Immunomodulatory Effects of Recombinant Mycobacterium smegmatis Expressing Antigen-85B Epitopes in Infected J774A.1 Murine Macrophages


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
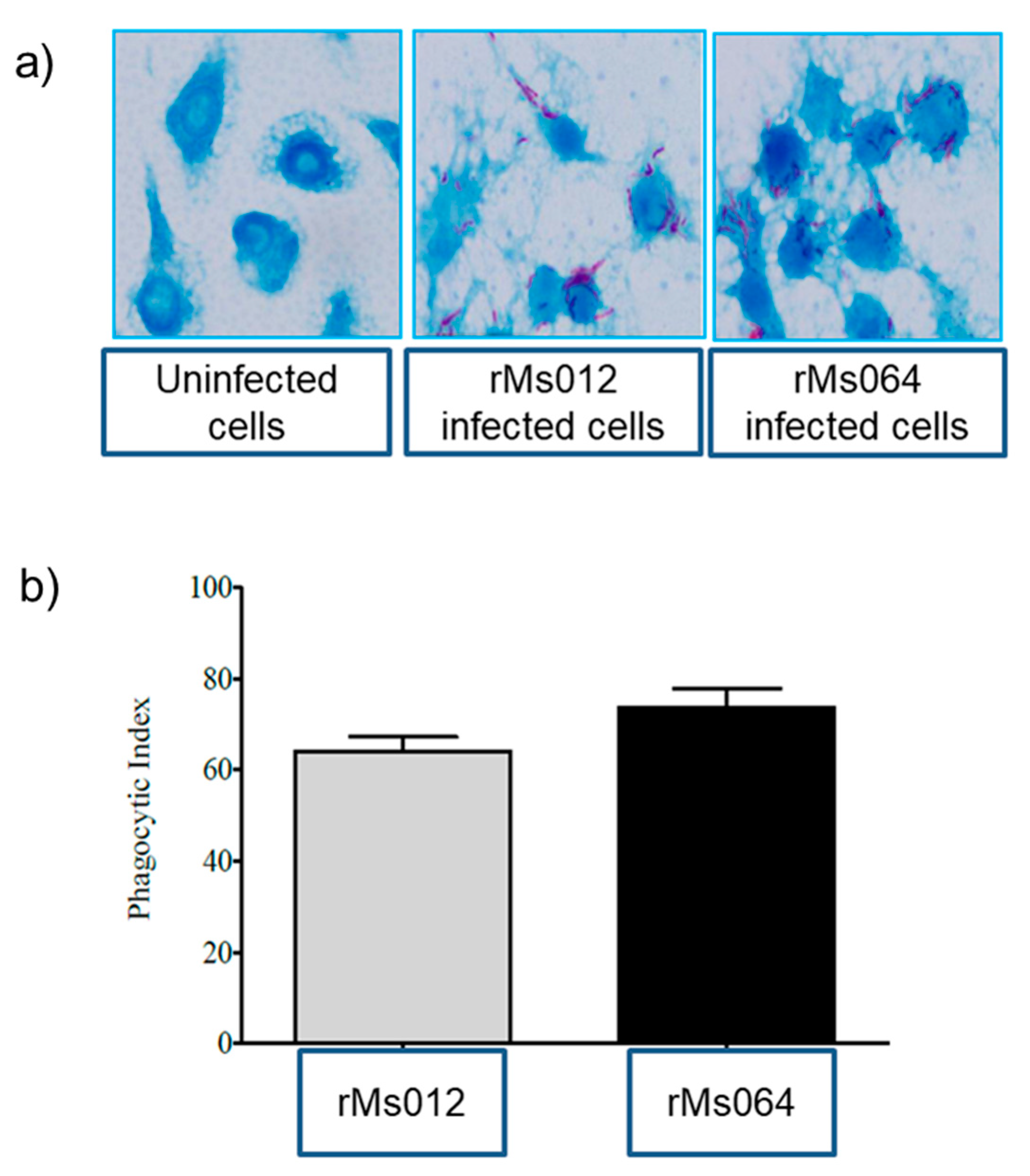
2.1. Phagocytic Uptake of rMs064 and rMs012 by J7474A.1 Macrophages
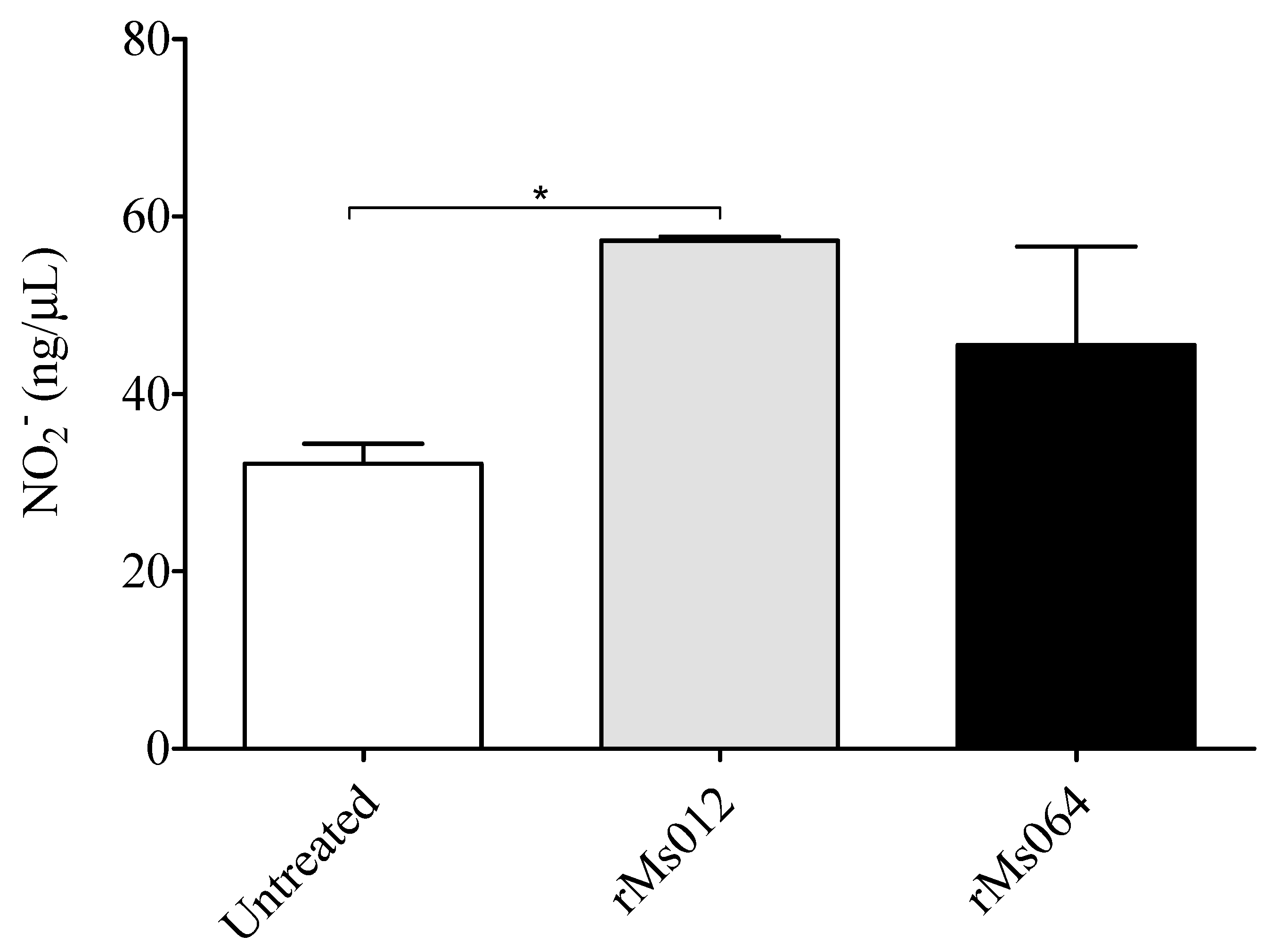
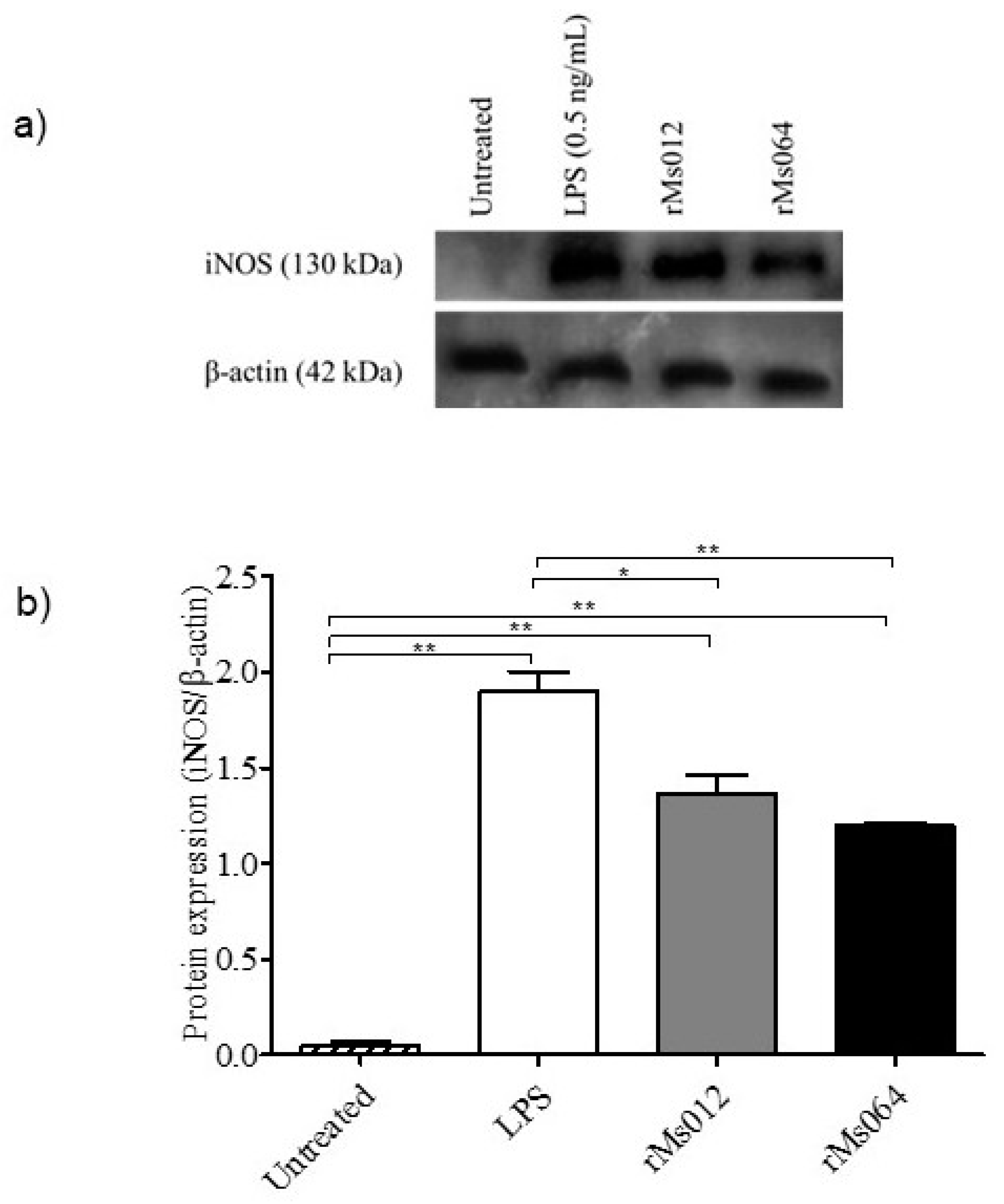
2.2. Production of Nitric Oxide and Inducible Forms of Nitric Oxide Synthase (iNOS) by Infected Macrophages
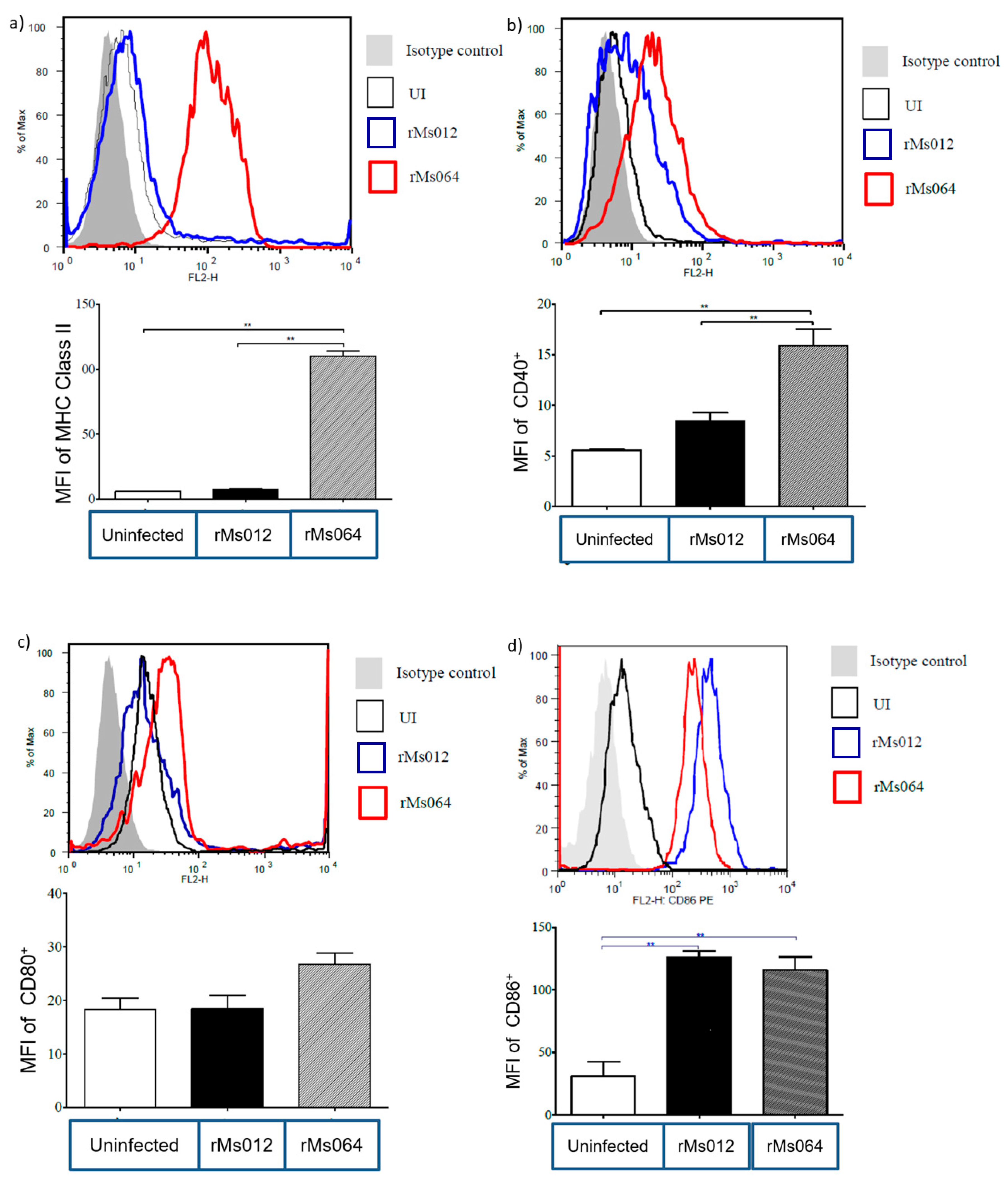
2.3. Macrophage Surface Molecule Expression
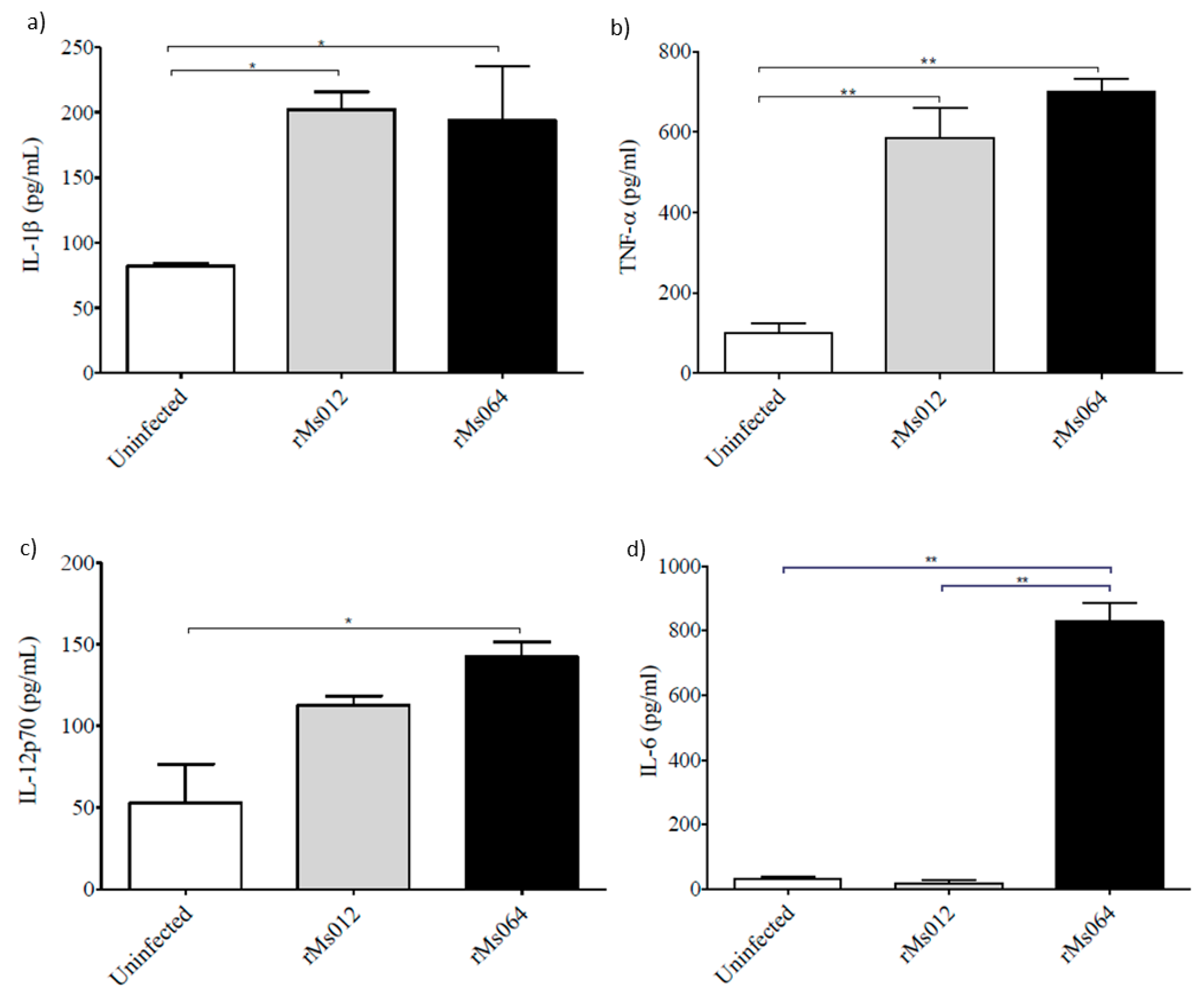
2.4. Cytokine Production
3. Discussion
4. Materials and Methods
4.1. Macrophage Cell Line
4.2. Mycobacterium Smegmatis Strains and Infection Procedure
4.3. Phagocytic Assay
4.4. Determination of NO2 Concentration and iNOS Assay
4.5. Immunofluorescent Staining of Cells
4.6. Cytokines Assay
4.7. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization. Global Tuberculosis Report 2019; WHO: Geneva, Switzerland, 2019. [Google Scholar]
- Sweeney, K.A.; Dao, D.N.; Goldberg, M.F.; Hsu, T.; Venkataswamy, M.M.; Henao-Tamayo, M.; Ordway, D.; Sellers, R.S.; Jain, P.; Chen, B. A recombinant Mycobacterium smegmatis induces potent bactericidal immunity against Mycobacterium tuberculosis. Nat. Med. 2011, 17, 1261–1268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junqueira-Kipnis, A.P.; De Oliveira, F.M.; Trentini, M.M.; Tiwari, S.; Chen, B.; Resende, D.P.; Silva, B.D.; Chen, M.; Tesfa, L.; Jacobs, W.R., Jr. Prime-boost with Mycobacterium smegmatis recombinant vaccine improves protection in mice infected with Mycobacterium tuberculosis. PLoS ONE 2013, 8, e78639. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Peng, P.; Miao, S.; Zhao, Y.; Mao, F.; Wang, L.; Bai, Y.; Xu, Z.; Wei, S.; Shi, C. Recombinant Mycobacterium smegmatis Expressing an ESAT6-CFP10 Fusion Protein Induces Anti-Mycobacterial Immune Responses and Protects Against Mycobacterium tuberculosis Challenge in Mice. Scand. J. Immunol. 2010, 72, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Nguyen Thi, L.T.; Borrero Maura, R.; Férnandez, S.; Reyes, G.; Perez, J.L.; Reyes, F.; de los Angeles García, M.; Fariñas, M.; Infante, J.F.; Tirado, Y.; et al. Evaluation of the potential of Mycobacterium smegmatis as vaccine candidate against tuberculosis by in silico and in vivo studies. VacciMonitor 2010, 19, 20–26. [Google Scholar]
