was thought to be tetrameric; however, based on crystallographic and electrophysiological investigations, it is now believed to be a hexamer of Orai subunits (Figure 1), [6,7,8,9].
In 2012, Hou et al. solved the X-ray crystal structure of a closed conformation of Drosophila melanogaster Orai, which provided the first glimpse of the channel’s molecular architecture [7]. The structure was resolved at 3.35 Å resolution and revealed that the channel is made up of a hexamer of Orai subunits instead of a tetrameric assembly, as had been predicted at the time [7]. Moreover, other groups confirmed that human Orai1 channels can likewise assemble and function as hexamers [10,11]. Today, it is well established that the channel is composed of six subunits, as shown in Figure 1, and each subunit comprises four transmembrane helices, TMHs (M1, M2, M3, and M4). Figure 1 illustrates the hexameric Orai channel structure and demonstrates that each subunit contains four transmembrane helices, as represented in different colours. Bioinformatics software Pymol was used to create structures based on known protein sequences and work from Hou et al. [12]. In more detail, the channel contains a single ion conduction pore along a central sixfold axis of symmetry, which in a cellular setting would be vertical to the membrane. Six M1 helices, one from each subunit, constitute the walls of the ion pore, which measures around 55 Å in length and is narrow in the closed conformation [12].
In brief, Yamashita et al. described how the pore opens in response to STIM1 binding [13]. In more detail, this group reported that V102 and F99, which are two rings of the pore-lining residues working as partners to form a hydrophobic gate. Moreover, mutations of residues to polar amino acids create channels with leaky gates operating ions in resting state. Pore helix rotation causes STIM1-mediated channel activation, which moves the F99 residues away from the pore axis, resulting in increased pore hydration. This enables ions to flow through the V102-F99 hydrophobic band. This rotation discloses the dynamic coupling between Ca2+ release-activated Ca2+ (CRAC) channel gating and ion selectivity. This hydrophobic gating mechanism plays a role in CRAC channel function, pharmacology, and disease-causing mutations [13].
Tiffner and colleagues very recently identified that isoform-specific nonconserved gating checkpoints in transmembrane domain 3 (TM3) of Orai1 and Orai3 maintain a closed state and an opening permissive channel conformation, whereas global conformational TM movements are required for pore opening of both channels [14]. That indicates that TM3 controls the pore opening in an isoform-specific manner for both Orai1 and Orai3 proteins [14].
1.2. Activation and Regulation of ORAI/STIM Channel Complexes
Calcium (Ca2+) ion is a universal second messenger that controls wide diversity and very critical functions in almost all cell types. These cellular functions involve muscle contraction, neuronal transmission, cell migration, cell growth, gene transcription, and cell death. Therefore, dysregulation of Ca2+ signals is linked to major diseases in humans such as cardiovascular and neurological diseases, cancer, and others [15]. Ion channels are pore-forming proteins that participate and play crucial roles in very vital physiological and pathological processes such as neuronal signalling and cardiac excitability; as a result, they serve as therapeutic drug targets such as in diabetes mellitus [4,16].
It is well established that the store-operated calcium entry is mediated by the two partners, the pore-forming proteins Orai (Orai1-3) and the calcium store sensor, stromal interaction molecule (STIM1-2). Briefly, in numerous mammal cells, the store-operated (CRAC) channels function as a vital path for Ca2+ entry and defects in CRAC channel utility are linked with severe human diseases such as immunodeficiency and autoimmunity [2].
Various extracellular signals can trigger GPCRs and/or Receptor Tyrosine Kinases (RTKs) that are localized on the cell membrane. This results from Phospholipase C-mediated enzymatic hydrolysis of Phosphatidyl inositol 4,5-bis phosphate (PIP2) and generation of the intracellular messenger inositol-1,4,5-triphosphate (IP3). Inositol-1,4,5-triphosphate receptor (IP3R): IP3 binds to the IP3R, which is largely found in the ER [17]. In brief, the IP3R is a ligand-activated Ca2+ channel. Following a ligand binding to the IP3R, Ca2+ flows from the ER to the cytosol along a favourable concentration gradient. Stromal interaction molecules (STIM) on the ER membrane sense the drop in ER Ca2+ because they are near the ER membrane [18]. STIM proteins operate as sensors for Ca2+ stored in the ER lumen and subsequently translocate into the ER-plasma membrane junctions, where they bind to and activate Orai channels. In other words, to generate the Ca2+-selective, CRAC current, the Orai channels are activated directly by STIM proteins, consequent to ER Ca2+ store depletion [19]. Notably, STIM proteins have a unique N-terminal calcium sensing domain that allows them to respond to changes in the ER luminal calcium concentration. This domain provides STIM proteins the ability to sense calcium levels in the ER [20]. STIM1 directly gates and activates the Orai1 after STIM1 binds to the C- and N- terminal cytosolic domains of Orai1. Notably, CRAC-activation domain, recognized as the STIM-Orai activating region (SOAR), contains ~100 amino acids and is essential for the activation of the channel [21,22].
As an example, the SOCE in VSMCs is mediated by the Orai proteins. Activation of Orai1 channels is induced by Ca2+ release from the ER via the IP3 receptor, which leads in store depletion and the formation of STIM1 oligomers. Somatic cell migration and proliferation (SOCE) mediated by Orai1 are related with the stimulation of the nuclear factor of the activated T-cell (NFAT), which increases the proliferation and migration of VSMCs. Additionally, Ca2+ influx through Orai1 stimulates mitochondrial Ca2+ uptake through the mitochondrial Ca2+ uniporter [23].
The involvement of classical transient receptor potential TRPC1, TRPC4, and TRPC5 (TRPC1/4/5) channels in SOCE has been controversial in the Ca2+ channel field [24]. Remarkably, the Pico145 blocker, which is a potent and selective inhibitor of TRPC1/4/5 channels, failed to produce any noticeable effect on the SOCE that was mediated by Orai1 [25,26,27,28,29]. These findings lead us to believe that the contribution of TRPC1/4/5 channels to store-operated Ca2+ entry should be disregarded in A498, HEK 293, and HUVEC cells [25,26,27,28,29].
2. ORAI/STIM Channel Complexes in Human Diseases
In recent years, the issue of Orai/STIM channels as a drug target in human diseases has received considerable attention. It is well-established that the store-operated calcium entry is mediated by the two partners: the pore-forming proteins Orai (Orai1-3) and the calcium store sensor, stromal interaction molecule (STIM1-2). Briefly, in numerous animal cells, the store-operated Ca2+ release-activated Ca2+ (CRAC) channels function as a vital path for Ca2+ entry, and defects in the CRAC channel event are linked with severe human diseases such as immunodeficiency and autoimmunity [30].
The intention of this section is to review, focus, and discuss the involvement of Orai/STIM channels in different and important human diseases and disorders such as immunodeficiency, myopathy, tubular aggregate, Stormorken syndrome, York platelet syndrome, cancers, and cardiovascular and metabolic disorders.
2.1. Immunodeficiency
Nearly two decades ago, Feske and his colleagues by applying two unbiased genetic methods reported a modified linkage analysis to categorize the gene mutated in the severe combined immune deficiency (SCID) patients, and a genome-wide RNA interference (RNAi) screen identified that a mutation in Orai1, R91W, produces immune deficiency by abolishing CRAC channel function [30]. This mutation denotes a nonfunctional CRAC channels as a result of a defect in gating than in an interaction with full-length STIM1 [31].
In addition, Orai1 is a vital element or mechanism of the CRAC channel complex [32]. Stimulation of immune cells by antigens triggers Ca2+ entry through CRAC channels, contributing to the immune response to pathogens by triggering the NFAT. The same group reported earlier that deficiency in SOCE and CRAC channel function is associated as hereditary to SCID syndrome in humans [32].
Immunodeficient patients with Orai1 deficiency suffer from persistent critical infections with viral bacterial, mycobacterial and fungal pathogens, which leads to pneumonia, meningitis, enteritis, GI candidiasis and sepsis due to a number of mutations, e.g., G98R and A103E, two loss-of-function mutations that abolish SOCE (Table 1) [33,34,35]. In patients lacking functional Orai expression, the immunodeficiency is dominated by a defect in T-cell activation and possibly B- and NK-cell activation, whereas lymphocyte development is normal [33]. The clinical symptoms resemble those found in patients with severe combined immunodeficiency. A majority of all patients with Orai1 deficiency exhibit increased susceptibility to a number of herpes virus infections.
2.2. Myopathy
Recently, as SOCE is the major mechanism that regulates the Ca2+ homeostasis that is facilitated by the two partners STIM1 and Orai1, Böhm et al. reported that Orai1 mutations, which have distinct channel gating defects, are associated beside Stormorken Syndrome and tubular aggregate myopathy (TAM) [36]. Böhm et al. suggested in their study on Orai1 channels that TAM and Stormorken syndrome are varieties of a similar disease, as their data stipulated a mutation-dependent pathomechanism and a genotype/phenotype correlation as the Orai1 mutations, which are related to the furthermost severe indications, producing the most significant functional influence on the cells (Table 1) [37]. The two syndromes are involved in a spectrum of clinical manifestations, including muscle weakness, skin disorder ichthyhosis, eye disorder miosis, blood disorder thrombocytopenia, and learning disorder dyslexia.
2.3. Tubular Aggregate Myopathy
Tubular aggregate myopathy (TAM) is a rare form of myopathy that affects the skeletal muscles. Muscle biopsies taken from TAM patients show densely packed membrane tubules, and symptoms include weakness, cramps, and muscle pains. TAM commonly affects the proximal muscles of the lower limbs, causing loss of mobility, including difficulties running and climbing stairs (Table 1) [36].
TAM and Stormorken syndrome are both caused by gain-of-function mutations. This type of mutation results from an alteration of gene products in which a new molecular function or new gene expression pattern are formed. The gain-of-function mutations in STIM1 or Orai1 and additional mutations in CASQ1 cause excessive Ca2+, leading to the two syndromes. These three genes are involved in the control of Ca2+ homeostasis [36]. All STIM1 mutations that are involved in TAM have shown to be missense mutations located in the EF hand, helix–loop–helix structural domain. It is suggested that these mutations prompt the constitutive ORAI1 activation and generate excessive Ca2+ entry into muscle cells [38]. Mutation STIM1-I484R has been located not in the EF hand but in STIM1′s cytoplasmic C-terminal inhibitory domain (CTID). It has been reported that cells transfected with STIM-I484R showed an aggregation-like appearance of STIM1 and reduced Ca2+ influx [38]. Further on, it was determined that frameshift STIM-I484R mutation induces a gain in CRAC channel function due to the loss of critical inhibitory C-terminal domains. This blocks STIM1 to bind to ORAI1, which then enables STIM1 trapping by microtubules and interferes slow Ca2+-dependent inhibition. This might provide a mechanistic explanation of why TAM patients with this mutation receive muscular defects [39].
ORAI-G98S, -V107M, and T184M are three other mutations of ORAI1 that are all involved in TAM. G98S and V107M mutations cause constitutive channel opening independently of STIM, while the third mutation, T184M, which is not located in the channel pore, overacts depending on STIM presence. This suggests that these mutations located in different ORAI1 transmembrane domains seem to involve different pathomechanisms revealing a genotype–phenotype correlation [37].
ORAI1-G98S is a heterozygous missense mutation that causes TAM with hypocalcemia due to constitutive activation of CRAC channels. A study involving ORAI1-G98S and other constitutive activity ORAI1 mutations—V107M, L138F, T184M, and P245L—showed when ORAI blocker GSK-7975A was applied the G98S mutant was resistant to inhibition. This is while GSK-7975A blocked the constitutive activity of the other mutants, indicating that it could possibly be used in therapy for TAM patients [40,41].
STIM mutations also play a role in TAM, e.g., EF hand mutation STIM1-D84G, which causes constitutive Ca2+ influx present in patients with nonsyndromic TAM. Interestingly, a study involving mice with STIM1-D84G developed a bleeding disorder with thrombocytopenia, a condition when blood platelet count is too low. Moreover, mutation STIM-H109N causes somewhat different disease symptoms, such as either predominantly postexercise fatigability and episodic double vision or weakness in lower limb muscles and contractures [42].
2.4. Stormorken Syndrome
Tubular aggregate myopathy is also present in a rare multiorgan disease, Stormorken syndrome, and this has led to discussions about considering the two as part of one clinical continuum. Stormorken syndrome is caused by autosomal dominant mutations, and its clinical features include miosis (permanent constriction of the eye pupils), thrombocytopenia (low blood platelet count), ichthyosis (widespread and persistent thick, dry, “fish-scale” skin), hyposplenism (physiological loss of spleen function), dyslexia, and short stature [43].
Stormorken syndrome is clinically fully expressed in patients who are affected by mutations of STIM and/or ORAI. STIM1-R304W mutation in functional studies showed a possible mutational influence on Orai1 channel deactivation; however, the specific pathomechanism of the syndrome is yet to be identified (Table 1) [44]. R304W in STIM1 activates allele more strongly than mutation P245L in ORAI1, which prevents slow Ca2+-dependent inactivation, however, does not cause a constitutively active CRAC channel. ORAI1-P245L is located with the fourth transmembrane helix, M4, of ORAI1, and it acts as a gain-of-function mutation [44].
