
Free Amino Acid Alterations in Patients with Gynecological and Breast Cancer: A Review

 , , , , , ,
, , , , , ,
Abstract
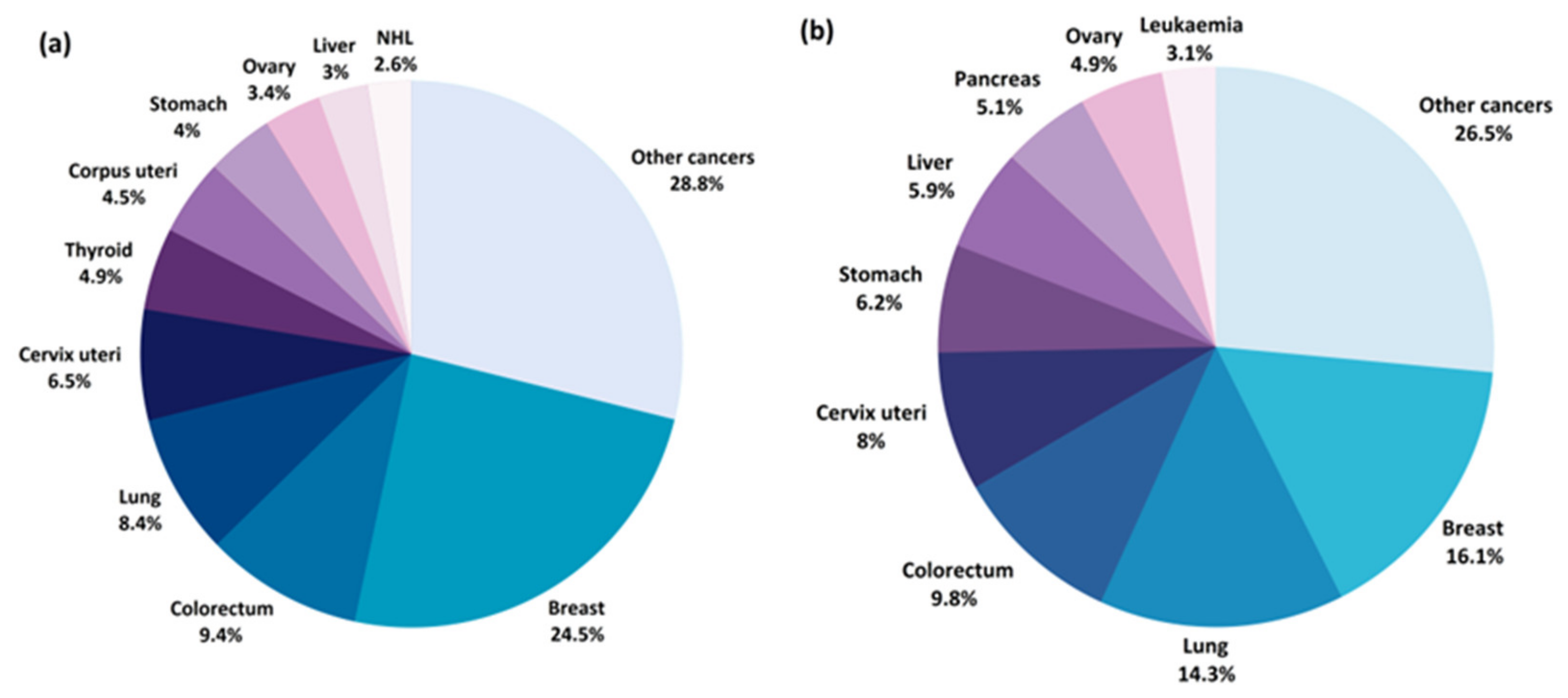
:1. Introduction
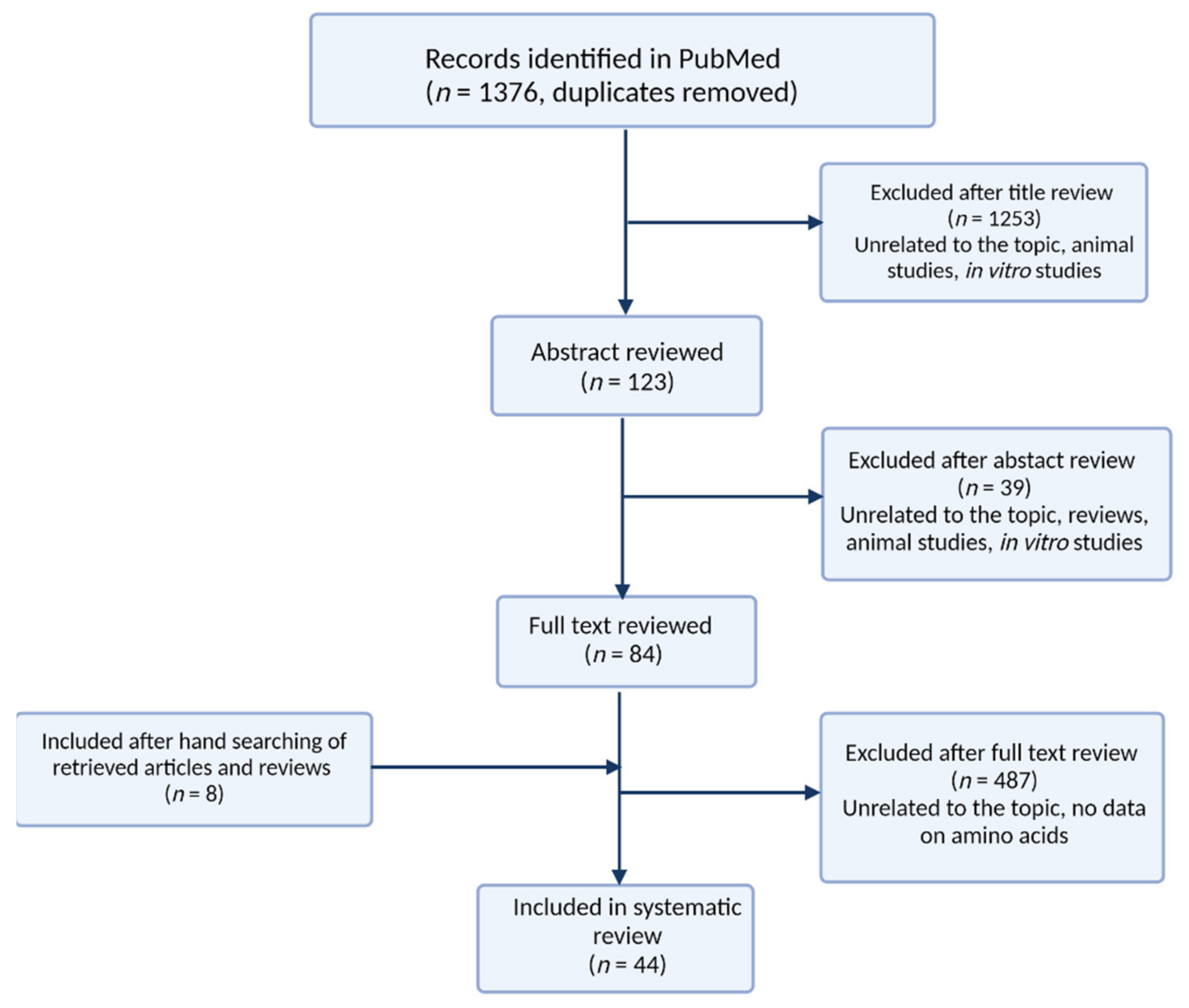
2. Methods
- original studies;
- studies focused on changes in amino acid profiles in patients with gynecological cancers (endometrial cancer, ovarian cancer, breast cancer, chorionic carcinoma, cervical cancer, vulvar cancer) and breast cancer
- articles in English.