- García, M.d.l.A.; Borrero, R.; Lanio, M.E.; Tirado, Y.; Alvarez, N.; Puig, A.; Aguilar, A.; Canet, L.; Mata Espinoza, D.; Barrios Payán, J. Protective effect of a lipid-based preparation from Mycobacterium smegmatis in a murine model of progressive pulmonary tuberculosis. BioMed Res. Int. 2014, 2014, 1–6. [Google Scholar]
- Lindsey, D.R.; Dhandayuthapani, S.; Jagannath, C. Anti-tuberculosis immunity induced in mice by vaccination with Mycobacterium smegmatis over-expressing Antigen 85B is due to the increased influx of IFNγ-positive CD4 T cells into the lungs. Tuberculosis 2009, 89, S46–S48. [Google Scholar] [CrossRef] [Green Version]
- Tirado, Y.; Puig, A.; Alvarez, N.; Borrero, R.; Aguilar, A.; Camacho, F.; Reyes, F.; Fernandez, S.; Perez, J.L.; Acevedo, R.; et al. Mycobacterium smegmatis proteoliposome induce protection in a murine progressive pulmonary tuberculosis model. Tuberculosis 2016, 101, 44–48. [Google Scholar] [CrossRef]
- Van Crevel, R.; Ottenhoff, T.H.; Van Der Meer, J.W. Innate immunity to Mycobacterium tuberculosis. Clin. Microbiol. Rev. 2002, 15, 294–309. [Google Scholar] [CrossRef] [Green Version]
- Tartey, S.; Takeuchi, O. Pathogen recognition and Toll-like receptor targeted therapeutics in innate immune cells. Int. Rev. Immunol. 2017, 36, 57–73. [Google Scholar] [CrossRef]
- Andreu, N.; Phelan, J.; Paola, F.; Cliff, J.M.; Clark, T.G.; Hibberd, M.L. Primary macrophages and J774 cells respond differently to infection with Mycobacterium tuberculosis. Sci. Rep. 2017, 7, 42225. [Google Scholar] [CrossRef] [Green Version]
- Underhill, D.M.; Ozinsky, A.; Smith, K.D.; Aderem, A. Toll-like receptor-2 mediates mycobacteria-induced proinflammatory signaling in macrophages. Proc. Natl. Acad. Sci. USA 1999, 96, 14459–14463. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Akira, S. Toll-like receptor signaling. J. Biol. Chem. 2003, 278, 38105–38108. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kremer, L.; Maughan, W.; Wilson, R.; Dover, L.; Besra, G. The M. tuberculosis antigen 85 complex and mycolyltransferase activity. Lett. Appl. Microbiol. 2002, 34, 233–237. [Google Scholar] [CrossRef] [PubMed]
- Belisle, J.T.; Vissa, V.D.; Sievert, T.; Takayama, K.; Brennan, P.J.; Besra, G.S. Role of the major antigen of Mycobacterium tuberculosis in cell wall biogenesis. Science 1997, 276, 1420–1422. [Google Scholar] [CrossRef]
