Genetic Variation Interacts with Selenium Exposure Regarding Breast Cancer Risk: Assessing Dietary Intake, Serum Levels and Genetically Elevated Selenium Levels

 , ,
, ,  and
and
Abstract
:1. Introduction
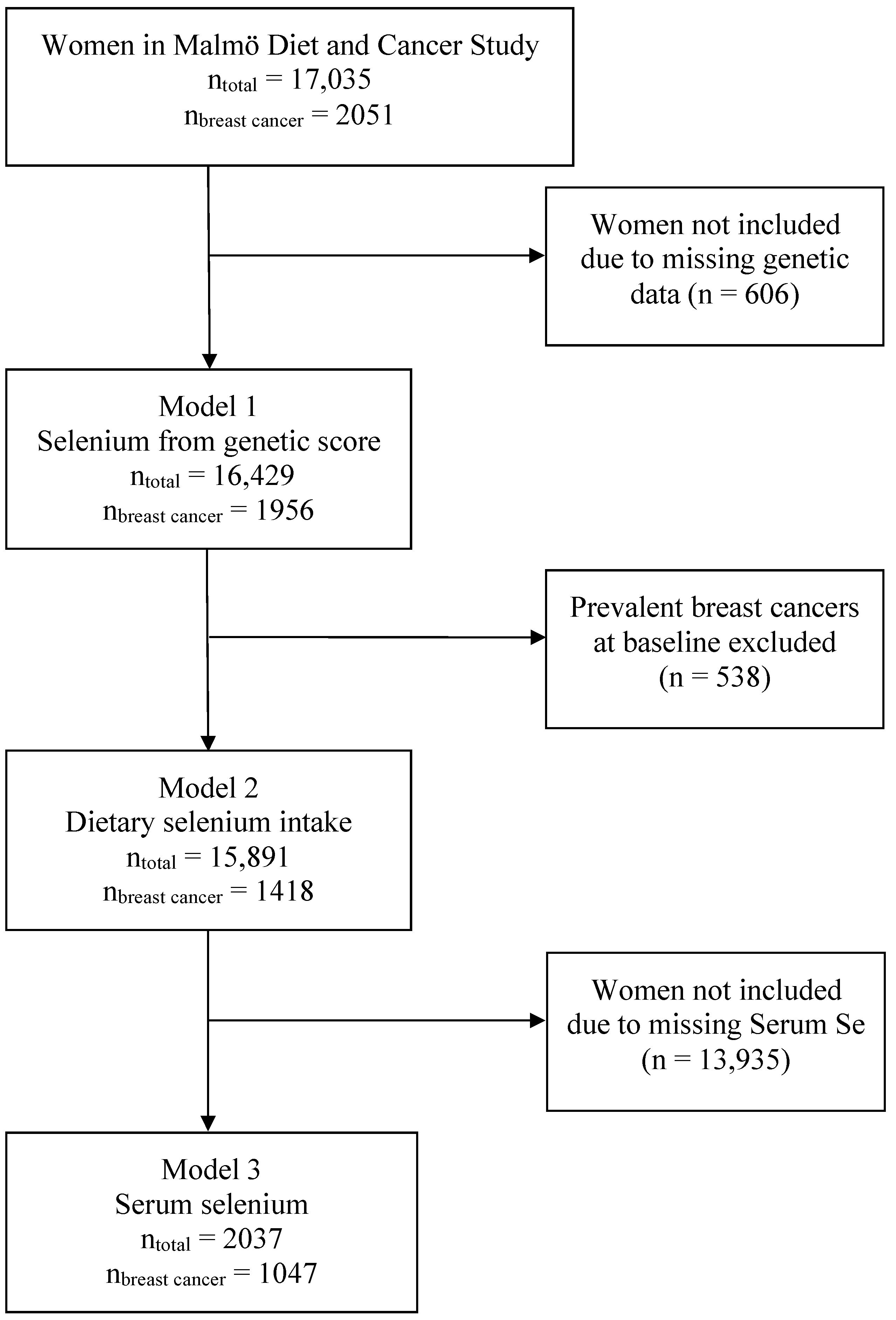
2. Materials and Methods
2.1. The Malmö Diet and Cancer Study (MDCS)
2.2. Inclusion and Exclusion
2.3. Genotyping
2.4. Selection of Candidate SNPs for Interaction
2.5. Allele Score for Genetically Elevated Selenium
2.6. Dietary Selenium Intake
2.7. Serum Selenium
2.8. Endpoint
2.9. Missing Values
2.10. Statistical Analysis
3. Results
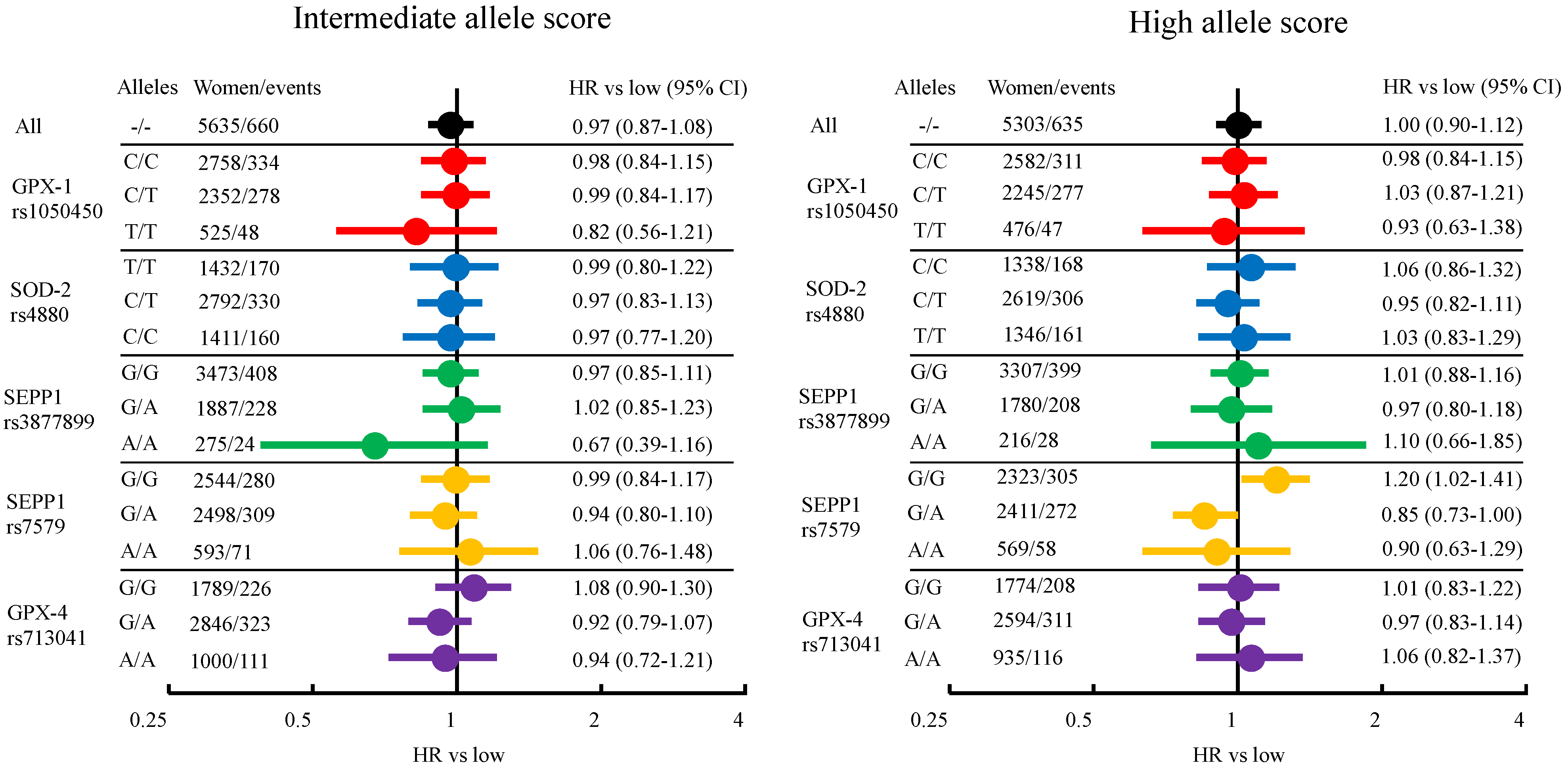
3.1. Breast Cancer Risk from Genetically Elevated Selenium and Individual SNPs
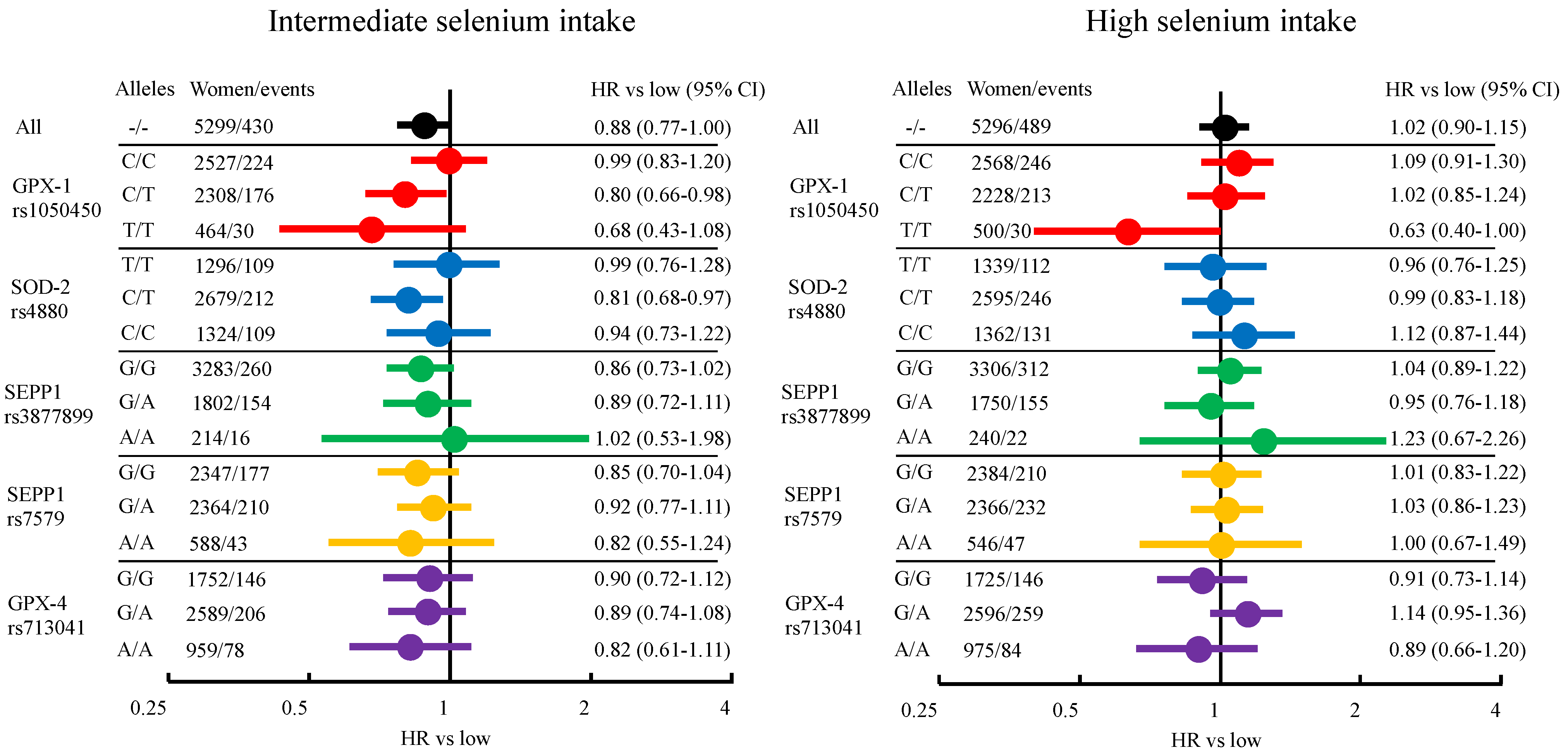
3.2. Breast Cancer Risk from Dietary Selenium Intake
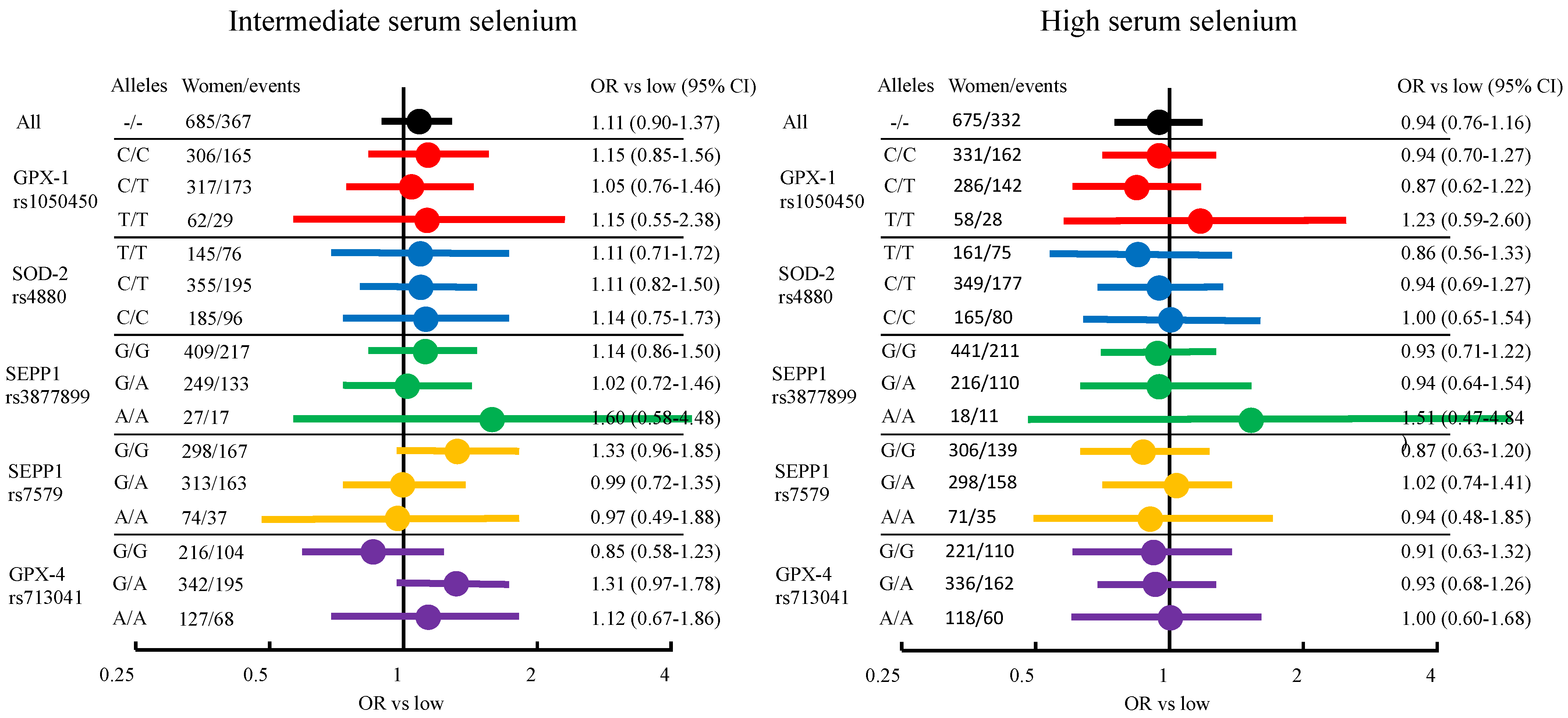
3.3. Breast Cancer Risk from Serum Selenium
4. Discussion
Strengths and Limitations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fontelles, C.C.; Ong, T.P. Selenium and breast cancer risk: Focus on cellular and molecular mechanisms. Adv. Cancer Res. 2017, 136, 173–192. [Google Scholar] [CrossRef] [PubMed]
- Vinceti, M.; Filippini, T.; Del Giovane, C.; Dennert, G.; Zwahlen, M.; Brinkman, M.P.; Zeegers, M.; Horneber, M.; D’Amico, R.; Crespi, C.M. Selenium for preventing cancer. Cochrane Database Syst. Rev. 2018, 1, CD005195. [Google Scholar] [CrossRef] [PubMed]
- Rejali, L.; Jaafar, M.H.; Ismail, N.H. Serum selenium level and other risk factors for breast cancer among patients in a Malaysian hospital. Environ. Health Prev. Med. 2007, 12, 105–110. [Google Scholar] [CrossRef]
