
A Milk-Fat Based Diet Increases Metastasis in the MMTV-PyMT Mouse Model of Breast Cancer

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Mouse Models
2.2. Tumor Monitoring
2.3. RNA Extraction and Quantitative Real-Time PCR
2.4. Statistical Analysis
3. Results
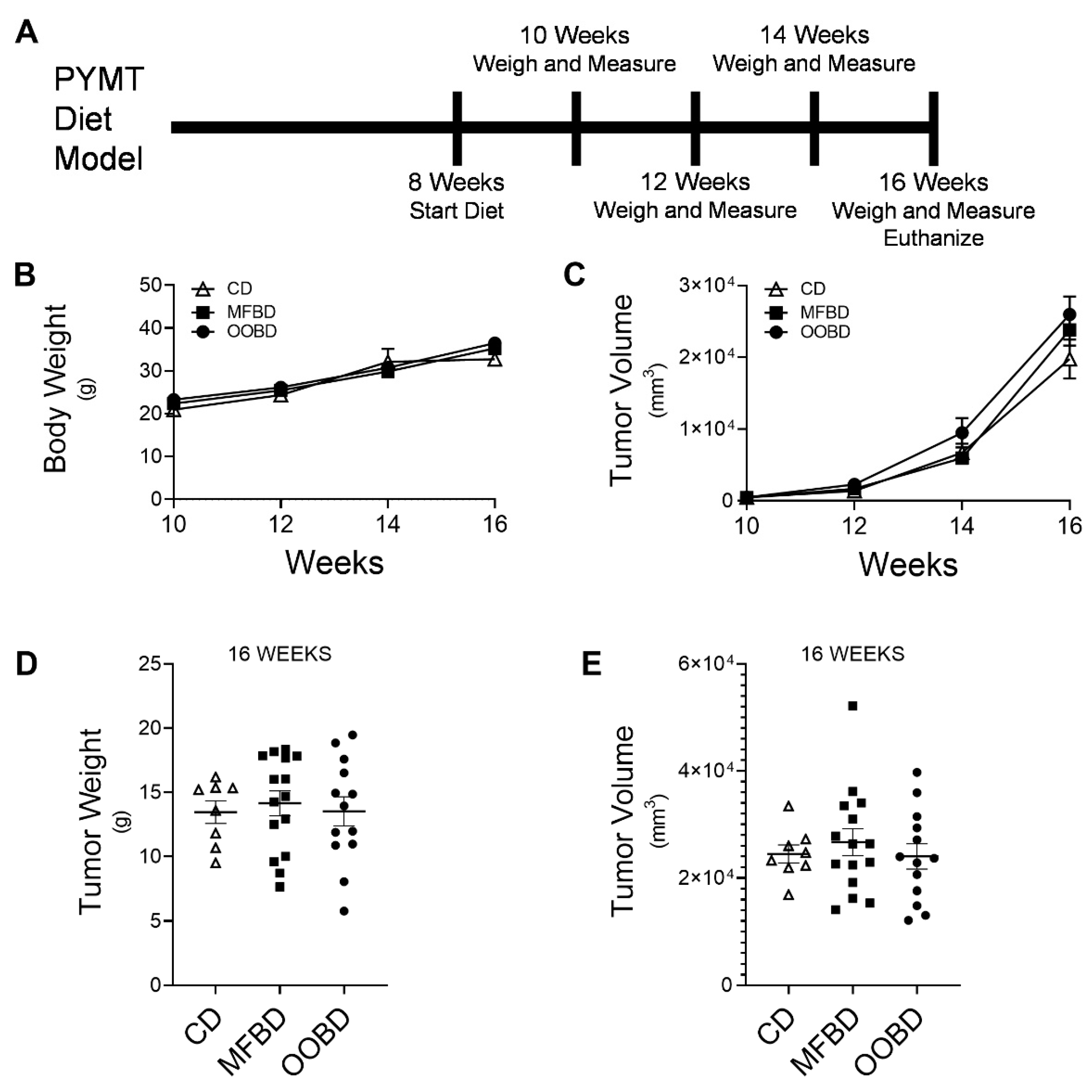
3.1. Milk Fat and Olive Oil Diets Do Not Alter Tumor Weight or Volume in MMTV-PyMT Mice
3.2. MFBD Fed Mice Exhibit Increased Lung Metastasis in MMTV-PyMT Mice
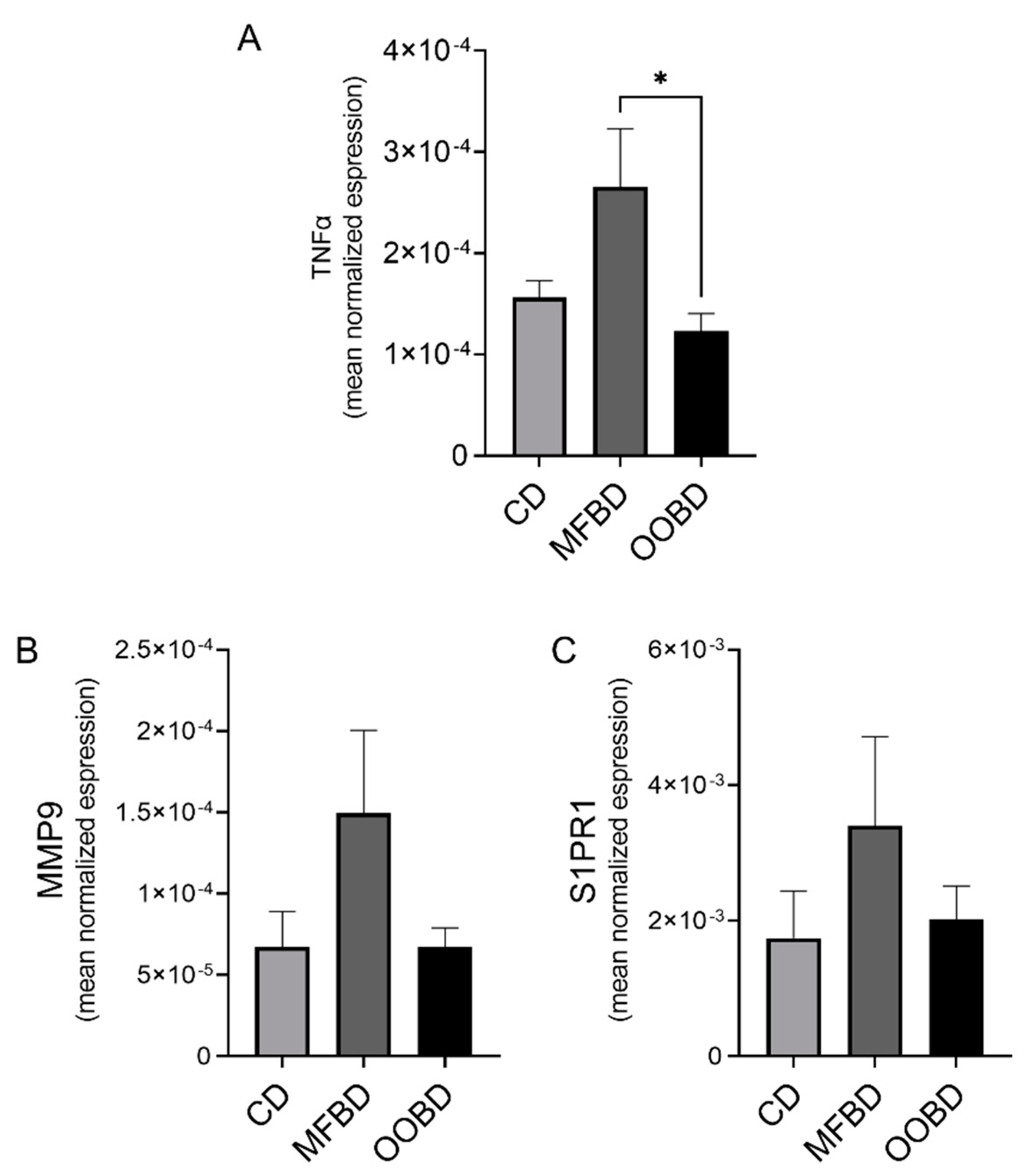
3.3. MFBD Increases TNFα Expression in Primary Tumors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Sun, Y.S.; Zhao, Z.; Yang, Z.N.; Xu, F.; Lu, H.J.; Zhu, Z.Y.; Shi, W.; Jiang, J.; Yao, P.P.; Zhu, H.P. Risk Factors and Preventions of Breast Cancer. Int. J. Biol. Sci. 2017, 13, 1387–1397. [Google Scholar] [CrossRef] [Green Version]
- Shetty, P.J.; Sreedharan, J. Breast Cancer and Dietary Fat Intake: A correlational study. Nepal. J. Epidemiol. 2019, 9, 812–816. [Google Scholar] [CrossRef]
- Miller, E.R.; Wilson, C.; Chapman, J.; Flight, I.; Nguyen, A.M.; Fletcher, C.; Ramsey, I. Connecting the dots between breast cancer, obesity and alcohol consumption in middle-aged women: Ecological and case control studies. BMC Public Health 2018, 18, 460. [Google Scholar] [CrossRef] [Green Version]
- Thiebaut, A.C.; Kipnis, V.; Chang, S.C.; Subar, A.F.; Thompson, F.E.; Rosenberg, P.S.; Hollenbeck, A.R.; Leitzmann, M.; Schatzkin, A. Dietary fat and postmenopausal invasive breast cancer in the National Institutes of Health-AARP Diet and Health Study cohort. J. Natl. Cancer Inst. 2007, 99, 451–462. [Google Scholar] [CrossRef]
- Hruby, A.; Hu, F.B. The Epidemiology of Obesity: A Big Picture. Pharmacoeconomics 2015, 33, 673–689. [Google Scholar] [CrossRef]
- Anderson, B.O.; Jakesz, R. Breast cancer issues in developing countries: An overview of the Breast Health Global Initiative. World J. Surg. 2008, 32, 2578–2585. [Google Scholar] [CrossRef]
- Porter, P. “Westernizing” women’s risks? Breast cancer in lower-income countries. N. Engl. J. Med. 2008, 358, 213–216. [Google Scholar] [CrossRef]
