Prevalence and Risk Factors of Intestinal Parasite Infections in Greek Swine Farrow-To-Finish Farms

 ,
,  ,
, 
Abstract
:1. Introduction
2. Results
2.1. Epizootiology of Intestinal Parasites in the Studied Population
2.2. Effects of Risk Factors on Parasitic Infections
2.2.1. B. coli
2.2.2. Entamoeba spp.
2.2.3. C. suis
2.2.4. A. suum
2.2.5. T. suis
2.2.6. Oesophagostomum spp.
3. Discussion
4. Materials and Methods
4.1. Sampling Process
4.2. Coprological Methods Used
4.3. Data Handling-Statistical Analyses
4.4. Ethics Approval and Consent to Participate
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Joachim, A.; Dülmer, N.; Daugschies, A.; Roepstorff, A. Occurrence of helminthes in pig fattening units with different management systems in Northern Germany. Vet. Parasitol. 2001, 96, 135–146. [Google Scholar] [CrossRef]
- Weng, Y.B.; Hu, Y.J.; Li, Y.; Li, B.S.; Lin, R.Q.; Xie, D.H.; Gasser, R.B.; Zhu, X.Q. Survey of intestinal parasites in pigs from intensive farms in Guangdong Province, People’s Republic of China. Vet. Parasitol. 2005, 127, 333–336. [Google Scholar] [CrossRef] [PubMed]
- Greve, J.H. Internal parasites: Helminths. In Diseases of Swine, 10th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; Wiley-Blackwell: West Sussex, UK, 2012; pp. 908–920. [Google Scholar]
- Lindsay, D.S.; Dubey, J.P.; Santín-Durán, M.; Fayer, R. Coccidia and other Protozoa. In Diseases of Swine, 10th ed.; Zimmerman, J.J., Karriker, L.A., Ramirez, A., Schwartz, K.J., Stevenson, G.W., Eds.; Wiley-Blackwell: West Sussex, UK, 2012; pp. 895–907. [Google Scholar]
- Roepstorff, A.; Nilsson, O.; Oksanen, A.; Gjerde, B.; Richter, S.H.; Ortenberg, E.; Christensson, D.; Martinsson, K.B.; Bartlett, P.C.; Nansen, P.; et al. Intestinal parasites in swine in the Nordic countries: Prevalence and geographical distribution. Vet. Parasitol. 1998, 76, 305–319. [Google Scholar] [CrossRef]
- Vandekerckhove, E. The Use of Serology in the Control of Ascaris Suum Infections in Pigs. Ph.D. Thesis, School of Veterinary Medicine, Ghent, Belgium, 2018. [Google Scholar]
- Papatsiros, V.; Tassis, P.; Christodoulopoulos, G.; Boutsini, S.; Tsirigotakis, G.; Tzika, E. Health and Production of Greek Organic Pig Farming: Current situation and perspectives. J. Hell. Vet. Med. Soc. 2017, 63, 37–44. [Google Scholar] [CrossRef] [Green Version]
- Pedersen, S.; Saeed, I.; Michaelsen, K.F.; Friis, H.; Murrell, K.D. Impact of protein energy malnutrition on Trichuris suis infection in pigs concomitantly infected with Ascaris suum. Parasitology 2002, 124, 561–568. [Google Scholar] [CrossRef]
- Knecht, D.; Popiołek, M.; Zaleśny, G. Does meatiness of pigs depend on the level of gastro-intestinal parasites infection? Prev. Vet. Med. 2011, 99, 234–239. [Google Scholar] [CrossRef] [PubMed]