Currently, there is no treatment for TAM and Stormorken syndrome; however, it has been shown that Ca2+ balance can be manipulated by chemical compounds. It has been indicated that reducing excessive Ca2+ influx through inhibition of CRAC channels could be a possible therapeutic approach in the treatment. Further studies are needed to determine if an imbalance of other ions could be a factor to the development of Stormorken syndrome and TAM [36].
2.5. York Platelet Syndrome
Within the same clinical continuum as TAM and Stormorken syndrome is York platelet syndrome (YPS). Clinical studies have shown that its muscular and nonmuscular phenotypes are consistent with the two other syndromes [45]. York platelet syndrome is a Ca2+ channelopathy caused by gain-of-function in STIM1 through heterozygous mutations, including c.343A > T and c.910C > T [45]. YPS represents a rare genetic form of the blood disorder thrombocytopenia, with striking ultrastructural platelet abnormalities such as giant electron opaque organelles and massive, multilayered target bodies, as well as a deficiency of platelet Ca2+ storage in delta granules.
A majority of patients suffering from YPS have a greater tendency to bleeding with epistaxis (nose bleeding), bruising, and postpartum haemorrhage. Notably, YPS and Stormorken syndrome are associated with same types of symptoms, and some suggest that the two may be the same clinical condition but with different names (Table 1) [46].
2.6. Cardiovascular and Metabolic Disorders
Calcium ions’ involvement in cardiovascular diseases is well known and was first introduced by Putney [1]. A number of studies have shown that vascular illnesses are significantly influenced by the dysregulation of the SOCE pathway. In most vascular disorders, there is an increased presence of STIM1, Orai1, and Orai3, which results in dysregulation of cellular Ca2+ homeostasis, which in turn leads to vascular pathologies. It has been discovered that STIM and Orai proteins are key molecular players of the SOCE pathway [47]. Because of these breakthroughs, the roles of STIM1 and Orai1 in mediating the increase in Ca2+ platelet concentration and thrombosis are now better understood.
Grosse et al. and Ahmad et al. have suggested that STIM1 is involved in thrombus formation and its stability. In a separate investigation, Xia et al. stated that an increased STIM1 function in patients with type 2 diabetic patients could be a contributing factor to their high tendency to thrombosis [48,49,50,51].
STIM1 and Orai1 also play a role in atherosclerosis, with studies showing that an increased presence SOCE in VSMCs prior to development of atherosclerosis plaques [52]. According to many studies, high-fat diets increase SOCE and STIM1 expression in coronary smooth cells, which is associated with enhanced atherosclerosis lesions [47]. It has also been shown that STIM1 is linked to the regulation of the endothelial barrier function and progenitor endothelial cells functions in atherosclerotic lesions [53]. STIM1 plays a key role in GPCR-activated endothelial barrier function alterations independent of Ca2+ and Orai1 [53]. A separate study has reported the significant role of Orai1 in atherosclerosis by its regulation of both vascular inflammation and foam cell formation [54]. In conclusion, these studies propose that STIM1 and Orai1 are involved in atherosclerosis plaque development. STIM and Orai proteins also play a critical role in pulmonary and vascular hypertension, as studies show a very strong association between hypertension and the augmented presence of the two proteins [55].
2.7. Cancers
Growing evidence has shown that numerous cancer types, including liver, lung, ovarian, breast, colon, and gastric cancer, as well as multiple myeloma, manifest increased SOCE and augmented expression of STIM1 or Orai1. Additionally, it has been claimed that SOCE could play a role in the oncogenic pathway, since its blockage, which can be achieved through the knockdown of STIM1 or Orai1, prevents the proliferation and spread of cancer cells [56,57,58,59,60,61]. It is suggested that SOCE stimulates the proliferation and metastasis of cancer cells, enhances tumour angiogenesis and the formation of a tumour-promoting inflammatory environment [58]. A study by Chen et al. demonstrated that STIM1 is able to control the production of vascular endothelial growth factor (VEGF) in cervical cancer by inhibiting STIM1, and the ability to govern the growth of blood vessels and tumours could be impaired [62]. Upregulation of STIM1 in primary cervical tumours was strongly linked with larger tumour size and elevated lymph node metastasis, and therefore poorer clinical outcomes [62]. A study involving immunostaining of overexpressed STIM1 and STIM2 in human cervical cancer revealed that STIM1 is the main endoplasmic reticulum Ca2+-sensing molecule found in the invasive tumour front, indicating that STIM1 plays a role in both tumour growth and invasion. This is while STIM2 is mainly associated with tumour growth (Table 1) [63]. In multiple myeloma patients, overexpression of STIM1/Orai1 was closely linked with shorter progression-free survival (Table 1) [60]. This was also observed in individuals with oesophageal squamous cell carcinoma, as increased expression of Orai1 in tumour tissue was associated with a worse prognosis overall and recurrence-free survival [64].
Schild et al. developed novel effective inhibitors: several monosubstituted and amino acid analogues of 2-APB and SOCE using a fluorometric imaging plate reader (FLIPR) assay format in MDA-MB-231 cells [65]. Compared to 2-APB, the compound series of monohalogenated 2-APB derivatives demonstrated potential inhibitory efficacy. Analogues m-Cl-2APB, m-Br-2APB, m-I-2APB, and p-I-2APB blocked SOCE at five times lower concentrations than 2-APB, and notably, 2-APB increased SOCE at this low concentration [65].
Recently, more and more evidence is indicating that Orai1′s two homologs, Orai2 and Orai3, play a role in the development of different types of cancer such as lung, breast, prostate, and leukaemia [66]. Studies have shown that Orai3 plays an essential role in lung cancer, with one study revealing that immunostaining of Orai3 was overexpressed in lung cancer tissues compared to nontumorous ones, and was linked to a high tumour grade [67]. In a separate study, Orai3 immunostaining showed a correlation between overexpressed Orai3 and the aggressiveness of lung adenocarcinoma [68].
The most commonly occurring cancer among women is breast cancer. In cancer cells, SOCE channels are extensively remodelled and are involved in the development and support of different cancer hallmarks. In all investigated breast cancer cell subtypes, Orai1 is overexpressed, and its cell biology is strongly dependent on CRAC channels. It has also been shown that Orai3 is selectively overexpressed in luminal A breast cancer cells where both SOCE and the development of different cancer features such as cell migration and proliferation are greatly dependent on Orai3. The role of Orai2 in breast cells was not determined until recently, when Sanchez-Collado et al. suggested that Orai2 is needed for cell cycle progression and apoptosis resistance in cells with high Orai2 expression [66]. The group indicated that based on their findings, Orai2 plays a crucial role in the pathophysiology of breast cancer subtypes with an elevated Orai2 expression profile [66]. Furthermore, Sanchez-Collado et al. suggested that Orai2′s role in cell cycle progression could be one factor contributing to chemoresistance in breast cancer cells [66].
Prostate cancer is the third most lethal tumour among men, with more than 500,000 new cases every year. Orai1, 2, and 3 are all expressed in normal and cancerous epithelial prostatic cells. It has been determined that in prostate cancer, remodelling of channel-forming Orai proteins induces an oncogenic switch [69]. It has also been shown that in prostate cancer samples, the relative expression of the Orai3 transcript is highly elevated in comparison to paired-match noncancerous tissue [66].
With the determination of the roles of Orai and STIM in cancer, studies involving the pharmacological inhibition of SOCE or downregulation of STIM1 have been revealed to enhance apoptosis induced by cisplatin in non-small cell lung cancer cells, and Orai1/STIM1 are associated with a protective antiapoptotic role in ovary carcinoma cells [70]. In cancer treatment, there is a great potential in inhibition of SOCE; however, its usage should be with caution due to the abundant expression of STIM and Orai proteins in the human body and their vital roles in the immune system. Therefore, one focus should be on developing SOCE modulators that target specific tumour cells or tumour-associated vascular endothelial cells by designing assays in the presence of both cancer and immune cells. Another focus should be to develop therapeutic approaches that target cancer cell-specific mechanisms of SOCE activation [71].
2.8. Nervous System
Orai and STIM proteins are present in normal brain function; however, they are also involved in neurological disorders such as Alzheimer’s disease, epilepsy, brain trauma, and cerebral infarction. The role of Orai proteins in neuronal calcium signalling is yet to be determined.
It has, however, been reported that STIM2 is abundantly active in hippocampal and cortical neurons, which activates neuronal SOCE. It is involved in regulating steady-state activity of phosphorylated Ca2+/calmodulin-dependent protein kinase II (CaMKII) and consequently stabilizes mushroom spines in hippocampal neurons. It is suggested that weakened STIM activity and loss of mushroom spines may play a role in memory deficiency and in Alzheimer’s disease. Regarding epilepsy, brain trauma, and cerebral infarction, STIM2 could possibly be a potential target for the treatment of glutamate-induced excitotoxicity, which leads to loss of neuronal function and cell death [72].
2.9. Kidney
Orai channels are also found in human kidney’s proximal tubules. Dysfunction of these proteins in the proximal tubules causes an absorption decrease in albumin by its cells and leads to proteinuria [2].
Furthermore, store-operated Ca2+ channel Orai1 activity plays a strong role in lymphocytes, especially TH17 cells, which regulate the severity of both acute injury and chronic kidney disease (Table 1) [73]. A study conducted by Basile revealed that inhibition Orai1 diminishes both acute and chronic kidney injury in rodent models [73]. Orai1+ cells are overexpressed in peripheral blood of patients suffering acute kidney injury, suggesting that Orai1 could possibly be a target in the acute kidney injury to chronic kidney disease transition. It is yet to be determined if inhibition of the endothelial STIM1/Orai1 pathway prevents kidney injury and where such activity plays a role in inflammation and/or haemodynamics [73].
In a separate study, Mai et al. revealed that the Orai1 calcium channel plays a role in renal fibrosis induced by a high-fat diet and by unilateral ureteral obstruction [74]. According to their results, Orai1-mediated Ca2+ entry is involved in the progression of renal fibrosis, which might lead to end-stage renal disease; however, the molecular mechanisms remain undetermined. Mai et al. suggest that blocking the Orai1 Ca2+ channel could possibly be a therapeutic option to curb the progress of kidney fibrosis [74].
3. Pharmacology of Store-Operated Calcium Entry Channels
Recent developments in the function of Orai/STIM channels in human physiology and pathology have led to a renewed interest in the identification of small molecules as modulators or pharmacological agents to target these channels. However, major problems in the field of the Orai/STIM channel pharmacology have been the identification of selective subtype-specific and potent modulators of these channels.
3.1. Activators of ORAI/STIM Channel Complexes
The activation process of Orai/STIM channels is very complex; therefore, several pharmacological tools can be developed to modulate these channels’ activity [76]. The Orai/STIM channels can be activated by phytohaemagglutinin (PHA), IP3, cyclopiazonic acid (CPA), ionomycin, BAPTA, EGTA, and thapsigargin (TG) [24,77]. In a nutshell, these substances induces ER Ca2+ release through IP3R, which blocks SERCA to make ER Ca2+ depletion, and which induces a passive ER Ca2+ depletion [78]. Moreover, conventionally nonspecific and low-potency blockers such as 2-APB, La3+, and SKF96365 have regularly been used to study the physiological function of Orai/STIM channels in different cells and tissues [79]. However, selective and potent pharmacological tools that can be used to assess the physiological and pathological role of Orai/STIM channels or develop as therapeutics drugs have been absent (Figure 2). Therefore, the inspiration to discover a novel selective and potent antagonist or agonist has been a necessity in recent years.
3.2. Thapsigargin
Thapsigargin, often known as TG, was first extracted from the roots of the Mediterranean umbelliferous plant Thapsia garganica in order to investigate the nature of its skin-irritant component. The Thapsia L. species, also known as deadly carrots, have been utilized in traditional medicine for thousands of years in the Mediterranean region. Furthermore, TG treatment of mammalian cells was demonstrated to result in increased calcium levels in the cytoplasm, and TG was established as blocker of the sarcoendoplasmic reticulum Ca2+-ATPase (SERCA) in 1990 [80]. TG has a unique chemical structure; in addition, the primary and most significant pharmacological effect of TG is its strong SERCA pump inhibition. Moreover, this effect is substantial at concentrations below one nanomolar [80,81].
3.3. 2T and 3G
Riva and colleagues, based on the Pyr family of chemicals (the pyrtriazole series), developed anti-inflammatory SOCE inhibitors [82]. Aside from that, there were two compounds in this series that were surprisingly discovered to be SOCE activators, activating the channel at 198–236% and 142–197% entry at 10 µM in three distinct cell lines (Hek, Jurkat, and BV-2), respectively. Furthermore, the modest differences identified between the two compounds were due to the fact that 2T was more potent than the other. The impact of 2T was obvious at concentrations more than 0.3 µM, while the effect of 3G was visible at concentrations greater than 3 µM, with the effect of 3G increasing up to the maximum concentration (100 µM) tested [82].