- Charalabopoulos, K.; Kotsalos, A.; Batistatou, A.; Vezyraki, P.; Peschos, D.; Kalfakakou, V.; Evangelou, A.; Charalabopoulos, A. Selenium in serum and neoplastic tissue in breast cancer: Correlation with CEA. Br. J. Cancer 2006, 95, 674–676. [Google Scholar] [CrossRef] [PubMed]
- Kadkol, S.; Diamond, A.M. The interaction between dietary selenium intake and genetics in determining cancer risk and outcome. Nutrients 2020, 12, 2424. [Google Scholar] [CrossRef]
- Méplan, C.; Hesketh, J. Selenium and cancer: A story that should not be forgotten—Insights from genomics. Cancer Treat. Res. 2014, 159, 145–166. [Google Scholar] [CrossRef]
- Kryukov, G.V.; Castellano, S.; Novoselov, S.V.; Lobanov, A.V.; Zehtab, O.; Guigoό, R.; Gladyshev, V.N. Characterization of mammalian selenoproteomes. Science 2003, 300, 1439–1443. [Google Scholar] [CrossRef] [Green Version]
- Davis, C.D.; Tsuji, P.A.; Milner, J.A. Selenoproteins and cancer prevention. Annu. Rev. Nutr. 2012, 32, 73–95. [Google Scholar] [CrossRef]
- Sunde, R.A.; Raines, A.M.; Barnes, K.M.; Evenson, J.K. Selenium status highly regulates selenoprotein mRNA levels for only a subset of the selenoproteins in the selenoproteome. Biosci. Rep. 2009, 29, 329–338. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.J.; Diamond, A.M. Role of glutathione peroxidase 1 in breast cancer: Loss of heterozygosity and allelic differences in the response to selenium. Cancer Res. 2003, 63, 3347–3351. [Google Scholar]
- Li, H.; Kantoff, P.; Giovannucci, E.; Leitzmann, M.F.; Gaziano, J.M.; Stampfer, M.J.; Ma, J. Manganese superoxide dismutase polymorphism, prediagnostic antioxidant status, and risk of clinical significant prostate cancer. Cancer Res. 2005, 65, 2498–2504. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meplan, C.; Dragsted, L.O.; Ravn-Haren, G.; Tjonneland, A.; Vogel, U.; Hesketh, J. Association between polymorphisms in glutathione peroxidase and selenoprotein P genes, glutathione peroxidase activity, HRT use and breast cancer risk. PLoS ONE 2013, 8, e73316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cox, D.G.; Tamimi, R.M.; Hunter, D.J. Gene × Gene interaction between MnSOD and GPX-1 and breast cancer risk: A nested case-control study. BMC Cancer 2006, 6, 217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jablonska, E.; Gromadzinska, J.; Peplonska, B.; Fendler, W.; Reszka, E.; Krol, M.B.; Wieczorek, E.; Bukowska, A.; Gresner, P.; Galicki, M.; et al. Lipid peroxidation and glutathione peroxidase activity relationship in breast cancer depends on functional polymorphism of GPX1. BMC Cancer 2015, 15, 657. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, J.; Vanderlelie, J.J.; Perkins, A.V.; Redman, C.W.G.; Ahmadi, K.R.; Rayman, M.P. Genetic polymorphisms that affect selenium status and response to selenium supplementation in United Kingdom pregnant women. Am. J. Clin. Nutr. 2016, 103, 100–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papadimitriou, N.; Dimou, N.; Gill, D.; Tzoulaki, I.; Murphy, N.; Riboli, E.; Lewis, S.J.; Martin, R.M.; Gunter, M.J.; Tsilidis, K.K. Genetically predicted circulating concentrations of micronutrients and risk of breast cancer: A Mendelian randomization study. Int. J. Cancer 2021, 148, 646–653. [Google Scholar] [CrossRef]
- Guo, D.; Hendryx, M.; Liang, X.; Manson, J.E.; He, K.; Vitolins, M.Z.; Li, Y.; Luo, J. Association between selenium intake and breast cancer risk: Results from the Women’s Health Initiative. Breast Cancer Res. Treat. 2020, 183, 217–226. [Google Scholar] [CrossRef]