- reviews
- meta-analyses
- letters
- comments
- articles unrelated to the topic
- studies on cell lines or animals
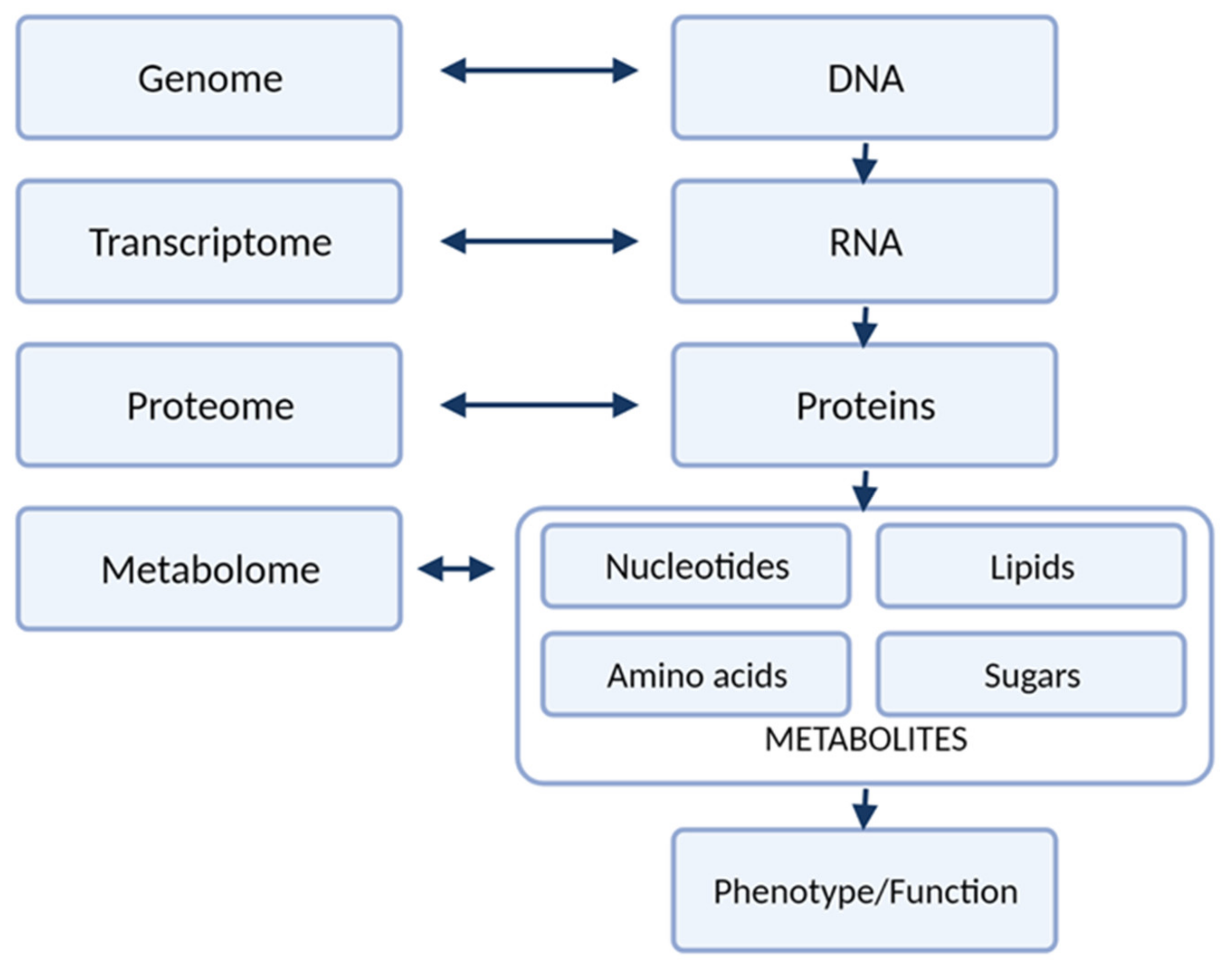
3. Metabolomic Platforms Used for Analysis of Amino Acids
Validation
4. Biological Matrices
4.1. Blood Based Matrices
4.2. Urine
4.3. Other Matrices
5. Metabolomic Studies of Gynecological Cancers
6. Amino Acid Profile Changes in Gynecological Cancers
6.1. Breast Cancer
6.2. Cervical Cancer
6.3. Endometrial Cancer
6.4. Ovarian Cancer
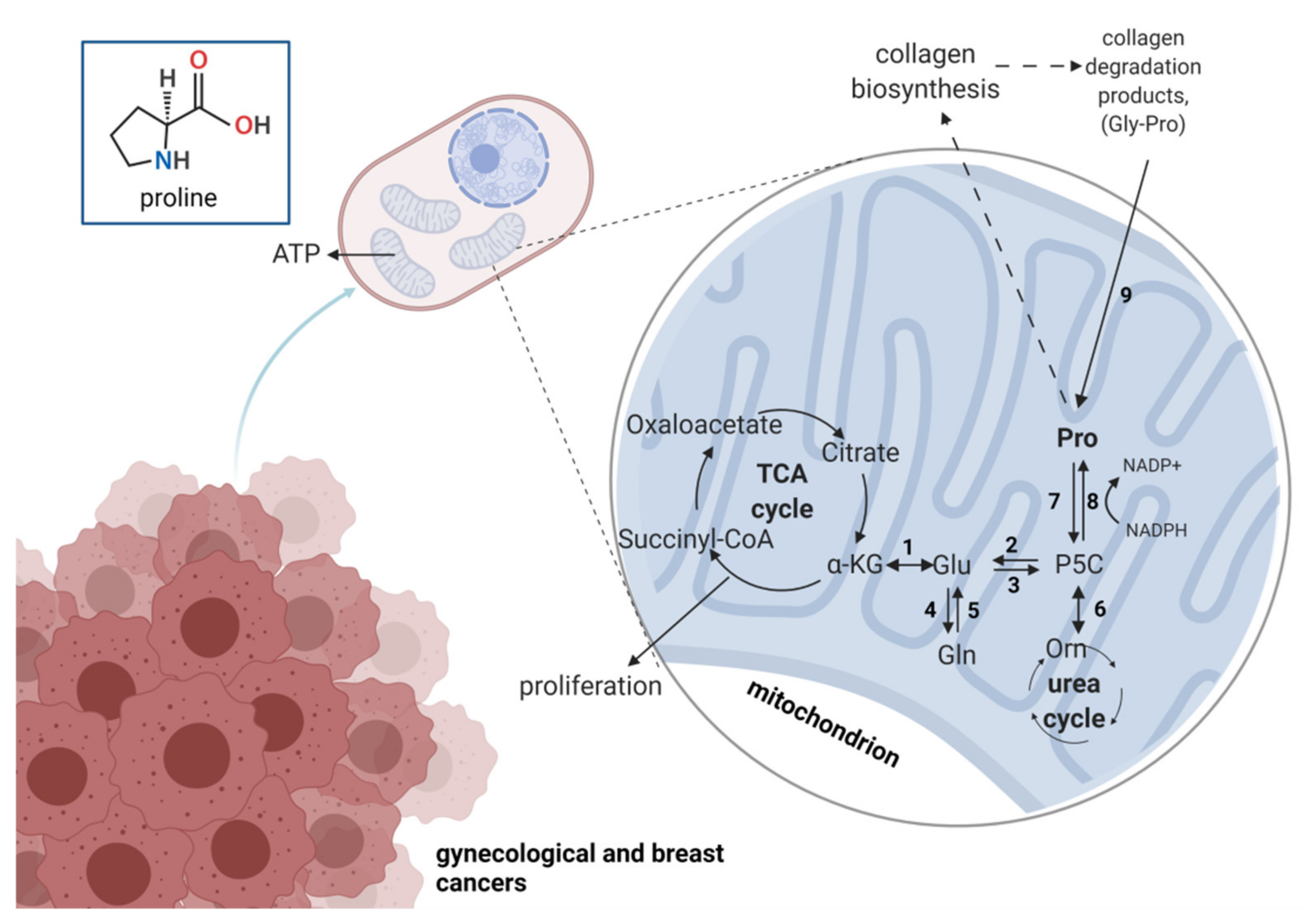
7. Role of Proline Metabolism in Gynecological Cancers
8. Concluding Remarks
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Louise Gaynor Metabolomics: Personalised Healthcare through Measuring Metabolites? Available online: https://www.phgfoundation.org/blog/what-is-metabolomics (accessed on 11 June 2021).
- Klupczynska, A.; Derezinski, P.; Kokot, Z.J. Metabolomics in medical sciences—Trends, challenges and perspectives. Acta Pol. Pharm. 2015, 72, 629–641. [Google Scholar]
- Plewa, S.; Dereziński, P.; Florczak-Wyspiańska, J.; Popławska-Domaszewicz, K.; Kozubski, W.; Sokół, B.; Jankowski, R.; Matysiak, J.; Kokot, Z.J. LC-MS/MS based targeted metabolomics method for analysis of serum and cerebrospinal fluid. J. Med. Sci. 2019, 88, 12–20. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef]
- Heer, E.; Harper, A.; Escandor, N.; Sung, H.; McCormack, V.; Fidler-Benaoudia, M.M. Global burden and trends in premenopausal and postmenopausal breast cancer: A population-based study. Lancet Glob. Health 2020, 8, e1027–e1037. [Google Scholar] [CrossRef]
- Paik, E.S.; Lee, Y.-Y.; Lee, E.-J.; Choi, C.H.; Kim, T.-J.; Lee, J.-W.; Bae, D.-S.; Kim, B.-G. Survival analysis of revised 2013 FIGO staging classification of epithelial ovarian cancer and comparison with previous FIGO staging classification. Obstet. Gynecol. Sci. 2015, 58, 124. [Google Scholar] [CrossRef] [PubMed]
- Partridge, E.; Kreimer, A.R.; Greenlee, R.T.; Williams, C.; Xu, J.L.; Church, T.R.; Kessel, B.; Johnson, C.C.; Weissfeld, J.L.; Isaacs, C.; et al. Results from four rounds of ovarian cancer screening in a randomized trial. Obstet. Gynecol. 2009, 113, 775–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Global Cancer Observatory. Available online: https://gco.iarc.fr/ (accessed on 25 May 2021).