3.4. IA65
IA65 or 4-((5-phenyl-4-(trifluoromethyl)-thiazol-2-yl)amino)benzoic acid has shown in recent studies to increase Orai1 activity in heterologous expression, as well as in vascular smooth muscle cells and skeletal muscle fibres [83]. In a concentration-dependent manner, IA65 also enhanced Ca2+-dependent inactivation (CDI) with an EC50 of 2 µM. IA65 has shown some selectivity by enhancing activity of ORAI1, while only slightly affecting Orai2 and 3. It is suggested that this Orai1 activator could be used as a tool to further understand the involvement of Orai1 in pathosphyisology [83]. Notably, IA65 has the reverse effect of Orai1 in that it suppresses Orai2 channel activation [72].
3.5. Inhibitors of ORAI/STIM Channel Complexes
SOCE is the predominant Ca2+ entry mechanism in mammalian cells and governs a number of cellular activities, including apoptosis, proliferation, motility, and death; therefore, inhibition of SOCE serves as potential drug target in several diseases and disorders [84].
3.6. Pyrazole Compounds: GSK-7975A and GSK-5503A
GSK-5503A and GSK-7975A are pyrazole derivatives that act as selective small molecule blockers of CRAC channels and TRPV6 channels (Table 2). GSK-7975A developed by GlaxoSmithKline inhibits ICRAC channels in HEK293 cells and both Orai1 and Orai3 mediated ICRAC in HEK293 cells with an IC50 of 4 μM (Table 2), respectively [85]. Studies have shown that the GSK compounds also potently block TRPV6 channels, as there are structural associations between CRAC and TRPV6 channels in the target site for the GSK compounds. This is while they have little or no influence on a number of other ion channels [86].
A separate study by Zhang et al. showed that GSK7975A largely inhibits Orai1 and Orai2 while it has a lower inhibitory effect against Orai3 [72]. As GSK7975A is unable to discriminate between Orai1 and Orai3 channels, as well as lacking the ability to block L-type Ca2+ and TRPV6 channels, the compound has a limited specificity as an Orai1 channel blocker [23].
GSK-7975A affects neither STIM1 oligomerization nor STIM1/Orai1 interaction [85]. These two pyrazole compounds could block STIM1-mediated Orai1 and Orai3 currents hypothetically via an allosteric effect on the Orai1 selectivity filter, having a gradual onset of action that effects STIM1-STIM1 oligomerization or STIM1-Orai1 coupling [85].
3.7. Compound 5J-4
Recently, Kim et al. noticed a novel family of compounds that block the activity of CRAC channels by using high-throughput chemical library screening targeting Orai1 [87]. The 5J-4 blocks the CRAC channel activity by preventing ion permeation, and this compound was more tolerated in vivo due to the removal of trichloride (-Cl3) [87]. The 5J-4 strongly blocked the SOCE in HeLa-O+S cells in addition to endogenous SOCE in TH17 cells [87]. The parent compound 5D had a half-maximum inhibitory concentration (IC50) of 807 nM and 195 nM (Table 2) for the peak and sustained levels of endogenous SOCE in primary murine effector T cells [87].
3.8. 2-APB and Its Analogues DPB162-AE and DPB163-AE
2-Aminoethyldiphenyl borate (2-APB) is nonselective Ca2+ entry blocker that has a biphasic effect on CRAC channel activation (Table 2). 2-APB at higher concentrations (20–100 μM, see Table 2) causes a transient activation of the CRAC channel, followed by a very potent inhibitory effect, which was observed even at low concentrations (1–10 μM), potentiating the CRAC channel activity [24,88]. The suggested mechanism of action for 2-APB is possibly through STIM1 multimerization, STIM1– Orai1 interaction, or on the Orai channel itself [31,89,90].
There have been studies on the mechanisms underlying the dual effects of 2-APB on SOCE, of which it has been reported that 2-APB might directly block STIM1 by enabling the connection between CC1 and SOAR. This is while it has been shown that 2-APB can also play a role in indirectly disrupting STIM1 functions by interfering the interaction between STIM1 and the mutant Orai1-V102C [91]. A number of research groups have also reported that 2-APB might directly gate and enlarge the pore diameter of Orai1 and Orai3 to control the SOCE pathway without interfering with STIM1 [2,92].
A number of studies have shown antineoplastic effects of 2-APB, making it a possible potent therapy for primary and metastatic cancers. Among others, when applied in high concentrations, 2-APB works as a blocker of SOCE, showing anticancer effects on various cancer types, including breast, ovarian, cervical, prostate, lung, melanoma, and glioblastoma [93,94,95,96,97].
Other studies have shown that 2-APB is effective in models of atherosclerosis, hypertension, and vascular calcification; however the effects are mostly attributed to the compound’s effects on TRP channels or IP3 receptor [23]. More studies are needed on the effects of 2-APB and its analogues on SOCE, as the mechanism of action is not fully understood.
The two 2-APB analogues, DPB162-AE and DPB163-AE, have quite similar chemical structures and only differ in the linker chain between their two diphenyl groups [90]. These two SOCE inhibitors are 100 times more effective than 2-APB, and they have the ability to block STIM1 clustering and inhibit the STIM1-triggered Orai1 and Orai2 activity by deactivating the STIM1-Orai activating region domain in STIM1.
DPB162-AE and DPB163-AE, like 2-APB, block the Orai1 currents and weaken the Orai2 currents. Nevertheless, in contrast to 2-APB, both fail to activate the Orai3 channels at higher concentrations in the absence of STIM1 [98]. However, the two mutants could both facilitate Orai3 currents at low concentration (~100 nM), and at high concentration (>300 nM) they transiently activated and then deactivated Orai3 currents. The DPB162-AE blocks the CRAC channel activity with IC50 = 200 nM and possibly acts directly on the coupling interface between the STIM1–Orai1 activating region (SOAR) and Orai1 [99,100]. Furthermore, studies have shown that DPB-162 could constantly block endogenous SOCE regardless of concentration strength, and when applying the greatest inhibitory effect on Ca2+ entry, little effect was applied on L-type Ca2+ channels, TRPC channels, or Ca2+ pumps [98,99].
To date, no studies have been reported on DPB compounds in regard to the specific SOCE inhibition and its effects on cancer treatment; however, DPB compounds are seen as potential anticancer drugs in the future.
3.9. Aspirin Metabolite Salicylate and Other NSAIDs
Nonsteroidal anti-inflammatory drugs (NSAIDs) such as aspirin and diclofenac are usually administered to relieve pain, fever, and inflammation; however, investigations have shown that this drug group likewise has cancer-protective effects, reducing the risk of a number of cancer types. A study by Núñez et al. showed that in colon cancer, NSAIDs could exert antiproliferative effects by inhibiting SOCE [101]. Burn et al. reported that patients suffering from Lynch syndrome are at high risk of developing CRC, and their study showed that aspirin protected against CRC [102]. Furthermore, the primary aspirin metabolite, salicylate, is regarded as a slight mitochondrial uncoupler that inhibits the proliferation of HT29 cells. A separate study has shown that applying ibuprofen together with indomethacin blocks COX-2, hindering STIM1-induced colorectal cancer development [103]. Muñoz et al. reported that NSAID drugs suppress vascular smooth muscle cell proliferation by facilitating the Ca2+-dependent inactivation of Orai channels, which mitochondria normally prevent [104]. Moreover, Muñoz et al. concluded that NSAIDs block Orai channels indirectly by preventing mitochondrial Ca2+ uptake, which facilitates the Ca2+-dependent inactivation of Orai channels [104].
3.10. SKF-96365
SKF-96365 is an antineoplastic drug, also known as a chemotherapy drug, and its effects are universal. This SOCE channel blocker is reportedly relatively nonselective, meaning it inhibits many other ion channels with similar potency. This compound is structurally different from other classic Ca2+ antagonists, which inhibits the entrance of Ca2+ by blocking its channels or affects its pools. The compound demonstrates selectivity as it inhibits receptor-mediated Ca2+ entry, causing a significant increase in [Ca2+]i while not affecting internal Ca2+ release in endothelial cells, platelets, or neutrophils [105].
Several studies have shown that SKF96365 inhibits cell proliferation by inducing cell apoptosis and cell cycle arrest in the G2/M phase in various types of cancers [60,72,100,106,107,108,109]. Furthermore, studies have shown that SKF 96354 could play a role in effectively blocking the metastasis of cancers by impairing the assembly and disassembly of focal adhesions of breast cancer cells, and block cell migration by deactivating nonmuscle myosin II, as well as decreasing actomyosin development and contractile force in cervical and non-small cell lung cancer (NSCLC) cells [100,110]. It has also been reported that by blocking TRPC channels in glioma cells, SKF 96365 could also be used as an adjuvant drug for radiotherapy. Analogues of SKF-96365 have been shown to have higher potency in blocking SOCE in B lymphocyte cells; however, further studies are needed to determine their specificity [111]. Azimi et al. found that SKF-96365 was active against Orai1-mediated Ca2+ influx; however, it was not selective with respect to inhibition of the CaV2.2 channel using MDA-MB-231 breast cancer cells [83].
Despite the universal usage of SKF 96365 amid its antineoplastic effects, they are nonspecific, and further studies are needed to determine its specific mechanisms.
3.11. Mibefradil
Mibefradil is a T-type Ca2+ channel inhibitor was first used as a cardiovascular drug. However, lately it has been shown that Mibefradil blocks SOCE by inhibiting Orai channels in a dose-dependent and reversible manner, causing a significant inhibition of cell proliferation, inciting cell apoptosis and halting the cell cycle in the S and G2/M phases in HEK-293T-Rex cells [112].
3.12. 4-Choloro-3-ethylphenol (4-CEP) and Its Analogues 4-Choloro-m-cresol (4-CmC) and 4-Chlorophenol (4-CIP)
4-Choloro-3-ethylphenol (4-CEP) and its analogues 4-choloro-m-cresol (4-CmC) and 4-chlorophenol (4-CIP) all belong to a group of ryanodine receptor (RyR) agonists. The function of RyRs is to release the Ca2+ from intracellular store during excitation–contraction coupling in cardiac and skeletal muscle. A study by Zeng et al. has shown that 4-CEP blocked SOCE in rat L6 myoblasts and induced a significant Ca2+ release, while its derivatives 4-CmC and 4-CIP at the same concentration only showed a small increase and no effect, respectively [113]. The study also showed that 4-CEP can evoke Ca2+ release through RyR activation, but significantly blocks the store-operated Ca2+ influx in thapsigargin-treated, store-depleted STIM1/Orai1 cells. The results showed IC50 values for 4-CEP, 4-CmC, and 4-CIP of 203.6, 830.9, and 1437.1 µM, respectively (Table 2). Furthermore, the study suggested that 500 µM of 4-CEP is inhibited in HEK293 T-REx cells and three store-operated Orai currents are evoked by thapsigargin, namely Orai1/STIM1, Orai2/STIM1, and Orai3/STIM1, while the Orai currents are partially blocked by 4-CmC and 4-CIP using the same concentration. Testing the effects of these three RyR agonists on 2-APB-induced Orai3 currents using 500 µM of 4-CEP, 4-CmC and 4-CIP, the current was significantly blocked with potencies in the order of 4-CEP > 4-CmC > 4-CIP. Whole-cell recordings of Orai3 showed that the current was induced by 50 µM of 2-APB and was blocked by 4-CEP with an IC50 value of 71.0 ± 4.2 µM (Table 2). This IC50 value is lower compared to other non-selective store-operated channel blockers, including 2-APB, SKF96365, and BTP2 [106,114,115]. Zeng et al. also suggested the action site for 4-CEP, as intracellular application of 4-CEP did not block the Orai3 channels, although the extracellular perfusion blocked the current [114].
3.13. Lanthanides
At low concentrations, lanthanides such as lanthanum and gadolinium (Gd3+) block CRAC channels and micromolar Gd3+ inhibits Ca2+ influx, which is often seen as evidence for store-operated Ca2+ entry. Lathanides also play a role in many other ion channels, meaning that Gd3+ even at submicromolar concentrations are not considered a specific CRAC channel inhibitor [116]. Lathanides can inhibit SOCE at low concentrations (<1 μM). Studies on Orai channels have shown that lathanides act by access of Ca2+ ions to the selectivity filter and pore [117]. While at concentrations of more than 100 μM, lathanides begin to inhibit plasma membrane Ca2+ATPase (PMCA), and at concentrations above 1 mM they completely block the activity, making it seem as if the cytoplasm is isolated or “insulated” from the extracellular space. At this state, the “insulation” is stopping both the entry and efflux of Ca2+ ions, creating an opportunity for investigations of complex intracellular calcium signalling events unassisted by constituents in the extracellular space [77].