- Bray, F.; Ferlay, J.; Laversanne, M.; Brewster, D.H.; Gombe Mbalawa, C.; Kohler, B.; Pineros, M.; Steliarova-Foucher, E.; Swaminathan, R.; Antoni, S.; et al. Cancer Incidence in Five Continents: Inclusion criteria, highlights from Volume X and the global status of cancer registration. Int. J. Cancer 2015, 137, 2060–2071. [Google Scholar] [CrossRef] [PubMed]
- Neuhouser, M.L.; Aragaki, A.K.; Prentice, R.L.; Manson, J.E.; Chlebowski, R.; Carty, C.L.; Ochs-Balcom, H.M.; Thomson, C.A.; Caan, B.J.; Tinker, L.F.; et al. Overweight, Obesity, and Postmenopausal Invasive Breast Cancer Risk: A Secondary Analysis of the Women’s Health Initiative Randomized Clinical Trials. JAMA Oncol. 2015, 1, 611–621. [Google Scholar] [CrossRef] [PubMed]
- Widschwendter, P.; Friedl, T.W.; Schwentner, L.; DeGregorio, N.; Jaeger, B.; Schramm, A.; Bekes, I.; Deniz, M.; Lato, K.; Weissenbacher, T.; et al. The influence of obesity on survival in early, high-risk breast cancer: Results from the randomized SUCCESS A trial. Breast Cancer Res. 2015, 17, 129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holmes, M.D.; Hunter, D.J.; Colditz, G.A.; Stampfer, M.J.; Hankinson, S.E.; Speizer, F.E.; Rosner, B.; Willett, W.C. Association of dietary intake of fat and fatty acids with risk of breast cancer. JAMA 1999, 281, 914–920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boyd, N.F.; Stone, J.; Vogt, K.N.; Connelly, B.S.; Martin, L.J.; Minkin, S. Dietary fat and breast cancer risk revisited: A meta-analysis of the published literature. Br. J. Cancer 2003, 89, 1672–1685. [Google Scholar] [CrossRef] [Green Version]
- Sieri, S.; Krogh, V.; Ferrari, P.; Berrino, F.; Pala, V.; Thiebaut, A.C.; Tjonneland, A.; Olsen, A.; Overvad, K.; Jakobsen, M.U.; et al. Dietary fat and breast cancer risk in the European Prospective Investigation into Cancer and Nutrition. Am. J. Clin. Nutr. 2008, 88, 1304–1312. [Google Scholar] [CrossRef]
- Wirfalt, E.; Mattisson, I.; Gullberg, B.; Johansson, U.; Olsson, H.; Berglund, G. Postmenopausal breast cancer is associated with high intakes of omega6 fatty acids (Sweden). Cancer Causes Control. 2002, 13, 883–893. [Google Scholar] [CrossRef]