- Sandsveden, M.; Manjer, J. Selenium and breast cancer risk: A prospective nested case-control study on serum selenium levels, smoking habits and overweight. Int. J. Cancer 2017, 141, 1741–1750. [Google Scholar] [CrossRef] [Green Version]
- Xia, Y.; Hill, K.E.; Li, P.; Xu, J.; Zhou, D.; Motley, A.K.; Wang, L.; Byrne, D.W.; Burk, R.F. Optimization of selenoprotein P and other plasma selenium biomarkers for the assessment of the selenium nutritional requirement: A placebo-controlled, double-blind study of selenomethionine supplementation in selenium-deficient Chinese subjects. Am. J. Clin. Nutr. 2010, 92, 525–531. [Google Scholar] [CrossRef] [Green Version]
- Nordic Nutrition Recommendations. Nordic Nutrition Recommendations 2012: Integrating Nutrition and Physical Activity, 5th ed.; Nordic Council of Ministers: Copenhagen, Denmark, 2014. [Google Scholar]
- European Food Safety Authority EFSA. Scientific opinion on dietary reference values for selenium. EFSA J. 2014, 12, 3846. [Google Scholar] [CrossRef]
- World Health Organization. Vitamin and Mineral Requirements in Human Nutrition, 2nd ed.; Report of a Joint FAO/WHO Expert Consultation, Bangkok, Thailand, 21–30 September 1998; World Health Organization: Geneva, Switzerland, 2004. [Google Scholar]
- Bengtsson, Y.; Sandsveden, M.; Manjer, J. Risk of breast cancer in relation to dietary intake of selenium and serum selenium as a marker of dietary intake: A prospective cohort study within The Malmö Diet and Cancer Study. Cancer Causes Control 2021, 32, 815–826. [Google Scholar] [CrossRef] [PubMed]
- Manjer, J.; Elmstahl, S.; Janzon, L.; Berglund, G. Invitation to a population-based cohort study: Differences between subjects recruited using various strategies. Scand. J. Public Health 2002, 30, 103–112. [Google Scholar] [CrossRef] [PubMed]
