A Review on Rhubarb-Derived Substances as Modulators of Cardiovascular Risk Factors—A Special Emphasis on Anti-Obesity Action


Department of General Biochemistry, Faculty of Biology and Environmental Protection, University of Lodz, 90-236 Lodz, Poland
*
Author to whom correspondence should be addressed.
Nutrients 2022, 14(10), 2053; https://doi.org/10.3390/nu14102053
Submission received: 11 April 2022
/
Revised: 9 May 2022
/
Accepted: 11 May 2022
/
Published: 13 May 2022
(This article belongs to the Section Phytochemicals and Human Health)
Abstract
:The currently available anti-obesity therapies encounter many associated risks and side effects often causing the ineffectiveness of treatment. Therefore, various plant-derived substances have been extensively studied as a promising support or even an alternative for existing anti-obesity therapies. This review is dealing with the anti-obesity potential of edible and ethnomedicinal rhubarb species and emerging possible role of the rhubarb-derived extracts or individual compounds in the prevention of obesity and perspectives for their use in an anti-obesity treatment. A special emphasis is put on the most popular edible specimens, i.e., Rheum rhabarbarum L. (garden rhubarb) and Rheum rhaponticum L. (rhapontic rhubarb, Siberian rhubarb); however, the anti-obesity potential of other rhubarb species (e.g., R. officinale, R. palmatum, and R. emodi) is presented as well. The significance of rhubarb-derived extracts and low-molecular specialized rhubarb metabolites of diversified chemical background, e.g., anthraquinones and stilbenes, as potential modulators of human metabolism is highlighted, including the context of cardiovascular disease prevention. The available reports present multiple encouraging rhubarb properties starting from the anti-lipidemic action of rhubarb fibre or its use as purgative medicines, through various actions of rhubarb-derived extracts and their individual compounds: inhibition of enzymes of cholesterol and lipid metabolism, targeting of key molecular regulators of adipogenesis, regulators of cell energy metabolism, the ability to inhibit pro-inflammatory signalling pathways and to regulate glucose and lipid homeostasis contributing to overall in vivo and clinical anti-obesity effects.
1. Introduction
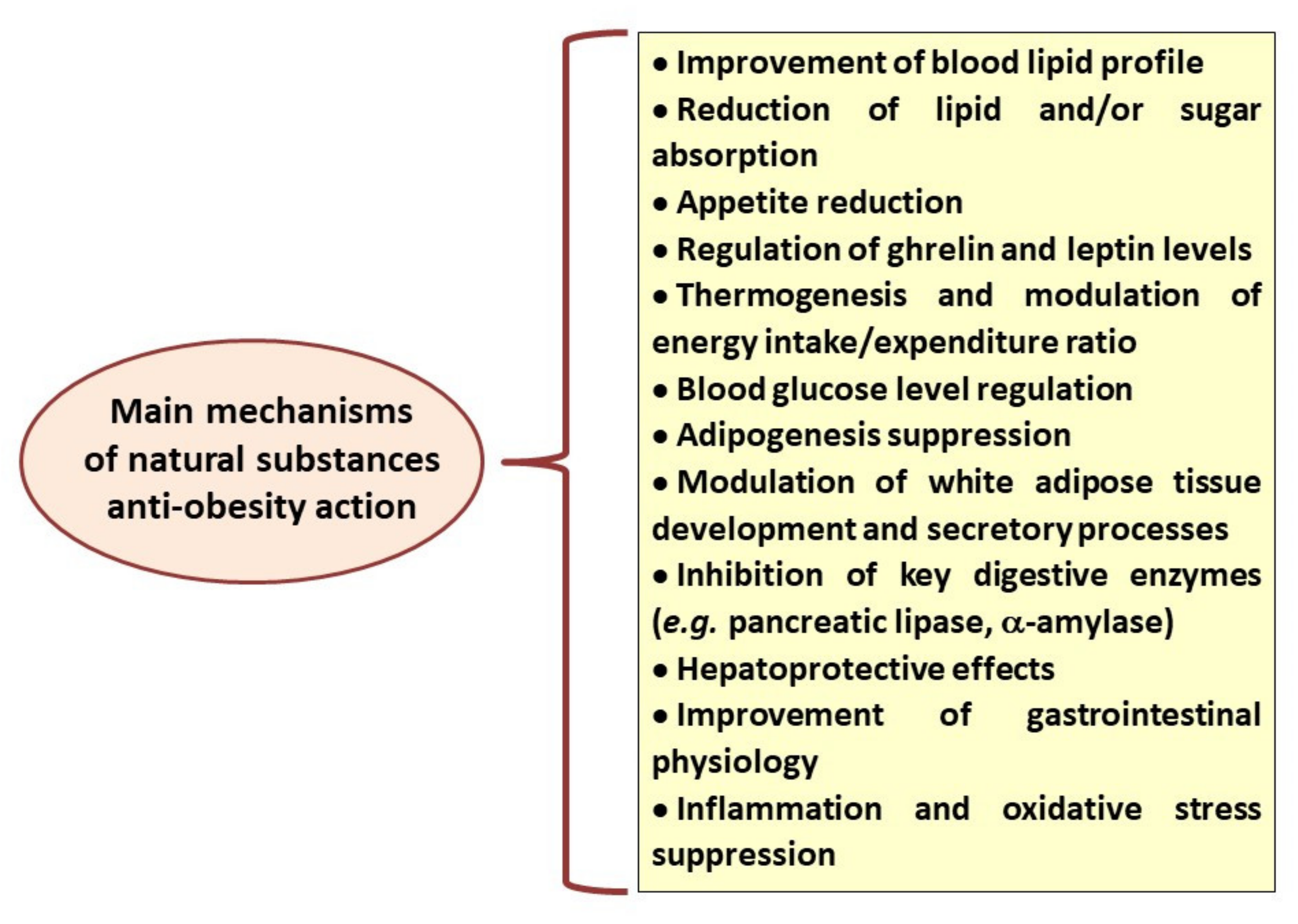
According to data reported by the World Health Organization, the number of obese subjects has nearly tripled since 1975, leading to more than 1.9 billion overweight adults in 2016 [1]. Obesity has become the largest global chronic health problem in Western civilization, and the available anti-obesity therapies are limited by many side effects, risks associated with surgical interventions and obesity relapse [2]. For that reason, lifestyle factors such as balanced diet and physical activity remain fundamental both at the stage of obesity prevention and treatment. In addition, many studies suggest that some of the plant-derived substances and herbal medicines can be helpful anti-obesity agents [3]. Their beneficial properties have been attributed to various mechanisms of action, including appetite reduction, stimulation of thermogenesis, pancreatic lipase inhibitory effects, reduction in dietary fat absorption, stimulation of lipolysis and reduction in lipogenesis (Figure 1) [4,5].
The present work covers a review of the available literature related to anti-lipidemic properties of the rhubarb-derived extracts and individual substances, use of rhubarbs in the prevention of obesity and prospects for their application in an anti-obesity treatment. A special emphasis is put on bioactive properties of two edible specimens, i.e., Rheum rhabarbarum L./syn. Rheum undulatum L. (garden rhubarb) and Rheum rhaponticum L. (rhapontic rhubarb, Siberian rhubarb) (www.theplantlist.org, accessed on 11 April 2022; www.worldfloraonline.org, accessed on 11 April 2022) as sources of dietary fibre and numerous bioactive (poly)phenolic compounds. However, biological properties of other rhubarb species are also presented and discussed in the context of their anti-lipidemic effects, body weight control and obesity prevention.
The review is based on in vitro and in vivo data (from both animal and human studies), originating from peer-reviewed journals indexed in international databases (i.e., Medline/Pubmed, Scopus, Science Direct/Elsevier and Springer Link/ICM) and published up to April 2022. In regard to the type of experimental design, the inclusion criteria covered conclusive studies with appropriate controls and methodology, with a special emphasis on randomized controlled trials (RCTs). To present general topics or to extend readers’ knowledge of particular aspects, well-written, comprehensive reviews were also included into this work. Editorials, conference communications, commentaries, papers from journals non-indexed in scientific databases as well as articles written in languages other than English were excluded. The main search criteria covered a combination of the “rheum” or “rhubarb” words with “obesity”, “obese” “hypecholesterolemia”, “adipocyte”, “adipogenesis” and “fibre”.
2. Rhubarbs as Ethnomedicinal Plants
The Rheum genus (rhubarb, Polygonaceae) includes over 60 species, commonly known as edible and medicinal plants in Asia, Europe and other regions of the world. Petioles of garden (R. rhabarbarum/R. undulatum) and rhapontic (R. rhaponticum) rhubarbs are primarily used as foods, while their underground parts provide herbal material. The current knowledge of phytochemical profile, ethnomedicinal relevance and main directions of physiological activity of individual compounds and extracts originated from R. rhaponticum and R. rhabarbarum and other rhubarbs have been extensively reviewed elsewhere [6,7,8,9]. Hence, in this paper, these issues have been only briefly mentioned. In traditional medicine, both species are used to alleviate various gastrointestinal disorders. Furthermore, mixtures based on R. rhaponticum were also used to cure heart problems, pulmonary system dysfunctions, and disorders related to the reproductive system, including uterine and breast pains. R. rhabarbarum was administered as a purgative and sedative remedy [9]. R. officinale Baill. (Chinese rhubarb) and R. palmatum L. are well-known medicinal plants widely used in traditional Chinese medicine and in many other regions of the world to treat gastrointestinal disorders, chronic renal failure inflammations and liver diseases [10]. Additionally, Rheum emodi Wall ex. Meissn has been known in Chinese and Ayurveda medicine, useful in the treatment of different types of cancer, ulcers, headaches, diarrhoea and liver disorders [11]. In Turkish folk medicine, R. officinale and R. ribes L. (Syrian rhubarb) are traditionally used for the treatment of obesity [12].
3. Rhubarb-Based Preparations in a Contemporary Pharmaceutical and Food Market
R. officinale (Chinese rhubarb), R. palmatum and other anthraquinone-rich rhubarb species, belonging to typical medicinal rhubarb specimens, are predominantly administered as laxatives [13]. They are commercially available as Radix Rhei or as components of different herbal mixtures, dietary supplements and tea dedicated to alleviating gastrointestinal disorders and to treat constipations.
Petioles of edible rhubarbs such as R. rhaponticum and R. rhabarbarum are used for culinary purposes, while their underground parts are a source of herbal material. The estrogenic activity of R. rhaponticum has been well-documented in animal and clinical studies. It has been primarily attributed to the presence of different hydroxystilbenes and their derivatives, including rhaponticin, desoxyrhaponticin, rhapontigenin, desoxrhapontigenin, resveratrol and piceatannol. A special extract (ERr 731®) prepared from roots of this plant specimen has been registered as a bioactive component of herbal preparation recommended to alleviate menopausal symptoms, as an alternative to the conventional hormone replacement therapy [14]. The stilbene-containing R. rhaponticum extracts are also available in different dietary supplements. In addition, the contemporary food and pharmaceutical market offers numerous dietary supplements based on powdered rhubarb rhizome and fibre. Components of R. rhabarbarum were also reported to bind to estrogen receptors ERα and ERβ [15], but the hormone-like activity of this species has been hitherto demonstrated using only in vitro experimental models.
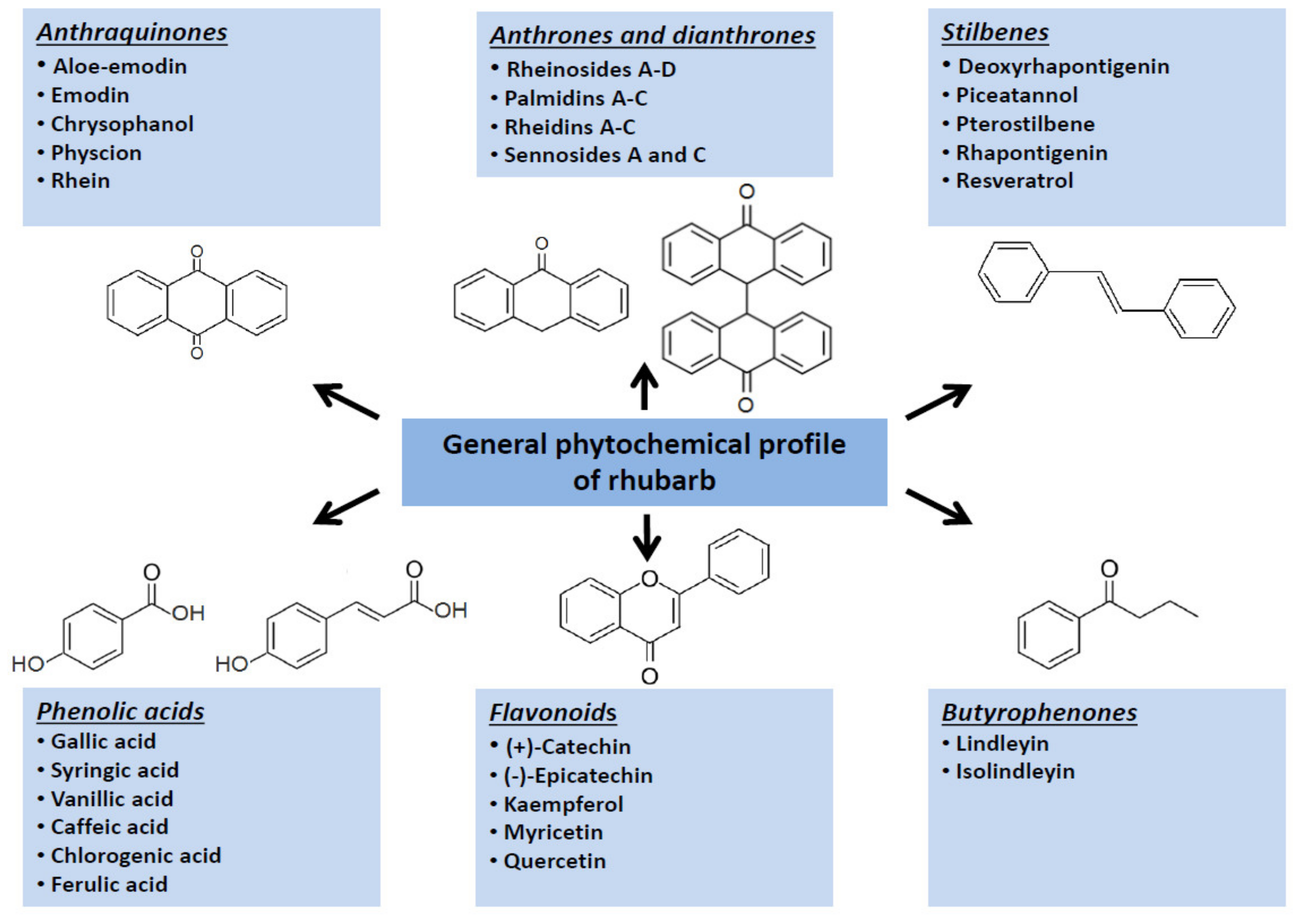
Owing to a protective activity of phytoestrogens on homeostasis [16,17], this activity of rhubarb-extracts, in some extent, may be helpful in maintaining of proper body weight and reduction in incidence and severity of cardiovascular diseases. However, in the context of a direct impact on development and progression of obesity, other molecular mechanisms as well as other bioactive phytochemicals seem to be more crucial. Originally, scientific attention was focused on the rhubarb fibre. Further studies (presented below) extended this approach by findings confirming that low-molecular bioactive metabolites from rhubarbs might affect different metabolic pathways related to obesity development, including the endocrine activity of adipose tissue, lipid metabolism and glucose uptake. Main groups of the low-molecular specialized metabolites synthesized by various rhubarb species are shown in the (Figure 2).