- Voorrips, L.E.; Brants, H.A.; Kardinaal, A.F.; Hiddink, G.J.; van den Brandt, P.A.; Goldbohm, R.A. Intake of conjugated linoleic acid, fat, and other fatty acids in relation to postmenopausal breast cancer: The Netherlands Cohort Study on Diet and Cancer. Am. J. Clin. Nutr. 2002, 76, 873–882. [Google Scholar] [CrossRef] [Green Version]
- Wolk, A.; Bergstrom, R.; Hunter, D.; Willett, W.; Ljung, H.; Holmberg, L.; Bergkvist, L.; Bruce, A.; Adami, H.O. A prospective study of association of monounsaturated fat and other types of fat with risk of breast cancer. Arch. Intern. Med. 1998, 158, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Murff, H.J.; Shu, X.O.; Li, H.; Yang, G.; Wu, X.; Cai, H.; Wen, W.; Gao, Y.T.; Zheng, W. Dietary polyunsaturated fatty acids and breast cancer risk in Chinese women: A prospective cohort study. Int. J. Cancer 2011, 128, 1434–1441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sczaniecka, A.K.; Brasky, T.M.; Lampe, J.W.; Patterson, R.E.; White, E. Dietary intake of specific fatty acids and breast cancer risk among postmenopausal women in the VITAL cohort. Nutr. Cancer 2012, 64, 1131–1142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attalla, S.; Taifour, T.; Bui, T.; Muller, W. Insights from transgenic mouse models of PyMT-induced breast cancer: Recapitulating human breast cancer progression In Vivo. Oncogene 2021, 40, 475–491. [Google Scholar] [CrossRef] [PubMed]
- Cowen, S.; McLaughlin, S.L.; Hobbs, G.; Coad, J.; Martin, K.H.; Olfert, I.M.; Vona-Davis, L. High-Fat, High-Calorie Diet Enhances Mammary Carcinogenesis and Local Inflammation in MMTV-PyMT Mouse Model of Breast Cancer. Cancers 2015, 7, 1125–1142. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, S.; Yan, L. High-fat Diet Enhances Mammary Tumorigenesis and Pulmonary Metastasis and Alters Inflammatory and Angiogenic Profiles in MMTV-PyMT Mice. Anticancer. Res. 2016, 36, 6279–6287. [Google Scholar] [CrossRef] [PubMed]
- Sundaram, S.; Yan, L. Adipose monocyte chemotactic protein-1 deficiency reduces high-fat diet-enhanced mammary tumorigenesis in MMTV-PyMT mice. J. Nutr. Biochem. 2020, 77, 108313. [Google Scholar] [CrossRef] [PubMed]