- Nejsum, P.; Betson, M.; Bendall, R.P.; Thamsborg, S.M.; Stothard, J.R. Assessing the zoonotic potential of Ascaris suum and Trichuris suis: Looking to the future from an analysis of the past. J. Helminthol. 2012, 86, 148–155. [Google Scholar] [CrossRef] [Green Version]
- Schubnell, F.; von Ah, S.; Graage, R.; Sydler, T.; Sidler, X.; Hadorn, D.; Basso, W. Occurrence, clinical involvement and zoonotic potential of endoparasites infecting Swiss pigs. Parasitol. Int. 2016, 65, 618–624. [Google Scholar] [CrossRef]
- Sharma, S.; Harding, G. Necrotizing lung infection caused by the protozoan Balantidium coli. Can. J. Infect. Dis. 2003, 14, 163–166. [Google Scholar] [CrossRef] [Green Version]
- Ferry, T.; Bouhour, D.; De Monbrison, F.; Laurent, F.; Dumouchel-Champagne, H.; Picot, S.; Piens, M.A.; Granier, P. Severe peritonitis due to Balantidium coli acquired in France. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 393–395. [Google Scholar] [CrossRef]
- Daugschies, A. Protozoen Infektionen des Schweines. In Veterinärmedizinische Parasitologie, 6th ed.; Schnieder, T., Ed.; Parey: Stuttgart, Germany, 2006; pp. 359–408. [Google Scholar]
- Xiao, L. Molecular epidemiology of cryptosporidiosis: An update. Exp. Parasitol. 2010, 124, 80–89. [Google Scholar] [CrossRef] [PubMed]
- Roepstorff, A.; Mejer, H.; Nejsum, P.; Thamsborg, S.M. Helminth parasites in pigs: New challenges in pig production and current research highlights. Vet. Parasitol. 2011, 180, 72–81. [Google Scholar] [CrossRef] [Green Version]
- Joachim, A.; Daugschies, A. Endoparasites in swine in different age groups and management systems. Berl. Munch. Tierarztl. Wochenschr. 2000, 113, 129–133. [Google Scholar] [PubMed]
- Damriyasa, I.M.; Bauer, C. Prevalence and age-dependent occurrence of intestinal protozoan infections in suckling piglets. Berl. Munch. Tierarztl. Wochenschr. 2006, 119, 287–290. [Google Scholar] [PubMed]
- Eijck, I.A.; Borgsteede, F.H. A survey of gastrointestinal pig parasites on free-range, organic and conventional pig farms in The Netherlands. Vet. Res. Commun. 2005, 29, 407–414. [Google Scholar] [CrossRef] [PubMed]
- Kochanowski, M.; Karamon, J.; Dąbrowska, J.; Dors, A.; Czyżewska-Dors, E.; Cencek, T. Occurrence of Intestinal Parasites in Pigs in Poland—The Influence of Factors Related to the Production System. J. Vet. Res. 2017, 61, 459–466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barbosa, A.S.; Bastos, O.M.P.; Dib, L.V.; de Siqueira, M.P.; Cardozo, M.L.; Ferreira, L.C.; Chaves, W.T.; Fonseca, A.B.M.; Uchôa, C.M.A.; Amendoeira, M.R.R. Gastrointestinal parasites of swine raised in different management systems in the State of Rio de Janeiro, Brazil. Pesq. Vet. Bras. 2015, 35, 941–946. [Google Scholar] [CrossRef] [Green Version]