4. Hypolipidemic Effects of Rhubarb: Not Only the Fibre
Edible varieties of rhubarb are considered a rich source of various types of dietary fibre. Comparative analyses of the fibre content in vegetables and fruits revealed that rhubarb stalks contained 26.7% of insoluble fibre, whereas 54.9% were residues after digestion with Driselase. In apple, insoluble fibre amounted to 9.8%, and digestion residue content was 46.2%. For carrot, the comparative values were 10.3% and 34.6%, respectively [18]. The total dietary fibre in a preparation from fresh and frozen rhubarb stalks amounted to 741 g/kg of dry weight (d.w.), including 659 g of insoluble dietary fibre/kg d.w. and 82 g of soluble dietary fibre/kg d.w. [19]. It has been also reported that 140 g of stewed rhubarb provided 3.8 g of the total NSP (non-starch polysaccharides), 2.2 g of insoluble NCP (non-cellulosic polysaccharides) and 0.7 g of soluble NCP fibre [20].
Although the hypolipidemic action of dietary fibre has been well established [21], the use of Rheum species as a source of natural fibre or other lipid-lowering substances is still under examination. Current knowledge of physiological effects of rhubarb-derived fibre is mainly based on results originated from experiments employing animal models of hypercholesterolemia and obesity; however, data from studies on human subjects are also available (for details see Table 1). The R. rhaponticum fibre has been shown to effectively decrease plasma cholesterol in animals fed a high-cholesterol diet, although its effect on cholesterol synthesis in hepatocytes was not found [22]. As evidenced, the fibre may influence secretory processes in the liver and gallbladder functions by both direct and indirect mechanisms. Directly, it binds bile acids [23]. The binding rate of taurocholic acid (TCA) for rhubarb fibre was established to be 6.2 µmol/100 mg and increased linearly with the fibre concentration. For a comparison, the TCA-binging rate for cereal bran ranged between 2.3 and 3.5 µmol/100 mg, and for cellulose, this rate amounted to 1.0 µmol TCA/100 mg [24]. The bile-binding rate for the different digested cereal products and alcohol-insoluble substances from fruits and vegetables such as apples, strawberries, rowan berries, carrots, white cabbage, red beets and sugar beet pulp attained 1.21–1.77 µmol bile acids/100 mg of the examined preparation [25].
Indirectly, rhubarb fibre regulates cholesterol metabolism. In vivo examination revealed a stimulatory effect of rhubarb fibre on the expression of cholesterol 7α-hydroxylase gene and excretion of bile acid in cholesterol-fed C57BL/6J mice [19]. Furthermore, the hypocholesterolemic effect of rhubarb fibre was also observed in hypercholesterolemic men, consuming 27 g of rhubarb stalk fibre daily for 4 weeks [23].
Contrary to the above results, a study on diabetic rats maintained on a diet rich in rhubarb fibre, i.e., 50 g of rhubarb stalk fibre/kg of body weight (b.w.), did not demonstrate a beneficial effect on the plasma cholesterol or triacylglycerol concentrations in the examined animals [26]. Moreover, the literature clearly indicates that the lipid-lowering action of rhubarb may be also dependent on the presence of other constituents, i.e., low -molecular phytochemicals as well as synergistic action of different types of phytocompounds. Based on studies from the 1990s, demonstrating the cholesterol-lowering properties of pectins [27,28,29], the hypolipidemic properties of rhubarb were strictly attributed to its fibre content. Later decades revised this point of view and revealed that stilbene compounds such as resveratrol [30,31,32], rhaponticin and rhapontigenin [33,34] display anti-hyperlipidemic effects in vivo (Table 2). Administration of rhaponticin and rhapontigenin to animals fed a high-cholesterol diet significantly improved the blood lipid profile and reduced the extent of pathological changes in fatty liver [33].
Among reports dealing with the anti-obesity properties of stilbene-type compounds, the action of resveratrol has been described and evidenced in the widest extent of examinations, including clinical trials. Although the anti-obesity [35,36] effects of resveratrol appeared to be a very promising research trend, results derived from studies involving human subjects are inconclusive, or even, in some cases, research hypotheses have failed [37,38,39,40,41,42,43,44,45,46,47,48,49,50,51,52] (Table 3). The synergistic activity of resveratrol and other substances (either polyphenolics [53,54] or drugs [55]) was examined to find an effective combination for treatment of obesity or its complications. In a randomized controlled trial, study participants were assigned into one of four groups, i.e., placebo, resveratrol (100 mg), orlistat (120 mg), or a combination of orlistat-resveratrol (O-R; 120 mg + 100 mg) group. After 6 months, results obtained for the resveratrol monotherapy group did not significantly differ from the placebo group. The anti-obesity action and weight loss were found in both the orlistat and O-R groups. However, the most effective reduction in BMI, fat mass and waist circumference was found in subjects taking the O-R combination [55].
Data obtained for the stilbenoids are also supported by results from studies on other individual compounds that are present in the rhubarb profile, e.g., aloe-emodin [56] and physcion [57]. Additionally, emodin displayed a hypocholesterolemic effect in vitro and in vivo, through the capability of bile salts binding and increasing the expression of cholesterol 7-alpha-monooxygenase (CYP7A1) [58]. In obese mice, emodin (40 or 80 mg/kg b.w., for 6 weeks) modulated the adipose tissue physiology, and decreased the body weight and the blood lipid level. As was demonstrated by the increased levels of markers of beige adipocytes (i.e., Cd137, Tmem26 and Tbx1 mRNA), the browning processes were stimulated in the white adipose tissue [59].
5. Molecular Targets for Rhubarb-Derived Substances
5.1. Inhibitory Effects on Key Enzymes Related to Lipid Absorption and Metabolism
Administration of different types of anti-obesity drugs have been proven to have a wide spectrum of side effects, including headache, insomnia, nervousness, constipation, hypertension as well as an increased risk of cardiovascular complications. For example, due to the prevalence of risk of adverse effects over the benefits, the European Medicines Agency (EMA) recommended the removal of sibutramine from the pharmaceutical market in the European Union after only several years of use [60]. Among synthetic anti-obesity drugs, orlistat (tetrahydrolipstatin), an inhibitor of pancreatic lipase (EC 3.1.1.3), is the only currently approved weight loss medicine for the long-term treatment of obesity [61,62]. The molecular target of this drug is the pancreatic lipase (PL), which is essential for digestion of dietary triglycerides. For that reason, the search for new and more effective inhibitors (including natural compounds) of this hydrolytic enzyme has gained increasing attention [63]. Moreover, numerous reports demonstrated the ability of phytochemicals belonging to various classes to inhibit PL activity [64,65,66,67,68,69,70,71,72,73]. Most of the examined plant-derived substances were weaker inhibitors of the pancreatic lipase than the reference drug, but some of them, i.e., ferulic acid and kaempferol-3-O-rutinoside, displayed comparable PL-inhibitory efficiency. Some of these potential inhibitors of PL, e.g., phenolic acids as well as quercetin and kaempferol derivatives, are present in rhubarb species (Table 4). Synergistic interactions of different compounds in the inhibition of 3T3-L1 PL activity and preadipocyte differentiation have been also evidenced [74]. An PL inhibitor screening of the 37 plant extracts revealed that only 6 of them had moderate or strong anti-lipase activity (more than 30% of inhibition). Extracts from Prunella vulgaris L. and Rheum palmatum were identified as the two most potent inhibitors, displaying 74.7% and 53.8% of inhibitory effectiveness towards PL, respectively [75]. In another screening, a methanolic extract from R. ribes rhizome inhibited the PL activity by 43%. The most effective plant preparation in that study, i.e., Quercus infectoria G. Olivier galls, reduced the PL activity by 85% [76]. Furthermore, galloyl glucosides and galloylproanthocyanidins isolated from rhubarb (R. palmatum) roots (Rhei Rhizoma) were described as promising inhibitors (acting at micromolar concentrations) of squalene epoxidase, a key enzyme of the cholesterol biosynthesis pathway [77].
According to the literature, rhubarb contains natural inhibitors of soluble epoxide hydrolase (sEH), which is believed to be one of the most important molecular targets in the therapy of cardiovascular diseases as well as other disorders. sEH is a major enzyme responsible for the hydrolysis of epoxy-fatty acids (incl. arachidonic, linoleic, eicosapentaenoic, and docosahexaenoic acid epoxides). The plant-derived compounds or extracts have been examined as potential inhibitors of this enzyme. For example, ethanol extract of Sophora flavescens Aiton roots inhibited this enzyme with IC50 = 2.07 µg/mL [78]. sEH-inhibitory effects were also found for phenolic compounds isolated from the rhizomes and roots of Gentiana scabra Bunge [79], extracts from aerial parts of Tetrastigma hemsleyanum (King) Chantaran. and J. Parn. [80] and compounds isolated from roots of Lycopus lucidus Turcz. ex Benth. [81].
Both the methanol extract, n-hexane, chloroform and butanol fractions from R. rhabarbarum displayed sEH-inhibitory efficiency, but mechanistic analyses revealed evident diversity in actions of individual components of the tested extracts. Piceatannol 3′-O-β-D-glucopyranoside was found to be a competitive sEH inhibitor. In contrast, resveratrol, desoxyrhaponticin, rhaponticin, isorhapontin, desoxyrhapontigenin, along with other compounds such as astringin, chrysophanol-8-O-β-D-glucopyranoside and aloe emodin acted as the mixed-type inhibitors. Rhapontigenin and emodin were non-competitive inhibitors of sEH. Among the examined compounds, astringin was the most effective sEH inhibitor (IC50 = 2.5 ± 0.5 µM), and the weakest one was desoxyrhaponticin (IC50 = 53.2 ± 4.4 µM) [82]. Unfortunately, in the case of the search for the rhubarb-derived inhibitors of sEH, only preliminary in vitro results are available, although similar works on sEH inhibitors from other plants are more advanced and have reached the level of in vivo studies [83].