- Devlin, M.J.; Robbins, A.; Cosman, M.N.; Moursi, C.A.; Cloutier, A.M.; Louis, L.; Van Vliet, M.; Conlon, C.; Bouxsein, M.L. Differential effects of high fat diet and diet-induced obesity on skeletal acquisition in female C57BL/6J vs. FVB/NJ Mice. Bone Rep. 2018, 8, 204–214. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.C.; Qing, K.; Chen, Y. Diet-induced changes in stearoyl-CoA desaturase 1 expression in obesity-prone and -resistant mice. Obes. Res. 2004, 12, 1264–1270. [Google Scholar] [CrossRef]
- Velazquez, F.N.; Zhang, L.; Viscardi, V.; Trocchia, C.; Hannun, Y.A.; Obeid, L.M.; Snider, A.J. Loss of sphingosine kinase 1 increases lung metastases in the MMTV-PyMT mouse model of breast cancer. PLoS ONE 2021, 16, e0252311. [Google Scholar] [CrossRef]
- Choi, S.; Snider, J.M.; Cariello, C.P.; Lambert, J.M.; Anderson, A.K.; Cowart, L.A.; Snider, A.J. Sphingosine kinase 1 is required for myristate-induced TNFalpha expression in intestinal epithelial cells. Prostaglandins Other Lipid Mediat. 2020, 149, 106423. [Google Scholar] [CrossRef]
- Ewertz, M.; Jensen, M.B.; Gunnarsdottir, K.A.; Hojris, I.; Jakobsen, E.H.; Nielsen, D.; Stenbygaard, L.E.; Tange, U.B.; Cold, S. Effect of obesity on prognosis after early-stage breast cancer. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2011, 29, 25–31. [Google Scholar] [CrossRef]
- Protani, M.; Coory, M.; Martin, J.H. Effect of obesity on survival of women with breast cancer: Systematic review and meta-analysis. Breast Cancer Res. Treat. 2010, 123, 627–635. [Google Scholar] [CrossRef]
- Davie, S.A.; Maglione, J.E.; Manner, C.K.; Young, D.; Cardiff, R.D.; MacLeod, C.L.; Ellies, L.G. Effects of FVB/NJ and C57Bl/6J strain backgrounds on mammary tumor phenotype in inducible nitric oxide synthase deficient mice. Transgenic Res. 2007, 16, 193–201. [Google Scholar] [CrossRef] [Green Version]
- Lin, E.Y.; Jones, J.G.; Li, P.; Zhu, L.; Whitney, K.D.; Muller, W.J.; Pollard, J.W. Progression to malignancy in the polyoma middle T oncoprotein mouse breast cancer model provides a reliable model for human diseases. Am. J. Pathol. 2003, 163, 2113–2126. [Google Scholar] [CrossRef] [Green Version]
- Nagahashi, M.; Yamada, A.; Katsuta, E.; Aoyagi, T.; Huang, W.C.; Terracina, K.P.; Hait, N.C.; Allegood, J.C.; Tsuchida, J.; Yuza, K.; et al. Targeting the SphK1/S1P/S1PR1 Axis That Links Obesity, Chronic Inflammation, and Breast Cancer Metastasis. Cancer Res. 2018, 78, 1713–1725. [Google Scholar] [CrossRef] [Green Version]