- Ihata, Y.; Miyagi, E.; Numazaki, R.; Muramatsu, T.; Imaizumi, A.; Yamamoto, H.; Yamakado, M.; Okamoto, N.; Hirahara, F. Amino acid profile index for early detection of endometrial cancer: Verification as a novel diagnostic marker. Int. J. Clin. Oncol. 2014, 19, 364–372. [Google Scholar] [CrossRef] [PubMed]
- Troisi, J.; Sarno, L.; Landolfi, A.; Scala, G.; Martinelli, P.; Venturella, R.; Di Cello, A.; Zullo, F.; Guida, M. Metabolomic Signature of Endometrial Cancer. J. Proteome Res. 2018, 17, 804–812. [Google Scholar] [CrossRef]
- Gaudet, M.M.; Falk, R.T.; Stevens, R.D.; Gunter, M.J.; Bain, J.R.; Pfeiffer, R.M.; Potischman, N.; Lissowska, J.; Peplonska, B.; Brinton, L.A.; et al. Analysis of Serum Metabolic Profiles in Women with Endometrial Cancer and Controls in a Population-Based Case-Control Study. J. Clin. Endocrinol. Metab. 2012, 97, 3216–3223. [Google Scholar] [CrossRef]
- Shi, K.; Wang, Q.; Su, Y.; Xuan, X.; Liu, Y.; Chen, W.; Qian, Y.; Lash, G.E. Identification and functional analyses of differentially expressed metabolites in early stage endometrial carcinoma. Cancer Sci. 2018, 109, 1032–1043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Suzuki, Y.; Tokinaga-uchiyama, A.; Mizushima, T.; Maruyama, Y.; Mogami, T.; Shikata, N. Normalization of abnormal plasma amino acid profile-based indexes in patients with gynecological malignant tumors after curative treatment. BMC Cancer 2018, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Bahado-Singh, R.O.; Lugade, A.; Field, J.; Al-Wahab, Z.; Han, B.S.; Mandal, R.; Bjorndahl, T.C.; Turkoglu, O.; Graham, S.F.; Wishart, D.; et al. Metabolomic prediction of endometrial cancer. Metabolomics 2018, 14, 6. [Google Scholar] [CrossRef]
- Audet-Delage, Y.; Villeneuve, L.; Grégoire, J.; Plante, M.; Guillemette, C. Identification of metabolomic biomarkers for endometrial cancer and its recurrence after surgery in postmenopausal women. Front. Endocrinol. 2018, 9, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strand, E.; Tangen, I.L.; Fasmer, K.E.; Jacob, H.; Halle, M.K.; Hoivik, E.A.; Delvoux, B.; Trovik, J.; Haldorsen, I.S.; Romano, A.; et al. Blood metabolites associate with prognosis in endometrial cancer. Metabolites 2019, 9, 302. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Altadill, T.; Dowdy, T.M.; Gill, K.; Reques, A.; Menon, S.S.; Moiola, C.P.; Lopez-Gil, C.; Coll, E.; Matias-Guiu, X.; Cabrera, S.; et al. Metabolomic and Lipidomic Profiling Identifies the Role of the RNA Editing Pathway in Endometrial Carcinogenesis. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Trousil, S.; Lee, P.; Pinato, D.J.; Ellis, J.K.; Dina, R.; Aboagye, E.O.; Keun, H.C.; Sharma, R. Alterations of choline phospholipid metabolism in endometrial cancer are caused by choline kinase alpha overexpression and a hyperactivated deacylation pathway. Cancer Res. 2014, 74, 6867–6877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, S.C.; Chen, K.; Chiu, C.Y.; Lu, K.Y.; Lu, H.Y.; Chiang, M.H.; Tsai, C.K.; Lo, C.J.; Cheng, M.L.; Chang, T.C.; et al. Metabolomic biomarkers in cervicovaginal fluid for detecting endometrial cancer through nuclear magnetic resonance spectroscopy. Metabolomics 2019, 15. [Google Scholar] [CrossRef]
- Zhou, M.; Guan, W.; Walker, L.D.E.; Mezencev, R.; Benigno, B.B.; Gray, A.; Fernández, F.M.; McDonald, J.F. Rapid mass spectrometric metabolic profiling of blood sera detects ovarian cancer with high accuracy. Cancer Epidemiol. Biomark. Prev. 2010, 19, 2262–2271. [Google Scholar] [CrossRef] [Green Version]
- Hilvo, M.; De Santiago, I.; Gopalacharyulu, P.; Schmitt, W.D.; Budczies, J.; Kuhberg, M.; Dietel, M.; Aittokallio, T.; Markowetz, F.; Denkert, C.; et al. Accumulated metabolites of hydroxybutyric acid serve as diagnostic and prognostic biomarkers of ovarian high-grade serous carcinomas. Cancer Res. 2016, 76, 796–804. [Google Scholar] [CrossRef] [Green Version]
- Garcia, E.; Andrews, C.; Hua, J.; Kim, H.L.; Sukumaran, D.K.; Szyperski, T.; Odunsi, K. Diagnosis of early stage ovarian cancer by 1 H NMR metabonomics of serum explored by use of a micro-flow NMR probe. J. Proteome Res. 2012, 10, 1765–1771. [Google Scholar] [CrossRef] [Green Version]
- Bachmayr-Heyda, A.; Aust, S.; Auer, K.; Meier, S.M.; Schmetterer, K.G.; Dekan, S.; Gerner, C.; Pils, D. Integrative systemic and local metabolomics with impact on survival in high-grade serous ovarian cancer. Clin. Cancer Res. 2017, 23, 2081–2092. [Google Scholar] [CrossRef] [Green Version]
- Buas, M.F.; Gu, H.; Djukovic, D.; Zhu, J.; Drescher, C.W.; Urban, N.; Raftery, D.; Li, C.I. Identification of novel candidate plasma metabolite biomarkers for distinguishing serous ovarian carcinoma and benign serous ovarian tumors. Gynecol. Oncol. 2016, 176, 139–148. [Google Scholar] [CrossRef] [Green Version]
- Ke, C.; Hou, Y.; Zhang, H.; Fan, L.; Ge, T.; Guo, B.; Zhang, F.; Yang, K.; Wang, J.; Lou, G.; et al. Large-scale profiling of metabolic dysregulation in ovarian cancer. Int. J. Cancer 2015, 136, 516–526. [Google Scholar] [CrossRef]
- Miyagi, E.; Maruyama, Y.; Mogami, T.; Numazaki, R.; Ikeda, A.; Yamamoto, H.; Hirahara, F. Comparison of plasma amino acid profile-based index and CA125 in the diagnosis of epithelial ovarian cancers and borderline malignant tumors. Int. J. Clin. Oncol. 2017, 22, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wu, X.; Ke, C.; Yin, M.; Li, Z.; Fan, L.; Zhang, W.; Zhang, H.; Zhao, F.; Zhou, X.; et al. Identification of potential biomarkers for ovarian cancer by urinary metabolomic profiling. J. Proteome Res. 2013, 12, 505–512. [Google Scholar] [CrossRef] [PubMed]
- Horala, A.; Plewa, S.; Derezinski, P.; Klupczynska, A.; Matysiak, J.; Nowak-Markwitz, E.; Kokot, Z.J. Serum free amino acid profiling in differential diagnosis of ovarian tumors—a comparative study with review of the literature. Int. J. Environ. Res. Public Health 2021, 18, 2167. [Google Scholar] [CrossRef]
- Plewa, S.; Horała, A.; Dereziński, P.; Klupczynska, A.; Nowak-Markwitz, E.; Matysiak, J.; Kokot, Z.J. Usefulness of amino acid profiling in ovarian cancer screening with special emphasis on their role in cancerogenesis. Int. J. Mol. Sci. 2017, 18, 2727. [Google Scholar] [CrossRef] [Green Version]
- Plewa, S.; Horała, A.; Dereziński, P.; Nowak-Markwitz, E.; Matysiak, J.; Kokot, Z.J. Wide spectrum targeted metabolomics identifies potential ovarian cancer biomarkers. Life Sci. 2019, 222, 235–244. [Google Scholar] [CrossRef]
- Slupsky, C.M.; Steed, H.; Wells, T.H.; Dabbs, K.; Schepansky, A.; Capstick, V.; Faught, W.; Sawyer, M.B. Urine metabolite analysis offers potential early diagnosis of ovarian and breast cancers. Clin. Cancer Res. 2010, 16, 5835–5841. [Google Scholar] [CrossRef] [Green Version]
- Ahn, H.S.; Yeom, J.; Yu, J.; Kwon, Y.I.; Kim, J.H.; Kim, K. Convergence of plasma metabolomics and proteomics analysis to discover signatures of high-grade serous ovarian cancer. Cancers 2020, 12, 3447. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Zhao, X.; Zhao, J.; Yang, T.; Zhang, F.; Liu, L. Serum metabolite signatures of epithelial ovarian cancer based on targeted metabolomics. Clin. Chim. Acta 2021, 518, 59–69. [Google Scholar] [CrossRef]
- His, M.; Viallon, V.; Dossus, L.; Gicquiau, A.; Achaintre, D.; Scalbert, A.; Ferrari, P.; Romieu, I.; Onland-Moret, N.C.; Weiderpass, E.; et al. Prospective analysis of circulating metabolites and breast cancer in EPIC. BMC Med. 2019, 17, 29. [Google Scholar] [CrossRef]