5.2. Modulation of the Adipose Tissue Physiology
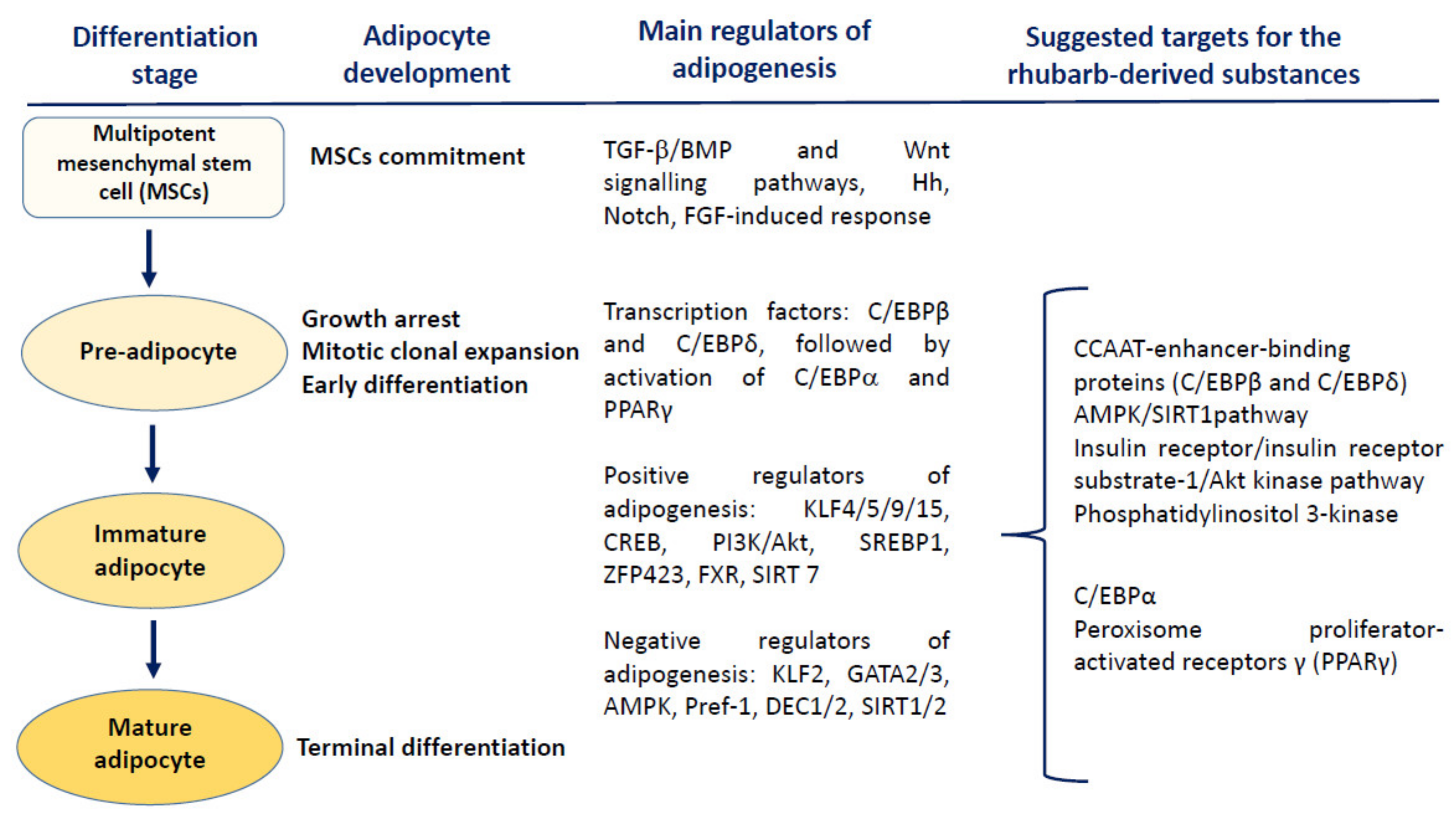
Although the energy storage is commonly believed to be the main role of white adipose tissue, current knowledge of human physiology evidently indicates its important endocrine activity, influence on numerous molecular pathways and modulatory effect on homeostasis [84]. Studies on anti-obesity actions of various natural substances and herbal remedies are focused on several points that may be critical in the regulation of either adipogenesis or secretory activity of the adipocytes. During this multistep process, phytochemicals (including compounds present in rhubarbs) may interact at the mitotic clonal expansion, early differentiation and terminal differentiation stages of adipogenesis (Figure 3). Due to multifactorial etiology and pathophysiology of obesity, many mechanisms underlying the anti-obesity effects of phytochemicals still remain only partly elucidated. Existing evidence indicates that the main targets for most plant-derived anti-obesity agents are key regulators of adipogenesis such as CCAAT/enhancer-binding proteins (C/EBPs) and the peroxisome proliferator-activated receptors (PPARs), as well as the AMP-activated protein kinase (AMPK), a regulator of energy metabolism [85,86,87]. An additional aspect of the beneficial action of phytochemicals in obese subjects may be their anti-inflammatory properties, including the ability to inhibit the nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB)-triggered signalling [88]. Exemplary molecular targets for phytochemicals are the phosphatidylinositol 3-kinase and Akt/Protein Kinase B (PI3K/Akt) pathway, the IκBα kinase/c-Jun N-terminal kinase (IKK/JNK), and Janus kinase/signal transducers and activators of transcription (JAK/STAT)-mediated response [89].
The literature provides very few data originating from studies on rhubarb extracts; however, some more information can be deduced from results reported by works on individual phytochemicals that are present in these plants. Inhibitory effects of natural substances on adipogenesis or metabolic activity of adipose tissue were found for compounds belonging to diverse phytochemical classes, e.g., stilbenes and anthraquinones. The newest literature emphasizes a pleiotropic activity of resveratrol, including the modulation of gene expression and affecting diverse molecular targets such as AMPK, the peroxisome proliferator-activated receptor co-activator-1α (PGC-1α) as well as sirtuin-1 (SIRT1) deacetylase [90]. The anti-adipogenic effects of another stilbene, i.e., piceatannol, seem to be mainly targeted to early phases of development of adipose tissue. The anti-adipogenic properties of piceatannol involved reduction in the mitotic clonal expansion, decrease in C/EBPβ, PPARγ, and C/EBPα mRNA levels and inhibition of the insulin receptor-dependent signalling (the insulin receptor/insulin receptor substrate-1/Akt kinase pathway) in 3T3-L1 preadipocytes. Piceatannol was an inhibitor of the insulin receptor kinase and phosphatidylinositol 3-kinase (PI3K) activity [91]. Its ability to suppress the macrophage interactions with adipocytes may be an important mechanism of alleviation of inflammatory processes in the adipose tissue [92]. Moreover, recent data suggest that piceatannol may be a more effective inhibitor of adipogenesis in human visceral adipose-derived stem cells than resveratrol [93].
Among rhubarb hydroxyanthraquinones, rhein is currently considered as the most promising anti-adipogenic and lipolysis-stimulating factor. The anti-obesity action of rhein is through different mechanisms, including activity on the transcriptional level, as an inhibitor of adipogenesis, including adipocyte differentiation. It rhein is able to downregulate the expression of adipogenesis-specific transcription factors such as PPARγ and CCAAT-enhancer-binding protein-α (C/EBPα) and their upstream regulator, i.e., CCAAT-enhancer-binding protein-β (C/EBPβ) [94]. The inhibition of PPARγ signalling was indicated as a potential mechanism of the reduction in obesity, fat mass and the size of white and brown adipocyte tissue in animals treated with rhein [95]. Another proposed mechanism of the anti-obesity action of rhein is a regulatory effect on cholesterol homeostasis, through the liver X receptors (LXRs) antagonism [96]. Recent studies on five rhubarb-derived compounds: chrysophanol, aloe emodin, emodin, physcion, and rhein suggested stronger inhibitory effects of emodin and rhein on lipid accumulation in 3T3-L1 adipocytes, compared to the remaining three hydroxyanthraquinones. Both emodin and rhein acted through mitogen-activated protein kinase (MAPK) signalling; however, their influence on adipogenesis and lipid metabolism were diverse. While rhein reduced lipid deposition by modulation of the adipogenic transcriptional factors and lipolytic enzymes, the lipid-lowering activity of emodin involved the reduction in lipogenic enzymes. The aforementioned diversity in molecular actions may result in divergent activity of both compounds in vivo. Experiments on animals demonstrated that rhein was a stronger anti-obesity agent than emodin, and reduced plasma cholesterol by 29% (versus a 14%-decrease caused by emodin) [97].
Danthron, a natural anthraquinone derivative synthesized by rhubarb, has been shown to reduce the obesity and associated metabolic fatty liver diseases (MAFLD) in animals. It has been established that this compound is able to stimulate the binding of the retinoid X receptor alpha (RXRα)/PPARα heterodimer to the promoter of the adiponectin receptor 2 (AdipoR2), resulting in the AMPKα and PPARα pathway activation [98]. The stimulation of the AMPK/SIRT1 pathway has been also suggested as the most likely mechanism of the anti-obesity activity of chrysophanol. In rats receiving a high-fat diet, this anthraquinone was found to stimulate the expression of lipolytic genes and suppress the lipogenic genes as well as to diminish the inflammatory response [99]. Anti-diabetic and anti-obesity effects in vitro and in vivo were also found for emodin. As a selective inhibitor of the 11β-hydroxysteroid dehydrogenase type 1 (11β-HSD1), emodin regulated adipogenesis and energy metabolism in 3T3-L1 adipocytes as well as ameliorated metabolic disorders in ob/ob (B6.V-Lepob/Lepob) mice [100]. In other animal study, the inhibition of proprotein convertase subtilisin/kexin type 9 was indicated as a key mechanism of the cholesterol-lowering effect of aloe-emodin [56].
5.3. Metabolism and Glucose Level Regulation
The ability to regulate glucose homeostasis has been demonstrated for various types of rhubarb-derived substances, i.e., mixtures extracted with the use of water or organic solvents, low-molecular isolates and the soluble fibre. Regarding the latter, it has been demonstrated that a high fibre diet might upregulate the proglucagon mRNA and secretion of both the glucagon-like peptide-1 [GLP-1(7–37)] and insulin [101]. In other work on animals, consumption of rhubarb fibre (50 g/kg of diet) influenced the ileal synthesis of proglucagon mRNA and reduced passive permeability, without affecting the glucose transport of the small intestine [102]. Furthermore, in patients with type 2 diabetes, an administration of essential oil preparation (400 mg capsules, 3 times daily, 90 capsules in total) isolated from rhubarb stem reduced the glycosylated haemoglobin and fasting blood glucose levels [103].
Another possible molecular target for rhubarb-derived substances is the protein tyrosine phosphatase 1B (PTP1B), a negative regulator of the leptin and insulin-dependent signalling pathways. Currently available works devoted to search for effective inhibitors of this enzyme reported examinations of over few hundreds of synthetic and natural compounds, including plant extracts [104,105]. Among them, one of the available medicinal plant screenings revealed that a hot water extract from R. rhabarbarum roots might reduce the PTP1B activity in vitro. Moreover, studies on high-fat diet-fed C57BL/6 mice and in vitro experiments demonstrated the metabolism-regulatory activity of R. rhabarbarum extracts and its anthraquinone compounds—chrysophanol and physcion [106]. Other authors reported the hypoglycaemic effects of Rheum tanguticum Maxim. ex Balf., based on the sucrase and maltase inhibitory activities of this plant metabolites such as flavan ((-)-epicatechin 3-O-gallate) and phenylbutanone (lindleyin) [107].
In animals, the glucose homeostasis regulatory ability was evidenced for both individual compounds and extracts isolated from different rhubarb species (Table 5). Beneficial effects of rhubarb-derived phytochemicals included the insulin-sensitizing activity reduction in the blood triglyceride and glucose [108,109,110,111,112] levels. Interestingly, some caution has been recommended in the case of Rheum turkestanicum, whose rhizomes are a traditional anti-diabetic medicine in Iran. An animal study revealed that this plant rhizome extract had anti-hypertriglyceridemic activity, but no hypoglycemic or hepatoprotective effects were found [113]. The combination of metformin and the anthraquinone glycoside preparation from R. palmatum rhizomes administered to rats (100 or 400 mg/kg b/w., for 6 weeks) ameliorated type 2 diabetes mellitus through the regulation of the gut microbiota and activation of the GLP-1/cAMP pathway, resulting in a decrease in insulin resistance [114]. A clinical trial on patients with type 2 diabetes mellitus demonstrated the reduction in fasting blood glucose level and glycosylated haemoglobin in the intervention group, treated with capsules containing rhubarb stem extract (90 capsules in total; 400 mg/capsule, three times a day) [103].
5.4. Anti-Obesity Action of Rhubarb Extracts in Animal Studies
The rhubarb extract (100 mg/kg b.w., administered for 8 weeks) modulated lipid metabolism (including a stimulation of adiponectin synthesis) and significantly reduced the body weight gain in the examined mice. In parallel experiments, the anthraquinones (30 µM) inhibited PTP1B activity and enhanced the insulin sensitivity in the serum-starved 3T3L1 cells [115]. In rats treated with a 690 mg/kg b.w. dose (for 6 weeks) of the R. palmatum extract (a concentrated decoction), established based on recommended clinical doses for humans (according to the Pharmacopoeia of the People’s Republic of China), a high-fat-diet-induced hepatosteatosis was significantly alleviated. The treatment significantly lowered the liver triglyceride levels, liver weight and improved glucose tolerance; however, a high dose of the extract, i.e., 1300 mg/kg b.w., was less efficient. The stimulating AMPK activity of the examined extract has been suggested as the most probable molecular mechanism of the observed effects [116].
The treatment with R. undulatum extracts not only reduced the low-density-lipoprotein cholesterol and increased the high-density-lipoprotein cholesterol in rats, but also mitigated vascular inflammation as confirmed by measurements of the vascular NF-κB-p65, adhesion molecules ICAM-1 and VCAM-1 as well as E-selectin levels [117]. In an animal model of atherosclerosis, the lipid-lowering and plaque-stabilizing properties associated with anti-inflammatory effects were found for aqueous extract from R. officinale [118]. A four-week therapy with hydroalcoholic extract from R. ribes L. root (150 mg/kg b.w./day) reduced the glucose, cholesterol and triglyceride levels in diabetic rats. Moreover, the reduction in low-density-lipoprotein (LDL) level achieved concentrations comparable to those recorded in the control group [119]. Likewise, the cholesterol-lowering effects were observed in rabbits having hypercholesterolemia, after a treatment (4 g/kg b.w./day, for 2 weeks) with ethanolic and aqueous extracts from R. ribes stalks [120]. Recent animal studies indicate that rhubarb supplementation may prevent diet-induced obesity. In C57BL6/J mice fed a high-fat and high-sucrose (HFHS) diet, enriched with 0.3% (g/g) of R. palmatum extract (for 8 weeks), the accumulated fat mass and weight gain were reduced, when compared to animals fed an HFHS diet, without the plant extract. Furthermore, in mice supplemented with the rhubarb extract, fat pads (visceral, epididymal and subcutaneous) were similar to those found in control animals (mice fed a control diet—D12450H; research diet). Rhubarb supplementation also resulted in the prevention of diet-induced liver steatosis as well as in increased hepatic cholesterol and inflammatory markers and beneficial effects on adipose tissue, including a trend towards a decreased infiltration by macrophages. Immunohistological analyses demonstrated that adipocyte morphology in mice supplemented with rhubarb extract was similar to adipocytes from control animals. In addition, those effects were associated with an increased amount of Akkermansia muciniphila in the colon [121], which is a commensal bacterium of the gut mucus, displaying beneficial effects for basal metabolism and associated with reduced risk of obesity [122].