- Gordon, R.R.; Hunter, K.W.; La Merrill, M.; Sorensen, P.; Threadgill, D.W.; Pomp, D. Genotype X diet interactions in mice predisposed to mammary cancer: II. Tumors and metastasis. Mamm Genome 2008, 19, 179–189. [Google Scholar] [CrossRef]
- Bousquenaud, M.; Fico, F.; Solinas, G.; Ruegg, C.; Santamaria-Martinez, A. Obesity promotes the expansion of metastasis-initiating cells in breast cancer. Breast Cancer Res. 2018, 20, 104. [Google Scholar] [CrossRef]
- Asiedu, M.K.; Ingle, J.N.; Behrens, M.D.; Radisky, D.C.; Knutson, K.L. TGFbeta/TNF(alpha)-mediated epithelial-mesenchymal transition generates breast cancer stem cells with a claudin-low phenotype. Cancer Res. 2011, 71, 4707–4719. [Google Scholar] [CrossRef] [Green Version]
- Li, C.W.; Xia, W.; Huo, L.; Lim, S.O.; Wu, Y.; Hsu, J.L.; Chao, C.H.; Yamaguchi, H.; Yang, N.K.; Ding, Q.; et al. Epithelial-mesenchymal transition induced by TNF-alpha requires NF-kappaB-mediated transcriptional upregulation of Twist1. Cancer Res. 2012, 72, 1290–1300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia-Tunon, I.; Ricote, M.; Ruiz, A.; Fraile, B.; Paniagua, R.; Royuela, M. Role of tumor necrosis factor-alpha and its receptors in human benign breast lesions and tumors (in situ and infiltrative). Cancer Sci. 2006, 97, 1044–1049. [Google Scholar] [CrossRef] [PubMed]
- Cohen, E.N.; Gao, H.; Anfossi, S.; Mego, M.; Reddy, N.G.; Debeb, B.; Giordano, A.; Tin, S.; Wu, Q.; Garza, R.J.; et al. Inflammation Mediated Metastasis: Immune Induced Epithelial-To-Mesenchymal Transition in Inflammatory Breast Cancer Cells. PLoS ONE 2015, 10, e0132710. [Google Scholar] [CrossRef]
- Monk, J.M.; Liddle, D.M.; Hutchinson, A.L.; Burns, J.L.; Wellings, H.; Cartwright, N.M.; Muller, W.J.; Power, K.A.; Robinson, L.E.; Ma, D.W.L. Fish oil supplementation increases expression of mammary tumor apoptosis mediators and reduces inflammation in an obesity-associated HER-2 breast cancer model. J. Nutr. Biochem. 2021, 95, 108763. [Google Scholar] [CrossRef] [PubMed]