3.14. Bistrifluoromethyl-Pyrazole Derivative (BTP-2) or Pyrazole Derivatives
Bistrifluoromethyl-pyrazole derivative, known as Pyr2, BTP2, or YM-58483, was initially reported as an SOCE inhibitor, reducing IL-2 production and NFAT dephosphorylation in Jurkat cells without altering the T-cell receptor signal transduction pathway [115]. BTP2 reportedly inhibits cytokine production and proliferation in electrically non-excitable T cells [118]. Studies have shown that YM-58483 does not cross-react with other ion channels, including voltage-operated Ca2+ entry, chloride (Cl−) channels, or potassium (K+) channels [115,119,120]. The exact molecular mechanism of this compound is unknown; nevertheless, YM-58483/BTP2 suppresses a number of ion channels such as CRAC, TRPC3, and TRPC5, as well as facilitating the TRPM4 channel [115,119,120]. YM-58483 is also a potent blocker of thapsigargin-induced SOCE Ca2+ influx. In a study by Azimi et al. using MDA-MB-231 breast cancer cells, YM-58483 inhibited SOCE mediated by Orai1 with an IC50 value of 2.8 µM, while the compound was inactive against TRPV1, TRPM8, and CaV2.2 at concentrations up to 100 µM (Table 2) [121].
He et al. studied 3,5-bis(trifluoromethyl)pyrazole derivative BTP2 and its effect on SOCE activation in various cell types and assessed its modification of TRPC3, TRPC5, and TRPV6 with an aim to find any link between the function of the TRP channel and SOCE [119]. Using human embryonic kidney (HEK) 293 cells, chicken DT40 B cell line, and A7r5 embryonic rat VSMCs, the group found that BTP2 inhibited SOCE within 10 min with an IC50 value of 0.1–0.3 µM. BTP also blocked the T3-65 clonal HEK293 cell line, stably expressing TRPC3 channels and TRPC3-mediated Sr2+-entry activated by muscarinic receptors with an IC50 value of <0.3 µM. The study also showed that BTP2 inhibited direct activation of TRPC3 channels by diacylglycerol, with an IC50 value of about 0.3 µM. BTP2 also blocked the whole-cell carbachol-induced TRPC3 current by 3 µM. Recordings of the single TRPC3 channel showed persistent short openings, indicating that BTP2 diminishes the channel’s open probability rather than its pore properties. In the case of TRPC5 channels transiently expressed in HEK293 cells, BTP2 blocked in the same range as TRPC3. This is while BTP2 did not affect the function of the highly Ca2+-selective TRPV6 channel at all, despite having as many channel properties as SOCs. He et al. concluded with these results that there is a strong functional relation between the operation of the expressed TRPC channel and endogenous SOC activity [119].
3.15. Synta-66
Synta-66 (N-(2′,5′-dimethoxy-[1,1′-biphenyl]-4-yl)-3-fluoroisonicotinamide) is a selective CRAC channel inhibitor developed by Synta Pharmaceuticals (Table 1). Synta-66 inhibits ICRAC in rat basophilic leukaemia (RBL) cells with an IC50 value of 1.4 μM [122], while not affecting plasma membrane Ca2+ ATPase pump and inwardly rectifying K+ channels.
A study by Li et al. showed that Synta-66 is a potent SOCE inhibitor in VSMCs, which are isolated from human saphenous veins [123]. The results showed a significant reduction in VSMC migration, with an IC50 of around 26 nM [123]. Despite various studies of Synta-66, the mechanism of action for the compound has not yet been fully clarified. Furthermore, due its poor aqueous solubility, Synta-66 probably will not be suitable for therapies [23]. It is worth noting that Synta66 inhibited Orai1 channel activity while potentiating Orai2, and had no impact on Orai3 [72].
3.16. RO2959
The CRAC inhibitor RO2959 (Carboxyamidotriazole; 2,6-Difluoro-N-tetrahydro-pyridin-3-yl]-pyrazin-2-yl)-benzamide) was characterized by Chen G. et al., and potently blocks human T-cell activation and effector functions (Table 2) [124]. In more detail, by using both electrophysiological and calcium-based fluorescence measurements, this group showed that the RO2959 was able to block the IP3-dependent current in CRAC-expressing RBL-2H3 cells and CHO cells stably expressing human Orai1 and Stim1, as well as SOCE in human primary CD4(+) T cells triggered by either TCR stimulation or thapsigargin treatment [124]. Originally, RO2959 was synthesized by Synta Pharmaceutical and has been identified as a novel, potent, and selective ICRAC inhibitor by Roche [90]. The ICRAC inhibitor is efficient at concentrations of nanomolars and demonstrates superior selectivity for Orai1 inhibition compared to Orai2 and Orai3. In a study by Chen et al., RO2959 at a 3 µM (Table 2) concentration inhibited the 5-hydroxytryptamine receptor 2B (5-HT2B) by 87% and the peripheral benzodiazepine (BZD) receptors by 89%. Despite this progress, the mechanism of action of RO2959 has yet to be determined [124].
3.17. CM4620, CM2489, CM3457, CM128 (CalciMedica Series)
CM4620 is a small molecule Orai1 inhibitor with a IC50 = 119 nM (Table 2) and displaying a weaker degree of effectiveness when used against Orai2 at 895 nM [125]. CM2489 and CM3457 are small molecule inhibitors of CRAC channels developed by CalciMedica (Table 2). The structure of CM2489 has not been revealed yet; however, it is the first CRAC channel blocker to be tested in humans for treating moderate-to-severe plaque psoriasis, and has completed Phase I clinical trials, although it was discontinued. This is while the efficacy of CM2489 as an inhibitor of different Orai isoforms and its mechanism of action have not been identified yet. The structure and mechanism of CM3457 remains unclear; although it has been shown that the compound inhibits T-cell proliferation, Th1, Th2, and Th17 induced cytokine production and mast cell degranulation in vitro. Studies on rats have revealed that orally administered CM3457 was effective in treating joint inflammation in a rat collagen-induced arthritis model, and the compound had an inhibitory effect on lung inflammation and eosinophilia in an ovalbumin-induced rat asthma model [126].
3.18. AnCoA4
AnCoA4 was discovered during a mass screening of 12,000 compounds, in which hits are found when small molecule compounds that bound to Orai1 and/or STIM1 in a microarray system containing minimal functional domains of STIM1and Orai1. In the study, AnCOA4 showed 80% inhibition of SOCE (HEK-293 cells, coexpressing STIM1/Orai1) at 20 µM [127]. Both in vitro and in vivo studies have shown that AnCoA4 can block CRACs and reduce the amount of T-cell activation. It carries this out by lowering the amount of Orai1 that is recruited into puncta and by inhibiting the activity of the constitutively active Orai1 V102C channels. Notably, this effect is completely independent of STIM1 [3].
3.19. ML-9
ML-9 (1-(5-chloronaphthalene-1-sulfonyl)-1H-hexahydro-1,4-diazepine), is a specific myosin light chain kinase (MLCK) inhibitor that reversibly inhibits SOCE with an IC50 value of 10 μM (Table 2). A study by Smyth et al. showed that ML-9 both prevented SOCE development and blocked SOCE following its activation [128]. It was also revealed that the effect of Ml-9 was reversible, meaning that the ICRAC current was restored after the compound was washed out [128,129]. According to a number of studies, ML-9 is one of a small set of pharmacological compounds that appear to have STIM1 as their target rather than Orai1, showing that STIM1 activation can be independently targeted to control the activity of CRAC Channel [79]. It is yet not determined where the action site of ML-9 on STIM1 is located.
3.20. JPIII
Recently, Bartoli et al. developed a new analogue of Synta-66 as a potent Orai1 inhibitor with superior pharmacokinetics over Synta-66 [130]. Moreover, in vivo, JPIII exhibited great effectiveness with no noticeable adverse effects in mouse models [130]. Notably, in HEK293 cells, JPIII demonstrated substantial nanomolar (IC50 = 399 nM) inhibitory effects of SOCE (Table 2). Furthermore, JPIII exhibited excellent selectivity towards Orai1 and had no effect on the activity of Orai3, TRPC4, TRPC5, TRPC6, TRPM2, or hERG channels [130].
3.21. Pyrtriazole Compound 39
SOCE inhibitors with anti-inflammatory properties that are derived from the Pyr family have been developed using the pyrtriazole series [82]. In HEK293 cells, lead Pyrtriazole compound (39) inhibited SOCE with an IC50 = 4.4 µM (Table 2) and was shown to be selective for SOCE over TRPM8, TRPV1, and voltage-gated Ca2+ channels, despite an analogue activating TRPV1. Moreover, Pyrtriazole 39 was tested in a mouse model of acute pancreatitis and was shown to decrease oedema, inflammation, and apoptosis, all of which are symptoms of the disease [82].
3.22. RP3128 and RP4010
Rhizen Pharmaceuticals has created SOCE inhibitors for cancer therapy, and two of them have progressed to the stage of clinical trials. In a guinea pig model of asthma, RP3128 was shown to be orally active and efficacious. It was also enrolled in a Phase I dosage escalation safety trial; however, it was later discontinued [131,132,133]. RP4010 has been explored for the treatment of oesophageal squamous cell carcinoma and has proven to be potent in various cancer cell lines as well as in xenograft animal models of the disease [3]. It needed approximately two hours to display peak inhibition of SOCE, indicating that it might have a less direct effect on the channel than immediately inhibiting Orai1 as previously thought [3]. It was enrolled in Phase I safety trials for the treatment of relapsed non-Hodgkin’s lymphoma; however the trial was stopped due to the failure of pharmacokinetic (PK) and safety considerations [134].
Only very recently, in duct cells, both in vitro and in vivo, Pallagi et al. discovered the effects of another ORAI1 inhibitor, CM5480 (CalciMedica Inc.), utilizing alcohol- or bile-induced acute pancreatitis (AP) [135]. In a cerulein-induced experimental animal model of AP, CM5480 dramatically reduced pancreatic damage while inhibiting Ca2+ influx in duct cells more slowly than GSK7975A [135].
A recent study regarding TAM, which involved wild-type and STIM1 p.I115F mice, showed that SOCE inhibitor CIC-39Na, restored platelet numbers and reverted the characteristic abnormal bleeding. CIC-39Na might be a pharmacological treatment strategy for thrombocytopenia in TAM patients [136]. Moreover, it is worth mentioning that 1,2,4-Oxadiazole-Bearing Pyrazoles was described as new SOCE inhibitor and as metabolically stable modulators of SOCE by [137]. The role of STIM2 in melanoma is still poorly understood; however, a study by Stanisz et al. described that STIM2 might act both as a tumour suppressor in highly proliferative cells and a tumour promoter in specific invasive cancers in which increased basal [Ca2+]i results in a more invasive phenotype [138].
4. Conclusions
A spike in cytosolic calcium concentration regulates biological responses ranging from sperm motility and egg fertilization to cell death via apoptosis and necrosis. Moreover, cytosolic Ca2+ controls a variety of processes occurring during the life–death cycle, including exocytosis, energy generation, gene transcription, and cell motility. In 1986, the concept of store-operated Ca2+ entry was introduced. Furthermore, over the past three decades, a large and growing body of literature has investigated the physiological and pathological role of the Orai/STIM channels in different cell types and tissues. These channels are distinctive Ca+2 signal mediators in most cell types, and their regulation and function are unmatched compared to other ion channels. They function with high selectivity for Ca+2 and remarkably mediate both short-term Ca+2 homeostasis and long-term Ca+2 signals [140].
It has been demonstrated that the Orai/STIM channels are involved in immunodeficiency, myopathy, tubular aggregate, Stormorken syndrome, York platelet syndrome, cardiovascular and metabolic disorders, cancers, kidney diseases, and other diseases and disorders [33,36,37,44,46,60,73,75].
SOCE signals are vital for cellular regulation and many cellular functions. The SOCE pathway mediated by CRAC channels regulates various physiological functions. It has been determined that ORAI1 protein is crucial for native mammalian CRAC channels, while ORAI2 and ORAI3 still remain unclear. While the crystal structure of the sole ORAI isoform in drosophilia has been determined, the relative contributions of ORAI1, ORAI2, and ORAI3 to the stoichiometry of native CRAC channels remain unresolved. Having more insight in the interaction between STIM and ORAI on the molecular level and the signal transduction that leads to channel activity would play a crucial role in further comprehending SOCE dynamics and its role in vital cellular functions, and furthermore, finding specific targets in the treatment of diseases and disorders.