5.5. Anti-Obesity Action of Rhubarb Extracts in Clinical Trials
A randomized, double-blind and placebo-controlled trial involving 83 patients with diagnosed atherosclerosis revealed that treatment with R. officinalis hot water extract (50 mg/kg b.w.) decreased the serum total cholesterol and the LDL cholesterol [123]. In contrast, administration of the R. emodi stem extract to patients with type 2 diabetes mellitus (taking a total amount of 90 extract capsules: 3 capsules/day, 400 mg of the extract/capsule) did not reveal any effect on body weight; however, a decrease in systolic and diastolic blood pressure was observed [124].
5.6. Laxative Effects of Rhubarb-Based Extracts and Preparations
In Western civilization, laxative abuse as a method supporting the weight loss and adverse effects of these substances constitutes a serious problem [125]. On the other hand, the laxative properties of natural compounds are an important element of the treatment of constipation associated with various diseases. The purgative effects of rhubarb are a result of the presence of different substances, mainly anthraquinones. For that reason, the anthraquinone-rich rhubarb specimens such as R. officinale and R. palmatum are often used as purgative medicines. The anthraquinone concentration in roots of these two rhubarb species typically ranges up to about 5% of dry mass [126,127]. Other purgative rhubarb components are anthrones and dianthrone compounds, including rheinosides A–D; palmidin A, B and C; rheidin A, B and C; as well as sennosides A–F [128]. In Asian countries, R. emodi is also traditionally recommended for purgative therapy. Interestingly, while larger doses of R. emodi are a natural laxative, at small doses, this plant was administered to treat dysentery and diarrhea [129].
According to the literature, the presence of anthraquinone glycosides with 1,8-dihydroxy groups and without hydroxyl groups in the 2, 3, 6 and 7 positions may be responsible for the effect of “watery diarrhea”, occurring after using of some rhubarb-based treatment [130]. Rhubarb contains both free anthraquinones (e.g., aloe-emodin, emodin, rhein, chrysophanol and physcion) and conjugated forms, containing the β-glycoside bonds. The anthraquinone conjugates are considered to be the exact medicinal components of the rhubarb-derived material. The presence of the β-type glycoside bonds in conjugates prevents their hydrolysis by α-glucosidase in the upper gastrointestinal tract and, as a consequence, they are hydrolysed to free anthraquinones by bacterial β-glucosidases in the colon. Recently, a Chinese research group designed oral colon-specific drug delivery granules, containing the total free anthraquinones to improve their purgative effect and to reduce anthraquinone nephrotoxicity [131,132]. The postprandial hyperlipidemia-lowering effect and improvement in gastrointestinal transit in diabetic rats was reported for aqueous extract from the rhizome of R. palmatum, administered at doses of 150 and 300 mg/kg b.w. The anthrone content in the examined rhubarb extract was not less than 0.5% [133]. A comparative study of three R. palmatum-containing traditional Chinese purgative medicines Ta-Cheng-Chi-Tang (TCCT), Xiao-Chen-Chi-Tang (XCCT) and Tiao-Wei-Chen-Chi-Tang (TWCCT) did not reveal their weight-lowering effects in rats fed on a high-fat diet. However, the examined extracts improved the blood serum lipid profile. In addition, both XCCT and TWCCT significantly attenuated hypercholesterolemia and the TWCCT preparation also reduced hypertriglyceridemia in rats [134].
Rhubarbs with lower content of anthraquinones (e.g., R. rhabarbarum) display mild laxative effects, but they also are used for production of different dietary supplements and food products dedicated to improving the intestine physiology and to prevent constipation. These products are available in a form of herbal mixtures, capsules or even candy-like bars enriched with the plant-derived fibre and low-molecular phytochemicals.
6. Concluding Remarks
Disrupted balance between an amount of taken calories and energy expenditure as well as complex physiological activity of both white and brown adipose tissue are essential pro-obesity factors. Different rhubarb species have been demonstrated to display a wide range of biological activities that may be relevant for reduction in cardiovascular risks, including improvement in glucose and lipid metabolism. The original findings on the health-promoting action of the rhubarb-derived substances were typically focused on the cholesterol-lowering role of the fibre. Currently, it is known that also other, low-molecular phytochemicals may be important for the cardioprotective, lipid-lowering and anti-adipogenic effects of these plants. However, the pharmacological significance of the rhubarb-based preparations in a context of the anti-obesity action still remains only partly recognized.
Literature evidence has indicated that the rhubarb-derived compounds and extracts may act at different molecular and physiological levels. The pathophysiology of obesity is multifactorial and includes many undesirable changes, such as alterations in blood lipid profile, enhanced adipogenesis, increased body fat mass, hyperglycemia, chronic inflammation in adipose tissue, oxidative stress, and others. Results from different types of examinations have demonstrated that rhubarb-derived substances have some potential to reduce cardiovascular risks. On the other hand, it should be noted that many works devoted to anti-obesity action of rhubarb-derived substances have stuck at a level of in vitro tests; thus, a reliable evaluation of their physiological and pharmacological effects is not possible yet. Moreover, findings from studies on individual phytochemicals (e.g., works on the anti-obesity properties of resveratrol) cannot be directly extrapolated to the effects of rhubarb preparations containing these phytochemicals. In addition, there is negligible evidence of using rhubarb extracts combined with other natural or synthetic substances (including drugs). The synergistic action of different substances seems to be a very promising research trend, and this approach could give better results. For example, a reduction in body weight gain was observed in an animal study on different combinations of extracts originating from other plants (i.e., preparations from leaves from Phyllostachys pubescens J. Houz. and roots of Scutellaria baicalensis Georgi) [135].
Furthermore, most in vivo studies have been conducted using animal models of obesity. The available data from clinical studies provide only a fragmentary insight into the pharmacological potential of these plants. Thus, the above-presented results point to the need for further and more extensive studies using standardized rhubarb extracts/preparations aimed at establishing their exact anti-obesity relevance in vivo and in humans.
Funding
This research was funded by the University of Lodz grant number [B2211 000 000 044.01].
Conflicts of Interest
The authors declare no conflict of interest.
References
- World Health Organization. Obesity and Overweight. 2022. Available online: https://www.who.int/news-room/fact-sheets/detail/obesity-and-overweight (accessed on 2 February 2022).
- Williams, D.M.; Nawaz, A.; Evans, M. Drug therapy in obesity: A review of current and emerging treatments. Diabetes Ther. 2020, 11, 1199–1216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bagherniya, M.; Mahdavi, A.; Abbasi, E.; Iranshahy, M.; Sathyapalan, T.; Sahebkar, A. The effects of phytochemicals and herbal bio-active compounds on tumour necrosis factor-α in overweight and obese individuals: A clinical review. Inflammopharmacology 2022, 30, 91–110. [Google Scholar] [CrossRef] [PubMed]
- Karri, S.; Sharma, S.; Hatware, K.; Patil, K. Natural anti-obesity agents and their therapeutic role in management of obesity: A future trend perspective. Biomed. Pharmacother. 2019, 110, 224–238. [Google Scholar] [CrossRef] [PubMed]
- Payab, M.; Hasani-Ranjbar, S.; Shahbal, N.; Qorbani, M.; Aletaha, A.; Haghi-Aminjan, H.; Soltani, A.; Khatami, F.; Nikfar, S.; Hassani, S.; et al. Effect of the herbal medicines in obesity and metabolic syndrome: A systematic review and meta-analysis of clinical trials. Phytother. Res. 2020, 34, 526–554. [Google Scholar] [CrossRef]
- Cao, Y.-J.; Pu, Z.-J.; Tang, Y.-P.; Shen, J.; Chen, Y.-Y.; Kang, A.; Zhou, G.-S.; Duan, J.-A. Advances in bioactive constituents, pharmacology and clinical applications of rhubarb. Chin. Med. 2017, 12, 36. [Google Scholar] [CrossRef] [Green Version]
- Xiang, H.; Zuo, J.; Guo, F.; Dong, D. What we already know about rhubarb: A comprehensive review. Chin. Med. 2020, 15, 88. [Google Scholar] [CrossRef]
- Mohtashami, L.; Amiri, M.S.; Ayati, Z.; Ramezani, M.; Jamialahmadi, T. Ethnobotanical uses, phytochemistry and pharmacology of different Rheum species (Polygonaceae): A review. Adv. Exp. Med. Biol. 2021, 1308, 309–352. [Google Scholar]
- Kolodziejczyk-Czepas, J.; Liudvytska, O. Rheum rhaponticum and Rheum rhabarbarum—A review of phytochemistry, biological activities and therapeutic potential. Phytochem. Rev. 2021, 20, 589–607. [Google Scholar] [CrossRef]
- Qin, X.; Wang, Y.; Li, A.; Sun, A.; Yu, L.; Liu, L. Separation and purification of six components from the roots of Rheum officinaleBaill. by supercritical fluid chromatography. J. Liq. Chrom. Relat. Tech. 2017, 40, 156–164. [Google Scholar] [CrossRef]
- Singh, R.; Tiwari, T.; Chaturvedi, P. Rheum emodi Wall ex. meissn (Indian Rhubarb): Highly endangered medicinal herb. J. Med. Plants Stud. 2017, 5, 13–16. [Google Scholar]
- Sargin, S.A. Plants used against obesity in Turkish folk medicine: A review. J. Ethnopharmacol. 2021, 270, 113841. [Google Scholar] [CrossRef]
- Sun, Y.; Chen, X.; Wang, Y. Meta analysis of umbilical area applying with rhubarb in treatment of constipation. Chin. Nurs. Res. 2016, 30, 4020–4024. [Google Scholar]
- Chang, J.-L.; Montalto, M.B.; Heger, P.W.; Thiemann, E.; Rettenberger, R.; Wacker, J. Rheum rhaponticum extract (ERr 731): Postmarketing data on safety surveillance and consumer complaints. Integr. Med. 2016, 15, 34–39. [Google Scholar]
- Park, S.; Kim, Y.N.; Kwak, H.J.; Jeong, E.J.; Kim, S.H. Estrogenic activity of constituents from the rhizomes of Rheum undulatumLinné. Bioorg. Med. Chem. Lett. 2018, 28, 552–557. [Google Scholar] [CrossRef]
- AsokanShibu, M.; Kuo, W.W.; Kuo, C.H.; Day, C.H.; Shen, C.Y.; Chung, L.C.; Lai, C.H.; Pan, L.F.; Vijaya Padma, V.; Huang, C.Y. Potential phytoestrogen alternatives exert cardio-protective mechanisms via estrogen receptors. Biomedicine 2017, 7, 11. [Google Scholar] [CrossRef] [Green Version]
- Domínguez-López, I.; Yago-Aragón, M.; Salas-Huetos, A.; Tresserra-Rimbau, A.; Hurtado-Barroso, S. Effects of dietary phytoestrogens on hormones throughout a human lifespan: A review. Nutrients 2020, 12, 2456. [Google Scholar] [CrossRef]
- Bunzel, M.; Seiler, A.; Steinhart, H. Characterization of dietary fiber lignins from fruits and vegetables using the DFRC method. J. Agric. Food Chem. 2005, 53, 9553–9559. [Google Scholar] [CrossRef]
- Goel, V.; Cheema, S.K.; Agellon, L.B.; Ooraikul, B.; Basu, T.K. Dietary rhubarb (Rheum rhaponticum) stalk fibre stimulates cholesterol 7a-hydroxylase gene expression and bile acid excretion in cholesterol-fed C57BL/6J mice. Br. J. Nutr. 1999, 81, 65–71. [Google Scholar] [CrossRef] [Green Version]
- Rana, V.; Bachheti, R.K.; Chand, T.; Barman, A. Dietary fibre and human health. Int. J. Food Saf. Nutr. Public Health 2011, 4, 101–118. [Google Scholar] [CrossRef]