- Naito, M.; Ohara, N.; Matsumoto, S.; Yamada, T. The novel fibronectin-binding motif and key residues of mycobacteria. J. Biol. Chem. 1998, 273, 2905–2909. [Google Scholar] [CrossRef] [Green Version]
- Kuo, C.-J.; Bell, H.; Hsieh, C.-L.; Ptak, C.P.; Chang, Y.-F. Novel mycobacteria antigen 85 complex binding motif on fibronectin. J. Biol. Chem. 2012, 287, 1892–1902. [Google Scholar] [CrossRef] [Green Version]
- Takamura, S.; Matsuo, K.; Takebe, Y.; Yasutomi, Y. Ag85B of mycobacteria elicits effective CTL responses through activation of robust Th1 immunity as a novel adjuvant in DNA vaccine. J. Immunol. 2005, 175, 2541–2547. [Google Scholar] [CrossRef]
- Yan, Q.; Cheng, Z.; Liu, H.; Shan, W.; Cheng, Z.; Dai, X.; Xue, Y.; Chen, F. Enhancement of Ag85B DNA vaccine immunogenicity against tuberculosis by dissolving microneedles in mice. Vaccine 2018, 36, 4471–4476. [Google Scholar] [CrossRef]
- Zhu, D.; Jiang, S.; Luo, X. Therapeutic effects of Ag85B and MPT64 DNA vaccines in a murine model of Mycobacterium tuberculosis infection. Vaccine 2005, 23, 4619–4624. [Google Scholar] [CrossRef]
- Kadir, N.-A.; Sarmiento, M.E.; Acosta, A.; Norazmi, M.-N. Cellular and humoral immunogenicity of recombinant Mycobacterium smegmatis expressing Ag85B epitopes in mice. Int. J. Mycobacteriol. 2016, 5, 7–13. [Google Scholar] [CrossRef] [Green Version]
- Sarmiento, M.E.; Alvarez, N.; Chin, K.; Bigi, F.; Tirado, Y.; García, M.; Anis, F.; Norazmi, M.N.; Acosta, A. Tuberculosis vaccine candidates based on mycobacterial cell envelope components. Tuberculosis 2019, 115, 26–41. [Google Scholar] [CrossRef] [PubMed]
- Reyrat, J.-M.; Kahn, D. Mycobacterium smegmatis: An absurd model for tuberculosis? Trends Microbiol. 2001, 9, 472–474. [Google Scholar] [CrossRef]
- Shiloh, M.U.; Champion, P.A.D. To catch a killer. What can mycobacterial models teach us about Mycobacterium tuberculosis pathogenesis? Curr. Opin. Microbiol. 