Influence of the FIV Status and Chronic Gingivitis on Feline Oral Microbiota

 , , and
, , and 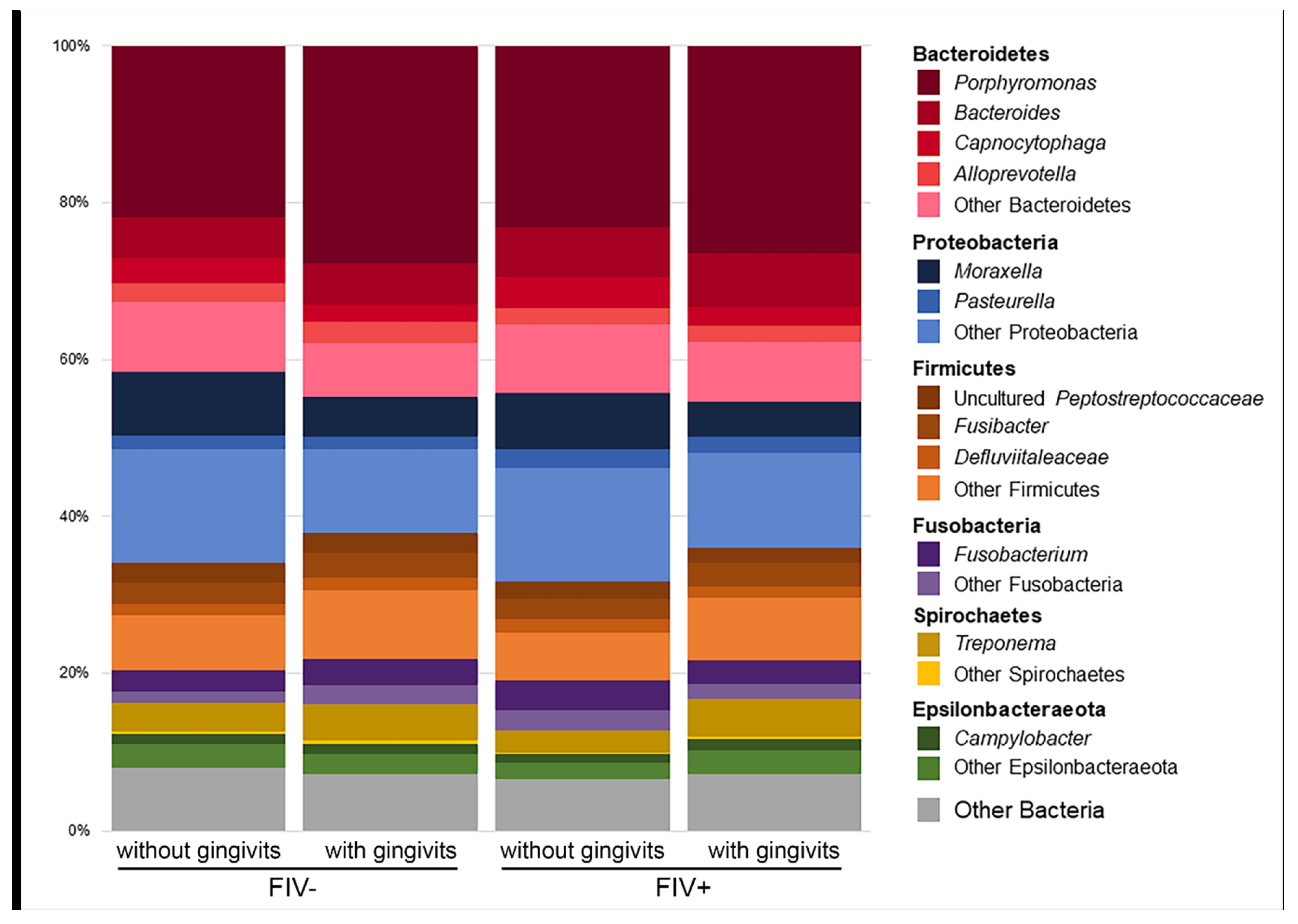{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Effect of the FIV Status
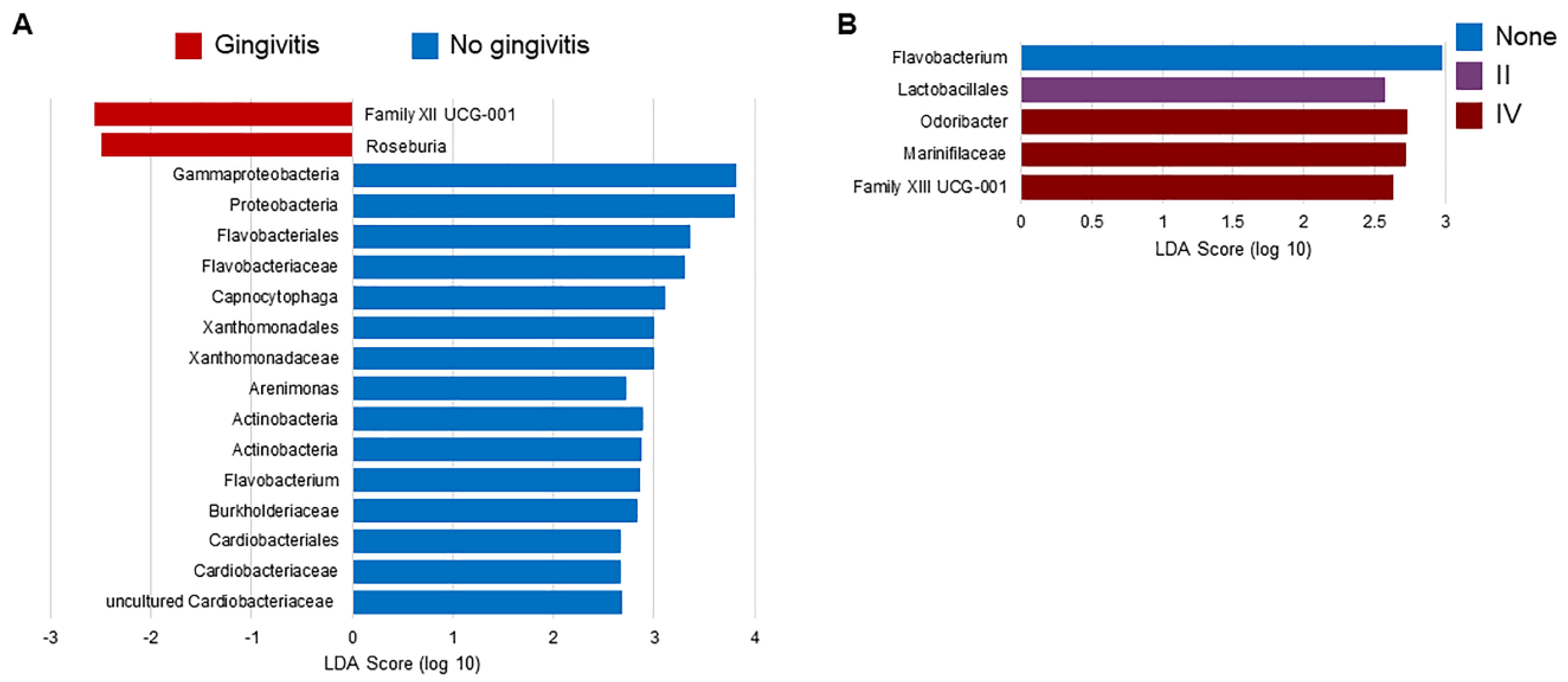
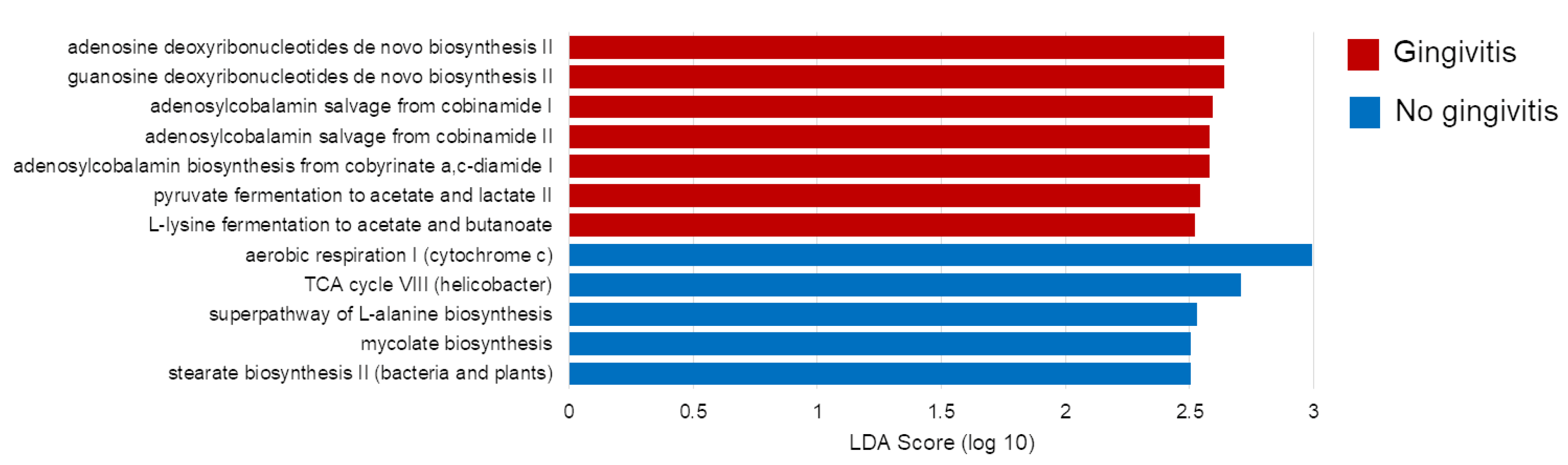
2.2. Effect of the Gingivitis Status
2.3. Effect of Diet
2.4. Other Factors
3. Discussion
4. Materials and Methods
4.1. Sample Collection and DNA Extraction
4.2. Next Generation Sequencing
4.3. Sequence Processing
4.4. Data Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Mallonee, D.H.; Harvey, C.E.; Venner, M.; Hammond, B.F. Bacteriology of periodontal disease in the cat. Arch. Oral Biol. 1988, 33, 677–683. [Google Scholar] [CrossRef]
- Love, D.N.; Johnson, J.L.; Moore, L.V. Bacteroides species from the oral cavity and oral-associated diseases of cats. Vet. Microbiol. 1989, 19, 275–281. [Google Scholar] [CrossRef]
- Dolieslager, S.M.; Riggio, M.P.; Lennon, A.; Lappin, D.F.; Johnston, N.; Taylor, D.; Bennett, D. Identification of bacteria associated with feline chronic gingivostomatitis using culture-dependent and culture-independent methods. Vet. Microbiol. 2011, 148, 93–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakanishi, H.; Furuya, M.; Soma, T.; Hayashiuchi, Y.; Yoshiuchi, R.; Matsubayashi, M.; Tani, H.; Sasai, K. Prevalence of microorganisms associated with feline gingivostomatitis. J. Feline Med. Surg. 2019, 21, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Costalonga, M.; Herzberg, M.C. The oral microbiome and the immunobiology of periodontal disease and caries. Immunol. Lett. 2014, 162, 22–38. [Google Scholar] [CrossRef] [Green Version]
- Rodrigues, M.X.; Bicalho, R.C.; Fiani, N.; Lima, S.F.; Peralta, S. The subgingival microbial community of feline periodontitis and gingivostomatitis: Characterization and comparison between diseased and healthy cats. Sci. Rep. 2019, 9, 12340. [Google Scholar] [CrossRef] [Green Version]