- Eniu, D.T.; Romanciuc, F.; Moraru, C.; Goidescu, I.; Eniu, D.; Staicu, A.; Rachieriu, C.; Buiga, R.; Socaciu, C. The decrease of some serum free amino acids can predict breast cancer diagnosis and progression. Scand. J. Clin. Lab. Invest. 2019, 79, 17–24. [Google Scholar] [CrossRef] [PubMed]
- Mitruka, M.; Gore, C.R.; Kumar, A.; Sarode, S.C.; Sharma, N.K. Undetectable Free Aromatic Amino Acids in Nails of Breast Carcin1. Mitruka, M.; Gore, C.R.; Kumar, A.; Sarode, S.C.; Sharma, N.K. Undetectable Free Aromatic Amino Acids in Nails of Breast Carcinoma: Biomarker Discovery by a Novel Metabolite Purification V. Front. Oncol. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Shen, J.; Yan, L.; Liu, S.; Ambrosone, C.B.; Zhao, H. Plasma metabolomic profiles in breast cancer patients and healthy controls: By race and tumor receptor subtypes. Transl. Oncol. 2013, 6, 757–765. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Budczies, J.; Brockmöller, S.F.; Müller, B.M.; Barupal, D.K.; Richter-Ehrenstein, C.; Kleine-Tebbe, A.; Griffin, J.L.; Orešič, M.; Dietel, M.; Denkert, C.; et al. Comparative metabolomics of estrogen receptor positive and estrogen receptor negative breast cancer: Alterations in glutamine and beta-alanine metabolism. J. Proteomics 2013, 94, 279–288. [Google Scholar] [CrossRef]
- Cala, M.P.; Aldana, J.; Medina, J.; Sánchez, J.; Guio, J.; Wist, J.; Meesters, R.J.W. Multiplatform plasma metabolic and lipid fingerprinting of breast cancer: A pilot control-case study in Colombian Hispanic women. PLoS ONE 2019, 13, 1–23. [Google Scholar] [CrossRef] [Green Version]
- Miyagi, Y.; Higashiyama, M.; Gochi, A.; Akaike, M.; Ishikawa, T.; Miura, T.; Saruki, N.; Bando, E.; Kimura, H.; Imamura, F.; et al. Plasma free amino acid profiling of five types of cancer patients and its application for early detection. PLoS ONE 2011, 6, e24143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moore, S.C.; Mazzilli, K.M.; Sampson, J.N.; Matthews, C.E.; Carter, B.D.; Playdon, M.C.; Wang, Y.; Stevens, V.L. A metabolomics analysis of postmenopausal breast cancer risk in the cancer prevention study II. Metabolites 2021, 11, 95. [Google Scholar] [CrossRef]
- Cao, Z.; Qin, X.; Liu, F.; Zhou, L. Tryptophan-induced pathogenesis of breast cancer. Afr. Health Sci. 2015, 15, 982–985. [Google Scholar] [CrossRef] [Green Version]
- Xie, G.; Zhou, B.; Zhao, A.; Qiu, Y.; Zhao, X.; Garmire, L.; Shvetsov, Y.B.; Yu, H.; Yen, Y.; Jia, W. Lowered circulating aspartate is a metabolic feature of human breast cancer. Oncotarget 2015, 6, 33369–33381. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Sun, T.; Cao, Y.; Gao, P.; Dong, J.; Fang, Y.; Fang, Z.; Sun, X.; Zhu, Z. Adried blood spot mass spectrometry metabolomic approach for rapid breast cancer detection. OncoTargets Ther. 2016, 9, 1389–1398. [Google Scholar] [CrossRef] [Green Version]
- Jasbi, P.; Wang, D.; Cheng, S.L.; Fei, Q.; Cui, J.Y.; Liu, L.; Wei, Y.; Raftery, D.; Gu, H. Breast cancer detection using targeted plasma metabolomics. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2019, 1105, 26–37. [Google Scholar] [CrossRef]
- Yuan, B.; Schafferer, S.; Tang, Q.; Scheffler, M.; Nees, J.; Heil, J.; Schott, S.; Golatta, M.; Wallwiener, M.; Sohn, C.; et al. A plasma metabolite panel as biomarkers for early primary breast cancer detection. Int. J. Cancer 2019, 144, 2833–2842. [Google Scholar] [CrossRef]
- Li, L.; Zheng, X.; Zhou, Q.; Villanueva, N.; Nian, W.; Liu, X.; Huan, T. Metabolomics-Based Discovery of Molecular Signatures for Triple Negative Breast Cancer in Asian Female Population. Sci. Rep. 2020, 10, 1–12. [Google Scholar] [CrossRef]
- Khan, I.; Nam, M.; Kwon, M.; Seo, S.S.; Jung, S.; Han, J.S.; Hwang, G.S.; Kim, M.K. Lc/ms-based polar metabolite profiling identified unique biomarker signatures for cervical cancer and cervical intraepithelial neoplasia using global and targeted metabolomics. Cancers 2019, 11, 511. [Google Scholar] [CrossRef] [Green Version]
- Abudula, A.; Rouzi, N.; Xu, L.; Yang, Y.; Hasimu, A. Tissue-based metabolomics reveals potential biomarkers for cervical carcinoma and HPV infection. Bosn. J. Basic Med. Sci. 2020, 20, 78–87. [Google Scholar] [CrossRef] [Green Version]
- Yang, K.; Xia, B.; Wang, W.; Cheng, J.; Yin, M.; Xie, H.; Li, J.; Ma, L.; Yang, C.; Li, A.; et al. A Comprehensive Analysis of Metabolomics and Transcriptomics in Cervical Cancer. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Chen, Y.; Xu, J.; Zhang, R.; Shen, G.; Song, Y.; Sun, J.; He, J.; Zhan, Q.; Abliz, Z. Assessment of data pre-processing methods for LC-MS/MS-based metabolomics of uterine cervix cancer. Analyst 2013, 138, 2669–2677. [Google Scholar] [CrossRef] [PubMed]
- Hasim, A.; Ali, M.; Mamtimin, B.; Ma, J.Q.; Li, Q.Z.; Abudula, A. Metabonomic signature analysis of cervical carcinoma and precancerous lesions in women by 1H NMR spectroscopy. Exp. Ther. Med. 2012, 3, 945–951. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Putri, S.P.; Yamamoto, S.; Tsugawa, H.; Fukusaki, E. Current metabolomics: Technological advances. J. Biosci. Bioeng. 2013, 116, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Gika, H.G.; Theodoridis, G.A.; Plumb, R.S.; Wilson, I.D. Current practice of liquid chromatography-mass spectrometry in metabolomics and metabonomics. J. Pharm. Biomed. Anal. 2014, 87, 12–25. [Google Scholar] [CrossRef]
- Deng, C.; Li, W.; Zhang, X. Rapid determination of amino acids in neonatal blood samples based on derivatization with isobutyl chloroformate followed by solid-phase microextraction and gas chromatography/mass spectrometry. Rapid Commun. Mass Spectrom. 2004, 18, 2558–2564. [Google Scholar] [CrossRef]
- de Paiva, M.J.N.; Menezes, H.C.; Christo, P.P.; Resende, R.R.; Cardeal, Z.D.L. An alternative derivatization method for the analysis of amino acids in cerebrospinal fluid by gas chromatography-mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2013, 931, 97–102. [Google Scholar] [CrossRef]
- Tang, D.Q.; Zou, L.; Yin, X.X.; Ong, C.N. HILIC-MS for metabolomics: An attractive and complementary approach to RPLC-MS. Mass Spectrom. Rev. 2016, 35, 574–600. [Google Scholar] [CrossRef]
- Teutsch, S.M.; Bradley, L.A.; Palomaki, G.E.; Haddow, J.E.; Piper, M.; Calonge, N.; Dotson, W.D.; Douglas, M.P.; Berg, A.O. The evaluation of genomic applications in practice and prevention (EGAPP) initiative: Methods of the EGAPP working group. Genet. Med. 2009, 11, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Hayes, D.F. Biomarker validation and testing. Mol. Oncol. 2015, 9, 960–966. [Google Scholar] [CrossRef] [PubMed]
- Lima, A.R.; Bastos, M.D.L.; Carvalho, M.; Pinho, P.G. De Biomarker Discovery in Human Prostate Cancer: An Update in Metabolomics Studies. Transl. Oncol. 2016, 9, 357–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Çakır, T.; Khatibipour, M.J. Metabolic Network Discovery by Top-Down and Bottom-Up Approaches and Paths for Reconciliation. Front. Bioeng. Biotechnol. 2014, 2, 62. [Google Scholar] [CrossRef] [Green Version]
- Yu, T.; Bai, Y. Analyzing LC/MS Metabolic Profiling Data in the Context of Existing Metabolic Networks. Curr. Metab. 2013, 1, 84–91. [Google Scholar] [CrossRef] [Green Version]