Taken together, the Orai/STIM channels are crucial protein complexes as drug targets. Moreover, the absence of trustworthy antibodies, channel potentiators, or blockers that are capable of distinguishing different Orai isoforms has impeded our knowledge of the physiological roles. On that account, the next step in the fields of Orai/STIM channels will be the identification of potent, selective, and subtype-specific small molecules as agents to treat human diseases and disorders for patient benefit.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data sharing not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Putney, J.W. A Model for Receptor-Regulated Calcium Entry. Cell Calcium 1986, 7, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Zeng, B.; Chen, G.-L.; Garcia-Vaz, E.; Bhandari, S.; Daskoulidou, N.; Berglund, L.M.; Jiang, H.; Hallett, T.; Zhou, L.-P.; Huang, L.; et al. ORAI Channels Are Critical for Receptor-Mediated Endocytosis of Albumin. Nat. Commun. 2017, 8, 1920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, C.; Merritt, R.; Fu, L.; Pan, Z. Targeting Calcium Signaling in Cancer Therapy. Acta Pharm. Sin. B 2017, 7, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Rubaiy, H.N. A Short Guide to Electrophysiology and Ion Channels. J. Pharm. Pharm. Sci. 2017, 20, 48–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mignen, O.; Thompson, J.L.; Shuttleworth, T.J. Orai1 Subunit Stoichiometry of the Mammalian CRAC Channel Pore. J. Physiol. 2008, 586, 419–425. [Google Scholar] [CrossRef] [PubMed]
- Balasuriya, D.; Srivats, S.; Murrell-Lagnado, R.D.; Edwardson, J.M. Atomic Force Microscopy (AFM) Imaging Suggests That Stromal Interaction Molecule 1 (STIM1) Binds to Orai1 with Sixfold Symmetry. FEBS Lett. 2014, 588, 2874–2880. [Google Scholar] [CrossRef] [Green Version]
- Hou, X.; Pedi, L.; Diver, M.M.; Long, S.B. Crystal Structure of the Calcium Release-Activated Calcium Channel Orai. Science 2012, 338, 1308–1313. [Google Scholar] [CrossRef] [Green Version]
- Thompson, J.L.; Shuttleworth, T.J. How Many Orai’s Does It Take to Make a CRAC Channel? Sci. Rep. 2013, 3, 1961. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.; Lewis, R.S. Numbers Count: How STIM and Orai Stoichiometry Affect Store-Operated Calcium Entry. Cell Calcium 2019, 79, 35–43. [Google Scholar] [CrossRef]
- Cai, X.; Zhou, Y.; Nwokonko, R.M.; Loktionova, N.A.; Wang, X.; Xin, P.; Trebak, M.; Wang, Y.; Gill, D.L. The Orai1 Store-Operated Calcium Channel Functions as a Hexamer * ♦. J. Biol. Chem. 2016, 291, 25764–25775. [Google Scholar] [CrossRef] [Green Version]
- Yen, M.; Lokteva, L.A.; Lewis, R.S. Functional Analysis of Orai1 Concatemers Supports a Hexameric Stoichiometry for the CRAC Channel. Biophys. J. 2016, 111, 1897–1907. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, X.; Outhwaite, I.R.; Pedi, L.; Long, S.B. Cryo-EM Structure of the Calcium Release-Activated Calcium Channel Orai in an Open Conformation. eLife 2020, 9, e62772. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, M.; Yeung, P.S.-W.; Ing, C.E.; McNally, B.A.; Pomès, R.; Prakriya, M. STIM1 Activates CRAC Channels through Rotation of the Pore Helix to Open a Hydrophobic Gate. Nat. Commun. 2017, 8, 14512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiffner, A.; Maltan, L.; Fahrner, M.; Sallinger, M.; Weiß, S.; Grabmayr, H.; Höglinger, C.; Derler, I. Transmembrane Domain 3 (TM3) Governs Orai1 and Orai3 Pore Opening in an Isoform-Specific Manner. Front. Cell Dev. Biol. 2021, 9, 635705. [Google Scholar] [CrossRef] [PubMed]
- Clapham, D.E. Calcium Signaling. Cell 2007, 131, 1047–1058. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubaiy, H.N. The Therapeutic Agents That Target ATP-Sensitive Potassium Channels. Acta Pharm. Zagreb Croat. 2016, 66, 23–34. [Google Scholar] [CrossRef] [Green Version]
- Berridge, M.J. Inositol Trisphosphate and Calcium Signalling Mechanisms. Biochim. Biophys. Acta BBA-Mol. Cell Res. 2009, 1793, 933–940. [Google Scholar] [CrossRef] [Green Version]
- Liou, J.; Kim, M.L.; Heo, W.D.; Jones, J.T.; Myers, J.W.; Ferrell, J.E.; Meyer, T. STIM Is a Ca2+ Sensor Essential for Ca2+-Store-Depletion-Triggered Ca2+ Influx. Curr. Biol. CB 2005, 15, 1235–1241. [Google Scholar] [CrossRef] [Green Version]
- Amcheslavsky, A.; Wood, M.L.; Yeromin, A.V.; Parker, I.; Freites, J.A.; Tobias, D.J.; Cahalan, M.D. Molecular Biophysics of Orai Store-Operated Ca2+ Channels. Biophys. J. 2015, 108, 237–246. [Google Scholar] [CrossRef] [Green Version]
- Fahrner, M.; Grabmayr, H.; Romanin, C. Mechanism of STIM Activation. Curr. Opin. Physiol. 2020, 17, 74–79. [Google Scholar] [CrossRef]
- Park, C.Y.; Hoover, P.J.; Mullins, F.M.; Bachhawat, P.; Covington, E.D.; Raunser, S.; Walz, T.; Garcia, K.C.; Dolmetsch, R.E.; Lewis, R.S. STIM1 Clusters and Activates CRAC Channels via Direct Binding of a Cytosolic Domain to Orai1. Cell 2009, 136, 876–890. [Google Scholar] [CrossRef] [Green Version]
- Yuan, J.P.; Zeng, W.; Dorwart, M.R.; Choi, Y.-J.; Worley, P.F.; Muallem, S. SOAR and the Polybasic STIM1 Domains Gate and Regulate the Orai Channels. Nat. Cell Biol. 2009, 11, 337–343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shawer, H.; Norman, K.; Cheng, C.W.; Foster, R.; Beech, D.J.; Bailey, M.A. ORAI1 Ca2+ Channel as a Therapeutic Target in Pathological Vascular Remodelling. Front. Cell Dev. Biol. 2021, 9, 653812. [Google Scholar] [CrossRef] [PubMed]
- Prakriya, M.; Lewis, R.S. Store-Operated Calcium Channels. Physiol. Rev. 2015, 95, 1383–1436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubaiy, H.N. Treasure Troves of Pharmacological Tools to Study Transient Receptor Potential Canonical 1/4/5 Channels. Br. J. Pharmacol. 2019, 176, 832–846. [Google Scholar] [CrossRef] [PubMed]
- Rubaiy, H.N.; Ludlow, M.J.; Bon, R.S.; Beech, D.J. Pico145 - Powerful New Tool for TRPC1/4/5 Channels. Channels 2017, 11, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Rubaiy, H.N.; Ludlow, M.J.; Siems, K.; Norman, K.; Foster, R.; Wolf, D.; Beutler, J.A.; Beech, D.J. Tonantzitlolone Is a Nanomolar Potency Activator of Transient Receptor Potential Canonical 1/4/5 Channels. Br. J. Pharmacol. 2018, 175, 3361–3368. [Google Scholar] [CrossRef] [Green Version]
- Rubaiy, H.N.; Ludlow, M.J.; Henrot, M.; Gaunt, H.J.; Miteva, K.; Cheung, S.Y.; Tanahashi, Y.; Hamzah, N.; Musialowski, K.E.; Blythe, N.M.; et al. Picomolar, Selective, and Subtype-Specific Small-Molecule Inhibition of TRPC1/4/5 Channels. J. Biol. Chem. 2017, 292, 8158–8173. [Google Scholar] [CrossRef] [Green Version]
- Rubaiy, H.N.; Seitz, T.; Hahn, S.; Choidas, A.; Habenberger, P.; Klebl, B.; Dinkel, K.; Nussbaumer, P.; Waldmann, H.; Christmann, M.; et al. Identification of an (−)-englerin A Analogue, Which Antagonizes (−)-englerin A at TRPC1/4/5 Channels. Br. J. Pharmacol. 2018, 175, 830–839. [Google Scholar] [CrossRef]
- Zeng, H.; Sanes, J.R. Neuronal Cell-Type Classification: Challenges, Opportunities and the Path Forward. Nat. Rev. Neurosci. 2017, 18, 530–546. [Google Scholar] [CrossRef]
- Navarro-Borelly, L.; Somasundaram, A.; Yamashita, M.; Ren, D.; Miller, R.J.; Prakriya, M. STIM1–Orai1 Interactions and Orai1 Conformational Changes Revealed by Live-Cell FRET Microscopy. J. Physiol. 2008, 586, 5383–5401. [Google Scholar] [CrossRef] [PubMed]
- Feske, S.; Gwack, Y.; Prakriya, M.; Srikanth, S.; Puppel, S.-H.; Tanasa, B.; Hogan, P.G.; Lewis, R.S.; Daly, M.; Rao, A. A Mutation in Orai1 Causes Immune Deficiency by Abrogating CRAC Channel Function. Nature 2006, 441, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Feske, S. CRAC Channelopathies. Pflugers Arch. 2010, 460, 417–435. [Google Scholar] [CrossRef] [PubMed]
- McCarl, C.-A.; Picard, C.; Khalil, S.; Kawasaki, T.; Röther, J.; Papolos, A.; Kutok, J.; Hivroz, C.; LeDeist, F.; Plogmann, K.; et al. ORAI1 Deficiency and Lack of Store-Operated Ca2+ Entry Cause Immunodeficiency, Myopathy and Ectodermal Dysplasia. J. Allergy Clin. Immunol. 2009, 124, 1311–1318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partiseti, M.; Le Deist, F.; Hivroz, C.; Fischer, A.; Korn, H.; Choquet, D. The Calcium Current Activated by T Cell Receptor and Store Depletion in Human Lymphocytes Is Absent in a Primary Immunodeficiency. J. Biol. Chem. 1994, 269, 32327–32335. [Google Scholar] [CrossRef] [PubMed]
- Böhm, J.; Laporte, J. Gain-of-Function Mutations in STIM1 and ORAI1 Causing Tubular Aggregate Myopathy and Stormorken Syndrome. Cell Calcium 2018, 76, 1–9. [Google Scholar] [CrossRef]
- Böhm, J.; Bulla, M.; Urquhart, J.E.; Malfatti, E.; Williams, S.G.; O’Sullivan, J.; Szlauer, A.; Koch, C.; Baranello, G.; Mora, M.; et al. ORAI1 Mutations with Distinct Channel Gating Defects in Tubular Aggregate Myopathy. Hum. Mutat. 2017, 38, 426–438. [Google Scholar] [CrossRef] [Green Version]
- Okuma, H.; Saito, F.; Mitsui, J.; Hara, Y.; Hatanaka, Y.; Ikeda, M.; Shimizu, T.; Matsumura, K.; Shimizu, J.; Tsuji, S.; et al. Tubular Aggregate Myopathy Caused by a Novel Mutation in the Cytoplasmic Domain of STIM1. Neurol. Genet. 2016, 2, e50. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.-H.; Carreras-Sureda, A.; Didier, M.; Henry, C.; Frieden, M.; Demaurex, N. The TAM-Associated STIM1I484R Mutation Increases ORAI1 Channel Function Due to a Reduced STIM1 Inactivation Break and an Absence of Microtubule Trapping. Cell Calcium 2022, 105, 102615. [Google Scholar] [CrossRef]
- Endo, Y.; Noguchi, S.; Hara, Y.; Hayashi, Y.K.; Motomura, K.; Miyatake, S.; Murakami, N.; Tanaka, S.; Yamashita, S.; Kizu, R.; et al. Dominant Mutations in ORAI1 Cause Tubular Aggregate Myopathy with Hypocalcemia via Constitutive Activation of Store-Operated Ca2+ Channels. Hum. Mol. Genet. 2015, 24, 637–648. [Google Scholar] [CrossRef] [Green Version]