2010, 13, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Prince, J.T.; Marcotte, E.M. Mass spectrometry of the M. smegmatis proteome: Protein expression levels correlate with function, operons, and codon bias. Genome Res. 2005, 15, 1118–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelley, V.A.; Schorey, J.S. Mycobacterium’s arrest of phagosome maturation in macrophages requires Rab5 activity and accessibility to iron. Mol. Biol. Cell 2003, 14, 3366–3377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheadle, E.J.; O’Donnell, D.; Selby, P.J.; Jackson, A.M. Closely related mycobacterial strains demonstrate contrasting levels of efficacy as antitumor vaccines and are processed for major histocompatibility complex class I presentation by multiple routes in dendritic cells. Infect. Immun. 2005, 73, 784–794. [Google Scholar] [CrossRef] [Green Version]
- Young, S.L.; Murphy, M.; Zhu, X.W.; Harnden, P.; O’Donnell, M.A.; James, K.; Patel, P.M.; Selby, P.J.; Jackson, A.M. Cytokine-modified Mycobacterium smegmatis as a novel anticancer immunotherapy. Int. J. Cancer 2004, 112, 653–660. [Google Scholar] [CrossRef]
- Kleinnijenhuis, J.; Oosting, M.; Joosten, L.A.; Netea, M.G.; Van Crevel, R. Innate immune recognition of Mycobacterium tuberculosis. Clin. Dev. Immunol. 2011, 2011, 405310. [Google Scholar] [CrossRef] [Green Version]
- Henao, J.; Sánchez, D.; Munoz, C.H.; Mejía, N.; Arias, M.A.; García, L.F.; Barrera, L.F. Human splenic macrophages as a model for in vitro infection with Mycobacterium tuberculosis. Tuberculosis 2007, 87, 509–517. [Google Scholar] [CrossRef]
- Estrella, J.; Kan-Sutton, C.; Gong, K.; Eissa, T.N.; Rajagopalan, M.; Lewis, D.; Hunter, R.; Jagannath, C. A novel in vitro human macrophage model to study the persistence of Mycobacterium tuberculosis using vitamin D3 and retinoic acid activated THP-1 macrophages. Front. Microbiol. 2011, 2, 67. [Google Scholar] [CrossRef] [Green Version]
- Shah, R.R.; Hassett, K.J.; Brito, L.A. Overview of vaccine adjuvants: Introduction, history, and current status. In Vaccine Adjuvants; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–13. [Google Scholar]
- Ding, C.; Ma, J.; Dong, Q.; Liu, Q. Live bacterial vaccine vector and delivery strategies of heterologous antigen: A review. Immunol. Lett. 2018, 197, 70–77. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, A.J.; Zangirolami, T.C.; Novo-Mansur, M.T.M.; Giordano, R.D.C.; Martins, E.A.L. Live bacterial vaccine vectors: An overview. Braz. J. Microbiol. 2014, 45, 1117–1129. [Google Scholar] [CrossRef] [PubMed]
- Faludi, I.; Szabó, Á.; Burián, K.; Endrész, V.; Miczák, A. Recombinant Mycobacterium smegmatis vaccine candidates. Acta Microbiol. Immunol. Hung. 2011, 58, 13–22. [Google Scholar] [CrossRef]