- Wedge, D.C.; Allwood, J.W.; Dunn, W.; Vaughan, A.A.; Simpson, K.; Brown, M.; Priest, L.; Blackhall, F.H.; Whetton, A.D.; Dive, C.; et al. Is serum or plasma more appropriate for intersubject comparisons in metabolomic studies? An assessment in patients with small-cell lung cancer. Anal. Chem. 2011, 83, 6689–6697. [Google Scholar] [CrossRef]
- Denery, J.R.; Nunes, A.A.K.; Dickerson, T.J. Characterization of differences between blood sample matrices in untargeted metabolomics. Anal. Chem. 2011, 83, 1040–1047. [Google Scholar] [CrossRef]
- Breier, M.; Wahl, S.; Prehn, C.; Fugmann, M.; Ferrari, U.; Weise, M.; Banning, F.; Seissler, J.; Grallert, H.; Adamski, J.; et al. Targeted metabolomics identifies reliable and stable metabolites in human serum and plasma samples. PLoS ONE 2014, 9, 1–11. [Google Scholar] [CrossRef]
- Pinto, J.; Domingues, M.R.M.; Galhano, E.; Pita, C.; Do Céu Almeida, M.; Carreira, I.M.; Gil, A.M. Human plasma stability during handling and storage: Impact on NMR metabolomics. Analyst 2014, 139, 1168–1177. [Google Scholar] [CrossRef]
- López-Bascón, M.A.; Priego-Capote, F.; Peralbo-Molina, A.; Calderón-Santiago, M.; Luque De Castro, M.D. Influence of the collection tube on metabolomic changes in serum and plasma. Talanta 2016, 150, 681–689. [Google Scholar] [CrossRef] [PubMed]
- Zhang, T.; Wu, X.; Yin, M.; Fan, L.; Zhang, H.; Zhao, F.; Zhang, W.; Ke, C.; Zhang, G.; Hou, Y.; et al. Discrimination between malignant and benign ovarian tumors by plasma metabolomic profiling using ultra performance liquid chromatography/mass spectrometry. Clin. Chim. Acta 2012, 413, 861–868. [Google Scholar] [CrossRef]
- Bi, H.; Guo, Z.; Jia, X.; Liu, H.; Ma, L.; Xue, L. The key points in the pre-analytical procedures of blood and urine samples in metabolomics studies. Metabolomics 2020, 16. [Google Scholar] [CrossRef]
- Burton, C.; Shi, H.; Ma, Y. Normalization of urinary pteridines by urine specific gravity for early cancer detection. Clin. Chim. Acta 2014, 435, 42–47. [Google Scholar] [CrossRef] [PubMed]
- Shu, I.; Jones, J.; Jones, M.; Lewis, D.; Negrusz, A. Detection of drugs in nails: Three-year experience. J. Anal. Toxicol. 2015, 39, 624–628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dutkiewicz, E.P.; Urban, P.L. Quantitative mass spectrometry of unconventional human biological matrices. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2016, 374. [Google Scholar] [CrossRef]
- Dereziński, P.; Klupczynska, A.; Sawicki, W.; Pałka, J.A.; Kokot, Z.J. Amino acid profiles of serum and urine in search for prostate cancer biomarkers: A pilot study. Int. J. Med. Sci. 2017, 14, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Synakiewicz, A.; Sawicka-Zukowska, M.; Adrianowska, N.; Galezowska, G.; Ratajczyk, J.; Owczarzak, A.; Konieczna, L.; Stachowicz-Stencel, T. Amino acid profiles as potential biomarkers for pediatric cancers: A preliminary communication. Biomark. Med. 2017, 11, 619–627. [Google Scholar] [CrossRef]
- Synakiewicz, A.; Stanislawska-Sachadyn, A.; Sawicka-Zukowska, M.; Galezowska, G.; Ratajczyk, J.; Owczarzak, A.; Skuza, M.; Wolska, L.; Stachowicz-Stencel, T. Plasma free amino acid profiling as metabolomic diagnostic and prognostic biomarker in paediatric cancer patients: A follow-up study. Amino Acids 2021, 53, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Simińska, E.; Koba, M. Amino acid profiling as a method of discovering biomarkers for early diagnosis of cancer. Amino Acids 2016, 48, 1339–1345. [Google Scholar] [CrossRef] [PubMed]
- Kimball, S.R.; Jefferson, L.S. Role of amino acids in the translational control of protein synthesis in mammals. Semin. Cell Dev. Biol. 2005, 16, 21–27. [Google Scholar] [CrossRef]
- Wei, Z.; Liu, X.; Cheng, C.; Yu, W.; Yi, P. Metabolism of Amino Acids in Cancer. Front. Cell Dev. Biol. 2021, 8, 603837. [Google Scholar] [CrossRef] [PubMed]
- Vučetić, M.; Cormerais, Y.; Parks, S.K.; Pouysségur, J. The central role of amino acids in cancer redox homeostasis: Vulnerability points of the cancer redox code. Front. Oncol. 2017, 7, 319. [Google Scholar] [CrossRef]
- Yang, L.; Wang, Y.; Cai, H.; Wang, S.; Shen, Y.; Ke, C. Application of metabolomics in the diagnosis of breast cancer: A systematic review. J. Cancer 2020, 11, 2540–2551. [Google Scholar] [CrossRef]
- Shin, C.-S.; Mishra, P.; Watrous, J.D.; Carelli, V.; D’Aurelio, M.; Jain, M.; Chan, D.C. The glutamate/cystine xCT antiporter antagonizes glutamine metabolism and reduces nutrient flexibility. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Suhane, S.; Ramanujan, V.K. Thyroid hormone differentially modulates Warburg phenotype in breast cancer cells. Biochem. Biophys. Res. Commun. 2011, 414, 73–78. [Google Scholar] [CrossRef] [Green Version]
- Kus, K.; Kij, A.; Zakrzewska, A.; Jasztal, A.; Stojak, M.; Walczak, M.; Chlopicki, S. Alterations in arginine and energy metabolism, structural and signalling lipids in metastatic breast cancer in mice detected in plasma by targeted metabolomics and lipidomics. Breast Cancer Res. 2018, 20, 1–13. [Google Scholar] [CrossRef]
- Brodsky, A.N.; Odenwelder, D.C.; Harcum, S.W. High extracellular lactate causes reductive carboxylation in breast tissue cell lines grown under normoxic conditions. PLoS ONE 2019, 14, e0213419. [Google Scholar] [CrossRef] [PubMed]
- Gkiouli, M.; Biechl, P.; Eisenreich, W.; Otto, A.M. Diverse Roads Taken by 13C-Glucose-Derived Metabolites in Breast Cancer Cells Exposed to Limiting Glucose and Glutamine Conditions. Cells 2019, 8, 1113. [Google Scholar] [CrossRef] [Green Version]
- Leij-Halfwerk, S.; Dagnelie, P.C.; Van Den Berg, J.W.O.; Wattimena, J.D.L.; Hordijk-Luijk, C.H.; Wilson, J.H.P. Weight loss and elevated gluconeogenesis from alanine in lung cancer patients. Am. J. Clin. Nutr. 2000, 71, 583–589. [Google Scholar] [CrossRef] [Green Version]
- Von Mach-Szczypiński, J.; Stanosz, S.; Sieja, K.; Stanosz, M. Metabolism of histamine in tissues of primary ductal breast cancer. Metabolism 2009, 58, 867–870. [Google Scholar] [CrossRef]
- Engel, N.; Cricco, G.; Davio, C.; Martin, G.; Croci, M.; Fitzsimons, C.; Bergoc, R.; Rivera, E. Histamine regulates the expression of histidine decarboxylase in N-nitroso-N-methylurea (NMU)-induced mammary tumors in rats. Inflamm. Res. 1996, 45, S64–S65. [Google Scholar] [CrossRef] [PubMed]
- Falus, A.; Grosman, N.; Darvas, Z. Histamine: Biology and Medical Aspects; SpringMed Pub: Budapest, Hungary, 2004; pp. 199–217. [Google Scholar]
- Hitosugi, T.; Fan, J.; Chung, T.W.; Lythgoe, K.; Wang, X.; Xie, J.; Ge, Q.; Gu, T.L.; Polakiewicz, R.D.; Roesel, J.L.; et al. Tyrosine Phosphorylation of Mitochondrial Pyruvate Dehydrogenase Kinase 1 Is Important for Cancer Metabolism. Mol. Cell 2011, 44, 864–877. [Google Scholar] [CrossRef] [Green Version]
- Fan, J.; Hitosugi, T.; Chung, T.-W.; Xie, J.; Ge, Q.; Gu, T.-L.; Polakiewicz, R.D.; Chen, G.Z.; Boggon, T.J.; Lonial, S.; et al. Tyrosine Phosphorylation of Lactate Dehydrogenase A Is Important for NADH/NAD+ Redox Homeostasis in Cancer Cells. Mol. Cell. Biol. 2011, 31, 4938–4950. [Google Scholar] [CrossRef] [Green Version]
- Belladonna, M.L.; Puccetti, P.; Orabona, C.; Fallarino, F.; Vacca, C.; Volpi, C.; Gizzi, S.; Pallotta, M.T.; Fioretti, M.C.; Grohmann, U. Immunosuppression via tryptophan catabolism: The role of kynurenine pathway enzymes. Transplantation 2007, 84, S17–S20. [Google Scholar] [CrossRef]
- Radu, C.A.; Bosch, N.; Bauer, T.M.; Kleist, C.; Jiga, L.; Terness, P.; Opelz, G.; Gebhard, M.M.; Germann, G.; Baumeister, S. Immunosuppressive effect of tryptophan metabolites in composite tissue allotransplantation. Plast. Reconstr. Surg. 2007, 119, 2023–2028. [Google Scholar] [CrossRef]