- Almquist, M.; Bondeson, A.-G.; Bondeson, L.; Malm, J.; Manjer, J. Serum levels of vitamin D, PTH and calcium and breast cancer risk-a prospective nested case-control study. Int. J. Cancer 2010, 127, 2159–2168. [Google Scholar] [CrossRef]
- Hindy, G.; Aragam, K.G.; Ng, K.; Chaffin, M.; Lotta, L.A.; Baras, A.; Drake, I.; Orho-Melander, M.; Melander, O.; Kathiresan, S.; et al. Genome-wide polygenic score, clinical risk factors, and long-term trajectories of coronary artery disease. Arter. Thromb. Vasc. Biol. 2020, 40, 2738–2746. [Google Scholar] [CrossRef] [PubMed]
- McCarthy, S.; Das, S.; Kretzschmar, W.; Delaneau, O.; Wood, A.R.; Teumer, A.; Kang, H.M.; Fuchsberger, C.; Danecek, P.; Sharp, K.; et al. A reference panel of 64,976 haplotypes for genotype imputation. Nat. Genet. 2016, 48, 1279–1283. [Google Scholar]
- Cornelis, M.C.; Fornage, M.; Foy, M.; Xun, P.; Gladyshev, V.N.; Morris, S.; Chasman, D.I.; Hu, F.B.; Rimm, E.B.; Kraft, P.; et al. Genome-wide association study of selenium concentrations. Hum. Mol. Genet. 2015, 24, 1469–1477. [Google Scholar] [CrossRef] [Green Version]
- Evans, D.M.; Zhu, G.; Dy, V.; Heath, A.C.; Madden, P.A.F.; Kemp, J.P.; McMahon, G.; St Pourcain, B.; Timpson, N.J.; Golding, J.; et al. Genome-wide association study identifies loci affecting blood copper, selenium and zinc. Hum. Mol. Genet. 2013, 22, 3998–4006. [Google Scholar] [CrossRef] [Green Version]
- Wirfält, E.; Sonestedt, E. The Modified Diet History Methodology of the Malmö Diet Cancer Cohort. Available online: https://www.malmo-kohorter.lu.se/sites/malmo-kohorter.lu.se/files/mdc_diet_history.pdf (accessed on 29 March 2021).