- Bulla, M.; Gyimesi, G.; Kim, J.H.; Bhardwaj, R.; Hediger, M.A.; Frieden, M.; Demaurex, N. ORAI1 Channel Gating and Selectivity Is Differentially Altered by Natural Mutations in the First or Third Transmembrane Domain. J. Physiol. 2019, 597, 561–582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lacruz, R.S.; Feske, S. Diseases Caused by Mutations in ORAI1 and STIM1. Ann. N. Y. Acad. Sci. 2015, 1356, 45–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morin, G.; Bruechle, N.O.; Singh, A.R.; Knopp, C.; Jedraszak, G.; Elbracht, M.; Brémond-Gignac, D.; Hartmann, K.; Sevestre, H.; Deutz, P.; et al. Gain-of-Function Mutation in STIM1 (P.R304W) Is Associated with Stormorken Syndrome. Hum. Mutat. 2014, 35, 1221–1232. [Google Scholar] [CrossRef] [PubMed]
- Nesin, V.; Wiley, G.; Kousi, M.; Ong, E.-C.; Lehmann, T.; Nicholl, D.J.; Suri, M.; Shahrizaila, N.; Katsanis, N.; Gaffney, P.M.; et al. Activating Mutations in STIM1 and ORAI1 Cause Overlapping Syndromes of Tubular Myopathy and Congenital Miosis. Proc. Natl. Acad. Sci. USA 2014, 111, 4197–4202. [Google Scholar] [CrossRef] [Green Version]
- Markello, T.; Chen, D.; Kwan, J.Y.; Horkayne-Szakaly, I.; Morrison, A.; Simakova, O.; Maric, I.; Lozier, J.; Cullinane, A.R.; Kilo, T.; et al. York Platelet Syndrome Is a CRAC Channelopathy Due to Gain-of-Function Mutations in STIM1. Mol. Genet. Metab. 2015, 114, 474–482. [Google Scholar] [CrossRef] [Green Version]
- Roman, J.; Palmer, M.I.; Palmer, C.A.; Johnson, N.E.; Butterfield, R.J. Myopathy in the York Platelet Syndrome: An Underrecognized Complication. Case Rep. Pathol. 2018, 2018, 5130143. [Google Scholar] [CrossRef]
- Pacheco, J.; Vaca, L. STIM-TRP Pathways and Microdomain Organization: Auxiliary Proteins of the STIM/Orai Complex. In Store-Operated Ca2+ Entry (SOCE) Pathways: Emerging Signaling Concepts in Human (Patho)physiology; Groschner, K., Graier, W.F., Romanin, C., Eds.; Advances in Experimental Medicine and Biology; Springer International Publishing: Cham, Switzerland, 2017; pp. 189–210. ISBN 978-3-319-57732-6. [Google Scholar]
- Xia, W.; Li, Y.; Wang, B.; Chen, J.; Wang, X.; Sun, Q.; Sun, F.; Li, Z.; Zhao, Z. Enhanced Store-Operated Calcium Entry in Platelets Is Associated with Peripheral Artery Disease in Type 2 Diabetes. Cell. Physiol. Biochem. 2015, 37, 1945–1955. [Google Scholar] [CrossRef]
- Grosse, J.; Braun, A.; Varga-Szabo, D.; Beyersdorf, N.; Schneider, B.; Zeitlmann, L.; Hanke, P.; Schropp, P.; Mühlstedt, S.; Zorn, C.; et al. An EF Hand Mutation in Stim1 Causes Premature Platelet Activation and Bleeding in Mice. J. Clin. Invest. 2007, 117, 3540–3550. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, F.; Boulaftali, Y.; Greene, T.K.; Ouellette, T.D.; Poncz, M.; Feske, S.; Bergmeier, W. Relative Contribution of STIM1 and CalDAG-GEFI to Calcium-Dependent Platelet Activation and Thrombosis. J. Thromb. Haemost. JTH 2011, 9, 2077–2086. [Google Scholar] [CrossRef] [Green Version]
- Xia, S.; Wang, L.; Fu, T.-M.; Wu, H. Mechanism of TRPM 2 Channel Gating Revealed by Cryo-EM. FEBS J. 2019, 286, 3333–3339. [Google Scholar] [CrossRef] [Green Version]
- Van Assche, T.; Fransen, P.; Guns, P.-J.; Herman, A.G.; Bult, H. Altered Ca2+ Handling of Smooth Muscle Cells in Aorta of Apolipoprotein E-Deficient Mice before Development of Atherosclerotic Lesions. Cell Calcium 2007, 41, 295–302. [Google Scholar] [CrossRef]
- Stolwijk, J.A.; Zhang, X.; Gueguinou, M.; Zhang, W.; Matrougui, K.; Renken, C.; Trebak, M. Calcium Signaling Is Dispensable for Receptor Regulation of Endothelial Barrier Function. J. Biol. Chem. 2016, 291, 22894–22912. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, H.-Y.; Kelsen, B. Influence of Personality and Motivation on Oral Presentation Performance. J. Psycholinguist. Res. 2018, 47, 755–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spinelli, A.M.; Trebak, M. Orai Channel-Mediated Ca2+ Signals in Vascular and Airway Smooth Muscle. Am. J. Physiol.-Cell Physiol. 2016, 310, C402–C413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goswamee, P.; Pounardjian, T.; Giovannucci, D.R. Arachidonic Acid-Induced Ca2+ Entry and Migration in a Neuroendocrine Cancer Cell Line. Cancer Cell Int. 2018, 18, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.-K.; Lin, Y.-H.; Chang, H.-A.; Lai, Y.-S.; Chen, Y.-C.; Huang, S.-C.; Chou, C.-Y.; Chiu, W.-T. Chemoresistant Ovarian Cancer Enhances Its Migration Abilities by Increasing Store-Operated Ca2+ Entry-Mediated Turnover of Focal Adhesions. J. Biomed. Sci. 2020, 27, 36. [Google Scholar] [CrossRef] [PubMed]
- Liang, X.; Zhang, N.; Pan, H.; Xie, J.; Han, W. Development of Store-Operated Calcium Entry-Targeted Compounds in Cancer. Front. Pharmacol. 2021, 12, 688244. [Google Scholar] [CrossRef] [PubMed]
- Schmid, E.; Stagno, M.J.; Yan, J.; Stournaras, C.; Lang, F.; Fuchs, J.; Seitz, G. Store-Operated Ca(2+) Entry in Rhabdomyosarcoma Cells. Biochem. Biophys. Res. Commun. 2016, 477, 129–136. [Google Scholar] [CrossRef]
- Wang, Y.; He, J.; Jiang, H.; Zhang, Q.; Yang, H.; Xu, X.; Zhang, C.; Xu, C.; Wang, J.; Lu, W. Nicotine Enhances Store-operated Calcium Entry by Upregulating HIF-1α and SOCC Components in Non-small Cell Lung Cancer Cells. Oncol. Rep. 2018, 40, 2097–2104. [Google Scholar] [CrossRef] [Green Version]
- Xia, J.; Wang, H.; Huang, H.; Sun, L.; Dong, S.; Huang, N.; Shi, M.; Bin, J.; Liao, Y.; Liao, W. Elevated Orai1 and STIM1 Expressions Upregulate MACC1 Expression to Promote Tumor Cell Proliferation, Metabolism, Migration, and Invasion in Human Gastric Cancer. Cancer Lett. 2016, 381, 31–40. [Google Scholar] [CrossRef]
- Chen, Y.-F.; Chiu, W.-T.; Chen, Y.-T.; Lin, P.-Y.; Huang, H.-J.; Chou, C.-Y.; Chang, H.-C.; Tang, M.-J.; Shen, M.-R. Calcium Store Sensor Stromal-Interaction Molecule 1-Dependent Signaling Plays an Important Role in Cervical Cancer Growth, Migration, and Angiogenesis. Proc. Natl. Acad. Sci. USA 2011, 108, 15225–15230. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Sanderson, M.J. Store-Operated Calcium Entry Is Required for Sustained Contraction and Ca2+ Oscillations of Airway Smooth Muscle. J. Physiol. 2017, 595, 3203–3218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, W.; Begum, G.; Pointer, K.; Clark, P.A.; Yang, S.-S.; Lin, S.-H.; Kahle, K.T.; Kuo, J.S.; Sun, D. WNK1-OSR1 Kinase-Mediated Phospho-Activation of Na+-K+-2Cl− Cotransporter Facilitates Glioma Migration. Mol. Cancer 2014, 13, 31. [Google Scholar] [CrossRef] [Green Version]
- Schild, A.; Bhardwaj, R.; Wenger, N.; Tscherrig, D.; Kandasamy, P.; Dernič, J.; Baur, R.; Peinelt, C.; Hediger, M.A.; Lochner, M. Synthesis and Pharmacological Characterization of 2-Aminoethyl Diphenylborinate (2-APB) Derivatives for Inhibition of Store-Operated Calcium Entry (SOCE) in MDA-MB-231 Breast Cancer Cells. Int. J. Mol. Sci. 2020, 21, 5604. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Collado, J.; Jardin, I.; López, J.J.; Ronco, V.; Salido, G.M.; Dubois, C.; Prevarskaya, N.; Rosado, J.A. Role of Orai3 in the Pathophysiology of Cancer. Int. J. Mol. Sci. 2021, 22, 11426. [Google Scholar] [CrossRef] [PubMed]
- Ay, B.; Prakash, Y.S.; Pabelick, C.M.; Sieck, G.C. Store-Operated Ca2+ Entry in Porcine Airway Smooth Muscle. Am. J. Physiol.-Lung Cell. Mol. Physiol. 2004, 286, L909–L917. [Google Scholar] [CrossRef] [PubMed]
- Benzerdjeb, N.; Sevestre, H.; Ahidouch, A.; Ouadid-Ahidouch, H. Orai3 Is a Predictive Marker of Metastasis and Survival in Resectable Lung Adenocarcinoma. Oncotarget 2016, 7, 81588–81597. [Google Scholar] [CrossRef] [Green Version]
- Dubois, C.; Vanden Abeele, F.; Lehen’kyi, V.; Gkika, D.; Guarmit, B.; Lepage, G.; Slomianny, C.; Borowiec, A.S.; Bidaux, G.; Benahmed, M.; et al. Remodeling of Channel-Forming ORAI Proteins Determines an Oncogenic Switch in Prostate Cancer. Cancer Cell 2014, 26, 19–32. [Google Scholar] [CrossRef] [Green Version]
- Fiorio Pla, A.; Kondratska, K.; Prevarskaya, N. STIM and ORAI Proteins: Crucial Roles in Hallmarks of Cancer. Am. J. Physiol.-Cell Physiol. 2016, 310, C509–C519. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.-F.; Lin, P.-C.; Yeh, Y.-M.; Chen, L.-H.; Shen, M.-R. Store-Operated Ca2+ Entry in Tumor Progression: From Molecular Mechanisms to Clinical Implications. Cancers 2019, 11, 899. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Xin, P.; Yoast, R.E.; Emrich, S.M.; Johnson, M.T.; Pathak, T.; Benson, J.C.; Azimi, I.; Gill, D.L.; Monteith, G.R.; et al. Distinct Pharmacological Profiles of ORAI1, ORAI2, and ORAI3 Channels. Cell Calcium 2020, 91, 102281. [Google Scholar] [CrossRef] [PubMed]
- Basile, D.P.; Collett, J.A. Orai1: A New Therapeutic Target for the Acute Kidney Injury-to-Chronic Kidney Disease Transition. Nephron 2022, 146, 264–267. [Google Scholar] [CrossRef] [PubMed]
- Mai, X.; Shang, J.; Liang, S.; Yu, B.; Yuan, J.; Lin, Y.; Luo, R.; Zhang, F.; Liu, Y.; Lv, X.; et al. Blockade of Orai1 Store-Operated Calcium Entry Protects against Renal Fibrosis. J. Am. Soc. Nephrol. JASN 2016, 27, 3063–3078. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanwar, J.; Motiani, R.K. Role of SOCE Architects STIM and Orai Proteins in Cell Death. Cell Calcium 2018, 69, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Luik, R.M.; Lewis, R.S. New Insights into the Molecular Mechanisms of Store-Operated Ca2+ Signaling in T Cells. Trends Mol. Med. 2007, 13, 103–107. [Google Scholar] [CrossRef]
- Bird, G.S.J.; Putney, J.W. Capacitative Calcium Entry Supports Calcium Oscillations in Human Embryonic Kidney Cells. J. Physiol. 2005, 562, 697–706. [Google Scholar] [CrossRef]
- Bird, G.S.; DeHaven, W.I.; Smyth, J.T.; Putney Jr, J.W. Methods for Studying Store-Operated Calcium Entry. Methods 2008, 46, 204–212. [Google Scholar] [CrossRef] [Green Version]
- Jairaman, A.; Prakriya, M. Molecular Pharmacology of Store-Operated CRAC Channels. Channels 2013, 7, 402–414. [Google Scholar] [CrossRef] [Green Version]