- Leslie, K.; Gao, S.P.; Berishaj, M.; Podsypanina, K.; Ho, H.; Ivashkiv, L.; Bromberg, J. Differential interleukin-6/Stat3 signaling as a function of cellular context mediates Ras-induced transformation. Breast Cancer Res. 2010, 12, R80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeNardo, D.G.; Coussens, L.M. Inflammation and breast cancer. Balancing immune response: Crosstalk between adaptive and innate immune cells during breast cancer progression. Breast Cancer Res. 2007, 9, 212. [Google Scholar] [CrossRef]
- Mei, Y.; Wang, M.; Lu, G.; Li, J.; Peng, L.; Lang, Y.; Yang, M.; Jiang, L.; Li, C.; Zheng, L.; et al. Postponing tumor onset and tumor progression can be achieved by alteration of local tumor immunity. Cancer Cell Int. 2021, 21, 97. [Google Scholar] [CrossRef]
- Lang, P.; Hasselwander, S.; Li, H.; Xia, N. Effects of different diets used in diet-induced obesity models on insulin resistance and vascular dysfunction in C57BL/6 mice. Sci. Rep. 2019, 9, 19556. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rose, D.P.; Vona-Davis, L. The cellular and molecular mechanisms by which insulin influences breast cancer risk and progression. Endocr. Relat. Cancer 2012, 19, R225–R241. [Google Scholar] [CrossRef] [PubMed]
- Del Giudice, M.E.; Fantus, I.G.; Ezzat, S.; McKeown-Eyssen, G.; Page, D.; Goodwin, P.J. Insulin and related factors in premenopausal breast cancer risk. Breast Cancer Res. Treat. 1998, 47, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Buckland, G.; Travier, N.; Cottet, V.; Gonzalez, C.A.; Lujan-Barroso, L.; Agudo, A.; Trichopoulou, A.; Lagiou, P.; Trichopoulos, D.; Peeters, P.H.; et al. Adherence to the mediterranean diet and risk of breast cancer in the European prospective investigation into cancer and nutrition cohort study. Int. J. Cancer. J. Int. Du Cancer 2013, 132, 2918–2927. [Google Scholar] [CrossRef]
- Demetriou, C.A.; Hadjisavvas, A.; Loizidou, M.A.; Loucaides, G.; Neophytou, I.; Sieri, S.; Kakouri, E.; Middleton, N.; Vineis, P.; Kyriacou, K. The mediterranean dietary pattern and breast cancer risk in Greek-Cypriot women: A case-control study. BMC Cancer 2012, 12, 113. [Google Scholar] [CrossRef] [Green Version]