- Tenorio, A.P.; Franti, C.E.; Madewell, B.R.; Pedersen, N.C. Chronic oral infections of cats and their relationship to persistent oral carriage of feline calici-, immunodeficiency, or leukemia viruses. Vet. Immunol. Immunopathol. 1991, 29, 1–14. [Google Scholar] [CrossRef]
- Knowles, J.O.; Gaskell, R.M.; Gaskell, C.J.; Harvey, C.E.; Lutz, H. Prevalence of feline calicivirus, feline leukaemia virus and antibodies to FIV in cats with chronic stomatitis. Vet. Rec. 1989, 124, 336–338. [Google Scholar] [CrossRef]
- Quimby, J.M.; Elston, T.; Hawley, J.; Brewer, M.; Miller, A.; Lappin, M.R. Evaluation of the association of Bartonella species, feline herpesvirus 1, feline calicivirus, feline leukemia virus and feline immunodeficiency virus with chronic feline gingivostomatitis. J. Feline Med. Surg. 2008, 10, 66–72. [Google Scholar] [CrossRef]
- Belgard, S.; Truyen, U.; Thibault, J.C.; Sauter-Louis, C.; Hartmann, K. Relevance of feline calicivirus, feline immunodeficiency virus, feline leukemia virus, feline herpesvirus and Bartonella henselae in cats with chronic gingivostomatitis. Berl. Munch Tierarztl. Wochenschr. 2010, 123, 369–376. [Google Scholar]
- Dowers, K.L.; Hawley, J.R.; Brewer, M.M.; Morris, A.K.; Radecki, S.V.; Lappin, M.R. Association of Bartonella species, feline calicivirus, and feline herpesvirus 1 infection with gingivostomatitis in cats. J. Feline Med. Surg. 2010, 12, 314–321. [Google Scholar] [CrossRef] [PubMed]
- Lommer, M.J. Oral inflammation in small animals. Vet. Clin. N. Am. Small Anim. Pract. 2013, 43, 555–571. [Google Scholar] [CrossRef] [PubMed]
- Kornya, M.R.; Little, S.E.; Scherk, M.A.; Sears, W.C.; Bienzle, D. Association between oral health status and retrovirus test results in cats. J. Am. Vet. Med. Assoc. 2014, 245, 916–922. [Google Scholar] [CrossRef] [PubMed]
- Taniwaki, S.A.; Figueiredo, A.S.; Araujo, J.P., Jr. Virus-host interaction in feline immunodeficiency virus (FIV) infection. Comp. Immunol. Microbiol. Infect. Dis. 2013, 36, 549–557. [Google Scholar] [CrossRef] [PubMed]