- Thastrup, O.; Cullen, P.J.; Drøbak, B.K.; Hanley, M.R.; Dawson, A.P. Thapsigargin, a Tumor Promoter, Discharges Intracellular Ca2+ Stores by Specific Inhibition of the Endoplasmic Reticulum Ca2(+)-ATPase. Proc. Natl. Acad. Sci. USA 1990, 87, 2466–2470. [Google Scholar] [CrossRef] [Green Version]
- Sagara, Y.; Inesi, G. Inhibition of the Sarcoplasmic Reticulum Ca2+ Transport ATPase by Thapsigargin at Subnanomolar Concentrations. J. Biol. Chem. 1991, 266, 13503–13506. [Google Scholar] [CrossRef]
- Riva, B.; Griglio, A.; Serafini, M.; Cordero-Sanchez, C.; Aprile, S.; Di Paola, R.; Gugliandolo, E.; Alansary, D.; Biocotino, I.; Lim, D.; et al. Pyrtriazoles, a Novel Class of Store-Operated Calcium Entry Modulators: Discovery, Biological Profiling and In Vivo Proof-of-Concept Efficacy in Acute Pancreatitis. J. Med. Chem. 2018, 61, 9756–9783. [Google Scholar] [CrossRef] [PubMed]
- Azimi, I.; Stevenson, R.J.; Zhang, X.; Meizoso-Huesca, A.; Xin, P.; Johnson, M.; Flanagan, J.U.; Chalmers, S.B.; Yoast, R.E.; Kapure, J.S.; et al. A New Selective Pharmacological Enhancer of the Orai1 Ca2+ Channel Reveals Roles for Orai1 in Smooth and Skeletal Muscle Functions. ACS Pharmacol. Transl. Sci. 2020, 3, 135–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hodeify, R.; Yu, F.; Courjaret, R.; Nader, N.; Dib, M.; Sun, L.; Adap, E.; Hubrack, S.; Machaca, K. Regulation and Role of Store-Operated Ca2+ Entry in Cellular Proliferation. In Calcium Entry Channels in Non-Excitable Cells; Kozak, J.A., Putney, J.W., Eds.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2018; ISBN 978-1-4987-5272-5. [Google Scholar]
- Derler, I.; Schindl, R.; Fritsch, R.; Heftberger, P.; Riedl, M.C.; Begg, M.; House, D.; Romanin, C. The Action of Selective CRAC Channel Blockers Is Affected by the Orai Pore Geometry. Cell Calcium 2013, 53, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Owsianik, G.; Talavera, K.; Voets, T.; Nilius, B. Permeation and Selectivity of TRP Channels. Annu. Rev. Physiol. 2006, 68, 685–717. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.-D.; Srikanth, S.; Yee, M.-K.W.; Mock, D.C.; Lawson, G.W.; Gwack, Y. ORAI1 Deficiency Impairs Activated T Cell Death and Enhances T Cell Survival. J. Immunol. 2011, 187, 3620–3630. [Google Scholar] [CrossRef] [Green Version]
- Ma, H.-T.; Venkatachalam, K.; Parys, J.B.; Gill, D.L. Modification of Store-Operated Channel Coupling and Inositol Trisphosphate Receptor Function by 2-Aminoethoxydiphenyl Borate in DT40 Lymphocytes. J. Biol. Chem. 2002, 277, 6915–6922. [Google Scholar] [CrossRef] [Green Version]
- DeHaven, W.I.; Smyth, J.T.; Boyles, R.R.; Putney, J.W. Calcium Inhibition and Calcium Potentiation of Orai1, Orai2, and Orai3 Calcium Release-Activated Calcium Channels *. J. Biol. Chem. 2007, 282, 17548–17556. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Du, L.; Zhou, Y.; Li, M. Store-Operated CRAC Channel Inhibitors: Opportunities and Challenges. Future Med. Chem. 2016, 8, 817–832. [Google Scholar] [CrossRef] [Green Version]
- Wei, D.; Mei, Y.; Xia, J.; Hu, H. Orai1 and Orai3 Mediate Store-Operated Calcium Entry Contributing to Neuronal Excitability in Dorsal Root Ganglion Neurons. Front. Cell. Neurosci. 2017, 11, 400. [Google Scholar] [CrossRef]
- Kappel, S.; Kilch, T.; Baur, R.; Lochner, M.; Peinelt, C. The Number and Position of Orai3 Units within Heteromeric Store-Operated Ca2+ Channels Alter the Pharmacology of ICRAC. Int. J. Mol. Sci. 2020, 21, 2458. [Google Scholar] [CrossRef] [Green Version]
- Bomben, V.C.; Sontheimer, H.W. Inhibition of Transient Receptor Potential Canonical Channels Impairs Cytokinesis in Human Malignant Gliomas. Cell Prolif. 2008, 41, 98–121. [Google Scholar] [CrossRef] [PubMed]
- Chokshi, R.; Fruasaha, P.; Kozak, J.A. 2-Aminoethyl Diphenyl Borinate (2-APB) Inhibits TRPM7 Channels through an Intracellular Acidification Mechanism. Channels 2012, 6, 362–369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kovacs, G.; Montalbetti, N.; Simonin, A.; Danko, T.; Balazs, B.; Zsembery, A.; Hediger, M.A. Inhibition of the Human Epithelial Calcium Channel TRPV6 by 2-Aminoethoxydiphenyl Borate (2-APB). Cell Calcium 2012, 52, 468–480. [Google Scholar] [CrossRef]
- Li, M.; Jiang, J.; Yue, L. Functional Characterization of Homo- and Heteromeric Channel Kinases TRPM6 and TRPM7. J. Gen. Physiol. 2006, 127, 525–537. [Google Scholar] [CrossRef] [Green Version]
- Singh, A.K.; Saotome, K.; McGoldrick, L.L.; Sobolevsky, A.I. Structural Bases of TRP Channel TRPV6 Allosteric Modulation by 2-APB. Nat. Commun. 2018, 9, 2465. [Google Scholar] [CrossRef] [PubMed]
- Goto, J.-I.; Suzuki, A.Z.; Ozaki, S.; Matsumoto, N.; Nakamura, T.; Ebisui, E.; Fleig, A.; Penner, R.; Mikoshiba, K. Two Novel 2-Aminoethyl Diphenylborinate (2-APB) Analogues Differentially Activate and Inhibit Store-Operated Ca2+ Entry via STIM Proteins. Cell Calcium 2010, 47, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Hendron, E.; Wang, X.; Zhou, Y.; Cai, X.; Goto, J.-I.; Mikoshiba, K.; Baba, Y.; Kurosaki, T.; Wang, Y.; Gill, D.L. Potent Functional Uncoupling between STIM1 and Orai1 by Dimeric 2-Aminodiphenyl Borinate Analogs. Cell Calcium 2014, 56, 482–492. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Wang, Y.; Zhou, Y.; Hendron, E.; Mancarella, S.; Andrake, M.D.; Rothberg, B.S.; Soboloff, J.; Gill, D.L. Distinct Orai-Coupling Domains in STIM1 and STIM2 Define the Orai-Activating Site. Nat. Commun. 2014, 5, 3183. [Google Scholar] [CrossRef] [Green Version]
- Núñez, L.; Valero, R.A.; Senovilla, L.; Sanz-Blasco, S.; García-Sancho, J.; Villalobos, C. Cell Proliferation Depends on Mitochondrial Ca2+ Uptake: Inhibition by Salicylate. J. Physiol. 2006, 571, 57–73. [Google Scholar] [CrossRef]
- Burn, J.; Mathers, J.C.; Bishop, D.T. Chemoprevention in Lynch Syndrome. Fam. Cancer 2013, 12, 707–718. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Sun, J.; Huang, M.-Y.; Wang, Y.-S.; Hou, M.-F.; Sun, Y.; He, H.; Krishna, N.; Chiu, S.-J.; Lin, S.; et al. STIM1 Overexpression Promotes Colorectal Cancer Progression, Cell Motility and COX-2 Expression. Oncogene 2015, 34, 4358–4367. [Google Scholar] [CrossRef] [Green Version]
- Muñoz, E.; Valero, R.A.; Quintana, A.; Hoth, M.; Núñez, L.; Villalobos, C. Nonsteroidal Anti-Inflammatory Drugs Inhibit Vascular Smooth Muscle Cell Proliferation by Enabling the Ca2+-Dependent Inactivation of Calcium Release-Activated Calcium/Orai Channels Normally Prevented by Mitochondria. J. Biol. Chem. 2011, 286, 16186–16196. [Google Scholar] [CrossRef] [Green Version]
- Cabello, O.A.; Schilling, W.P. Vectorial Ca2+ Flux from the Extracellular Space to the Endoplasmic Reticulum via a Restricted Cytoplasmic Compartment Regulates Inositol 1,4,5-Trisphosphate-Stimulated Ca2+ Release from Internal Stores in Vascular Endothelial Cells. Biochem. J. 1993, 295, 357–366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jing, Z.; Sui, X.; Yao, J.; Xie, J.; Jiang, L.; Zhou, Y.; Pan, H.; Han, W. SKF-96365 Activates Cytoprotective Autophagy to Delay Apoptosis in Colorectal Cancer Cells through Inhibition of the Calcium/CaMKIIγ/AKT-Mediated Pathway. Cancer Lett. 2016, 372, 226–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvaraj, S.; Sun, Y.; Sukumaran, P.; Singh, B.B. Resveratrol Activates Autophagic Cell Death in Prostate Cancer Cells via Downregulation of STIM1 and the MTOR Pathway. Mol. Carcinog. 2016, 55, 818–831. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Ren, Y.; Wang, L.; Zhao, W.; Dong, X.; Pan, J.; Gao, H.; Tian, Y. Orai1 and Stim1 Mediate the Majority of Store-Operated Calcium Entry in Multiple Myeloma and Have Strong Implications for Adverse Prognosis. Cell. Physiol. Biochem. 2018, 48, 2273–2285. [Google Scholar] [CrossRef]
- Zeng, B.; Yuan, C.; Yang, X.; Atkin, S.; Xu, S.-Z. TRPC Channels and Their Splice Variants Are Essential for Promoting Human Ovarian Cancer Cell Proliferation and Tumorigenesis. Curr. Cancer Drug Targets 2012, 13. [Google Scholar] [CrossRef]
- Yang, S.; Zhang, J., J.; Huang, X.-Y. Orai1 and STIM1 Are Critical for Breast Tumor Cell Migration and Metastasis | Elsevier Enhanced Reader. Available online: https://reader.elsevier.com/reader/sd/pii/S1535610808004388?token=AB3AA70BCD2C6D456CAC6DC1FAB05D8586491B411FAAA4921EEC653FF41FE3379709A8FF19E4A80ECCEBB79CB59359B3&originRegion=eu-west-1&originCreation=20220514185214 (accessed on 14 May 2022).
- Dago, C.D.; Maux, P.L.; Roisnel, T.; Brigaudeau, C.; Bekro, Y.-A.; Mignen, O.; Bazureau, J.-P. Preliminary Structure-Activity Relationship (SAR) of a Novel Series of Pyrazole SKF-96365 Analogues as Potential Store-Operated Calcium Entry (SOCE) Inhibitors. Int. J. Mol. Sci. 2018, 19, 856. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Rubaiy, H.N.; Chen, G.; Hallett, T.; Zaibi, N.; Zeng, B.; Saurabh, R.; Xu, S. Mibefradil, a T-type Ca2+ Channel Blocker Also Blocks Orai Channels by Action at the Extracellular Surface. Br. J. Pharmacol. 2019, 176, 3845–3856. [Google Scholar] [CrossRef]
- Zeng, B.; Chen, G.-L.; Daskoulidou, N.; Xu, S.-Z. The Ryanodine Receptor Agonist 4-Chloro-3-Ethylphenol Blocks ORAI Store-Operated Channels. Br. J. Pharmacol. 2014, 171, 1250–1259. [Google Scholar] [CrossRef] [Green Version]
- Xu, S.-Z.; Muraki, K.; Zeng, F.; Li, J.; Sukumar, P.; Shah, S.; Dedman, A.M.; Flemming, P.K.; McHugh, D.; Naylor, J.; et al. A Sphingosine-1–Phosphate-Activated Calcium Channel Controlling Vascular Smooth Muscle Cell Motility. Circ. Res. 2006, 98, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Zitt, C.; Strauss, B.; Schwarz, E.C.; Spaeth, N.; Rast, G.; Hatzelmann, A.; Hoth, M. Potent Inhibition of Ca2+ Release-Activated Ca2+ Channels and T-Lymphocyte Activation by the Pyrazole Derivative BTP2. J. Biol. Chem. 2004, 279, 12427–12437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putney, J.W. Pharmacology of Capacitative Calcium Entry. Mol. Interv. 2001, 1, 84–94. [Google Scholar]