- Wu, A.H.; Yu, M.C.; Tseng, C.C.; Stanczyk, F.Z.; Pike, M.C. Dietary patterns and breast cancer risk in Asian American women. Am. J. Clin. Nutr. 2009, 89, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Murtaugh, M.A.; Sweeney, C.; Giuliano, A.R.; Herrick, J.S.; Hines, L.; Byers, T.; Baumgartner, K.B.; Slattery, M.L. Diet patterns and breast cancer risk in Hispanic and non-Hispanic white women: The Four-Corners Breast Cancer Study. Am. J. Clin. Nutr. 2008, 87, 978–984. [Google Scholar] [CrossRef] [Green Version]
- Trichopoulou, A.; Lagiou, P.; Kuper, H.; Trichopoulos, D. Cancer and Mediterranean dietary traditions. Cancer Epidemiol. Biomark. Prev. 2000, 9, 869–873. [Google Scholar]
- Bessaoud, F.; Daures, J.P.; Gerber, M. Dietary factors and breast cancer risk: A case control study among a population in Southern France. Nutr. Cancer 2008, 60, 177–187. [Google Scholar] [CrossRef]
- Toledo, E.; Salas-Salvado, J.; Donat-Vargas, C.; Buil-Cosiales, P.; Estruch, R.; Ros, E.; Corella, D.; Fito, M.; Hu, F.B.; Aros, F.; et al. Mediterranean Diet and Invasive Breast Cancer Risk Among Women at High Cardiovascular Risk in the PREDIMED Trial: A Randomized Clinical Trial. JAMA Intern. Med. 2015, 175, 1752–1760. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Diet Formula | Control | MFBD | OOBD | |||
|---|---|---|---|---|---|---|
| (g/Kg) | (g/Kg) | (g/Kg) | ||||
| Casein | 195.00 | 195.00 | 195.00 | |||
| DL-Methionine | 3.00 | 3.00 | 3.00 | |||
| Sucrose | 341.00 | 341.46 | 345.20 | |||
| Corn Starch | 211.99 | 150.00 | 147.82 | |||
| Maltodextrin | 100.00 | - | - | |||
| Anhydrous Milkfat | 37.20 | 210.00 | 72.50 | |||
| Olive Oil | - | - | 135.50 | |||
| Soybean Oil | 12.80 | - | - | |||
| Cholesterol | - | 1.50 | 1.85 | |||
| Cellulose | 50.00 | 50.00 | 50.00 | |||
| Mineral Mix, AIN-76 | 35.00 | 35.00 | 35.00 | |||
| Vitamin Mix, Teklad | 10.00 | 10.00 | 10.00 | |||
| Ethioxyquin, antioxidant | 0.01 | 0.04 | 0.04 | |||
| Nutrient Composition | % Weight | % Kcal | % Weight | % Kcal | % Weight | % Kcal |
| Protein | 17.30 | 18.70 | 17.30 | 15.20 | 17.30 | 15.30 |
| Carbohydrate | 63.50 | 68.60 | 48.50 | 42.70 | 48.70 | 43.00 |
| Fat | 5.20 | 12.60 | 21.10 | 42.00 | 21.00 | 41.70 |
| Energy (Kcal/g) | 3.70 | 4.50 | 4.50 | |||
| Diet Formula | g/Kg | % | g/Kg | % | g/Kg | % |