- Weese, S.J.; Nichols, J.; Jalali, M.; Litster, A. The oral and conjunctival microbiotas in cats with and without feline immunodeficiency virus infection. Vet. Res. 2015, 46, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goncalves, L.S.; Soares Ferreira, S.M.; Souza, C.O.; Souto, R.; Colombo, A.P. Clinical and microbiological profiles of human immunodeficiency virus (HIV)-seropositive Brazilians undergoing highly active antiretroviral therapy and HIV-seronegative Brazilians with chronic periodontitis. J. Periodontol. 2007, 78, 87–96. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Chandra, J.; Retuerto, M.; Sikaroodi, M.; Brown, R.E.; Jurevic, R.; Salata, R.A.; Lederman, M.M.; Gillevet, P.M.; Ghannoum, M.A. Oral mycobiome analysis of HIV-infected patients: Identification of Pichia as an antagonist of opportunistic fungi. PLoS Pathog. 2014, 10, e1003996. [Google Scholar] [CrossRef]
- Li, Y.; Saxena, D.; Chen, Z.; Liu, G.; Abrams, W.R.; Phelan, J.A.; Norman, R.G.; Fisch, G.S.; Corby, P.M.; Dewhirst, F.; et al. HIV infection and microbial diversity in saliva. J. Clin. Microbiol. 2014, 52, 1400–1411. [Google Scholar] [CrossRef] [Green Version]
- Kistler, J.O.; Arirachakaran, P.; Poovorawan, Y.; Dahlen, G.; Wade, W.G. The oral microbiome in human immunodeficiency virus (HIV)-positive individuals. J. Med. Microbiol. 2015, 64, 1094–1101. [Google Scholar] [CrossRef]
- Beck, J.M.; Schloss, P.D.; Venkataraman, A.; Twigg, H., III; Jablonski, K.A.; Bushman, F.D.; Campbell, T.B.; Charlson, E.S.; Collman, R.G.; Crothers, K.; et al. Multicenter Comparison of Lung and Oral Microbiomes of HIV-infected and HIV-uninfected Individuals. Am. J. Respir. Crit. Care Med. 2015, 192, 1335–1344. [Google Scholar] [CrossRef]
- Frisan, T.; Nagy, N.; Chioureas, D.; Terol, M.; Grasso, F.; Masucci, M.G. A bacterial genotoxin causes virus reactivation and genomic instability in Epstein-Barr virus infected epithelial cells pointing to a role of co-infection in viral oncogenesis. Int. J. Cancer 2019, 144, 98–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gonzalez, O.A.; Li, M.; Ebersole, J.L.; Huang, C.B. HIV-1 reactivation induced by the periodontal pathogens Fusobacterium nucleatum and Porphyromonas gingivalis involves Toll-like receptor 2 [corrected] and 9 activation in monocytes/macrophages. Clin. Vaccine Immunol. 