- Hindsbo, O.; Nielsen, C.V.; Andreassen, J.; Willingham, A.L.; Bendixen, M.; Nielsen, M.A.; Nielsen, N.O. Age-dependent occurrence of the intestinal ciliate Balantidium coli in pigs at a Danish research farm. Acta Vet. Scand. 2000, 41, 79–83. [Google Scholar]
- McDonald, J.D. On Balantidium coli (Malmsten) and Balantidium suis (sp. nov.) with an account of their neuromotor apparatus. Univ. Cal. Pub. Zool. 1922, 20, 243–300. [Google Scholar]
- Ponce-Gordo, F.; Fonseca-Salamanca, F.; Martínez-Díaz, R.A. Genetic Heterogeneity in Internal Transcribed Spacer Genes of Balantidium coli (Litostomatea, Ciliophora). Protist 2011, 162, 774–794. [Google Scholar] [CrossRef]
- Ponce-Gordo, F.; Jirků-Pomajbíková, K. Balantidium coli. In Global Water Pathogens Project; Rose, J.B., Jiménez-Cisneros, B., Eds.; Michigan State University Press: East Lansing, MI, USA, 2017; pp. 3–14. [Google Scholar]
- Schuster, F.L.; Visvesvara, G.S. Amebae and ciliated protozoa as causal agents of waterborne zoonotic disease. Vet. Parasitol. 2004, 126, 91–120. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.G.; Jordan, H.E.; Luce, W.G.; Coburn, T.C.; Maxwell, C.V. Prevalence of gastrointestinal parasitism in Oklahoma swine. Am. J. Vet. Res. 1984, 45, 2421–2423. [Google Scholar]
- Schuster, F.L.; Ramirez-Avila, L. Current world status of Balantidium coli. Clin. Microbiol. Rev. 2008, 21, 626–638. [Google Scholar] [CrossRef] [Green Version]
- Ezenwa, V.O.; Jolles, A.E. From Host Immunity to Pathogen Invasion: The effects of helminth coinfection on the dynamics of microparasites. Integr. Comp. Biol. 2011, 51, 540–551. [Google Scholar] [CrossRef]
- Mundt, H.C.; Cohnen, A.; Daugschies, A.; Joachim, A.; Prosl, H.; Schmäschke, R.; Westphal, B. Occurrence of Isospora suis in Germany, Switzerland and Austria. J. Vet. Med. B. Infect. Dis. Vet. Public Health 2005, 52, 93–97. [Google Scholar] [CrossRef] [PubMed]
- Worliczek, H.L.; Buggelsheim, M.; Saalmüller, A.; Joachim, A. Porcine isosporosis: Infection dynamics, pathophysiology and immunology of experimental infections. Wien. Klin. Wochenschr. 2007, 119, 33–39. [Google Scholar] [CrossRef]
- Chae, C.; Kwon, D.; Kim, O.; Min, K.; Cheon, D.S.; Choi, C.; Kim, B.; Suh, J. Diarrhoea in nursing piglets associated with coccidiosis: Prevalence, microscopic lesions and coexisting microorganisms. Vet. Rec. 1998, 143, 417–420. [Google Scholar] [CrossRef]
- Shrestha, A.; Abd-Elfattah, A.; Freudenschuss, B.; Hinney, B.; Palmieri, N.; Ruttkowski, B.; Joachim, A. Cystoisospora suis—A Model of Mammalian Cystoisosporosis. Front. Vet. Sci. 2015, 2, 68. [Google Scholar] [CrossRef] [Green Version]