- Kendall, C.W.C.; Esfahani, A.; Jenkins, D.J.A. The link between dietary fibre and human health. Food Hydrocoll. 2010, 24, 42–48. [Google Scholar] [CrossRef]
- Basu, T.K.; Ooraikul, B.; Garg, M. The lipid lowering effects of rhubarb stalk fibre: A new source of dietary fibre. Nutr. Res. 1993, 13, 1017–1024. [Google Scholar] [CrossRef]
- Goel, V.; Ooraikul, B.; Basu, T.K. Cholesterol lowering effects of rhubarb fibre in hypercholesterolemic men. J. Am. Coll. Nutr. 1997, 16, 600–604. [Google Scholar]
- Goel, V.; Cheema, S.K.; Agellon, L.B.; Ooraikul, B.; McBurney, M.I.; Basu, T.K. In vitro binding of bile salt to rhubarb stalk powder. Nutr. Res. 1998, 18, 893–903. [Google Scholar] [CrossRef]
- Dongowski, G. Interactions between dietary fibre-rich preparations and glycoconjugated bile acids in vitro. Food Chem. 2007, 104, 390–397. [Google Scholar] [CrossRef]
- Cheema, S.K.; Goel, V.; Basu, T.K.; Agellon, L.B. Dietary rhubarb (Rheum rhaponticum) stalk fibre does not lower plasma cholesterol levels in diabetic rats. Br. J. Nutr. 2003, 89, 201–216. [Google Scholar] [CrossRef]
- Fernandez, M.L. Distinct mechanisms of plasma LDL lowering by dietary fibre in the guinea pig: Specific effects of pectin, guar gum and psyllium. J. Lipid Res. 1995, 36, 2394–2404. [Google Scholar] [CrossRef]
- Fernandez, M.L.; Lin, E.C.; Trejo, A.; McNamara, D.J. Prickly pear (Opuntia sp.) pectin reverses low density lipoprotein receptor suppression induced by a hypercholesterolemic diet in guinea pigs. J. Nutr. 1992, 122, 2330–2340. [Google Scholar] [CrossRef]
- Shen, H.; He, L.; Price, R.L.; Fernandez, M.L. Dietary soluble fiber lowers plasma LDL cholesterol concentrations by altering lipoprotein metabolism in female guinea pigs. J. Nutr. 1998, 128, 1434–1441. [Google Scholar] [CrossRef] [Green Version]
- Chen, Q.; Wang, E.; Ma, L.; Zhai, P. Dietary resveratrol increases the expression of hepatic 7α-hydroxylase and ameliorates hypercholesterolemia in high-fat fed C57BL/6J mice. Lipids Health Dis. 2012, 11, 56. [Google Scholar] [CrossRef] [Green Version]
- Jeon, S.-M.; Lee, S.-A.; Choi, M.-S. Antiobesity and vasoprotective effects of resveratrol in ApoE-deficient mice. J. Med. Food. 2014, 17, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Rašković, A.; Ćućuz, V.; Torović, L.; Tomas, A.; Gojković-Bukarica, L.; Ćebović, T.; Milijašević, B.; Stilinović, N.; CvejićHogervorst, J. Resveratrol supplementation improves metabolic control in rats with induced hyperlipidemia and type 2 diabetes. Saudi Pharm. J. 2019, 27, 1036–1043. [Google Scholar] [CrossRef] [PubMed]
- Jo, S.P.; Kim, J.K.; Lim, Y.H. Antihyperlipidemic effects of rhapontin and rhapontigenin from rheum undulatum in rats fed a high-cholesterol diet. Planta Med. 2014, 80, 1067–1071. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, J.; Ma, M.; Lu, Y.; Wang, L.; Wu, C.; Duan, H. Rhaponticin from rhubarb rhizomes alleviates liver steatosis and improves blood glucose and lipid profiles in KK/Ay diabetic mice. Planta Med. 2009, 75, 472–477. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira Andrade, J.M.; Frade, A.C.; Guimarães, J.B.; Freitas, K.M.; Lopes, M.T.; Guimarães, A.L.; de Paula, A.M.; Coimbra, C.C.; Santos, S.H. Resveratrol increases brown adipose tissue thermogenesis markers by increasing SIRT1 and energy expenditure and decreasing fat accumulation in adipose tissue of mice fed a standard diet. Eur. J. Nutr. 2014, 53, 1503–1510. [Google Scholar] [CrossRef]
- Ding, S.; Jiang, J.; Wang, Z.; Zhang, G.; Yin, J.; Wang, X.; Wang, S.; Yu, Z. Resveratrol reduces the inflammatory response in adipose tissue and improves adipose insulin signaling in high-fat diet-fed mice. PeerJ 2018, 6, e5173. [Google Scholar] [CrossRef]
- Timmers, S.; Konings, E.; Bilet, L.; Houtkooper, R.H.; van deWeijer, T.; Goossens, G.H.; Hoeks, J.; van der Krieken, S.; Ryu, D.; Kersten, S.; et al. Calorie restriction-like effects of 30 days of resveratrol supplementation on energy metabolism and metabolic profile in obese humans. Cell Metab. 2011, 14, 612–622. [Google Scholar] [CrossRef] [Green Version]
- Poulsen, M.M.; Vestergaard, P.F.; Clasen, B.F.; Radko, Y.; Christensen, L.P.; Stødkilde-Jørgensen, H.; Møller, N.; Jessen, N.; Pedersen, S.B.; Jørgensen, J.O. High-dose resveratrol supplementation in obese men: An investigator-initiated, randomized, placebo-controlled clinical trial of substrate metabolism, insulin sensitivity, and body composition. Diabetes 2013, 62, 1186–1195. [Google Scholar] [CrossRef] [Green Version]
- Wong, R.H.; Berry, N.M.; Coates, A.M.; Buckley, J.D.; Bryan, J.; Kunz, I.; Howe, P.R. Chronic resveratrol consumption improves brachial flow-mediated dilatation in healthy obese adults. J. Hypertens. 2013, 31, 18191827. [Google Scholar] [CrossRef]
- Van der Made, S.M.; Plat, J.; Mensink, R.P. Resveratrol does not influence metabolic risk markers related to cardiovascular health in overweight and slightly obese subjects: A randomized, placebo-controlled crossover trial. PLoS ONE 2015, 10, e0118393. [Google Scholar] [CrossRef] [Green Version]
- Faghihzadeh, F.; Adibi, P.; Hekmatdoost, A. The effects of resveratrol supplementation on cardiovascular risk factors in patients with non-alcoholic fatty liver disease: A randomised, double-blind, placebo-controlled study. Br. J. Nutr. 2015, 114, 796–803. [Google Scholar] [CrossRef]
- Dash, S.; Xiao, C.; Morgantini, C.; Szeto, L.; Lewis, G.F. High-dose resveratrol treatment for 2 weeks inhibits intestinal and hepatic lipoprotein production in overweight/obese men. Arter. Thromb. Vasc. Biol. 2013, 33, 2895–2901. [Google Scholar] [CrossRef] [Green Version]
- Knop, F.K.; Konings, E.; Timmers, S.; Schrauwen, P.; Holst, J.J.; Blaak, E.E. Thirty days of resveratrol supplementation does not affect postprandial incretin hormone responses, but suppresses postprandial glucagon in obese subjects. Diabet. Med. 2013, 30, 1214–1218. [Google Scholar] [CrossRef]
- Konings, E.; Timmers, S.; Boekschoten, M.V.; Goossens, G.H.; Jocken, J.W.; Afman, L.A.; Müller, M.; Schrauwen, P.; Mariman, E.C.; Blaak, E.E. The effects of 30-days resveratrol supplementation on adipose tissue morphology and gene expression patterns in obese men. Int. J. Obes. 2014, 38, 470–473. [Google Scholar] [CrossRef]
- Witte, A.V.; Kerti, L.; Margulies, D.S.; Floel, A. Effects of resveratrol on memory performance, hippocampal functional connectivity, and glucose metabolism in healthy older adults. J. Neurosci. 2014, 34, 7862–7870. [Google Scholar] [CrossRef] [Green Version]
- Kjær, T.N.; Ornstrup, M.J.; Poulsen, M.M.; Stødkilde-Jørgensen, H.; Jessen, N.; Jørgensen, J.O.L.; Richelsen, B.; Pedersen, S.B. No beneficial effects of resveratrol on the metabolic syndrome: A randomized placebo-controlled clinical trial. J. Clin. Endocrinol. Metab. 2017, 102, 1642–1651. [Google Scholar] [CrossRef] [Green Version]
- Anton, S.D.; Embry, C.; Marsiske, M.; Lu, X.; Doss, H.; Leeuwenburgh, C.; Manini, T.M. Safety and metabolic outcomes of resveratrol supplementation in older adults: Results of a twelve-week, placebo-controlled pilot study. Exp. Gerontol. 2014, 57, 181–187. [Google Scholar] [CrossRef] [Green Version]
- Kantartzis, K.; Fritsche, L.; Bombrich, M.; Machann, J.; Schick, F.; Staiger, H.; Kunz, I.; Schoop, R.; Lehn-Stefan, A.; Heni, M.; et al. Effects of resveratrol supplementation on liver fat content in overweight and insulin-resistant subjects: A randomized, double-blind, placebo-controlled clinical trial. Diabetes Obes. Metab. 2018, 20, 1793–1797. [Google Scholar] [CrossRef]
- Van der Made, S.M.; Plat, J.; Mensink, R.P. Trans-resveratrol supplementation and endothelial function during the fasting and postprandial phase: A randomized placebo-controlled trial in overweight and slightly obese participants. Nutrients 2017, 9, 596. [Google Scholar] [CrossRef]
- Walker, J.M.; Eckardt, P.; Aleman, J.O.; da Rosa, J.C.; Liang, Y.; Iizumi, T.; Etheve, S.; Blaser, M.J.L.; Breslow, J.; Holt, P.R. The effects of trans-resveratrol on insulin resistance, inflammation, and microbiota in men with the metabolic syndrome: A pilot randomized, placebo-controlled clinical trial. J. Clin. Transl. Res. 2018, 4, 122–135. [Google Scholar]
- De Ligt, M.; Bergman, M.; Fuentes, R.M.; Essers, H.; Moonen-Kornips, E.; Havekes, B.; Schrauwen-Hinderling, V.B.; Schrauwen, P. No effect of resveratrol supplementation after 6 months on insulin sensitivity in overweight adults: A randomized trial. Am. J. Clin. Nutr. 2020, 12, 1029–1038. [Google Scholar] [CrossRef]
- Batista-Jorge, G.C.; Barcala-Jorge, A.S.; Silveira, M.F.; Lelis, D.F.; Andrade, J.M.O.; de Paula, A.M.B.; Guimarães, A.L.S.; Santos, S.H.S. Oral resveratrol supplementation improves metabolic syndrome features in obese patients submitted to a lifestyle changing program. Life Sci. 2020, 256, 117962. [Google Scholar] [CrossRef]
- Most, J.; Timmers, S.; Warnke, I.; Jocken, J.W.; van Boekschoten, M.; de Groot, P.; Bendik, L.; Schrauwen, P.; Goossens, G.H.; Blaak, E.E. Combined epigallocatechin-3-gallate and resveratrol supplementation for 12 wk increases mitochondrial capacity and fat oxidation, but not insulin sensitivity, in obese humans: A randomized controlled trial. Am. J. Clin. Nutr. 2016, 104, 215–227. [Google Scholar] [CrossRef] [Green Version]
- Most, J.; Warnke, I.; van Boekschoten, M.; Jocken, J.W.; de Groot, P.; Friedel, A.; Bendik, I.; Goossens, G.H.; Blaak, E.E. The effects of polyphenol supplementation on adipose tissue morphology and gene expression in overweight and obese humans. Adipocyte 2018, 7, 190–196. [Google Scholar] [CrossRef]
- Arzola-Paniagua, M.A.; García-Salgado López, E.R.; Calvo-Vargas, C.G.; Guevara-Cruz, M. Efficacy of an orlistat-resveratrol combination for weight loss in subjects with obesity: A randomized controlled trial. Obesity 2016, 24, 1454–1463. [Google Scholar] [CrossRef] [Green Version]
- Su, Z.-L.; Hang, P.-Z.; Hu, J.; Zheng, Y.-Y.; Sun, H.-Q.; Guo, J.; Liu, K.-Y.; Du, Z.-M. Aloe-emodin exerts cholesterol-lowering effects by inhibiting proprotein convertase subtilisin/kexin type 9 in hyperlipidemic rats. Acta Pharmacol. Sin. 2020, 41, 1085–1092. [Google Scholar] [CrossRef]
- Lee, S.J.; Cho, S.J.; Kwon, E.Y.; Choi, M.S. Physcion reduces lipid accumulation and prevents the obesity in mice. Nutr. Metab. 2019, 16, 31. [Google Scholar] [CrossRef]
- Wang, J.; Ji, J.; Song, Z.; Zhang, W.; He, X.; Li, F.; Zhang, C.; Guo, C.; Wang, C.; Yuan, C. Hypocholesterolemic effect of emodin by simultaneous determination of in vitro and in vivo bile salts binding. Fitoterapia 2016, 110, 116–122. [Google Scholar] [CrossRef]
- Cheng, L.; Zhang, S.; Shang, F.; Ning, Y.; Huang, Z.; He, R.; Sun, J.; Dong, S. Emodin improves glucose and lipid metabolism disorders in obese mice via activating brown adipose tissue and inducing browning of white adipose tissue. Front. Endocrinol. 2021, 12, 618037. [Google Scholar] [CrossRef]
- Czernichow, S.; Batty, G.D. Withdrawal of sibutramine for weight loss: Where does this leave clinicians? Obes. Facts 2010, 3, 155–156. [Google Scholar] [CrossRef]
- Mohamed, G.A.; Ibrahim, S.R.M.; Elkhayat, E.S.; El Dine, R.S. Natural anti-obesity agents. Bull. Fac. Pharm. Cairo Univ. 2014, 52, 269–284. [Google Scholar] [CrossRef] [Green Version]
- Qi, X. Review of the clinical effect of orlistat. In Proceedings of the IOP Conference Series: Materials Science and Engineering, Xiamen, China, 15–17 December 2018. [Google Scholar] [CrossRef]