2010, 17, 1417–1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishengallis, G.; Lamont, R.J. Beyond the red complex and into more complexity: The polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol. Oral Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lloret, A.; Egberink, H.; Addie, D.; Belák, S.; Boucraut-Baralon, C.; Frymus, T.; Pennisi, M.G.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; et al. Capnocytophaga Canimorsus Infection in Cats: ABCD guidelines on prevention and management. J. Feline Med. Surg. 2013, 15, 588–590. [Google Scholar] [CrossRef] [PubMed]
- Dolieslager, S.M.; Bennett, D.; Johnston, N.; Riggio, M.P. Novel bacterial phylotypes associated with the healthy feline oral cavity and feline chronic gingivostomatitis. Res. Vet. Sci. 2013, 94, 428–432. [Google Scholar] [CrossRef] [PubMed]
- Lloret, A.; Egberink, H.; Addie, D.; Belak, S.; Boucraut-Baralon, C.; Frymus, T.; Gruffydd-Jones, T.; Hartmann, K.; Hosie, M.J.; Lutz, H.; et al. Pasteurella multocida infection in cats: ABCD guidelines on prevention and management. J. Feline Med. Surg. 2013, 15, 570–572. [Google Scholar] [CrossRef]
- Love, D.N.; Vekselstein, R.; Collings, S. The obligate and facultatively anaerobic bacterial flora of the normal feline gingival margin. Vet. Microbiol. 1990, 22, 267–275. [Google Scholar] [CrossRef]
- Whyte, A.; Gracia, A.; Bonastre, C.; Tejedor, M.T.; Whyte, J.; Monteagudo, L.V.; Simon, C. Oral Disease and Microbiota in Free-Roaming Cats. Top. Companion Anim Med. 2017, 32, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Qiqiang, L.; Huanxin, M.; Xuejun, G. Longitudinal study of volatile fatty acids in the gingival crevicular fluid of patients with periodontitis before and after nonsurgical therapy. J. Periodontal. Res. 2012, 47, 740–749. [Google Scholar] [CrossRef]