- Komatsu, T.; Matsubayashi, M.; Murakoshi, N.; Sasai, K.; Shibahara, T. Retrospective and Histopathological Studies of Entamoeba spp. and Other Pathogens Associated with Diarrhea and Wasting in Pigs in Aichi Prefecture, Japan. Jpn. Agric. Res. Q. 2019, 53, 59–67. [Google Scholar] [CrossRef] [Green Version]
- Koudela, B.; Kucerová, S. Role of acquired immunity and natural age resistance on course of Isospora suis coccidiosis in nursing piglets. Vet. Parasitol. 1999, 82, 93–99. [Google Scholar] [CrossRef]
- Wieler, L.H.; Ilieff, A.; Herbst, W.; Bauer, C.; Vieler, E.; Bauerfeind, R.; Failing, K.; Klös, H.; Wengert, D.; Baljer, G.; et al. Prevalence of enteropathogens in suckling and weaned piglets with diarrhoea in southern Germany. J. Vet. Med. B. Infect. Dis. Vet. Public Health 2001, 48, 151–159. [Google Scholar] [CrossRef]
- Joachim, A.; Mundt, H.C. Efficacy of sulfonamides and Baycox® against Isospora suis in experimental infections of suckling piglets. Parasitol. Res. 2011, 109, 1653–1659. [Google Scholar] [CrossRef]
- Kreiner, T.; Worliczek, H.L.; Tichy, A.; Joachim, A. Influence of toltrazuril treatment on parasitological parameters and health performance of piglets in the field— An Austrian experience. Vet. Parasitol. 2011, 183, 14–20. [Google Scholar] [CrossRef]
- Rodríguez-Vivas, R.I.; Domínguez-Alpizar, J.L.; Torres-Acosta, J.F. Epidemiological factors associated to bovine coccidiosis in calves (Bos indicus) in a subhumid tropical climate. Rev. Biomed. 1996, 7, 211–218. [Google Scholar]
- Balicka-Ramisz, A.; Ramisz, A.; Vovk, S.; Snitynskyj, V. Prevalence of coccidia infection in goats in Western Pomerania (Poland) and West Ukraine Region. Ann. Parasitol. 2012, 58, 167–171. [Google Scholar]
- Awais, M.M.; Akhtar, M.A.; Muhammad, F.; Anwar, M.I. Seasonal prevalence of coccidiosis in industrial broiler chickens in Faisalabad, Pakistan. Trop. Anim. Health Prod. 2012, 44, 323–328. [Google Scholar] [CrossRef]
- Raue, K.; Heuer, L.; Böhm, C.; Wolken, S.; Epe, C.; Strube, C. 10-year parasitological examination results (2003 to 2012) of faecal samples from horses, ruminants, pigs, dogs, cats, rabbits and hedgehogs. Parasitol. Res. 2017, 116, 3315–3330. [Google Scholar] [CrossRef]
- Rzeżutka, A.; Kaupke, A.; Kozyra, I.; Pejsak, Z. Molecular studies on pig cryptosporidiosis in Poland. Pol. J. Vet. Sci. 2014, 17, 577–582. [Google Scholar] [CrossRef] [Green Version]
- Nansen, P.; Roepstorff, A. Parasitic helminthes of the pig: Factors influencing transmission and infection levels. Int. J. Parasitol. 1999, 29, 877–891. [Google Scholar] [CrossRef]