- Da Costa, A.C.; Nogueira, S.V.; Kipnis, A.; Junqueira-Kipnis, A.P. Recombinant BCG: Innovations on an old vaccine. Scope of BCG strains and strategies to improve long-lasting memory. Front. Immunol. 2014, 5, 152. [Google Scholar] [CrossRef] [Green Version]
- Anes, E.; Peyron, P.; Staali, L.; Jordao, L.; Gutierrez, M.G.; Kress, H.; Hagedorn, M.; Maridonneau-Parini, I.; Skinner, M.A.; Wildeman, A.G. Dynamic life and death interactions between Mycobacterium smegmatis and J774 macrophages. Cell. Microbiol. 2006, 8, 939–960. [Google Scholar] [CrossRef] [PubMed]
- Jordao, L.; Bleck, C.K.; Mayorga, L.; Griffiths, G.; Anes, E. On the killing of mycobacteria by macrophages. Cell. Microbiol. 2008, 10, 529–548. [Google Scholar] [CrossRef] [PubMed]
- Weiss, G.; Schaible, U.E. Macrophage defense mechanisms against intracellular bacteria. Immunol. Rev. 2015, 264, 182–203. [Google Scholar] [CrossRef] [Green Version]
- Rich, F.J.; Kuhn, S.; Hyde, E.J.; Harper, J.L.; Ronchese, F.; Kirman, J.R. Induction of T cell responses and recruitment of an inflammatory dendritic cell subset following tumor immunotherapy with Mycobacterium smegmatis. Cancer Immunol. Immunother. 2012, 61, 2333–2342. [Google Scholar] [CrossRef]
- Lee, B.-Y.; Horwitz, M.A. T-cell epitope mapping of the three most abundant extracellular proteins of Mycobacterium tuberculosis in outbred guinea pigs. Infect. Immun. 1999, 67, 2665–2670. [Google Scholar] [CrossRef] [Green Version]
- Roche, P.W.; Peake, P.W.; Billman-Jacobe, H.; Doran, T.; Britton, W.J. T-cell determinants and antibody binding sites on the major mycobacterial secretory protein MPB59 of Mycobacterium bovis. Infect. Immun. 1994, 62, 5319–5326. [Google Scholar] [CrossRef] [Green Version]
- Gaudino, S.J.; Kumar, P. Cross-talk between antigen presenting cells and T cells impacts intestinal homeostasis, bacterial infections, and tumorigenesis. Front. Immunol. 2019, 10, 360. [Google Scholar] [CrossRef] [Green Version]
- Unanue, E.R. Perspective on antigen processing and presentation. Immunol. Rev. 2002, 185, 86–102. [Google Scholar] [CrossRef] [PubMed]