- McNally, B.A.; Yamashita, M.; Engh, A.; Prakriya, M. Structural Determinants of Ion Permeation in CRAC Channels. Proc. Natl. Acad. Sci. USA 2009, 106, 22516–22521. [Google Scholar] [CrossRef] [Green Version]
- Ohga, K.; Takezawa, R.; Arakida, Y.; Shimizu, Y.; Ishikawa, J. Characterization of YM-58483/BTP2, a Novel Store-Operated Ca2+ Entry Blocker, on T Cell-Mediated Immune Responses in Vivo. Int. Immunopharmacol. 2008, 8, 1787–1792. [Google Scholar] [CrossRef] [PubMed]
- He, L.-P.; Hewavitharana, T.; Soboloff, J.; Spassova, M.A.; Gill, D.L. A Functional Link between Store-Operated and TRPC Channels Revealed by the 3,5-Bis(Trifluoromethyl)Pyrazole Derivative, BTP2 *. J. Biol. Chem. 2005, 280, 10997–11006. [Google Scholar] [CrossRef] [Green Version]
- Takezawa, R.; Cheng, H.; Beck, A.; Ishikawa, J.; Launay, P.; Kubota, H.; Kinet, J.-P.; Fleig, A.; Yamada, T.; Penner, R. A Pyrazole Derivative Potently Inhibits Lymphocyte Ca2+ Influx and Cytokine Production by Facilitating Transient Receptor Potential Melastatin 4 Channel Activity. Mol. Pharmacol. 2006, 69, 1413–1420. [Google Scholar] [CrossRef]
- Azimi, I.; Flanagan, J.U.; Stevenson, R.J.; Inserra, M.; Vetter, I.; Monteith, G.R.; Denny, W.A. Evaluation of Known and Novel Inhibitors of Orai1-Mediated Store Operated Ca 2+ Entry in MDA-MB-231 Breast Cancer Cells Using a Fluorescence Imaging Plate Reader Assay. Bioorg. Med. Chem. 2017, 25, 440–449. [Google Scholar] [CrossRef] [Green Version]
- Ng, L.C.; McCormack, M.D.; Airey, J.A.; Singer, C.A.; Keller, P.S.; Shen, X.-M.; Hume, J.R. TRPC1 and STIM1 Mediate Capacitative Ca2+ Entry in Mouse Pulmonary Arterial Smooth Muscle Cells. J. Physiol. 2009, 587, 2429–2442. [Google Scholar] [CrossRef]
- Li, J.; McKeown, L.; Ojelabi, O.; Stacey, M.; Foster, R.; O’Regan, D.; Porter, K.E.; Beech, D.J. Nanomolar Potency and Selectivity of a Ca2+ Release-Activated Ca2+ Channel Inhibitor against Store-Operated Ca2+ Entry and Migration of Vascular Smooth Muscle Cells. Br. J. Pharmacol. 2011, 164, 382–393. [Google Scholar] [CrossRef] [Green Version]
- Chen, G.; Panicker, S.; Lau, K.-Y.; Apparsundaram, S.; Patel, V.A.; Chen, S.-L.; Soto, R.; Jung, J.K.C.; Ravindran, P.; Okuhara, D.; et al. Characterization of a Novel CRAC Inhibitor That Potently Blocks Human T Cell Activation and Effector Functions. Mol. Immunol. 2013, 54, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Velicelebi, G.; Stauderman, K.; Dunn, M.; Roos, J. Pancreatitis Treatment 2016. WO2016138472A1, 26 February 2016. [Google Scholar]
- Ramos, S.; Grigoryev, S.; Rogers, E.; Roos, J.; Whitten, J.; Stauderman, K.; Velicelebi, G. CM3457, a Potent and Selective Oral CRAC Channel Inhibitor, Suppresses T and Mast Cell Function and Is Efficacious in Rat Models of Arthritis and Asthma (72.3). J. Immunol. 2012, 188, 72–73. [Google Scholar] [CrossRef]
- Sadaghiani, A.M.; Lee, S.M.; Odegaard, J.I.; Leveson-Gower, D.B.; McPherson, O.M.; Novick, P.; Kim, M.R.; Koehler, A.N.; Negrin, R.; Dolmetsch, R.E.; et al. Identification of Orai1 Channel Inhibitors by Using Minimal Functional Domains to Screen Small Molecule Microarrays. Chem. Biol. 2014, 21, 1278–1292. [Google Scholar] [CrossRef] [PubMed]
- Smyth, J.T.; DeHaven, W.I.; Bird, G.S.; Putney, J.W. Calcium store-dependent and independent reversal of stim1 localization and function. J. Cell Sci. 2008, 121, 762–772. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, T.; Kitani, A.; Murray, P.J.; Wakatsuki, Y.; Fuss, I.J.; Strober, W. Nucleotide Binding Oligomerization Domain 2 Deficiency Leads to Dysregulated TLR2 Signaling and Induction of Antigen-Specific Colitis. Immunity 2006, 25, 473–485. [Google Scholar] [CrossRef] [Green Version]
- Bartoli, F.; Bailey, M.A.; Rode, B.; Mateo, P.; Antigny, F.; Bedouet, K.; Gerbaud, P.; Gosain, R.; Plante, J.; Norman, K.; et al. Orai1 Channel Inhibition Preserves Left Ventricular Systolic Function and Normal Ca2+ Handling After Pressure Overload. Circulation 2020, 141, 199–216. [Google Scholar] [CrossRef]
- Barde, P.J.; Viswanadha, S.; Veeraraghavan, S.; Vakkalanka, S.V.; Nair, A. A First-in-Human Study to Evaluate the Safety, Tolerability and Pharmacokinetics of RP3128, an Oral Calcium Release-Activated Calcium (CRAC) Channel Modulator in Healthy Volunteers. J. Clin. Pharm. Ther. 2021, 46, 677–687. [Google Scholar] [CrossRef] [PubMed]
- Sutovska, M.; Kocmalova, M.; Franova, S.; Vakkalanka, S.; Viswanadha, S. Pharmacodynamic Evaluation of RP3128, a Novel and Potent CRAC Channel Inhibitor in Guinea Pig Models of Allergic Asthma. Eur. J. Pharmacol. 2016, 772, 62–70. [Google Scholar] [CrossRef] [PubMed]
- Vakkalanka, S.; Merikapudi, G.; Babu, G.; Routhu, K.; Veeraraghavan, S.; Viswanadha, S. Pre-Clinical Characterization of RP3128, a Novel and Potent CRAC Channel Inhibitor for the Treatment of Respiratory Disorders . Available online: https://www.ers-education.org/lr/show-details/?idP=127076 (accessed on 16 May 2022).
- Rhizen Pharmaceuticals SA. A Phase I/Ib, Study to Evaluate Safety and Efficacy of RP4010, in Patients With Relapsed or Refractory Lymphomas. 2019. Available online: https://cinicaltrials.gov (accessed on 14 May 2022).
- Pallagi, P.; Görög, M.; Papp, N.; Madácsy, T.; Varga, Á.; Crul, T.; Szabó, V.; Molnár, M.; Dudás, K.; Grassalkovich, A.; et al. Bile Acid- and Ethanol-Mediated Activation of Orai1 Damages Pancreatic Ductal Secretion in Acute Pancreatitis. J. Physiol. 2022, 600, 1631–1650. [Google Scholar] [CrossRef]
- Cordero-Sanchez, C.; Pessolano, E.; Riva, B.; Vismara, M.; Trivigno, S.M.G.; Clemente, N.; Aprile, S.; Ruffinatti, F.A.; Portararo, P.; Filigheddu, N.; et al. CIC-39Na Reverses the Thrombocytopenia That Characterizes Tubular Aggregate Myopathy. Blood Adv. 2022, 6, 4471–4484. [Google Scholar] [CrossRef]
- Aprile, S.; Riva, B.; Bhela, I.P.; Cordero-Sanchez, C.; Avino, G.; Genazzani, A.A.; Serafini, M.; Pirali, T. 1,2,4-Oxadiazole-Bearing Pyrazoles as Metabolically Stable Modulators of Store-Operated Calcium Entry. ACS Med. Chem. Lett. 2021, 12, 640–646. [Google Scholar] [CrossRef] [PubMed]
- Stanisz, H.; Saul, S.; Müller, C.S.L.; Kappl, R.; Niemeyer, B.A.; Vogt, T.; Hoth, M.; Roesch, A.; Bogeski, I. Inverse Regulation of Melanoma Growth and Migration by Orai1/STIM2-Dependent Calcium Entry. Pigment Cell Melanoma Res. 2014, 27, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Chung, S.C.; McDonald, T.V.; Gardner, P. Inhibition by SK&F 96365 of Ca2+ Current, IL-2 Production and Activation in T Lymphocytes. Br. J. Pharmacol. 1994, 113, 861–868. [Google Scholar] [PubMed]
- Rothberg, B.S.; Wang, Y.; Gill, D.L. Orai Channel Pore Properties and Gating by STIM: Implications from the Orai Crystal Structure. Sci. Signal. 2013, 6, pe9. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
The hexameric Orai channel structure. The figure illustrates a side view of the overall structure of Orai channel complex from the perspective of the membrane (A), a side view of just the four TMs (B), a top view (C), and intracellular view (D). Each subunit contains four transmembrane helices as represented in different colours. The red ball in the centre of the pore represents the Ca2+ ion.
Figure 1.
The hexameric Orai channel structure. The figure illustrates a side view of the overall structure of Orai channel complex from the perspective of the membrane (A), a side view of just the four TMs (B), a top view (C), and intracellular view (D). Each subunit contains four transmembrane helices as represented in different colours. The red ball in the centre of the pore represents the Ca2+ ion.

Figure 2.
The chemical structure of Orai/STIM channel blockers. (A) Synta-66, (B) RO2959, (C) BTP2, (D) GSK-7975A, (E) CM4620, (F) JPIII.
Figure 2.
The chemical structure of Orai/STIM channel blockers. (A) Synta-66, (B) RO2959, (C) BTP2, (D) GSK-7975A, (E) CM4620, (F) JPIII.


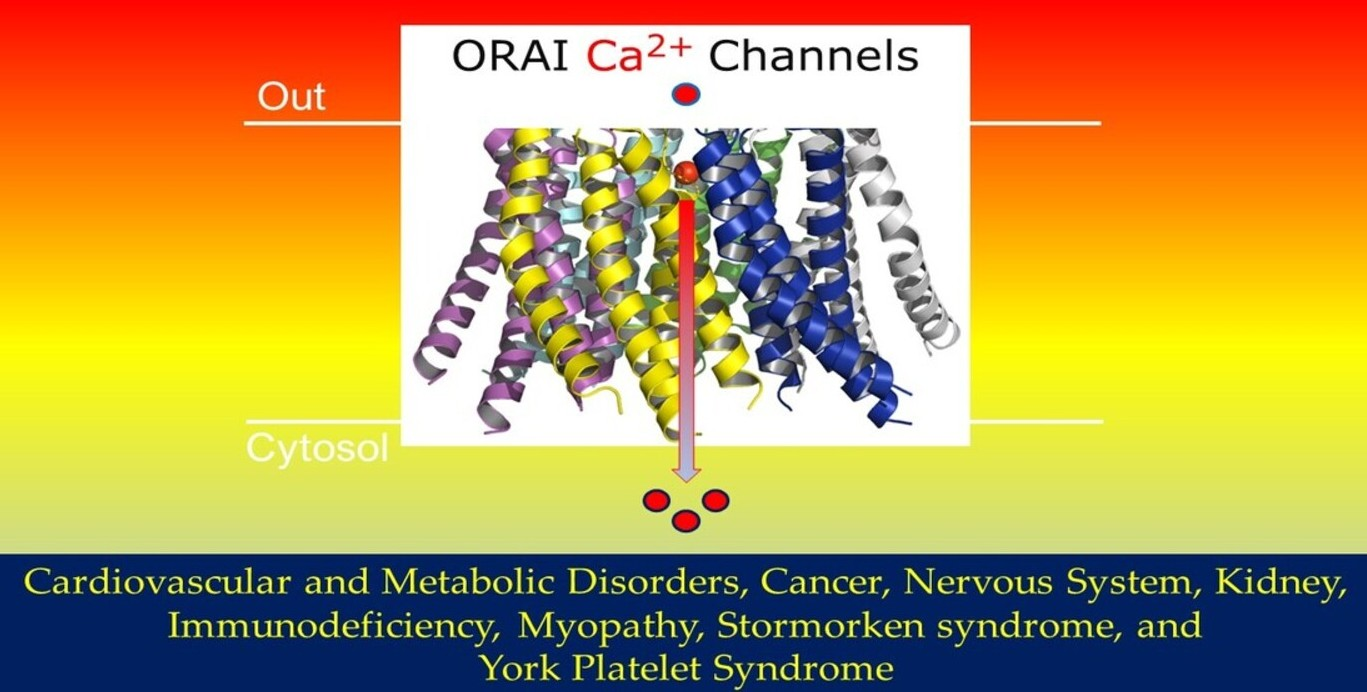{kind=link}
{kind=link}
{kind=link}
Table 1.
Orai/STIM channel complexes in human diseases.
| Diseases | Subunit Combination | Reference(s) |
|---|---|---|
| Immunodeficiency | Orai1/STIM1 | [33] |
| Myopathy | Orai1/STIM1 | [37] |
| Tubular aggregate myopathy | Orai1/STIM1 | [36] |
| Stormorken syndrome | Orai1/STIM1 | [44] |
| York platelet syndrome | Orai1/STIM1 | [46] |
| Cardiovascular and metabolic disorders | Oria1/STIM1 | [75] |
| Cancers | Oria1/STIM1 ORAI3 (lung cancer) STIM2 (tumour growth) | [60] [68] [63] |
| Kidney | Orai1 | [73] |
Table 2.
Selected Orai-STIM channel blockers.
| Name | IC50 | Cell Types | Reference |
|---|---|---|---|
| GSK-5503A & GSK-7975A | 4 μM | HEK-293 | [85] |
| 5J-4 | 807 nM (peak) 195 nM (sustained) | Primary murine effector T TH17 HeLa-O + S cells | [87] |
| DPB162-AE | 200 nM | HEK-293 | [99] |
| 4-CEP | 203.6 µM | HEK-293 | [113] |
| 4-CmC | 830.9 1 µM | HEK-293 | [113] |
| 4-CIP | 1437.1 µM | HEK-293 | [113] |
| SKF96365 | 12 µM | T leukaemic | [139] |
| BTP2 (YM-58483) | 2.8 µM | MDA-MB-231 breast cancer cells | [121] |
| Synta-66 | 1.4 μM | Rat basophilic leukaemia (RBL) | [122] |
| ML-9 | 10 μM | HEK-293 | [128,129] |
| JPIII | 399 nM | HEK-293 | [130] |
| Pyr 39 | 4.4 μM | HEK-293 | [82] |
| RO2959 | 265 nM | CD4+T lymphocytes | [124] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Rubaiy, H.N. ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets. Pharmaceuticals 2023, 16, 162. https://doi.org/10.3390/ph16020162
AMA Style
Rubaiy HN. ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets. Pharmaceuticals. 2023; 16(2):162. https://doi.org/10.3390/ph16020162
Chicago/Turabian StyleRubaiy, Hussein N. 2023. "ORAI Calcium Channels: Regulation, Function, Pharmacology, and Therapeutic Targets" Pharmaceuticals 16, no. 2: 162. https://doi.org/10.3390/ph16020162
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.