- Carstensen, L.; Vaarst, M.; Roepstorff, A. Helminth infections in Danish organic swine herds. Vet. Parasitol. 2002, 106, 253–264. [Google Scholar] [CrossRef]
- Katakam, K.K.; Thamsborg, S.M.; Dalsgaard, A.; Kyvsgaard, N.C.; Mejer, H. Environmental contamination and transmission of Ascaris suum in Danish organic pig farms. Parasit. Vectors 2016, 9, 80. [Google Scholar] [CrossRef] [Green Version]
- De Silva, N.R.; Brooker, S.; Hotez, P.J.; Montresor, A.; Engels, D.; Savioli, L. Soil-transmitted helminth infections: Updating the global picture. Trends Parasitol. 2003, 19, 547–551. [Google Scholar] [CrossRef]
- Takata, I. Experimental infection of man with Ascaris of man and the pig. Kitasato Arch. Exp. Med. 1951, 23, 49–59. [Google Scholar]
- Galvin, T.J. Development of human and pig Ascaris in the pig and rabbit. J. Parasitol. 1968, 54, 1085–1091. [Google Scholar] [CrossRef] [PubMed]
- Peng, W.D.; Yuan, K.; Hu, M.; Zhou, X.M.; Gasser, R.B. Mutation scanning-coupled analysis of haplotypic variability in mitochondrial DNA regions reveals low gene flow between human and porcine Ascaris in endemic regions of China. Electrophoresis 2005, 26, 4317–4326. [Google Scholar] [CrossRef]
- Betson, M.; Halstead, F.D.; Nejsum, P.; Imison, E.; Khamis, I.S.; Sousa-Figueiredo, J.C.; Rollinson, D.; Stothard, J.R. A molecular epidemiological investigation of Ascaris on Unguja, Zanzibar using isoenyzme analysis, DNA barcoding and microsatellite DNA profiling. Trans. R. Soc. Trop. Med. Hyg. 2011, 105, 370–379. [Google Scholar] [CrossRef] [PubMed]
- Nissen, S. Morphological and Genetic Diversity of Whipworms (Trichuris spp.) Recovered from Pigs and Humans. Master’s Thesis, University of Copenhagen, Copenhagen, Denmark, 2009. [Google Scholar]
- Cutillas, C.; Callejón, R.; de Rojas, M.; Tewes, B.; Ubeda, J.M.; Ariza, C.; Guevara, D.C. Trichuris suis and Trichuris trichiura are different nematode species. Acta Trop. 2009, 111, 299–307. [Google Scholar] [CrossRef]
- Roepstorff, A. Natural Ascaris suum infections in swine diagnosed by coprological and serological (ELISA) methods. Parasitol. Res. 1998, 84, 537–543. [Google Scholar] [CrossRef]
- Stephenson, L.S.; Pond, W.G.; Nesheim, M.C.; Krook, L.P.; Crompton, D.W.T. Ascaris suum: Nutrient absorption, growth, and intestinal pathology in young pigs experimentally infected with 15-day-old larvae. Exp. Parasitol. 1980, 49, 15–25. [Google Scholar] [CrossRef]
- Forsum, E.; Nesheim, M.C.; Crompton, D.W.T. Nutritional aspects of Ascaris infection in young protein-deficient pigs. Parasitology 1981, 83, 497–512. [Google Scholar] [CrossRef]
- Ronéus, O. Studies on aetiology and pathogenesis of white spots in the liver of pigs. Acta Vet. Scand. 1966, 7 (Suppl. 16), 1–112. [Google Scholar]