- Geeganage, S.; Sams, L.; Henry, J.; Dorsey, F.; Roth, K.; Nikolayev, A.; Benhadji, K.; Gilmour, R.; Cerezo, A.; Peregrina, S.; et al. Abstract 2187: Tryptophan Metabolism Plays a Central Role in Immunosuppression. Am. Assoc. Cancer Res. (AACR) 2019, 79, 2187. [Google Scholar]
- Kang, J.S. Dietary restriction of amino acids for Cancer therapy. Nutr. Metab. 2020, 17, 1–12. [Google Scholar] [CrossRef]
- Popovic, P.J.; Zeh, H.J.; Ochoa, J.B. Arginine and immunity. J. Nutr. 2007, 137, 1681–1686. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Fan, J.; Venneti, S.; Cross, J.R.; Takagi, T.; Bhinder, B.; Djaballah, H.; Kanai, M.; Cheng, E.H.; Judkins, A.R.; et al. Asparagine plays a critical role in regulating cellular adaptation to glutamine depletion. Mol. Cell 2014, 56, 205–218. [Google Scholar] [CrossRef] [Green Version]
- Bonifácio, V.D.B.; Pereira, S.A.; Serpa, J.; Vicente, J.B. Cysteine metabolic circuitries: Druggable targets in cancer. Br. J. Cancer 2020, 124, 124. [Google Scholar]
- Felig, P.; Pozefsk, T.; Cahill, G.F. Alanine: Key Role in Gluconeogenesis. Science 1970, 167, 7–9. [Google Scholar] [CrossRef]
- Liberti, M.V.; Locasale, J.W. The Warburg Effect: How Does it Benefit Cancer Cells? Trends Biochem. Sci. 2016, 41, 211–218. [Google Scholar] [CrossRef] [Green Version]
- Sato, M.; Kawana, K.; Adachi, K.; Fujimoto, A.; Yoshida, M.; Nakamura, H.; Nishida, H.; Inoue, T.; Taguchi, A.; Takahashi, J.; et al. Spheroid cancer stem cells display reprogrammed metabolism and obtain energy by actively running the tricarboxylic acid (TCA) cycle. Oncotarget 2016, 7, 33297–33305. [Google Scholar] [CrossRef] [Green Version]
- González-Domínguez, R.; González-Domínguez, Á.; Sayago, A.; Fernández-Recamales, Á. Recommendations and best practices for standardizing the pre-analytical processing of blood and urine samples in metabolomics. Metabolites 2020, 10, 229. [Google Scholar] [CrossRef] [PubMed]
- Krall, A.S.; Xu, S.; Graeber, T.G.; Braas, D.; Christofk, H.R. Asparagine promotes cancer cell proliferation through use as an amino acid exchange factor. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.; Ko, B.; Hensley, C.T.; Jiang, L.; Wasti, A.T.; Kim, J.; Sudderth, J.; Calvaruso, M.A.; Lumata, L.; Mitsche, M.; et al. Glutamine oxidation maintains the TCA cycle and cell survival during impaired mitochondrial pyruvate transport. Mol. Cell 2014, 56, 414–424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, H.; Vandekeere, S.; Kalucka, J.; Bierhansl, L.; Zecchin, A.; Brüning, U.; Visnagri, A.; Yuldasheva, N.; Goveia, J.; Cruys, B.; et al. Role of glutamine and interlinked asparagine metabolism in vessel formation. EMBO J. 2017, 36, 2334–2352. [Google Scholar] [CrossRef]
- Luo, S.; Levine, R.L. Methionine in proteins defends against oxidative stress. FASEB J. 2009, 23, 464–472. [Google Scholar] [CrossRef] [Green Version]
- Fratoni, V.; Brandi, M.L. B vitamins, Homocysteine and bone health. Nutrients 2015, 7, 2176–2192. [Google Scholar] [CrossRef] [Green Version]
- Clare, C.E.; Brassington, A.H.; Kwong, W.Y.; Sinclair, K.D. One-Carbon Metabolism: Linking Nutritional Biochemistry to Epigenetic Programming of Long-Term Development. Annu. Rev. Anim. Biosci. 2019, 7, 263–287. [Google Scholar] [CrossRef]
- Schmandt, R.E.; Iglesias, D.A.; Co, N.N.; Lu, K.H. Understanding obesity and endometrial cancer risk: Opportunities for prevention. Am. J. Obstet. Gynecol. 2011, 205, 518–525. [Google Scholar] [CrossRef] [Green Version]
- Onstad, M.A.; Schmandt, R.E.; Lu, K.H. Addressing the role of obesity in endometrial cancer risk, prevention, and treatment. J. Clin. Oncol. 2016, 34, 4225–4230. [Google Scholar] [CrossRef] [PubMed]
- Beavis, A.L.; Smith, A.J.B.; Fader, A.N. Lifestyle changes and the risk of developing endometrial and ovarian cancers: Opportunities for prevention and management. Int. J. Womens Health 2016, 8, 151–167. [Google Scholar]
- Amelio, I.; Cutruzzolá, F.; Antonov, A.; Agostini, M.; Melino, G. Serine and glycine metabolism in cancer. Trends Biochem. Sci. 2014, 39, 191–198. [Google Scholar] [CrossRef]
- Rossi Fanelli, F.; Cangiano, C.; Ceci, F.; Cellerino, R.; Franchi, F.; Menichetti, E.T.; Muscaritoli, M.; Cascino, A. Plasma tryptophan and anorexia in human cancer. Eur. J. Cancer Clin. Oncol. 1986, 22, 89–95. [Google Scholar] [CrossRef]
- Muscaritoli, M.; Conversano, L.; Petti, M.C.; Torelli, G.F.; Cascino, A.; Mecarocci, S.; Annicchiarico, M.A.; Rossi Fanelli, F. Plasma amino acid concentrations in patients with acute myelogenous leukemia. Nutrition 1999, 15, 195–199. [Google Scholar] [CrossRef]
- Cangiano, C.; Cascino, A.; Ceci, F.; Laviano, A.; Mulieri, M.; Muscaritoli, M.; Rossi-Fanelli, F. Plasma and CSF tryptophan in cancer anorexia. J. Neural Transm. Gen. Sect. 1990, 81, 225–233. [Google Scholar] [CrossRef] [PubMed]
- Fallarino, F.; Grohmann, U.; Vacca, C.; Bianchi, R.; Orabona, C.; Spreca, A.; Fioretti, M.C.; Puccetti, P. T cell apoptosis by tryptophan catabolism. Cell Death Differ. 2002, 9, 1069–1077. [Google Scholar] [CrossRef]
- Badawy, A.A.-B. Targeting tryptophan availability to tumors: The answer to immune escape? Immunol. Cell Biol. 2018, 96, 1026–1034. [Google Scholar] [CrossRef]
- Medina, V.A.; Rivera, E.S. Histamine receptors and cancer pharmacology. Br. J. Pharmacol. 2010, 161, 755–767. [Google Scholar] [CrossRef] [Green Version]
- Cory, J.G.; Cory, A.H. Critical roles of glutamine as nirogen donors in purine and pyrimidine nucleotide synthesis: Asparaginase treatment in childhood acute lymphoblastic leukemia. In Vivo 2006, 20, 587–590. [Google Scholar] [PubMed]
- Eagle, H. Nutrition needs of mammalian cells in tissue culture. Science 1955, 122, 501–504. [Google Scholar] [CrossRef]
- Yang, J.S.; Wang, C.C.; Qiu, J.D.; Ren, B.; You, L. Arginine metabolism: A potential target in pancreatic cancer therapy. Chin. Med. J. 2020, 134, 28–37. [Google Scholar] [CrossRef]
- Miolo, G.; Muraro, E.; Caruso, D.; Crivellari, D.; Ash, A.; Scalone, S.; Lombardi, D.; Rizzolio, F.; Giordano, A.; Corona, G. Phamacometabolomics study identifies circulating spermidine and tryptophan as potential biomarkers associated with the complete pathological response to trastuzumab-paclitaxel neoadjuvant therapy in HER-2 positive breast cancer. Oncotarget 2016, 7, 39809–39822. [Google Scholar] [CrossRef] [Green Version]
- Pusapati, R.V.; Daemen, A.; Wilson, C.; Sandoval, W.; Gao, M.; Haley, B.; Baudy, A.R.; Hatzivassiliou, G.; Evangelista, M.; Settleman, J. mTORC1-Dependent Metabolic Reprogramming Underlies Escape from Glycolysis Addiction in Cancer Cells. Cancer Cell 2016, 29, 548–562. [Google Scholar] [CrossRef] [Green Version]
- Caneba, C.A.; Bellance, N.; Yang, L.; Pabst, L.; Nagrath, D. Pyruvate uptake is increased in highly invasive ovarian cancer cells under anoikis conditions for anaplerosis, mitochondrial function, and migration. Am. J. Physiol. Endocrinol. Metab. 2012, 303, 1036–1052. [Google Scholar] [CrossRef] [Green Version]
- Caneba, C.A.; Yang, L.; Baddour, J.; Curtis, R.; Win, J.; Hartig, S.; Marini, J.; Nagrath, D. Nitric oxide is a positive regulator of the Warburg effect in ovarian cancer cells. Cell Death Dis. 2014, 5, e1302. [Google Scholar] [CrossRef] [PubMed]