- Rzeznik, M.; Triba, M.N.; Levy, P.; Jungo, S.; Botosoa, E.; Duchemann, B.; Le Moyec, L.; Bernaudin, J.F.; Savarin, P.; Guez, D. Identification of a discriminative metabolomic fingerprint of potential clinical relevance in saliva of patients with periodontitis using 1H nuclear magnetic resonance (NMR) spectroscopy. PLoS ONE 2017, 12, e0182767. [Google Scholar] [CrossRef] [Green Version]
- Niederman, R.; Buyle-Bodin, Y.; Lu, B.Y.; Robinson, P.; Naleway, C. Short-chain carboxylic acid concentration in human gingival crevicular fluid. J. Dent. Res. 1997, 76, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Jeng, J.H.; Chan, C.P.; Ho, Y.S.; Lan, W.H.; Hsieh, C.C.; Chang, M.C. Effects of butyrate and propionate on the adhesion, growth, cell cycle kinetics, and protein synthesis of cultured human gingival fibroblasts. J. Periodontol. 1999, 70, 1435–1442. [Google Scholar] [CrossRef] [PubMed]
- Hardham, J.M.; King, K.W.; Dreier, K.; Wong, J.; Strietzel, C.; Eversole, R.R.; Sfintescu, C.; Evans, R.T. Transfer of Bacteroides splanchnicus to Odoribacter gen. nov. as Odoribacter splanchnicus comb. nov., and description of Odoribacter denticanis sp. nov., isolated from the crevicular spaces of canine periodontitis patients. Int. J. Syst. Evol. Microbiol. 2008, 58, 103–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perez-Salcedo, L.; Laguna, E.; Sanchez, M.C.; Marin, M.J.; O’Connor, A.; Gonzalez, I.; Sanz, M.; Herrera, D. Molecular identification of black-pigmented bacteria from subgingival samples of cats suffering from periodontal disease. J. Small Anim. Pract. 2015, 56, 270–275. [Google Scholar] [CrossRef] [PubMed]
- Adler, C.J.; Malik, R.; Browne, G.V.; Norris, J.M. Diet may influence the oral microbiome composition in cats. Microbiome 2016, 4, 23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harley, R.; Gruffydd-Jones, T.J.; Day, M.J. Salivary and serum immunoglobulin levels in cats with chronic gingivostomatitis. Vet. Rec. 2003, 152, 125–129. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet. J. 2011, 17, 3. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahe, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. UCHIME improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.; Pedregosa, F.; Eickenberg, M.; Gervais, P.; Mueller, A.; Kossaifi, J.; Gramfort, A.; Thirion, B.; Varoquaux, G. Machine learning for neuroimaging with scikit-learn. Front. Neuroinform. 2014, 8, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, G.M.; Maffei, V.J.; Zaneveld, J.; Yurgel, S.N.; Brown, J.R.; Taylor, C.M.; Huttenhower, C.; Langille, M.G.I. PICRUSt2: An improved and extensible approach for metagenome inference. bioRxiv 2019. [Google Scholar] [CrossRef] [Green Version]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lozupone, C.; Knight, R. UniFrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-5. 2019. Available online: https://cran.r-project.org/web/packages/vegan/vegan.pdf (accessed on 24 April 2020).
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R. Stat. Soc. Ser. B (Stat. Methodol.) 1995, 57, 289–300. [Google Scholar] [CrossRef]



© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Older, C.E.; Gomes, M.d.O.S.; Hoffmann, A.R.; Policano, M.D.; Reis, C.A.C.d.; Carregaro, A.B.; Ambrósio, C.E.; Carregaro, V.M.L. Influence of the FIV Status and Chronic Gingivitis on Feline Oral Microbiota. Pathogens 2020, 9, 383. https://doi.org/10.3390/pathogens9050383
Older CE, Gomes MdOS, Hoffmann AR, Policano MD, Reis CACd, Carregaro AB, Ambrósio CE, Carregaro VML. Influence of the FIV Status and Chronic Gingivitis on Feline Oral Microbiota. Pathogens. 2020; 9(5):383. https://doi.org/10.3390/pathogens9050383
Chicago/Turabian StyleOlder, Caitlin E., Márcia de Oliveira Sampaio Gomes, Aline Rodrigues Hoffmann, Mariel Dalmédico Policano, Camila Aparecida Cruz dos Reis, Adriano Bonfim Carregaro, Carlos Eduardo Ambrósio, and Valéria Maria Lara Carregaro. 2020. "Influence of the FIV Status and Chronic Gingivitis on Feline Oral Microbiota" Pathogens 9, no. 5: 383. https://doi.org/10.3390/pathogens9050383