- Yoshihara, S.; Nakagawa, M.; Suda, H.; Ikeda, K.; Hanashiro, K. White spots of the liver in pigs experimentally infected with Ascaris suum. Natl. Inst. Anim. Health Q. 1983, 23, 127–137. [Google Scholar]
- Deplazes, P.; Eckert, J.; von Samson-Himmelstjerna, G.; Zahner, H. Lehrbuch der Parasitologie für Tiermedizin, 3rd ed.; Enke Verlag: Stuttgart, Germany, 2013. [Google Scholar]
- Roepstorff, A. Helminth surveillance as a prerequisite for anthelmintic treatment in intensive sow herds. Vet. Parasitol. 1997, 73, 139–151. [Google Scholar] [CrossRef]
- Wharton, D.A. Ascaris sp.: Water loss during dessication of embryonating eggs. Exp. Parasitol. 1979, 48, 398–406. [Google Scholar] [CrossRef]
- Larsen, M.N.; Roepstorff, A. Seasonal variation in development and survival of Ascaris suum and Trichuris suis eggs on pastures. Parasitology 1999, 119, 209–220. [Google Scholar] [CrossRef]
- Mejer, H.; Roepstorff, A. Long-term survival of Ascaris suum and Trichuris suis eggs in relation to pasture management. In Proceedings of the 23rd International Conference of the World Association for the Advancement of Veterinary Parasitology, Buenos Aires, Argentina, 21–25 August 2011; AAPAVET: Buenos Aires, Argentina, 2011; p. 113. [Google Scholar]
- Stewart, T.B.; Hoyt, P.G. Internal parasites. In Diseases of Swine, 9th ed.; Straw, B.E., Zimmerman, J.J., D’Allaire, S., Taylor, D.J., Eds.; Blackwell Publishing: Ames, IA, USA, 2006; pp. 901–914. [Google Scholar]
- Thomsen, L.E.; Bach Knudsen, K.E.; Hedemann, M.S.; Roepstorff, A. The effect of dietary carbohydrates and Trichuris suis infection on pig large intestine tissue structure, epithelial cell proliferation and mucin characteristics. Vet. Parasitol. 2006, 142, 112–122. [Google Scholar] [CrossRef] [PubMed]
- Li, R.W.; Wu, S.; Li, W.; Navarro, K.; Couch, R.D.; Hill, D.; Urban, J.F., Jr. Alterations in the porcine colon microbiota induced by the gastrointestinal nematode Trichuris suis. Infect. Immun. 2012, 80, 2150–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mansfield, L.S.; Gauthier, D.T.; Abner, S.R.; Jones, K.M.; Wilder, S.R.; Urban, J.F. Enhancement of disease and pathology by synergy of Trichuris suis and Campylobacter jejuni in the colon of immunologically naive swine. Am. J. Trop. Med. Hyg. 2003, 68, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Shin, J.L.; Gardiner, G.W.; Deitel, W.; Kandel, G. Does whipworm increase the pathogenicity of Campylobacter jejuni? A clinical correlate of an experimental observation. Can. J. Gastroenterol. 2004, 18, 175–177. [Google Scholar] [CrossRef] [Green Version]