- Phang, J.M.; Liu, W.; Hancock, C.N.; Fischer, J.W. Proline metabolism and cancer: Emerging links to glutamine and collagen. Curr. Opin. Clin. Nutr. Metab. Care 2015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, X.; Zhang, L.; Natarajan, S.K.; Becker, D.F. Proline Mechanisms of Stress Survival. Antioxid. Redox Signal. 2013, 19, 998–1011. [Google Scholar] [CrossRef] [Green Version]
- Tanner, J.J.; Fendt, S.M.; Becker, D.F. The Proline Cycle as a Potential Cancer Therapy Target. Biochemistry 2018, 57, 3433–3444. [Google Scholar] [CrossRef]
- Palka, J.; Oscilowska, I.; Szoka, L. Collagen metabolism as a regulator of proline dehydrogenase/proline oxidase-dependent apoptosis/autophagy. Amino Acids 2021. [Google Scholar] [CrossRef] [PubMed]
- Phang, J.M.; Yeh, G.C.; Hagedorn, C.H. The intercellular proline cycle. Life Sci. 1981, 28, 53–58. [Google Scholar] [CrossRef]
- Adams, E.; Frank, L. Metabolism of proline and the hydroxyprolines. Annu. Rev. Biochem. 1980, 49, 1005–1061. [Google Scholar] [CrossRef]
- Phang, J.M.; Pandhare, J.; Liu, Y. The metabolism of proline as microenvironmental stress substrate. J. Nutr. 2008, 138, 2008S–2015S. [Google Scholar] [CrossRef]
- Mock, W.L.; Green, P.C.; Boyer, K.D. Specificity and pH dependence for acylproline cleavage by prolidase. J. Biol. Chem. 1990, 265, 19600–19605. [Google Scholar] [CrossRef]
- Zbucka, M.; Miltyk, W.; Bielawski, T.; Surazynski, A.; Palka, J.; Wolczynski, S. Mechanism of collagen biosynthesis up-regulation in cultured leiomyoma cells. Folia Histochem. Cytobiol. 2007, 45, 181–185. [Google Scholar] [CrossRef]
- Phang, J.M. Proline metabolism in cell regulation and cancer biology: Recent advances and hypotheses. Antioxid. Redox Signal. 2019, 30, 635–649. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Le, A.; Hancock, C.; Lane, A.N.; Dang, C.V.; Fan, T.W.M.; Phang, J.M. Reprogramming of proline and glutamine metabolism contributes to the proliferative and metabolic responses regulated by oncogenic transcription factor c-MYC. Proc. Natl. Acad. Sci. USA 2012, 109, 8983–8988. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Hancock, C.N.; Fischer, J.W.; Harman, M.; Phang, J.M. Proline biosynthesis augments tumor cell growth and aerobic glycolysis: Involvement of pyridine nucleotides. Sci. Rep. 2015, 5, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Surazynski, A.; Donald, S.P.; Cooper, S.K.; Whiteside, M.A.; Salnikow, K.; Liu, Y.; Phang, J.M. Extracellular matrix and HIF-1 signaling: The role of prolidase. Int. J. Cancer 2008, 122, 1435–1440. [Google Scholar] [CrossRef]
- Phang, J.M.; Liu, W.; Hancock, C.; Christian, K.J. The proline regulatory axis and cancer. Front. Oncol. 2012, 2, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Borchert, G.L.; Donald, S.P.; Diwan, B.A.; Anver, M.; Phang, J.M. Proline oxidase functions as a mitochondrial tumor suppressor in human cancers. Cancer Res. 2009, 69, 6414–6422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoque, M.A.; Banu, M.N.A.; Nakamura, Y.; Shimoishi, Y.; Murata, Y. Proline and glycinebetaine enhance antioxidant defense and methylglyoxal detoxification systems and reduce NaCl-induced damage in cultured tobacco cells. J. Plant Physiol. 2008, 165, 813–824. [Google Scholar] [CrossRef]
- Elia, I.; Broekaert, D.; Christen, S.; Boon, R.; Radaelli, E.; Orth, M.F.; Verfaillie, C.; Grünewald, T.G.P.; Fendt, S.M. Proline metabolism supports metastasis formation and could be inhibited to selectively target metastasizing cancer cells. Nat. Commun. 2017, 8, 1–11. [Google Scholar] [CrossRef]
- Craze, M.L.; Cheung, H.; Jewa, N.; Coimbra, N.D.M.; Soria, D.; El-Ansari, R.; Aleskandarany, M.A.; Wai Cheng, K.; Diez-Rodriguez, M.; Nolan, C.C.; et al. MYC regulation of glutamine-proline regulatory axis is key in luminal B breast cancer. Br. J. Cancer 2018, 118, 258–265. [Google Scholar] [CrossRef] [Green Version]
- Ding, J.; Kuo, M.L.; Su, L.; Xue, L.; Luh, F.; Zhang, H.; Wang, J.; Lin, T.G.; Zhang, K.; Chu, P.; et al. Human mitochondrial pyrroline-5-carboxylate reductase 1 promotes invasiveness and impacts survival in breast cancers. Carcinogenesis 2017, 38, 519–531. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reference | Disease | Design | Matrix | Method | Strategy | Differentiating Amino Acids | Validation 1 |
|---|---|---|---|---|---|---|---|
| Ihata et al., 2014 [9] | Endometrial cancer | Endometrial cancer (n = 80); Gynecological benign diseases (n = 122); Healthy controls (n = 240) | Plasma | HPLC-MS/MS | Targeted | Asparagine (↑), glutamine (↑), histidine (↓), isoleucine (↑), leucine (↓), methionine (↓), ornithine (↑), phenylalanine (↓), proline (↑), serine (↓), tryptophan (↓), valine (↓) | Yes |
| Troisi et al., 2018 [10] | Endometrial cancer | Healthy subjects (n = 130); Endometrial Cancer (n = 118); Ovarian Cancer (n = 30); Benign endometrial disease (n = 10) | Serum | GC-MS | Untargeted | Homocysteine (↑), threonine (↓), valine (↓) | Yes |
| Gaudet et al., 2012 [11] | Endometrial cancer | Endometrial cancer(n = 250); Controls (n = 250) | Serum | FIA-MS/MS | Targeted | Valine (↑) | No |
| Shi et al., 2018 [12] | Endometrial cancer | Endometrial cancer (n = 46); Healthy controls (n = 46) | Serum | UHPLC-MS | Untargeted | Phenylalanine (↑) | No |
| Suzuki et al., 2018 [13] | Endometrial cancer | Endometrial cancer (n = 53); pre-surgery vs. post-surgery | Plasma | HPLC-MS/MS | Targeted | Citrulline (↓), histidine (↓), isoleucine (↑), tryptophan (↓), valine (↓) | No |
| Bahado-Singh et al., 2017 [14] | Endometrial cancer | Endometrial cancer (n = 56); Controls (n = 60) | Serum | 1H-NMR; LC-MS/MS; FIA-MS/MS | Untargeted and targeted | Asparagine (↓), glutamate (↑), methionine (↓) | Yes |
| Audet-Delage et al., 2018 [15] | Endometrial cancer | Control women (n = 18); Type I endometrioid (n = 24); Type II serous carcinomas (n = 12) | Serum | UHPLC-MS/MS | Untargeted | Glycine (↓) | No |
| Strand et al., 2019 [16] | Endometrial cancer | Short survival (n = 20); Long survival (n = 20) | Plasma | LC-MS/MS; FIA-MS/MS | Targeted | Methionine sulfoxide (↓) | No |
| Altadill et al., 2017 [17] | Endometrial cancer | Endometrial cancer (n = 39); Healthy controls (n = 17) | Tissue | UHPLC-MS | Untargeted | Arginine (↓), glutamate (↓), phenylalanine (↓), tryptophan (↓) | Yes |
| Trousil et al., 2014 [18] | Endometrial cancer | Control group (n = 10); Endometrial cancer (n = 8) | Tissue | 1H-NMR | Untargeted | Alanine (↑), leucine (↑), proline (↑), valine (↑), tyrosine (↑) | No |
| Cheng et al., 2019 [19] | Endometrial cancer | Endometrial cancer (n = 21); non-endometrial cancer (n = 33) | Cervicovaginal fluid | 1H-NMR | Untargeted | Aspartate (↓), asparagine (↓), isoleucine (↓), phenylalanine (↓) | Yes |
| Zhou et al., 2010 [20] | Ovarian cancer | Ovarian cancer (n = 44); Healthy women or with benign condition (n = 50) | Serum | DART-MS | Untargeted | Alanine (↑), cystine (↑), glycine (↑), serine (↑), threonine (↑) | No |
| Hilvo et al., 2015 [21] | Ovarian cancer | Ovarian cancer (high grade) (n = 158); Benign ovarian tumors and healthy control (n = 100) | Serum, tissue | GC-MS | Untargeted | Alanine (↓), glutamate (↑), glutamine (↑), glycine (↑), methionine (↓), phenylalanine (↓), proline (↓), serine (↓), threonine (↓), tryptophan (↓), tyrosine (↓), valine (↓) | Yes |
| Garcia et al., 2011 [22] | Ovarian cancer | Ovarian cancer (early stage FIGO I/II) (n = 170); Healthy controls (n = 182) | Serum | 1H-NMR | Untargeted | Alanine (↓), valine (↓) | Yes |