- Willett, W.C.; Howe, G.R.; Kushi, L.H. Adjustment for total energy intake in epidemiologic studies. Am. J. Clin. Nutr. 1997, 65, 1220S–1228S. [Google Scholar] [CrossRef]
- Hu, Y.; Benya, R.V.; Carroll, R.E.; Diamond, A.M. Allelic loss of the gene for the GPX1 selenium-containing protein is a common event in cancer. J. Nutr. 2005, 135, 3021S–3024S. [Google Scholar] [CrossRef]
- Moscow, J.A.; Schmidt, L.; Ingram, D.T.; Gnarra, J.; Johnson, B.; Cowan, K.H. Loss of heterozygosity of the human cytosolic glutathione peroxidase I gene in lung cancer. Carcinogenesis 1994, 15, 2769–2773. [Google Scholar] [CrossRef] [Green Version]
- Bera, S.; Weinberg, F.; Ekoue, D.N.; Ansenberger-Fricano, K.; Mao, M.; Bonini, M.G.; Diamond, A.M. Natural allelic variations in glutathione peroxidase-1 affect its subcellular localization and function. Cancer Res. 2014, 74, 5118–5126. [Google Scholar] [CrossRef] [Green Version]
- Brigelius-Flohé, R.; Maiorino, M. Glutathione peroxidases. Biochim. Biophys. Acta (BBA)—Gen. Subj. 2013, 1830, 3289–3303. [Google Scholar] [CrossRef] [PubMed]
- Miriyala, S.; Spasojevic, I.; Tovmasyan, A.; Salvemini, D.; Vujaskovic, Z.; Clair, D.S.; Batinic-Haberle, I. Manganese superoxide dismutase, MnSOD and its mimics. Biochim. Biophys. Acta (BBA)—Mol. Basis Dis. 2012, 1822, 794–814. [Google Scholar] [CrossRef] [Green Version]
- Ekoue, D.N.; He, C.; Diamond, A.M.; Bonini, M.G. Manganese superoxide dismutase and glutathione peroxidase-1 contribute to the rise and fall of mitochondrial reactive oxygen species which drive oncogenesis. Biochim. Biophys. Acta Bioeneg. 2017, 1858, 628–632. [Google Scholar] [CrossRef] [PubMed]
- Ravn-Haren, G.; Olsen, A.; Tjønneland, A.; Dragsted, L.O.; Nexø, B.A.; Wallin, H.; Overvad, K.; Raaschou-Nielsen, O.; Vogel, U. Associations between GPX1 Pro198Leu polymorphism, erythrocyte GPX activity, alcohol consumption and breast cancer risk in a prospective cohort study. Carcinogenesis 2006, 27, 820–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Zhou, G.-W.; Wang, N.; Wang, Y.-J. GPX1 Pro198Leu polymorphism and breast cancer risk: A meta-analysis. Breast Cancer Res. Treat. 2010, 124, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Shu, X.; Shu, X.-O.; Bolla, M.K.; Kweon, S.-S.; Cai, Q.; Michailidou, K.; Wang, Q.; Dennis, J.; Park, B.; et al. Re-evaluating genetic variants identified in candidate gene studies of breast cancer risk using data from nearly 280,000 women of Asian and European ancestry. eBioMedicine 2019, 48, 203–211. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Blein, S.; Berndt, S.; Joshi, A.D.; Campa, D.; Ziegler, R.G.; Riboli, E.; Cox, D.G.; Gaudet, M.M.; Stevens, V.L.; Diver, W.R.; et al. Factors associated with oxidative stress and cancer risk in the Breast and Prostate Cancer Cohort Consortium. Free Radic. Res. 2014, 48, 380–386. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.; Zhang, R.; Chen, N.; Yang, L.; Wang, Y.; Sun, Y.; Huang, L.; Zhu, M.; Ji, Y.; Li, W. Association between glutathione peroxidase-1 (GPX1) Rs1050450 polymorphisms and cancer risk. Int. J. Clin. Exp. Pathol. 2017, 10, 9527–9540. [Google Scholar]