- Mansfield, L.S.; Urban, J.F., Jr. The pathogenesis of necrotic proliferative colitis in swine is linked to whipworm induced suppression of mucosal immunity to resident bacteria. Vet. Immunol. Immunopathol. 1996, 50, 1–17. [Google Scholar] [CrossRef]
- Burden, D.J.; Hammet, N.C.; Brookes, P.A. Field observations on the longevity of Trichuris suis ova. Vet. Rec. 1987, 121, 43. [Google Scholar] [CrossRef] [PubMed]
- Roepstorff, A.; Murrell, K.D. Transmission dynamics of helminth parasites of pigs on continuous pasture: Oesophagostomum dentatum and Hyostrongylus rubidus. Int. J. Parasitol. 1997, 27, 553–562. [Google Scholar] [CrossRef]
- Faust, E.C.; D’Antonio, J.S.; Odom, V.; Miller, M.J.; Peres, C.; Sawitz, W.; Walker, J.H. A critical study of clinical laboratory techniques for the diagnosis of protozoan cysts and helminth eggs in feces. Am. J. Trop. Med. Hyg. 1938, 18, 169–183. [Google Scholar] [CrossRef]
- Henriksen, S.A.; Pohlenz, J.F.L. Staining of Cryptosporidia by a modified Ziehl-Neelsen technique. Acta Vet. Scand. 1981, 22, 594–596. [Google Scholar] [PubMed]
- Taylor, M.A.; Coop, R.L.; Wall, R.L. Veterinary Parasitology, 3rd ed.; Blackwell Publishing Ltd.: Oxford, UK, 2007. [Google Scholar]
- Zajac, A.M.; Conboy, G.A. Veterinary Clinical Parasitology, 8th ed.; Wiley Blackwell: West Sussex, UK, 2012. [Google Scholar]



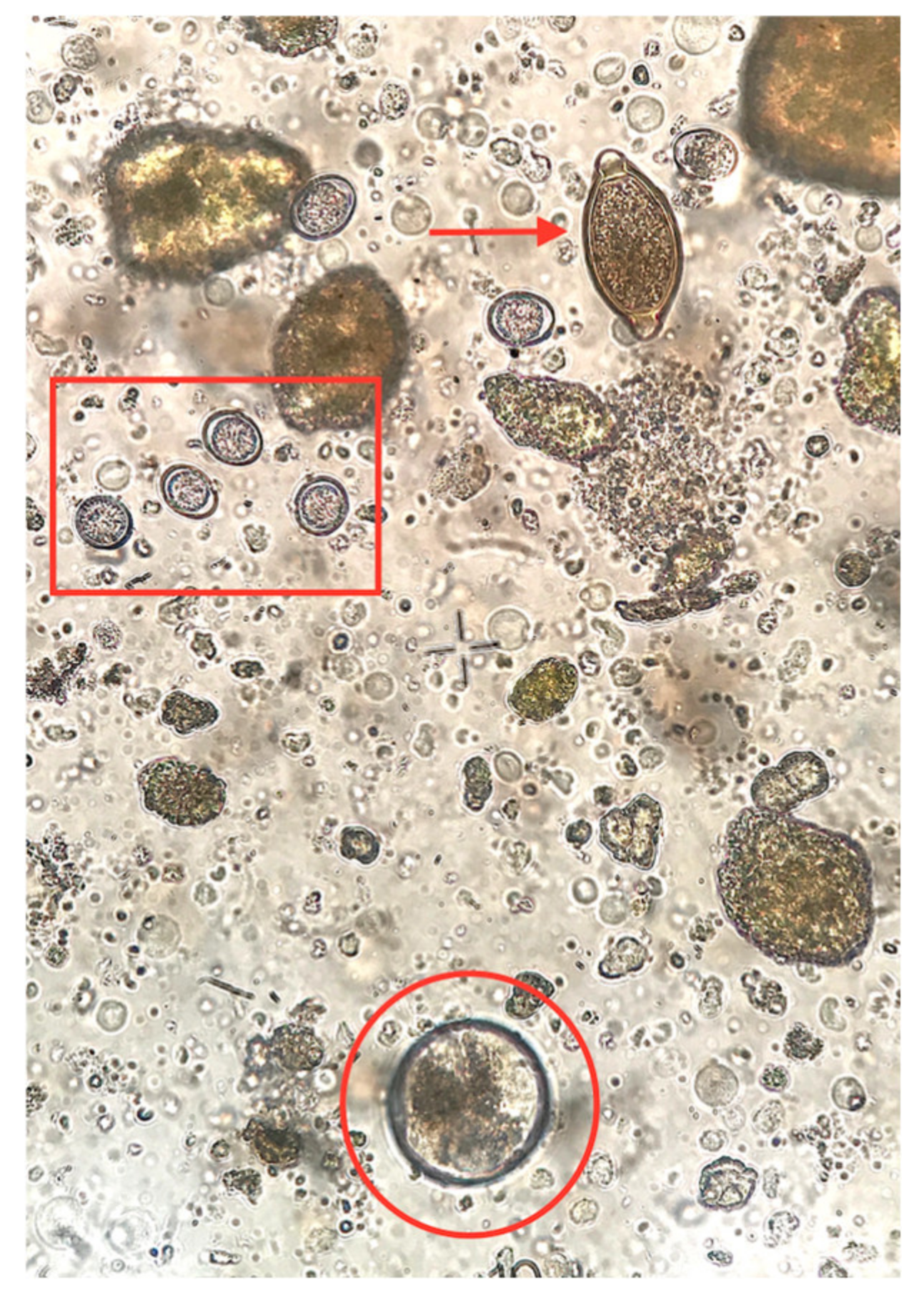{kind=link}
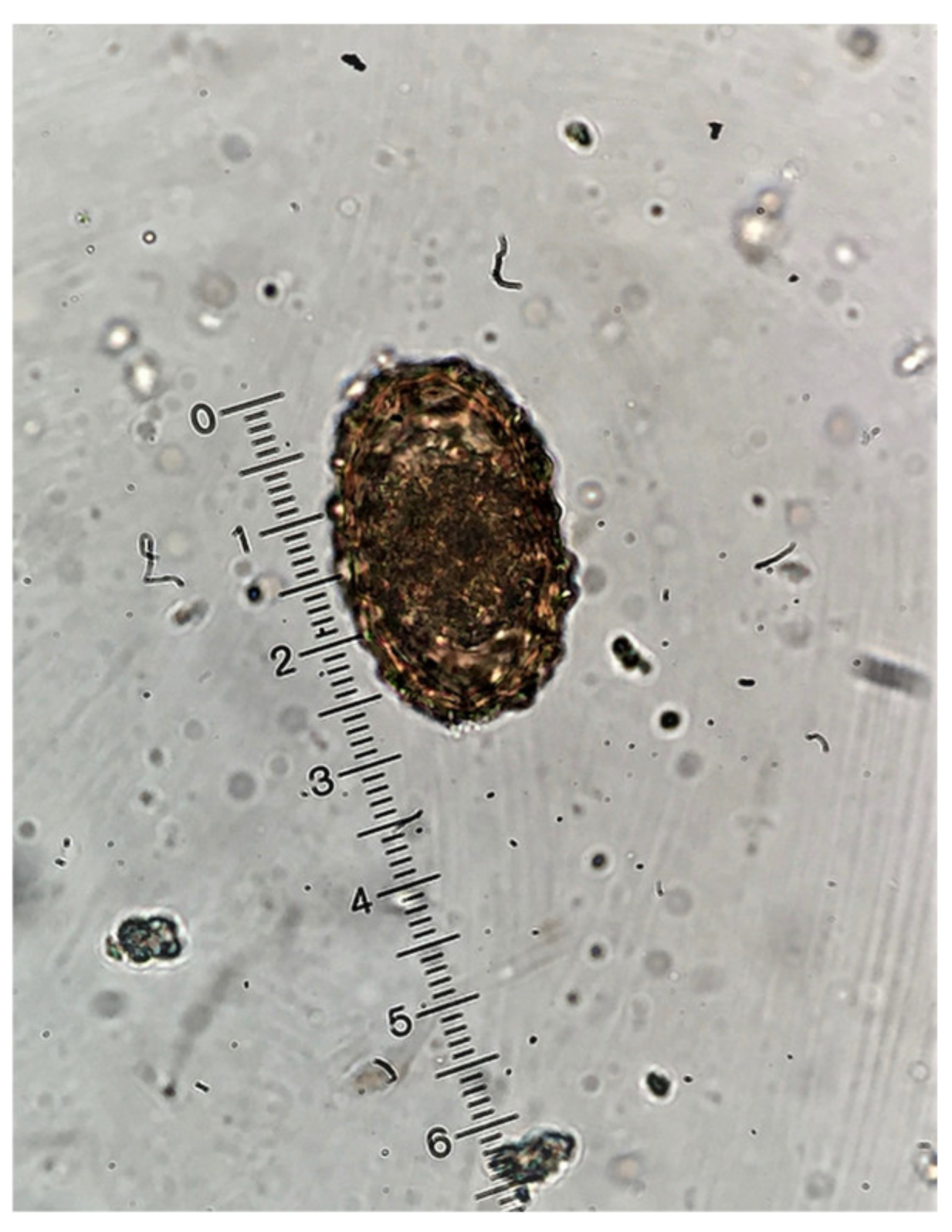{kind=link}
| Parasite | Suckling Piglets | Weaners | Growers | Fatteners | Sows | Total |
|---|---|---|---|---|---|---|
| Balantidium coli | 6 (2.6%) | 31 (13.5%) | 86 (37.4%) | 125 (54.3%) | 187 (81.3%) | 435 (37.8%) |
| Entamoeba spp. | 0 (0.0%) | 0 (0.0%) | 3 (1.3%) | 8 (3.5%) | 84 (36.5%) | 95 (8.3%) |
| Cystoisospora suis | 44 (19.1%) | 21 (9.1%) | 0 (0.0%) | 0 (0.0%) | 4 (1.7%) | 69 (6.0%) |
| Ascaris suum | 0 (0.0%) | 0 (0.0%) | 2 (0.9%) | 2 (0.9%) | 38 (16.5%) | 42 (3.7%) |
| Trichuris suis | 0 (0.0%) | 0 (0.0%) | 1 (0.5%) | 0 (0.0%) | 28 (12.2%) | 29 (2.5%) |
| Oesophagostomum spp. | 0 (0.0%) | 0 (0.0%) | 2 (0.9%) | 2 (0.9%) | 12 (5.2%) | 16 (1.4%) |
| Single infection | 44 (19.1%) | 48 (20.9%) | 83 (36.1%) | 119 (51.7%) | 80 (34.8%) | 374 (32.5%) |
| Double infection | 3 (1.3%) | 2 (0.9%) | 4 (1.7%) | 9 (3.9%) | 94 (40.9%) | 112 (9.7%) |
| Triple infection | 0 (0.0%) | 0 (0.0%) | 1 (0.4%) | 0 (0.0%) | 23 (10.0%) | 24 (2.1%) |
| Quadriple infection | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 0 (0.0%) | 4 (1.7%) | 4 (0.3%) |
| Parasite | Autumn (n = 400) | Winter (n = 350) | Spring (n = 400) |
|---|---|---|---|
| Balantidium coli | 157 (39.3%) | 141 (40.3%) | 137 (34.3%) |
| Entamoeba spp. | 25 (6.3%) | 23 (6.6%) | 47 (11.8%) |