| Total Fat | 58.98 | 216.55 | 223.64 | |||
| SFA | 26.10 | 52.4 | 136.71 | 65.2 | 70.40 | 33.85 |
| MUFA | 14.60 | 29.3 | 65.73 | 31.3 | 120.70 | 70.40 |
| PUFA | 9.10 | 18.3 | 7.35 | 3.5 | 16.90 | 13.95 |
| 4:0 | 1.40 | 2.37 | 7.98 | 3.69 | 2.76 | 1.23 |
| 6:0 | 0.86 | 1.46 | 4.83 | 2.23 | 1.67 | 0.75 |
| 8:0 | 0.41 | 0.70 | 2.31 | 1.07 | 0.80 | 0.36 |
| 10:0 | 0.74 | 1.25 | 4.20 | 1.94 | 1.45 | 0.65 |
| 12:0 | 1.20 | 2.03 | 6.51 | 3.01 | 2.25 | 1.01 |
| 14:0 | 4.40 | 7.46 | 24.57 | 11.35 | 8.48 | 3.79 |
| 14:1 | 0.30 | 0.51 | 1.68 | 0.78 | 0.58 | 0.26 |
| 15:0 | 0.60 | 1.02 | 3.36 | 1.55 | 1.16 | 0.52 |
| 16:0 | 11.20 | 18.99 | 55.02 | 25.41 | 37.60 | 16.81 |
| 16:1 | 0.71 | 1.20 | 3.99 | 1.84 | 3.00 | 1.34 |
| 17:0 | 0.26 | 0.44 | 1.47 | 0.68 | 0.51 | 0.23 |
| 17:1 | - | 0.00 | 0.04 | 0.02 | 0.15 | 0.07 |
| 18:0 | 5.20 | 8.82 | 26.25 | 12.12 | 12.50 | 5.59 |
| 18:1 | 13.50 | 22.89 | 59.22 | 27.35 | 116.80 | 52.23 |
| 18:2 | 7.90 | 13.39 | 6.09 | 2.81 | 15.70 | 7.02 |
| 18:3 | 1.20 | 2.03 | 1.05 | 0.48 | 1.18 | 0.53 |
| 20:1 | - | 0.00 | 0.42 | 0.19 | 0.15 | 0.07 |
| 20:4 | - | 0.00 | 0.21 | 0.10 | 0.00 | 0.00 |
| n-6 | 1.20 | 2.03 | 6.30 | 2.91 | 15.70 | 7.02 |
| n-3 | 7.90 | 13.39 | 1.05 | 0.48 | 1.20 | 0.54 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Velazquez, F.N.; Viscardi, V.; Montemage, J.; Zhang, L.; Trocchia, C.; Delamont, M.M.; Ahmad, R.; Hannun, Y.A.; Obeid, L.M.; Snider, A.J. A Milk-Fat Based Diet Increases Metastasis in the MMTV-PyMT Mouse Model of Breast Cancer. Nutrients 2021, 13, 2431. https://doi.org/10.3390/nu13072431
Velazquez FN, Viscardi V, Montemage J, Zhang L, Trocchia C, Delamont MM, Ahmad R, Hannun YA, Obeid LM, Snider AJ. A Milk-Fat Based Diet Increases Metastasis in the MMTV-PyMT Mouse Model of Breast Cancer. Nutrients. 2021; 13(7):2431. https://doi.org/10.3390/nu13072431
Chicago/Turabian StyleVelazquez, Fabiola N., Valentina Viscardi, Julia Montemage, Leiqing Zhang, Carolena Trocchia, Megan M. Delamont, Rasheed Ahmad, Yusuf A. Hannun, Lina M. Obeid, and Ashley J. Snider. 2021. "A Milk-Fat Based Diet Increases Metastasis in the MMTV-PyMT Mouse Model of Breast Cancer" Nutrients 13, no. 7: 2431. https://doi.org/10.3390/nu13072431