- Jablonska, E.; Gromadzinska, J.; Reszka, E.; Wasowicz, W.; Sobala, W.; Szeszenia-Dabrowska, N.; Boffetta, P. Association between GPx1 Pro198Leu polymorphism, GPx1 activity and plasma selenium concentration in humans. Eur. J. Nutr. 2009, 48, 383–386. [Google Scholar] [CrossRef]
- Zhuo, P.; Goldberg, M.; Herman, L.; Lee, B.-S.; Wang, H.; Brown, R.L.; Foster, C.B.; Peters, U.; Diamond, A.M. Molecular consequences of genetic variations in the glutathione peroxidase 1 selenoenzyme. Cancer Res. 2009, 69, 8183–8190. [Google Scholar] [CrossRef] [Green Version]
- Schumacker, P.T. Reactive oxygen species in cancer cells: Live by the sword, die by the sword. Cancer Cell 2006, 10, 175–176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barlow, L.; Westergren, K.; Holmberg, L.; Talbäck, M. The completeness of the Swedish Cancer Register—A sample survey for year 1998. Acta Oncol. 2009, 48, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Riboli, E.; Elmståhl, S.; Saracci, R.; Gullberg, B.; Lindgärde, F. The Malmo Food Study: Validity of two dietary assessment methods for measuring nutrient intake. Int. J. Epidemiol. 1997, 26, 161S–173S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Combs, G.F., Jr. Biomarkers of selenium status. Nutrients 2015, 7, 2209–2236. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vinceti, M.; Grill, P.; Malagoli, C.; Filippini, T.; Storani, S.; Malavolti, M.; Michalke, B. Selenium speciation in human serum and its implications for epidemiologic research: A cross-sectional study. J. Trace Elem. Med. Biol. 2015, 31, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yarmolinsky, J.; Bonilla, C.; Haycock, P.C.; Langdon, R.J.Q.; Lotta, L.A.; Langenberg, C.; Relton, C.L.; Lewis, S.J.; Evans, D.M.; Smith, G.D.; et al. Circulating selenium and prostate cancer risk: A Mendelian randomization analysis. J. Natl. Cancer Inst. 2018, 110, 1035–1038. [Google Scholar] [CrossRef] [Green Version]
- Lawlor, D.A.; Tilling, K.; Smith, G.D. Triangulation in aetiological epidemiology. Int. J. Epidemiol. 2017, 45, 1866–1886. [Google Scholar] [CrossRef] [Green Version]
- Vittinghoff, E.; McCulloch, C.E. Relaxing the rule of ten events per variable in logistic and cox regression. Am. J. Epidemiol. 2007, 165, 710–718. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No Breast Cancer (n = 14,473) | Incident Breast Cancer (n = 1418) | Breast Cancer at Baseline (n = 538) | No Genetic Data (n = 606) | ||
|---|---|---|---|---|---|
| Valid Column % | Valid Column % | Valid Column % | Valid Column % | ||
| Age at baseline * (SD) | 57.4 (7.9) | 56.0 (7.3) | 60.7 (7.4) | 57.1 (7.3) | |
| Age at menarche * (SD) | 13.6 (1.5) | 13.5 (1.4) | 13.7 (1.4) | 13.6 (1.5) | |
| Parity | 0 | 12.7 | 14.0 | 16.7 | 14.8 |
| 1 | 21.7 | 19.5 | 28.4 | 22.5 | |
| 2 | 41.5 | 46.5 | 35.8 | 41.9 | |
| 3 | 17.2 | 15.1 | 13.4 | 13.0 | |
| 4 or more | 6.8 | 4.8 | 5.7 | 7.9 | |
| Missing | 1.6 | 2.0 | 1.9 | 3.8 | |
| Ever use of oral | Never | 51.1 | 45.3 | 56.8 | 54.1 |
| contraceptives | Ever | 48.9 | 54.7 | 43.2 | 45.9 |
| Married or cohabiting | No | 33.1 | 31.5 | 36.2 | 36.5 |
| Yes | 66.9 | 68.5 | 63.8 | 63.5 | |
| Bilateral | No | 98.6 | 98.6 | 98.0 | 97.7 |
| oophorectomy | Yes | 1.4 | 1.4 | 2.0 | 2.3 |
| BMI | BMI < 25 | 53.7 | 51.0 | 50.4 | 50.4 |