- Rajan, L.; Palaniswamy, D.; Mohankumar, S.K. Targeting obesity with plant-derived pancreatic lipase inhibitors: A comprehensive review. Pharmacol. Res. 2020, 155, 104681. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Gonzalez, A.I.; Alvarez-Parrilla, E.; Díaz-Sánchez, A.G.; de la Rosa, L.; Núñez-Gastélum, J.A.; Vazquez-Flores, A.A.; Gonzalez-Aguilar, G.A. In vitro inhibition of pancreatic lipase by polyphenols: A kinetic, fluorescence spectroscopy and molecular docking study. Food Technol. Biotechnol. 2017, 55, 519–530. [Google Scholar] [CrossRef] [PubMed]
- Kasabri, V.; Al-Hallaq, E.K.; Bustanji, Y.K.; Abdul-Razzak, K.K.; Abaza, I.F.; Afifi, F.U. Antiobesity and antihyperglycaemic effects of Adiantumcapillus-veneris extracts: In-vitro and in-vivo evaluations. Pharm. Biol. 2017, 55, 164–172. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, Y.; Zhang, D.; Yang, G.; Zheng, Y.; Guo, L. Screening of antilipase components of Artemisia argyi leaves based on spectrum-effect relationships and HPLC-MS/MS. Front. Pharmacol. 2021, 12, 675396. [Google Scholar] [CrossRef]
- Vijayaraj, P.; Nakagawa, H.; Yamaki, K. Cyanidin and cyanidin-3-glucoside derived from Vignaunguiculata act as noncompetitive inhibitors of pancreatic lipase. J. Food Biochem. 2019, 43, e12774. [Google Scholar] [CrossRef]
- Thilavech, T.; Adisakwattana, S. Cyanidin-3-rutinoside acts as a natural inhibitor of intestinal lipid digestion and absorption. BMC Complement. Altern. Med. 2019, 19, 242. [Google Scholar] [CrossRef]
- Nakai, M.; Fukui, Y.; Asami, S.; Toyoda-Ono, T.; Iwashita, T.; Shibata, H.; Mitsunaga, T.; Hashimoto, F.; Kiso, Y. Inhibitory effects of oolong tea polyphenols on pancreatic lipase in vitro. J. Agric. Food Chem. 2005, 53, 4593–4598. [Google Scholar] [CrossRef]
- Kim, Y.-M.; Lee, E.-W.; Eom, S.-H.; Kim, T.H. Pancreatic lipase inhibitory stilbenoids from the roots of Vitisvinifera. Int. J. Food Sci. Nutr. 2014, 65, 97–100. [Google Scholar] [CrossRef]
- Habtemariam, S. Antihyperlipidemic components of Cassia auriculata aerial parts: Identification through in vitro studies. Phytother. Res. 2013, 27, 152–155. [Google Scholar] [CrossRef]
- Batubara, L.; Kuspradini, H.; Muddathir, A.M.; Mitsunaga, T. Intsiapalembanica wood extracts and its isolated compounds as Propionibacterium acnes lipase inhibitor. J. Wood Sci. 2014, 60, 169–174. [Google Scholar] [CrossRef]
- Tao, Y.; Zhang, Y.; Wang, Y.; Cheng, Y. Hollow fiber based affinity selection combined with high performance liquid chromatography-mass spectroscopy for rapid screening lipase inhibitors from lotus leaf. Anal. Chim. Acta 2013, 785, 75–81. [Google Scholar] [CrossRef]
- Guo, X.; Liu, J.; Cai, S.; Wang, O.; Ji, B. Synergistic interactions of apigenin, naringin, quercetin and emodin on inhibition of 3T3-L1 preadipocyte differentiation and pancreas lipase activity. Obes. Res. Clin. Pract. 2016, 10, 327–339. [Google Scholar] [CrossRef]
- Zheng, C.-D.; Duan, Y.-Q.; Gao, J.-M.; Ruan, Z.-G. Screening for anti-lipase properties of 37 traditional Chinese medicinal herbs. J. Chin. Med. Assoc. 2020, 73, 319–324. [Google Scholar] [CrossRef] [Green Version]
- Gholamhoseinian, A.; Shahouzebi, B.; Sharifi-Far, F. Inhibitory effect of some plant extracts on pancreatic lipase. Int. J. Pharmacol. 2010, 6, 18–24. [Google Scholar] [CrossRef] [Green Version]
- Abe, I.; Seki, T.; Noguchi, H.; Kashiwada, Y. Galloyl esters from rhubarb are potent inhibitors of squalene epoxidase, a key enzyme in cholesterol biosynthesis. Planta Med. 2000, 66, 753–756. [Google Scholar] [CrossRef]
- Shi, D.H.; Xu, C.; Guo, B.X.; Wang, X.T.; Chen, Y.X.; Tan, R.X. Inhibition of soluble epoxide hydrolase by extracts derived from inflammation-treating Chinese medicinal herbs. Phytother. Res. 2008, 22, 1264–1268. [Google Scholar] [CrossRef]
- Li, W.; Kim, J.H.; Zhou, W.; Shim, S.H.; Ma, J.Y.; Kim, Y.H. Soluble epoxide hydrolase inhibitory activity of phenolic components from the rhizomes and roots of Gentiana scabra. Biosci. Biotechnol. Biochem. 2015, 79, 907–911. [Google Scholar] [CrossRef]
- Wang, C.Y.; Lee, S.; Jang, H.J.; Su, X.D.; Wang, H.S.; Kim, Y.H.; Yang, S.Y. Inhibition potential of phenolic constituents from the aerial parts of Tetrastigmahemsleyanum against soluble epoxide hydrolase and nitric oxide synthase. J. Enzyme Inhib. Med. Chem. 2019, 34, 753–760. [Google Scholar] [CrossRef]
- Han, Y.K.; Lee, J.S.; Yang, S.Y.; Lee, K.Y.; Kim, Y.H. In Vitro and in silico studies of soluble epoxide hydrolase inhibitors from the roots of lycopuslucidus. Plants 2021, 10, 356. [Google Scholar] [CrossRef]
- Jo, A.R.; Kim, J.H.; Yan, X.T.; Yang, S.Y.; Kim, Y.H. Soluble epoxide hydrolase inhibitory components from Rheum undulatum and in silico approach. J. Enzyme Inhib. Med. Chem. 2016, 31, 70–78. [Google Scholar] [CrossRef] [Green Version]
- Kitamura, S.; Morisseau, C.; Inceoglu, B.; Kamita, S.G.; De Nicola, G.R.; Nyegue, M.; Hammock, B.D. Potent natural soluble epoxide hydrolase inhibitors from Pentadiplandrabrazzeanabaillon: Synthesis, quantification, and measurement of biological activities in vitro and in vivo. PLoS ONE 2015, 10, e0117438. [Google Scholar] [CrossRef]
- Gómez-Hernández, A.; Beneit, N.; Díaz-Castroverde, S.; Escribano, Ó. Differential role of adipose tissues in obesity and related metabolic and vascular complications. Int. J. Endocrinol. 2016, 2016, 1216783. [Google Scholar] [CrossRef] [Green Version]
- Jakab, J.; Miškić, B.; Mikšić, S.; Juranić, B.; Ćosić, V.; Schwarz, D.; Včev, A. Adipogenesis as a potential anti-obesity target: A review of pharmacological treatment and natural products. Diabetes Metab. Syndr. Obes. 2021, 14, 67–83. [Google Scholar] [CrossRef]
- Chang, E.; Kim, C.Y. Natural products and obesity: A focus on the regulation of mitotic clonal expansion during adipogenesis. Molecules 2019, 24, 1157. [Google Scholar] [CrossRef] [Green Version]
- Bhardwaj, M.; Yadav, P.; Vashishth, D.; Sharma, K.; Kumar, A.; Chahal, J.; Dalal, S.; Kataria, S.K. A review on obesity management through natural compounds and a green nanomedicine-based approach. Molecules 2021, 26, 3278. [Google Scholar] [CrossRef]
- Miao, J.; He, X.; Hu, J.; Cai, W. Emodin inhibits NF-κB signaling pathway to protect obese asthmatic rats from pathological damage via Visfatin. Tissue Cell 2022, 74, 101713. [Google Scholar] [CrossRef]
- Yahfoufi, N.; Alsadi, N.; Jambi, M.; Matar, C. The immunomodulatory and anti-inflammatory role of polyphenols. Nutrients 2018, 10, 1618. [Google Scholar] [CrossRef] [Green Version]
- Mongioì, L.M.; La Vignera, S.; Cannarella, R.; Cimino, L.; Compagnone, M.; Condorelli, R.A.; Calogero, A.E. The role of resveratrol administration in human obesity. Int. J. Mol. Sci. 2021, 22, 4362. [Google Scholar] [CrossRef]
- Kwon, J.Y.; Seo, S.G.; Heo, Y.S.; Yue, S.; Cheng, J.-X.; Lee, K.W.; Kim, K.-H. Piceatannol, natural polyphenolic stilbene, inhibits adipogenesis via modulation of mitotic clonal expansion and insulin receptor-dependent insulin signaling in early phase of differentiation. J. Biol. Chem. 2012, 287, 11566–11578. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Li, Y.; Hanafusa, Y.; Yeh, Y.-S.; Maruki-Uchida, H.; Kawakami, S.; Sai, M.; Goto, T.; Ito, T.; Kawada, T. Piceatannol exhibits anti-inflammatory effects on macrophages interacting with adipocytes. Food Sci. Nutr. 2017, 5, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Park, I.S.; Han, Y.; Jo, H.; Lee, K.W.; Song, Y.S. Piceatannol is superior to resveratrol at suppressing adipogenesis in human visceral adipose-derived stem cells. Plants 2021, 10, 366. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Zhang, X.L.; Tao, R.Y.; Niu, Y.J.; Chen, X.G.; Tian, J.Y.; Ye, F. Rhein, an inhibitor of adipocyte differentiation and adipogenesis. J. Asian Nat. Prod. Res. 2011, 13, 714–723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Fan, S.; Hu, N.; Gu, M.; Chu, C.; Li, Y.; Lu, X.; Huang, C. Rhein reduces fat weight in db/db mouse and prevents diet-induced obesity in C57Bl/6 mouse through the inhibition of PPARγ signaling. PPAR Res. 2012, 2012, 374936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheng, X.; Zhu, X.; Zhang, Y.; Cui, G.; Peng, L.; Lu, X.; Zang, Y.Q. Rhein protects against obesity and related metabolic disorders through liver X receptor-mediated uncoupling protein 1 upregulation in brown adipose tissue. Int. J. Biol. Sci. 2012, 8, 1375–1384. [Google Scholar] [CrossRef]
- Fang, J.Y.; Huang, T.H.; Chen, W.J.; Aljuffali, I.A.; Hsu, C.Y. Rhubarb hydroxyanthraquinones act as antiobesity agents to inhibit adipogenesis and enhance lipolysis. Biomed. Pharmacother. 2022, 146, 112497. [Google Scholar] [CrossRef]
- Ma, C.; Wang, Z.; Xia, R.; Wei, L.; Zhang, C.; Zhang, J.; Zhao, L.; Wu, H.; Kang, L.; Yang, S. Danthron ameliorates obesity and MAFLD through activating the interplay between PPARα/RXRα heterodimer and adiponectin receptor 2. Biomed. Pharmacother. 2021, 137, 111344. [Google Scholar] [CrossRef]
- Zhang, J.; Kang, H.; Wang, L.; Zhao, X.; He, L. Chrysophanol ameliorates high-fat diet-induced obesity and inflammation in neonatal rats. Pharmazie 2018, 73, 228–233. [Google Scholar]
- Wang, Y.-J.; Huang, S.-L.; Feng, Y.; Ning, M.-M.; Leng, Y. Emodin, an 11β-hydroxysteroid dehydrogenase type 1 inhibitor, regulates adipocyte function in vitro and exerts anti-diabetic effect in ob/ob mice. Acta Pharmacol. Sin. 2012, 33, 1195–1203. [Google Scholar] [CrossRef] [Green Version]
- Reimer, R.A.; McBurney, M.I. Dietary fiber modulates intestinal pro-glucagon messenger ribonucleic acid and postprandial secretion of glucagon-like peptide-1 and insulin in rats. Endocrinology 1996, 137, 3948–3956. [Google Scholar] [CrossRef] [Green Version]
- Reimer, R.A.; Thomson, A.B.; Rajotte, R.V.; Basu, T.K.; Ooraikul, B.; McBurney, M.I. A physiological level of rhubarb fiber increases proglucagon gene expression and modulates intestinal glucose uptake in rats. J. Nutr. 1997, 127, 1923–1928. [Google Scholar] [CrossRef]
- Shojaei Shad, F.; Haghighi, M.J. Study of the effect of the essential oil (extract) of rhubarb stem (shoot) on glycosylated hemoglobin and fasting blood glucose levels in patients with type II diabetes. Biomedicine 2018, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Jiang, C.S.; Liang, L.F.; Guo, Y.W. Natural products possessing protein tyrosine phosphatase 1B (PTP1B) inhibitory activity found in the last decades. Acta Pharmacol. Sin. 2012, 33, 1217–1245. [Google Scholar] [CrossRef]
- Saidu, Y.; Muhammad, S.A.; Abbas, A.Y.; Onu, A.; Tsado, I.M.; Muhammad, L. In vitro screening for protein tyrosine phosphatase 1B and dipeptidyl peptidase IV inhibitors from selected Nigerian medicinal plants. J. Intercult. Ethnopharmacol. 2016, 6, 154–157. [Google Scholar]
- Lee, W.; Kim, S.N.; Yoon, G. Screening of medicinal herbs for inhibitory activity against protein tyrosine phosphatase B. Korean J. Pharmacogn. 2010, 41, 227–231. [Google Scholar]
- Yue, H.; Jiang, S.; Wang, L.; Banma, C.; Zhou, G.; Shao, Y.; Tao, Y.; Zhao, X. Hypoglycemic ingredients identification of Rheum tanguticum Maxim. ex Balf. by UHPLC-triple-TOF-MS/MS and interrelationships between ingredients content and glycosidase inhibitory activities. Ind. Crops Prod. 2022, 178, 114595. [Google Scholar] [CrossRef]
- Choi, S.B.; Ko, B.S.; Park, S.K.; Jang, J.S.; Park, S. Insulin sensitizing and a-glucoamylase inhibitory action of sennosides, rheins and rhaponticin in RheiRhizoma. Life Sci. 2006, 78, 934–942. [Google Scholar] [CrossRef]
- Choi, S.Z.; Lee, S.O.; Jang, K.U.; Chung, S.H.; Park, S.H.; Kang, H.C.; Yang, E.Y.; Cho, H.J.; Lee, K.R. Antidiabetic stilbene and anthraquinone derivatives from Rheum undulatum. Arch. Pharm. Res. 2005, 28, 1027–1030. [Google Scholar] [CrossRef]