- Kaufmann, S.H. Protection against tuberculosis: Cytokines, T cells, and macrophages. Ann. Rheum. Dis. 2002, 61 (Suppl. S2), ii54–ii58. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.; Gowthaman, U.; Pahari, S.; Agrewala, J.N. Manipulation of costimulatory molecules by intracellular pathogens: Veni, vidi, vici!! PLoS Pathog. 2012, 8, e1002676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Flores-Batista, V.; Boechat, N.; Lago, P.; Lazzarini, L.; Pessanha, L.; Almeida, A.; Mafort, T.; Kritski, A.; Ho, J.; Lapa-e-Silva, J. Low expression of antigen-presenting and costimulatory molecules by lung cells from tuberculosis patients. Braz. J. Med Biol. Res. 2007, 40, 1671–1679. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grewal, I.S.; Flavell, R.A. The role of CD40 ligand in costimulation and T-cell activation. Immunol. Rev. 1996, 153, 85–106. [Google Scholar] [CrossRef]
- Schreiber, H.A.; Hulseberg, P.D.; Lee, J.; Prechl, J.; Barta, P.; Szlavik, N.; Harding, J.S.; Fabry, Z.; Sandor, M. Dendritic cells in chronic mycobacterial granulomas restrict local anti-bacterial T cell response in a murine model. PLoS ONE 2010, 5, e11453. [Google Scholar] [CrossRef]
- Hubo, M.; Trinschek, B.; Kryczanowsky, F.; Tuettenberg, A.; Steinbrink, K.; Jonuleit, H. Costimulatory molecules on immunogenic versus tolerogenic human dendritic cells. Front. Immunol. 2013, 4, 82. [Google Scholar] [CrossRef] [Green Version]
- Holling, T.M.; Schooten, E.; van Den Elsen, P.J. Function and regulation of MHC class II molecules in T-lymphocytes: Of mice and men. Hum. Immunol. 2004, 65, 282–290. [Google Scholar] [CrossRef]
- Leitner, J.; Kuschei, W.; Grabmeier-Pfistershammer, K.; Woitek, R.; Kriehuber, E.; Majdic, O.; Zlabinger, G.; Pickl, W.F.; Steinberger, P. T cell stimulator cells, an efficient and versatile cellular system to assess the role of costimulatory ligands in the activation of human T cells. J. Immunol. Methods 2010, 362, 131–141. [Google Scholar] [CrossRef] [Green Version]
- Podojil, J.R.; Miller, S.D. Targeting the B7 family of co-stimulatory molecules. BioDrugs 2013, 27, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, J.-C.D.; Zhang, X.; Fedorov, A.A.; Nathenson, S.G.; Almo, S.C. Structural basis for co-stimulation by the human CTLA-4/B7-2 complex. Nature 2001, 410, 604–608. [Google Scholar] [CrossRef] [PubMed]
- Greenwald, R.J.; Freeman, G.J.; Sharpe, A.H. The B7 family revisited. Annu. Rev. Immunol. 2005, 23, 515–548. [Google Scholar] [CrossRef] [PubMed]