| Cystoisospora suis | 26 (6.5%) | 13 (3.7%) | 30 (7.5%) |
| Ascaris suum | 13 (3.3%) | 16 (4.6%) | 13 (3.3%) |
| Trichuris suis | 7 (1.8%) | 9 (2.6%) | 13 (3.3%) |
| Oesophagostomum spp. | 0 (0.0%) | 10 (2.9%) | 6 (1.5%) |
| B 1 | S.E. 2 | P | Odds Ratio | 95% C.I.3 for EXP(B) | |||
|---|---|---|---|---|---|---|---|
| Lower | Upper | ||||||
| Balantidium coli | Autumn | 0.43 | 0.19 | 0.023 | 1.53 | 1.06 | 2.21 |
| Winter | 0.15 | 0.22 | 0.477 | 1.17 | 0.76 | 1.78 | |
| Spring | Ref. | ||||||
| Suckling piglets | −5.30 | 0.454 | 0 | 0.01 | 0 | 0.01 | |
| Weaners | −3.51 | 0.267 | 0 | 0.03 | 0.02 | 0.05 | |
| Growers | −2.11 | 0.226 | 0 | 0.12 | 0.08 | 0.19 | |
| Fatteners | −1.39 | 0.222 | 0 | 0.25 | 0.16 | 0.39 | |
| Adult | Ref. | ||||||
| ATI (<120 days) | −0.65 | 0.223 | 0.004 | 0.53 | 0.34 | 0.81 | |
| ATI (>120 days) | Ref. | ||||||
| Entamoeba spp. | Autumn | −0.72 | 0.262 | 0.006 | 0.49 | 0.29 | 0.82 |
| Winter | −0.87 | 0.364 | 0.017 | 0.42 | 0.21 | 0.86 | |
| Spring | Ref. | ||||||
| ATI (<120 days) | −0.24 | 0.362 | 0.501 | 0.78 | 0.39 | 1.59 | |
| ATI (>120 days) | Ref. | ||||||
| Cystoisospora suis | Autumn | −0.10 | 0.293 | 0.733 | 0.91 | 0.51 | 1.61 |
| Winter | −0.65 | 0.373 | 0.083 | 0.52 | 0.25 | 1.09 | |
| Spring | Ref. | ||||||
| ATI (<120 days) | −0.26 | 0.357 | 0.462 | 0.77 | 0.38 | 1.55 | |
| ATI (>120 days) | Ref. | ||||||
| Ascaris suum | Autumn | 0.27 | 0.47 | 0.562 | 1.31 | 0.52 | 3.3 |
| Winter | −0.15 | 0.633 | 0.814 | 0.86 | 0.25 | 2.98 | |
| Spring | Ref. | ||||||
| ATI (<120 days) | −0.97 | 0.753 | 0.2 | 0.38 | 0.09 | 1.67 | |
| ATI (>120 days) | Ref. | ||||||
| Trichuris suis | Autumn | 0.06 | 0.606 | 0.921 | 1.06 | 0.32 | 3.49 |
| Winter | −0.85 | 0.59 | 0.151 | 0.43 | 0.14 | 1.36 | |
| Spring | Ref. | ||||||
| ATI (<120 days) | −1.55 | 0.723 | 0.032 | 0.21 | 0.05 | 0.88 | |
| ATI (>120 days) | Ref. | ||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Symeonidou, I.; Tassis, P.; Gelasakis, A.Ι.; Tzika, E.D.; Papadopoulos, E. Prevalence and Risk Factors of Intestinal Parasite Infections in Greek Swine Farrow-To-Finish Farms. Pathogens 2020, 9, 556. https://doi.org/10.3390/pathogens9070556
Symeonidou I, Tassis P, Gelasakis AΙ, Tzika ED, Papadopoulos E. Prevalence and Risk Factors of Intestinal Parasite Infections in Greek Swine Farrow-To-Finish Farms. Pathogens. 2020; 9(7):556. https://doi.org/10.3390/pathogens9070556
Chicago/Turabian StyleSymeonidou, Isaia, Panagiotis Tassis, Athanasios Ι. Gelasakis, Eleni D. Tzika, and Elias Papadopoulos. 2020. "Prevalence and Risk Factors of Intestinal Parasite Infections in Greek Swine Farrow-To-Finish Farms" Pathogens 9, no. 7: 556. https://doi.org/10.3390/pathogens9070556