| BMI 25–30 | 32.7 | 35.2 | 35.5 | 35.3 | |
| BMI ≥ 30 | 13.5 | 13.8 | 14.1 | 14.3 | |
| Education | O-level college | 70.0 | 67.3 | 72.6 | 69.2 |
| A-level college | 7.1 | 6.5 | 5.4 | 6.8 | |
| University | 23.0 | 26.2 | 22.0 | 24.0 | |
| Socio-economic index | Manual | 38.6 | 33.4 | 37.2 | 38.7 |
| Non-manual | 53.7 | 59.7 | 57.0 | 53.8 | |
| Employer | 7.7 | 6.9 | 5.8 | 7.5 | |
| Missing | 1.0 | 1.1 | 1.5 | 1.5 | |
| HRT use at baseline | No | 82.4 | 73.7 | 95.0 | 81.3 |
| Yes | 17.6 | 26.3 | 5.0 | 18.7 | |
| Alcohol intake | No alcohol | 7.8 | 5.6 | 9.1 | 5.8 |
| <15 g/day | 64.2 | 62.8 | 61.5 | 65.8 | |
| 15–30 g/day | 14.0 | 15.5 | 13.0 | 12.7 | |
| >30 g/day | 2.2 | 4.2 | 3.4 | 2.1 | |
| Infrequent use | 11.8 | 12.0 | 13.0 | 13.5 | |
| Age at menopause ** | Hysterectomy | 0.5 | 0.6 | 0.9 | 0.5 |
| Pre-/peri | 33.3 | 41.5 | 17.6 | 31.7 | |
| ≤44 | 10.1 | 8.5 | 8.2 | 12.9 | |
| 45–54 | 50.2 | 43.7 | 62.1 | 49.6 | |
| ≥55 | 5.9 | 5.8 | 11.2 | 5.3 | |
| Missing | 0.7 | 0.8 | 0.6 | 1.2 | |
| Age at first childbirth | ≤20 | 17.1 | 15.9 | 13.3 | 18.4 |
| 21–25 | 36.0 | 35.2 | 31.3 | 34.0 | |
| 26–30 | 24.8 | 24.5 | 27.3 | 22.5 | |
| ≥31 | 9.3 | 10.4 | 11.6 | 10.5 | |
| Nullipara | 12.8 | 14.0 | 16.7 | 14.8 | |
| Missing | 1.7 | 2.0 | 1.9 | 3.8 |
| Women with Genetic Data (n = 16,429) | ||||
|---|---|---|---|---|
| SNP | Alleles | Events/Women (%) |
Events/ 100,000 py | HR (95% CI) |
| GPX-1 | C/C | 965/7951 (12.1%) | 156 | 1 |
| rs1050450 | C/T | 843/6988 (12.1%) | 155 | 0.99 (0.90–1.08) |
| T/T | 148/1490 (9.9%) | 127 | 0.81 (0.68–0.96) | |
| SOD-2 | T/T | 498/4113 (12.1%) | 156 | 1 |
| rs4880 | T/C | 980/8216 (11.9%) | 153 | 0.98 (0.88–1.09) |
| C/C | 478/4100 (11.7%) | 149 | 0.96 (0.84–1.08) | |
| SEPP1 | G/G | 1220/10208 (12.0%) | 153 | 1 |
| rs3877899 | G/A | 655/5494 (11.9%) | 153 | 1.00 (0.91–1.10) |
| A/A | 81/727 (11.1%) | 144 | 0.95 (0.76–1.19) | |
| SEPP1 | G/G | 864/7370 (11.7%) | 150 | 1 |
| rs7579 | G/A | 898/7328 (12.3%) | 157 | 1.04 (0.95–1.14) |
| A/A | 194/1731 (11.2%) | 144 | 0.96 (0.82–1.12) | |
| GPX-4 | G/G | 651/5405 (12.0%) | 154 | 1 |
| rs713041 | G/A | 959/8089 (11.9%) | 152 | 0.99 (0.90–1.10) |
| A/A | 346/2935 (11.8%) | 151 | 0.98 (0.68–1.12) | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sandsveden, M.; Bengtsson, Y.; Melander, O.; Rosendahl, A.H.; Manjer, J. Genetic Variation Interacts with Selenium Exposure Regarding Breast Cancer Risk: Assessing Dietary Intake, Serum Levels and Genetically Elevated Selenium Levels. Nutrients 2022, 14, 826. https://doi.org/10.3390/nu14040826
Sandsveden M, Bengtsson Y, Melander O, Rosendahl AH, Manjer J. Genetic Variation Interacts with Selenium Exposure Regarding Breast Cancer Risk: Assessing Dietary Intake, Serum Levels and Genetically Elevated Selenium Levels. Nutrients. 2022; 14(4):826. https://doi.org/10.3390/nu14040826
Chicago/Turabian StyleSandsveden, Malte, Ylva Bengtsson, Olle Melander, Ann H. Rosendahl, and Jonas Manjer. 2022. "Genetic Variation Interacts with Selenium Exposure Regarding Breast Cancer Risk: Assessing Dietary Intake, Serum Levels and Genetically Elevated Selenium Levels" Nutrients 14, no. 4: 826. https://doi.org/10.3390/nu14040826