- Grewal, I.S.; Borrow, P.; Pamer, E.G.; Oldstone, M.B.; Flavell, R.A. The CD40–CD154 system in anti-infective host defense. Curr. Opin. Immunol. 1997, 9, 491–497. [Google Scholar] [CrossRef]
- Bonato, V.; Medeiros, A.; Lima, V.; Dias, A.; Faccioli, L.; Silva, C. Downmodulation of CD18 and CD86 on Macrophages and VLA-4 on Lymphocytes in Experimental Tuberculosis. Scand. J. Immunol. 2001, 54, 564–573. [Google Scholar] [CrossRef] [PubMed]
- Leal, I.S.; Flórido, M.; Andersen, P.; Appelberg, R. Interleukin-6 regulates the phenotype of the immune response to a tuberculosis subunit vaccine. Immunology 2001, 103, 375–381. [Google Scholar] [CrossRef] [PubMed]
- Leal, I.S.; Smedegård, B.; Andersen, P.; Appelberg, R. Interleukin-6 and interleukin-12 participate in induction of a type 1 protective T-cell response during vaccination with a tuberculosis subunit vaccine. Infect. Immun. 1999, 67, 5747–5754. [Google Scholar] [CrossRef] [Green Version]
- Zuñiga, J.; Torres-García, D.; Santos-Mendoza, T.; Rodriguez-Reyna, T.S.; Granados, J.; Yunis, E.J. Cellular and humoral mechanisms involved in the control of tuberculosis. Clin. Dev. Immunol. 2012, 2012, 1–18. [Google Scholar] [CrossRef]
- Su, B.; Wang, J.; Wang, X.; Jin, H.; Zhao, G.; Ding, Z.; Kang, Y.; Wang, B. The effects of IL-6 and TNF-α as molecular adjuvants on immune responses to FMDV and maturation of dendritic cells by DNA vaccination. Vaccine 2008, 26, 5111–5122. [Google Scholar] [CrossRef]
- Singh, V.; Jain, S.; Gowthaman, U.; Parihar, P.; Gupta, P.; Gupta, U.D.; Agrewala, J.N. Co-administration of IL-1+ IL-6+ TNF-α with Mycobacterium tuberculosis infected macrophages vaccine induces better protective T cell memory than BCG. PLoS ONE 2011, 6, e16097. [Google Scholar] [CrossRef]
- Rodríguez, T.; Pérez, O.; Ménager, N.; Ugrinovic, S.; Bracho, G.; Mastroeni, P. Interactions of proteoliposomes from serogroup B Neisseria meningitidis with bone marrow-derived dendritic cells and macrophages: Adjuvant effects and antigen delivery. Vaccine 2005, 23, 1312–1321. [Google Scholar] [CrossRef] [PubMed]
- Bohsali, A.; Abdalla, H.; Velmurugan, K.; Briken, V. The non-pathogenic mycobacteria M. smegmatis and M. fortuitum induce rapid host cell apoptosis via a caspase-3 and TNF dependent pathway. BMC Microbiol. 2010, 10, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.; Mosser, D. Macrophage activation by endogenous danger signals. J. Pathol. 2008, 214, 161–178. [Google Scholar] [CrossRef]