| Bachmayr-Heyda et al., 2017 [23] | Ovarian cancer | Ovarian cancer (high-grade serous) (n = 65); Healthy controls (n = 62) | Serum, ascites, tissue | LC-MS/MS; FIA-MS/MS | Targeted | Asparagine (↓), histidine (↓), lysine (↓), threonine (↓), tryptophan (↓) | Yes |
| Buas et al., 2016 [24] | Ovarian cancer | Ovarian cancer (serous) (n = 50); Benign ovarian tumors (serous) (n = 50) | Plasma | HPLC-MS | Targeted | Alanine (↓) | No |
| Ke et al., 2014 [25] | Ovarian cancer | Ovarian cancer (n = 140); Benign ovarian tumors/uterine fibromas (n = 308) | Plasma | UHPLC-MS | Untargeted | Histidine (↓), lysine (↓), phenylalanine (↓), tryptophan (↓) | Validation of previous research |
| Miyagi et al., 2017 [26] | Ovarian cancer | Ovarian cancer + borderline tumors (n = 80); Benign ovarian tumors (n = 97) | Plasma | HPLC-MS/MS | Targeted | Histidine (↓), isoleucine (↑), proline (↑), tryptophan (↓) | Validation of previous research |
| Zhang et al., 2012 [27] | Ovarian cancer | Ovarian cancer (n = 80); Benign ovarian tumors (n = 90) | Urine | UHPLC-MS | Untargeted | Tryptophan (↓) | Yes |
| Horala et al., 2021 [28] | Ovarian cancer | Ovarian cancer + borderline tumors (n = 44); Benign ovarian tumors (n = 62) | Serum | HPLC-MS/MS | Targeted | Aminoadipic acid (↓), asparagine (↓), citrulline (↓), cystine (↓), glutamine (↑), histidine (↓), isoleucine (↑), leucine (↑), phenylalanine (↑), threonine (↓), tryptophan (↓) | No |
| Plewa et al., 2017 [29] | Ovarian cancer | Ovarian cancer (n = 38); Benign ovarian tumors (n = 62); Healthy controls (n = 50) | Serum | HPLC-MS/MS | Targeted | Citrulline (↓), histidine (↓), lysine (↓), phenylalanine (↓), threonine (↓), tryptophan (↓) | No |
| Plewa et al., 2019 [30] | Ovarian cancer | Ovarian cancer (n = 26); Benign ovarian tumors (n = 25); Healthy controls (n = 25) | Serum | HPLC-MS/MS | Targeted | Citrulline (↓), histidine (↓) | Validation of previous research |
| Slupsky et al., 2010 [31] | Ovarian cancer | Ovarian cancer (n = 40); Healthy controls (n = 62) | Urine | 1H-NMR | Untargeted | Alanine (↓), asparagine (↓), isoleucine (↓), leucine (↓), valine (↓) | No |
| Ahn et al., 2020 [32] | Ovarian cancer | Ovarian cancer (n = 10); Healthy controls (n = 10) | Plasma | UHPLC-MS/MS; FIA-MS/MS | Targeted | Ornithine (↓), tryptophan (↓) | No |
| Wang et al., 2021 [33] | Ovarian cancer | Ovarian cancer (n = 39); Healthy controls (n = 31) | Serum | UHPLC-MS/MS | Targeted | Asparagine (↑), glutamine (↑), methionine (↑) | Yes |
| His et al., 2019 [34] | Breast cancer | Invasive breast cancer (n = 1624); Control group (n = 1624) | Plasma | LC-MS/MS; FIA-MS/MS | Targeted | Arginine (↓), asparagine (↓) | No |
| Eniu et al., 2018 [35] | Breast cancer | Breast cancer (n = 30); Healthy controls (n = 26) | Serum | UHPLC-MS | Targeted | Alanine (↓), arginine (↓), glutamine (↓), isoleucine (↓), leucine (↓), tyrosine (↓) | No |
| Mitruka et al., 2020 [36] | Breast cancer | Breast cancer (n = 10); Healthy controls (n = 12) | Nails | HPLC-MS | Untargeted | Histidine (↓), phenylalanine (↓), tryptophan (↓), tyrosine (↓) | No |
| Shen et al., 2013 [37] | Breast cancer | Breast cancer (n = 60); Healthy controls (n = 60) | Plasma | UHPLC-MS/MS; GC-MS | Untargeted | Alanine (↓), glutamine (↓), histidine (↓), methionine (↓), proline (↓) | No |
| Budczies et al., 2013 [38] | Breast cancer | Estrogen receptor positive (ER+) (n = 204); Estrogen receptor negative (ER−) (n = 67) | Tissue | GC-MS | Untargeted | Alanine (↑), glutamate (↑), glutamine (↓) | Yes |
| Cala et al., 2018 [39] | Breast cancer | Breast cancer (n = 29); Healthy controls (n = 29) | Plasma | GC–MS; LC-MS; 1H-NMR | Untargeted | Alanine (↑), cystine (↓), isoleucine (↓), threonine (↓), tryptophan (↓) | No |
| Miyagi et al., 2011 [40] | Breast cancer | Breast cancer (n = 196); Healthy controls (n = 976) | Plasma | HPLC-MS/MS | Targeted | Alanine (↑), glutamine (↓), glycine (↑), histidine (↓), ornithine (↑), phenylalanine (↓), proline (↑), serine (↑), tryptophan (↓), tyrosine (↓) | No |
| Moore et al., 2021 [41] | Breast cancer | Breast cancer (n = 782); Control group (n = 782) | Serum | UHPLC-MS | Untargeted | Cystine (↑) | Validation of previous research |
| Cao et al., 2015 [42] | Breast cancer | Breast cancer (n = 20); Healthy controls (n = 50) | Serum | FIA-MS/MS | Targeted | Tryptophan (↑) | No |
| Xie et al., 2015 [43] | Breast cancer | Breast cancer (n = 35); Control group (n = 35) | Plasma, serum, tissue | HPLC-MS; GC-MS | Untargeted | Aspartate (↓) | Yes |
| Wang et al., 2016 [44] | Breast cancer | Breast cancer (n = 258); Control group (n = 159) | Dried blood spot | DIMS | Targeted | Asparagine (↓), cystine (↓), histidine (↓), homocysteine (↓), lysine (↓), proline (↓), tyrosine (↑), tryptophan (↓) | Yes |
| Jasbi et al., 2018 [45] | Breast cancer | Breast cancer (n = 102); Healthy controls (n = 99) | Plasma | UHPLC-MS/MS | Targeted | Proline (↓) | No |
| Yuan et al., 2018 [46] | Breast cancer | Breast cancer (n = 80); Healthy controls (n = 100) | Plasma | LC-MS/MS; FIA-MS/MS | Targeted | Alanine (↓), asparagine (↓), glutamine (↓), histidine (↓), leucine (↓), lysine (↓), methionine (↓), ornithine (↓), phenylalanine (↓), threonine (↓), tryptophan (↓), tyrosine (↓), valine (↓) | Yes |
| Li et al., 2020 [47] | Breast cancer | Breast cancer (n = 31); Healthy controls (n = 31) | Serum | HPLC-MS | Untargeted | Leucine (↑), proline (↑), threonine (↑), tyrosine (↑), valine (↑) | Yes |
| Khan et al., 2019 [48] | Cervical cancer | Cervical cancer (n = 60); Healthy controls (n = 69); CIN1 (n = 55); CIN2/3 (n = 42) | Plasma | UHPLC-MS | Untargeted | Aspartate (↑), glutamate (↑), proline (↑) | No |
| Abudula et al., 2020 [49] | Cervical cancer | Negative controls (n = 11); Cervical cancer (n = 21) | Tissue | 1H-NMR | Untargeted | Alanine (↓), isoleucine (↓), methylproline (↓), phenylalanine (↓), tyrosine (↓) | Yes |
| Yang et al., 2017 [50] | Cervical cancer | Negative controls (n = 149); Cervical cancer (n = 136) | Plasma | UHPLC-MS | Untargeted | Lysine (↓) | No |
| Chen et al., 2013 [51] | Cervical cancer | Negative control (n = 23); Cervical cancer (n = 22) | Urine | LC-MS/MS | Untargeted | Tryptophan (↓), tyrosine (↓) | No |
| Hasim et al., 2012 [52] | Cervical cancer | Negative control (n = 38); Cervical cancer (n = 38) | Plasma | 1H-NMR | Untargeted | Alanine (↓), isoleucine (↓), leucine (↓), valine (↓) | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pietkiewicz, D.; Klupczynska-Gabryszak, A.; Plewa, S.; Misiura, M.; Horala, A.; Miltyk, W.; Nowak-Markwitz, E.; Kokot, Z.J.; Matysiak, J. Free Amino Acid Alterations in Patients with Gynecological and Breast Cancer: A Review. Pharmaceuticals 2021, 14, 731. https://doi.org/10.3390/ph14080731
Pietkiewicz D, Klupczynska-Gabryszak A, Plewa S, Misiura M, Horala A, Miltyk W, Nowak-Markwitz E, Kokot ZJ, Matysiak J. Free Amino Acid Alterations in Patients with Gynecological and Breast Cancer: A Review. Pharmaceuticals. 2021; 14(8):731. https://doi.org/10.3390/ph14080731
Chicago/Turabian StylePietkiewicz, Dagmara, Agnieszka Klupczynska-Gabryszak, Szymon Plewa, Magdalena Misiura, Agnieszka Horala, Wojciech Miltyk, Ewa Nowak-Markwitz, Zenon J. Kokot, and Jan Matysiak. 2021. "Free Amino Acid Alterations in Patients with Gynecological and Breast Cancer: A Review" Pharmaceuticals 14, no. 8: 731. https://doi.org/10.3390/ph14080731