- Hosseini, A.; Mollazadeh, H.; Amiri, M.S.; Sadeghnia, H.R.; Ghorbani, A. Effects of a standardized extract of Rheum turkestanicumJanischew root on diabetic changes in the kidney, liver and heart of streptozotocin-induced diabetic rats. Biomed. Pharmacother. 2017, 86, 605–611. [Google Scholar] [CrossRef]
- Bahnasy, R.M. Rhubarb (Rheum emodi) improves some immunity markers and blood glucose in rats exposed to glucocorticoids. Home Econ. J. 2020, 36, 111–128. [Google Scholar] [CrossRef]
- Cheng, F.R.; Cui, H.X.; Fang, J.L.; Yuan, K.; Guo, Y. Ameliorative effect and mechanism of the purified anthraquinone-glycoside preparation from Rheum Palmatum L. on type 2 diabetes mellitus. Molecules 2019, 24, 1454. [Google Scholar] [CrossRef] [Green Version]
- Hadjzadeh, M.A.; Rajaei, Z.; Khodaei, E.; Malek, M.; Ghanbari, H. Rheum turkestanicum rhizomes possess anti-hypertriglyceridemic, but not hypoglycemic or hepatoprotective effect in experimental diabetes. Avicenna J. Phytomed. 2017, 7, 1–9. [Google Scholar]
- Cui, H.-X.; Zhang, L.-S.; Luo, Y.; Yuan, K.; Huang, Z.-Y.; Guo, Y. A purified anthraquinone-glycoside preparation from rhubarb ameliorates type 2 diabetes mellitus by modulating the gut microbiota and reducing inflammation. Front. Microbiol. 2019, 10, 1423. [Google Scholar] [CrossRef]
- Lee, W.; Yoon, G.; Hwang, Y.R.; Kim, Y.K.; Kim, S.N. Anti-obesity and hypolipidemic effects of Rheum undulatum in high-fat diet-fed C57BL/6 mice through protein tyrosine phosphatase 1B inhibition. BMB Rep. 2012, 45, 141–146. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Li, X.; Zeng, X.; Ou, Z.; Xue, M.; Gao, D.; Liu, S.; Li, S.; Yang, S. Rheum palmatum L. attenuates high fat diet-induced hepatosteatosis by activating AMP-activated protein kinase. Am. J. Chin. Med. 2016, 44, 551–564. [Google Scholar] [CrossRef]
- Moon, M.K.; Kang, D.G.; Lee, A.S.; Yeom, K.B.; Kim, J.S.; Lee, H.S. Anti-atherogenic effects of the aqueous extract of rhubarb in rats fed an atherogenic diet. Am. J. Chin. Med. 2008, 36, 555–568. [Google Scholar] [CrossRef]
- Liu, Y.; Yan, F.; Liu, Y.; Zhang, C.; Yu, H.; Zhang, Y.; Zhao, Y. Aqueous extract of rhubarb stabilizes vulnerable atherosclerotic plaques due to depression of inflammation and lipid accumulation. Phytother. Res. 2008, 22, 935–942. [Google Scholar] [CrossRef]
- Shokri, H.; Farokhi, F.; Heydari, R.; Manaffar, R. Renoprotective effect of hydroalcoholic extract of Rheum ribes root in diabetic female rats. Avicenna J. Phytomed. 2014, 4, 392–401. [Google Scholar]
- Hadjzadeh, M.A.R.; Parsaee, H.; Sadeghian, A. Cholesterol lowering effects of Rheum ribs in hypercholesterolemic rabbits. Med. J. Islamic Repub. Iran 2004, 18, 277–280. [Google Scholar]
- Régnier, M.; Rastelli, M.; Morissette, A.; Suriano, F.; Le Roy, T.; Pilon, G.; Delzenne, N.M.; Marette, A.; van Hul, M.; Cani, P.D. Rhubarb supplementation prevents diet-induced obesity and diabetes in association with increased Akkermansiamuciniphila in mice. Nutrients 2020, 12, 2932. [Google Scholar] [CrossRef]
- Zhou, Q.; Zhang, Y.; Wang, X.; Yang, R.; Zhu, X.; Zhang, Y.; Chen, C.; Yuan, H.; Yang, Z.; Sun, L. Gut bacteria Akkermansia is associated with reduced risk of obesity: Evidence from the American Gut Project. Nutr. Metab. 2020, 17, 90. [Google Scholar] [CrossRef]
- Liu, Y.-F. Treatment with rhubarb improves brachial artery endothelial function in patients with atherosclerosis: A randomized, double-blind, placebo-controlled clinical trial. Am. J. Chin. Med. 2007, 35, 583–595. [Google Scholar] [CrossRef] [PubMed]
- Shojaei-Shad, F.; Jahantigh-Haghighi, M.; Mansouri, A.; Jahantigh-Haghighi, M. The effect of Rhubarb stem extract on blood pressure and weight of type 2 diabetic patients. Med. Sci. 2019, 23, 159–162. [Google Scholar]
- Roerig, J.L.; Steffen, K.J.; Mitchell, J.E.; Zunker, C. Laxative abuse. Epidemiology, diagnosis and management. Drugs 2010, 70, 1487–1503. [Google Scholar] [CrossRef] [PubMed]
- Cui, T.; Zhou, Q.-X.; Zhang, D.; Jiang, X.-H. Investigation the distributions and pharmacokinetics of five rhubarb anthraquinones in rabbits and rats. J. Anal. Pharm. Res. 2016, 3, 00048. [Google Scholar] [CrossRef]
- Cai, Y.; Sun, M.; Xing, J.; Corke, H. Antioxidant phenolic constituents in roots of Rheum officinale and Rubiacordifolia: Structure-radical scavenging activity relationships. J. Agric. Food Chem. 2004, 52, 7884–7890. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.S.; Chen, F.; Liu, X.H.; Xu, H.; Zhou, Y.Z. Progress in research of chemical constituents and pharmacological actions of Rhubarb. Chin. J. New Drugs 2011, 20, 1534–1548. [Google Scholar]
- Rehman, H.; Begum, W.; Anjum, F.; Tabasum, H. Rheum emodi (Rhubarb): A fascinating herb. J. Pharmacogn. Phytochem. 2014, 3, 89–94. [Google Scholar]
- Li, F.; Wang, S.-C.; Wang, X.; Ren, Q.-Y.; Wang, W.; Shang, G.-W.; Zhang, L.; Zhang, S.-H. Novel exploration of cathartic pharmacology induced by rhubarb. ZhongguoZhong Yao ZaZhi 2008, 33, 481–484. [Google Scholar]
- Wei, H.; Chang, J.; Liu, P.; Li, Z.; Miao, G.; Liu, X.; Liu, C.; Zhang, X. Design and evaluation of rhubarb total free anthraquinones oral colon-specific drug delivery granules to improve the purgative effect. Braz. J. Pharm. Sci. 2019, 55, e17110. [Google Scholar] [CrossRef] [Green Version]
- Liu, P.; Wei, H.; Chang, J.; Miao, G.; Liu, X.; Li, Z.; Liu, L.; Zhang, X.; Liu, C. Oral colon-specific drug delivery system reduces the nephrotoxicity of rhubarb anthraquinones when they produce purgative efficacy. Exp. Ther. Med. 2017, 14, 3589–3601. [Google Scholar] [CrossRef] [Green Version]
- Xie, W.; Xing, D.; Zhao, Y.; Su, H.; Meng, Z.; Chen, Y.; Du, L. A new tactic to treat postprandial hyperlipidemia in diabetic rats with gastroparesis by improving gastrointestinal transit. Eur. J. Pharmacol. 2005, 510, 113–120. [Google Scholar] [CrossRef]
- Tseng, S.-H.; Chien, T.-Y.; Chen, J.-R.; Lin, I.-H.; Wang, C.-C. Hypolipidemic effects of three purgative decoctions. Evid.-Based Complement. Altern. Med. 2011, 2011, 249254. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Kim, S.H.; Cha, J. Antiobesity effects of the combined plant extracts varying the combination ratio of phyllostachyspubescens leaf extract and scutellariabaicalensis root extract. Evid.-Based Complement. Altern. Med. 2016, 2016, 9735276. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Main mechanisms of anti-obesity action recognised for different natural, plant-derived substances.
Figure 1.
Main mechanisms of anti-obesity action recognised for different natural, plant-derived substances.

Figure 2.
Graphical summary of main groups of the specialized metabolites synthesized by rhubarb species.
Figure 2.
Graphical summary of main groups of the specialized metabolites synthesized by rhubarb species.

Figure 3.
Modulatory effects of the plant-derived substances on different steps of adipogenesis. The wingless-type MMTV integration site (Wnt) signalling, the growth factor-beta (TGF-β)/bone morphogenic protein (BMP) signalling, Hedgehogs (Hh), Notch, and the fibroblast growth factor (FGF)-dependent response are the main mechanisms regulating the commitment of multipotent mesenchymal stem cells (MSCs). Early stages of adipocyte differentiation are primarily regulated by C/EBPβ and C/EBPδ, responsible for the induction of C/EBPα and PPARγ—the central positive modulators of adipogenesis. The rhubarb-originated substances may interfere with different pathways controlling the adipocyte differentiation and maturation, including the blockade of clonal expansion as well as downregulation of PPARγ and C/EBPα. Abbreviations and explanations: AMPK—AMP-activated protein kinase; KLF 2—Krüppel-like factor; Pref-1—preadipocyte factor 1; DEC1,2—transcription factors DEC1,2; GATA 2/3—GATA-binding factor 2/3; SIRT1/2—sirtuin 1,2; TGF-β—tumour growth factor beta; PI3K/Akt—phosphatidylinositol 3-kinase and Akt/Protein Kinase B pathway; KLF4/5/9/15—Krüppel-like factors 4,5,9, and 15; CREB—cAMP response element-binding protein; SIRT7—sirtuin 7; SREBP1—sterol regulatory element-binding protein 1; ZFP423—C2H2 zinc-finger protein; FXR—farnesoid X receptor; C/EBPα, C/EBPβ and C/EBPδ—CCAAT/enhancer-binding protein alpha, beta and delta; PPARγ—peroxisome proliferator-activated receptor gamma.
Figure 3.
Modulatory effects of the plant-derived substances on different steps of adipogenesis. The wingless-type MMTV integration site (Wnt) signalling, the growth factor-beta (TGF-β)/bone morphogenic protein (BMP) signalling, Hedgehogs (Hh), Notch, and the fibroblast growth factor (FGF)-dependent response are the main mechanisms regulating the commitment of multipotent mesenchymal stem cells (MSCs). Early stages of adipocyte differentiation are primarily regulated by C/EBPβ and C/EBPδ, responsible for the induction of C/EBPα and PPARγ—the central positive modulators of adipogenesis. The rhubarb-originated substances may interfere with different pathways controlling the adipocyte differentiation and maturation, including the blockade of clonal expansion as well as downregulation of PPARγ and C/EBPα. Abbreviations and explanations: AMPK—AMP-activated protein kinase; KLF 2—Krüppel-like factor; Pref-1—preadipocyte factor 1; DEC1,2—transcription factors DEC1,2; GATA 2/3—GATA-binding factor 2/3; SIRT1/2—sirtuin 1,2; TGF-β—tumour growth factor beta; PI3K/Akt—phosphatidylinositol 3-kinase and Akt/Protein Kinase B pathway; KLF4/5/9/15—Krüppel-like factors 4,5,9, and 15; CREB—cAMP response element-binding protein; SIRT7—sirtuin 7; SREBP1—sterol regulatory element-binding protein 1; ZFP423—C2H2 zinc-finger protein; FXR—farnesoid X receptor; C/EBPα, C/EBPβ and C/EBPδ—CCAAT/enhancer-binding protein alpha, beta and delta; PPARγ—peroxisome proliferator-activated receptor gamma.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Hypolipidemic action of the rhubarb-derived fibre.
| The Examined Substances | Type of Study | Experimental Model, Doses and Concentrations | Main Effects of the Rhubarb Fibre and Stilbenes Administration | References |
|---|---|---|---|---|
| ANIMAL STUDIES | ||||
| R. rhaponticum stalk-derived preparation containing 74% of dietary fibre/dry mass (incl. 66% insoluble and 8% soluble) | animal | mice fed with cholesterol-enriched diet with 5% of rhubarb stalk fibre, for 4 weeks | ↓ the acyl CoA: cholesterol acyltransferase (ACAT) activity; no effects on the cholesterol-rich diet enhancement of the β-hydroxyβ-methyl coenzyme A reductase (HMGR) activity | [22] |
| R. rhaponticum stalk fibre | animal | cholesterol-fed C57BL/6J mice receiving the fibre-rich diet (50 g/kg b.w.) for 4 weeks | ↓ plasma cholesterol (−13%); ↓ the hepatic concentrations of total cholesterol (by 34%) and cholesteryl esters (by 34%); ↓ acyl CoA: cholesterol acyltransferase activity; ↓ the faecal bile acid loss; ↓the gallbladder bile acid pool | [19] |
| R. rhaponticum stalk fibre | animal | the diabetes-prone and the streptozotocin-induced diabetic rats receiving the fibre-rich diet (50 g/kg b.w.) for 2 weeks | No effect on the plasma cholesterol and triacylglycerol levels in diabetic rats | [26] |
| HUMAN STUDIES | ||||
| the rhubarb-stalk-derived preparation containing 74% dietary fibre/dry mass (incl. 66% insoluble and 8% soluble) | human | Ten hypercholesterolemic men (BMI of 27.9 ± 3.8 kg/m2); 27 g of rhubarb fibre/day, for 4 weeks | ↓ serum total cholesterol (−8%) and LDL cholesterol (−9%); no changes in HDL cholesterol level; a return of the cholesterol-lowering effect to baseline after the fibre supplementation withdrawal for one month | [23] |
Table 2.
Exemplary data originated from animal studies on the hypolipidemic and anti-obesity properties of stilbenes that are present in rhubarb.
Table 2.