- Sutherland, J.S.; Adetifa, I.M.; Hill, P.C.; Adegbola, R.A.; Ota, M.O. Pattern and diversity of cytokine production differentiates between Mycobacterium tuberculosis infection and disease. Eur. J. Immunol. 2009, 39, 723–729. [Google Scholar] [CrossRef] [PubMed]
- Cooper, A.M.; Khader, S.A. The role of cytokines in the initiation, expansion, and control of cellular immunity to tuberculosis. Immunol. Rev. 2008, 226, 191–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jasenosky, L.D.; Scriba, T.J.; Hanekom, W.A.; Goldfeld, A.E. T cells and adaptive immunity to Mycobacterium tuberculosis in humans. Immunol. Rev. 2015, 264, 74–87. [Google Scholar] [CrossRef]
- Snapper, S.; Melton, R.; Mustafa, S.; Kieser, T.; Jacobs, W.R., Jr. Isolation and characterization of efficient plasmid transformation mutants of Mycobacterium smegmatis. Mol. Microbiol. 1990, 4, 1911–1919. [Google Scholar] [CrossRef]
- Bettencourt, P.; Carmo, N.; Pires, D.; Timóteo, P.; Anes, E. Mycobacterial Infection of Macrophages: The Effect of the Multiplicity of Infection. In Antimicrobial Research: Novel Bioknowledge and Educational Programs; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2017; pp. 651–664. [Google Scholar]
- Gopinath, V.; Musa, M.; Samsudin, A.; Lalitha, P.; Sosroseno, W. Role of nitric oxide in hydroxyapatite-induced phagocytosis by murine macrophage cell line (RAW264. 7). Arch. Oral Biol. 2006, 51, 339–344. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kadir, N.-A.; Acosta, A.; Sarmiento, M.E.; Norazmi, M.-N. Immunomodulatory Effects of Recombinant Mycobacterium smegmatis Expressing Antigen-85B Epitopes in Infected J774A.1 Murine Macrophages. Pathogens 2020, 9, 1000. https://doi.org/10.3390/pathogens9121000
Kadir N-A, Acosta A, Sarmiento ME, Norazmi M-N. Immunomodulatory Effects of Recombinant Mycobacterium smegmatis Expressing Antigen-85B Epitopes in Infected J774A.1 Murine Macrophages. Pathogens. 2020; 9(12):1000. https://doi.org/10.3390/pathogens9121000
Chicago/Turabian StyleKadir, Nur-Ayuni, Armando Acosta, Maria E. Sarmiento, and Mohd-Nor Norazmi. 2020. "Immunomodulatory Effects of Recombinant Mycobacterium smegmatis Expressing Antigen-85B Epitopes in Infected J774A.1 Murine Macrophages" Pathogens 9, no. 12: 1000. https://doi.org/10.3390/pathogens9121000