Exemplary data originated from animal studies on the hypolipidemic and anti-obesity properties of stilbenes that are present in rhubarb.
| The Examined Substances | Experimental Model, Doses and Concentrations | Main Effects of the Stilbene Administration | References |
|---|---|---|---|
| Rhaponticin and rhapontigenin isolated from R. rhabarbarum roots | rats fed a high-cholesterol diet, followed by oral rhapontin or rhapontigenin treatment (1, 2.5 and 5 mg/kg b.w. (body weight)/day) | ↓ the serum lipid level; ↑ HDL cholesterol; improvement in the degenerating fatty liver structure; the aspartate aminotransferase (AST) and the alanine aminotransferase (ALT) levels comparable to the control group | [33] |
| Rhaponticin from R. rhabarbarum roots | KK/Ay type 2 diabetic mice treated with rhaponticin (125 mg/kg b.w., 4 weeks) | ↓ the plasma triglyceride, LDL, cholesterol, non-esterified free fatty acids; ↓ lactate dehydrogenase, creatine kinase, AST and ALT activities | [34] |
| Resveratrol | high-fat diet (HFD)-fed C57BL/6 J mice, a daily dose of 200 mg/kg b.w. of resveratrol, for 8 weeks | anti-hypercholesterolemic effects: improvement in serum lipid parameters, ↓ hepatic cholesterol, ↓ body weight, ↑ bile acid pool size, ↑ liver CYP7A1 mRNA expression and CYP7A1 enzyme activity | [30] |
| apoE-deficient mice fed an atherogenic diet containing 0.02% resveratrol (w/w), for 12 weeks | ↓ the plasma total cholesterol, LDL cholesterol, non-high-density-lipoprotein cholesterol, apoB/apoA1 ratio, hepatic cholesterol and triglyceride; ↑ the plasma HDL cholesterol | [31] | |
| mice fed standard diet plus resveratrol (4 g/kg of food to provide a 400-mpk dose), for 8 weeks | ↑ brown adipose tissue thermogenesis; ↑ mRNA of thermogenesis-related genes, incl. uncoupling protein 1 (UCP1), sirtuin 1 (SIRT1), phosphatase and tensin homolog (PTEN) and bone morphogenetic protein 7 (BMP-7) expression; ↓ fat accumulation in adipose tissue; ↓ total cholesterol and glucose levels in plasma | [35] | |
| C57BL/6 mice fed a high-fat diet with a low dose of resveratrol, i.e., 200 mg/kg b.w./day (HFD-RES/L) or with a high dose of resveratrol, i.e., 400 mg/kg b.w./day (HFD-RES/H) | ↓ insulin resistance; ↑ expressions of pAkt, glucose transporter type 4 (GLUT4) and insulin receptor substrate 1 (IRS-1) in white adipose tissue (WAT); ↓ proinflammatory cytokine levels in serum; ↓ macrophage infiltration and C-C chemokine receptor type 2 (CCR2) chemokine expression in white adipose tissue (WAT) | [36] | |
| rats with hyperlipidemia; a daily dose of 20 mg/kg b.w., of resveratrol, for 30 days | ↓ LDL and triglyceride levels; ↑ HDL levels in animals; | [32] |
Table 3.
Evaluation of hypolipidemic effects and anti-obesity action of resveratrol in clinical trials.
Table 3.
Evaluation of hypolipidemic effects and anti-obesity action of resveratrol in clinical trials.
| Number of Participants (n), Resveratrol Doses and Study Duration | Participant Diagnosis | Main Effects of Resveratrol Supplementation in the Context of an Anti-Obesity Action | References |
|---|---|---|---|
| n = 11; 150 mg/day, for 30 days; a randomized, placebo-controlled, double-blind and cross-over study | obesity | Calorie-restriction-like effects; reduction in the sleeping and resting metabolic rate; ↓ intrahepatic lipid content, circulating glucose and triglycerides; ↓ inflammation markers; ↓ the systolic blood pressure; improvement in the HOMA index | [37] |
| n = 24; 500 mg, 3 times/day, for 4 weeks; a randomized, placebo-controlled study | obesity (BMI > 30 kg/m2) | No effect on the total cholesterol, HDL, LDL, plasma triglyceride and blood pressure; no changes in the resting metabolic rate and lipid oxidation | [38] |
| n = 28; 75 mg/day, for 6 weeks; a randomized, placebo-controlled, double-blind, cross-over study | obesity (BMI of 33.3 ± 0.6 kg/m2) | No effects on blood pressure; the flow-mediated dilatation (FMD) increased by 23% | [39] |
| n = 45; 150 mg/day, for 4 weeks; a randomized, placebo-controlled, cross-over study | overweight or obesity (BMI of 25–35 kg/m2) | No effects on metabolic risk markers related to cardiovascular health | [40] |
| n = 50; 500 mg/day, for 12 weeks; a randomized, placebo-controlled, double-blind study | overweight (BMI of 28.35 ± 3.49 and 28.75 ± 3.50 kg/m2, for the intervention and placebo group, respectively); non-alcoholic fatty liver disease | Reduction in BMI, waist circumference, HDL cholesterol and apo A1 both in intervention and placebo group; no differences in the above parameters between these groups; ↓ alanine transferase (ALT) and hepatic steatosis, compared to placebo | [41] |
| n = 8; 1000 mg once a day for a week, then 2000 mg/day for the next week; a randomized, placebo-controlled study | overweight or obesity (BMI of 27.0–40.0 kg/m2), mild hypertriglyceridemia | No effects on insulin sensitivity and blood plasma triglyceride level; ↓ apoB-48 and apoB-100 production rate | [42] |
| n = 10; 150 mg/a day, for 30 days; a randomized, placebo-controlled, double-blind, cross-over study | obesity (BMI of 32 ± 1 kg/m2) | Suppression of postprandial glucagon responses; no changes in fasting glucagon levels | [43] |
| n = 32; 300 mg/day or 1000 mg/day for 90 days; a randomized, placebo controlled, double-blind study | overweight or obesity (BMI of 25.0–34.9 kg/m2) | ↓ glucose levels compared to placebo; no changes in blood pressure, body weight and waist circumference | [47] |
| n = 11; 150 mg/day for 30 days; a randomized, placebo-controlled, double-blind, cross-over study | obesity (BMI of 28–36 kg/m2) | ↓ adipocyte size; changes in the adipose tissue morphology: reduction in the proportion of large and very large adipocytes; increase in small adipocytes; enhanced adipogenesis | [44] |
| n = 23; 200 mg/day for 26 weeks, a placebo-controlled study | overweight (BMI of 25–30 kg/m2), healthy older adults (50–80 years) | ↓ body fat and leptin increase compared to placebo; no significant changes in body weight, BMI or blood pressure compared to placebo | [45] |
| n = 161; 100 mg of resveratrol or 120 mg orlistat + 100 mg resveratrol (O-R group), 3 times a day, for 6 months; the participants consumed 500 k calories fewer than the usual diet; a randomized, placebo-controlled study | obesity (BMI of 30.0–39.9 kg/m2) | No significant changes in the group treated with resveratrol solely; ↓ BMI, waist circumference and fat mass in the orlistat-treated and O-R groups; the most effective one was the O-R combination | [55] |
| n = 74; 150 mg or 1000 mg/day, for 16 weeks; a randomized, placebo-controlled study | obesity (BMI of 33.8 ± 0.44 kg/m2) | No effect on blood pressure, body composition, lipid deposition in the liver or striated muscle; no beneficial effect on glucose and lipid metabolism; 1000 mg dose increased the total cholesterol and LDL compared to placebo group | [46] |
| n = 45; 75 mg twice a day, for 4 weeks; a randomized, placebo-controlled study | overweight or slight obesity (BMI of 28.3 ± 3.2 kg/m2) | No changes in plasma biomarkers of endothelial function or inflammation (both in the fasting state and postprandial phase); no changes in serum triglyceride and insulin level | [49] |
| n = 38; a combination of 282 or 80 mg/day of the epigallocatechin-3-gallate and resveratrol (EGCG + RES), for 12 weeks; a randomized, placebo-controlled study | overweight or obesity (BMI of 29.7 ± 0.5 kg/m2) | ↓ visceral adipose tissue mass; no effect on insulin-stimulated glucose disposal, endogenous glucose production or lipolysis; | [53] |
| n = 25; 282 or 80 mg/day of the EGCG + RES, for 12 weeks; a randomized, placebo-controlled study | overweight or obesity (BMI of 29.7 ± 1.1 kg/m2) | No changes in adipocyte size or surface area in abdominal subcutaneous adipose tissue; EGCG + RES downregulated pathways contributing to adipogenesis, cell cycle and apoptosis in the abdominal subcutaneous adipose tissue | [54] |
| n = 112; 75 mg, twice a day, for 12 weeks; a randomized, placebo-controlled study | overweight or obesity (BMI ≥ 27 kg/m2), insulin resistance | No effects on cardiometabolic risk parameters and liver fat content | [48] |
| n = 28; 1000 mg, twice a day, for 30 days; a randomized, placebo-controlled study | obesity (BMI of 30–40 kg/m2), metabolic syndrome | No changes in insulin resistance; no changes in adipose tissue metabolism | [50] |
| n = 41; 150 mg/day, for 6 months; a randomized, placebo-controlled study | overweight or obesity (BMI of 27–35 kg/m2) | No effects on intrahepatic lipid level, energy metabolism, blood pressure, physical performance, quality of life and sleep | [51] |
| n = 25; 250 mg/day, with physical training and diet, for 3 months; a randomized, placebo controlled, double-blind study | obesity (BMI ≥ 30 kg/m2), metabolic syndrome | Resveratrol potentiated beneficial effects of diet and physical training; ↓ VLDL and the total cholesterol in blood plasma | [52] |
BMI—body mass index.
Table 4.
Pancreatic lipase inhibitory effects of exemplary phytochemicals, present also in rhubarbs.
Table 4.
Pancreatic lipase inhibitory effects of exemplary phytochemicals, present also in rhubarbs.
| Compound | Phytochemical Classification | Pancreatic Lipase Inhibitory Effects | References | |
|---|---|---|---|---|
| IC50 for the Examined Compound | IC50 for Orlistat | |||
| Caffeic acid | Phenolic acids | 401.5 μM | 4.0 μM | [64] |
| Chlorogenic acid | 110.0 μM 114.0 μM | 0.23 μM ND | [65] | |
| p-Coumaric acid | 170.2 μM | 4.0 μM | [64] | |
| Ellagic acid | 44.78 μM | 0.23 μM | [65] | |
| Ferulic acid | 2.49 μM | 4.0 μM | [65] | |
| Cyanidin-3-rutinoside | Anthocyanidins and their derivatives | 188.28 μM | ND | [67] |
| 59.4 μM | 31.7 μM | [68] | ||
| Delphinidin-3-glucoside | 223.26 μM | ND | [68] | |
| Procyanidin B2 | Proanthocyanidins | 7.96 μM | ND | [69] |
| cis-Piceid | Stilbene derivatives | 76.1 μM | 0.7 μM | [70] |
| trans-Piceid | 121.5 μM | 0.7 μM | [70] | |
| trans-Resveratrol | >200 μM | 0.7 μM | [70] | |
| Kaempferol-3-O-rutinoside | Flavonoids and their glycosides | 2.9 μM | 1.45 μM | [71] |
| Quercetin | 421.1 μM | ND | [72] | |
| 146 μM | 1.45 μM | [71] | ||
| Quercetin-3-O-β-D-glucuronide | 94 μM | ND | [73] | |
| Rutin | 149 μM | 1.45 μM | [71] | |
ND—not determined.
Table 5.
The ability of rhubarb-derived substances to regulate the glucose homeostasis demonstrated in animal studies.
Table 5.
The ability of rhubarb-derived substances to regulate the glucose homeostasis demonstrated in animal studies.
| The Examined Rhubarb Compounds or Extracts | Experimental Model/Doses | Main Findings | References |
|---|---|---|---|
| desoxyrhapontigenin, emodin and chrysophanol, from roots of R. rhabarbarum | mice | ↓ postprandial hyperglycaemia by 35.8, 29.5, 42.3%, respectively | [109] |
| 70% ethanol Rhei Rhizoma extract | streptozotocin-induced diabetes in mice/5 mg/kg b.w. (body weight), 8 weeks | ↑ insulin-stimulated glucose uptake, ↓ carbohydrate digestion via inhibiting alpha-glucoamylase | [109] |
| decoction from R. turkestanicum rhizome | diabetic rats/200–600 mg/kg b.w, 3 weeks | no effects on serum glucose, ↓ serum triglyceride level | [113] |
| standardized extract from R. turkestanicum roots | streptozotocin-induced diabetes in rats/100, 200 and 300 mg/kg b.w., 4 weeks | ↓ blood glucose, ↓ diabetic changes in kidneys, liver and heart | [110] |
| R. emodi extract | rats treated with glucocorticoids/10, 20 and 30 g of rhubarb powder/kg of diet, 8 weeks | ↓ blood glucose and immunity markers | [111] |
| anthraquinone-glycoside preparation from R. palmatum | high-fat diet-induced type 2 diabetes mellitus in rats/100, 200, and 400 mg/kg b.w., 6 weeks | ↓ fasting blood glucose, ↓ total cholesterol and triglyceride levels, improvement in pathological changes in the liver, kidney and pancreatic tissues | [112] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liudvytska, O.; Kolodziejczyk-Czepas, J. A Review on Rhubarb-Derived Substances as Modulators of Cardiovascular Risk Factors—A Special Emphasis on Anti-Obesity Action. Nutrients 2022, 14, 2053. https://doi.org/10.3390/nu14102053
AMA Style
Liudvytska O, Kolodziejczyk-Czepas J. A Review on Rhubarb-Derived Substances as Modulators of Cardiovascular Risk Factors—A Special Emphasis on Anti-Obesity Action. Nutrients. 2022; 14(10):2053. https://doi.org/10.3390/nu14102053
Chicago/Turabian StyleLiudvytska, Oleksandra, and Joanna Kolodziejczyk-Czepas. 2022. "A Review on Rhubarb-Derived Substances as Modulators of Cardiovascular Risk Factors—A Special Emphasis on Anti-Obesity Action" Nutrients 14, no. 10: 2053. https://doi.org/10.3390/nu14102053
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.