A Standardized Extract of Asparagus officinalis Stem (ETAS®) Ameliorates Cognitive Impairment, Inhibits Amyloid β Deposition via BACE-1 and Normalizes Circadian Rhythm Signaling via MT1 and MT2


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Diet
2.2. Cognitive Evaluation
2.3. Measurements of Amyloid β (Aβ)
2.4. Morphometric Analysis of the Suprachiasmatic Nucleus (SCN)
2.5. Western Blotting
2.6. Statistics
3. Results
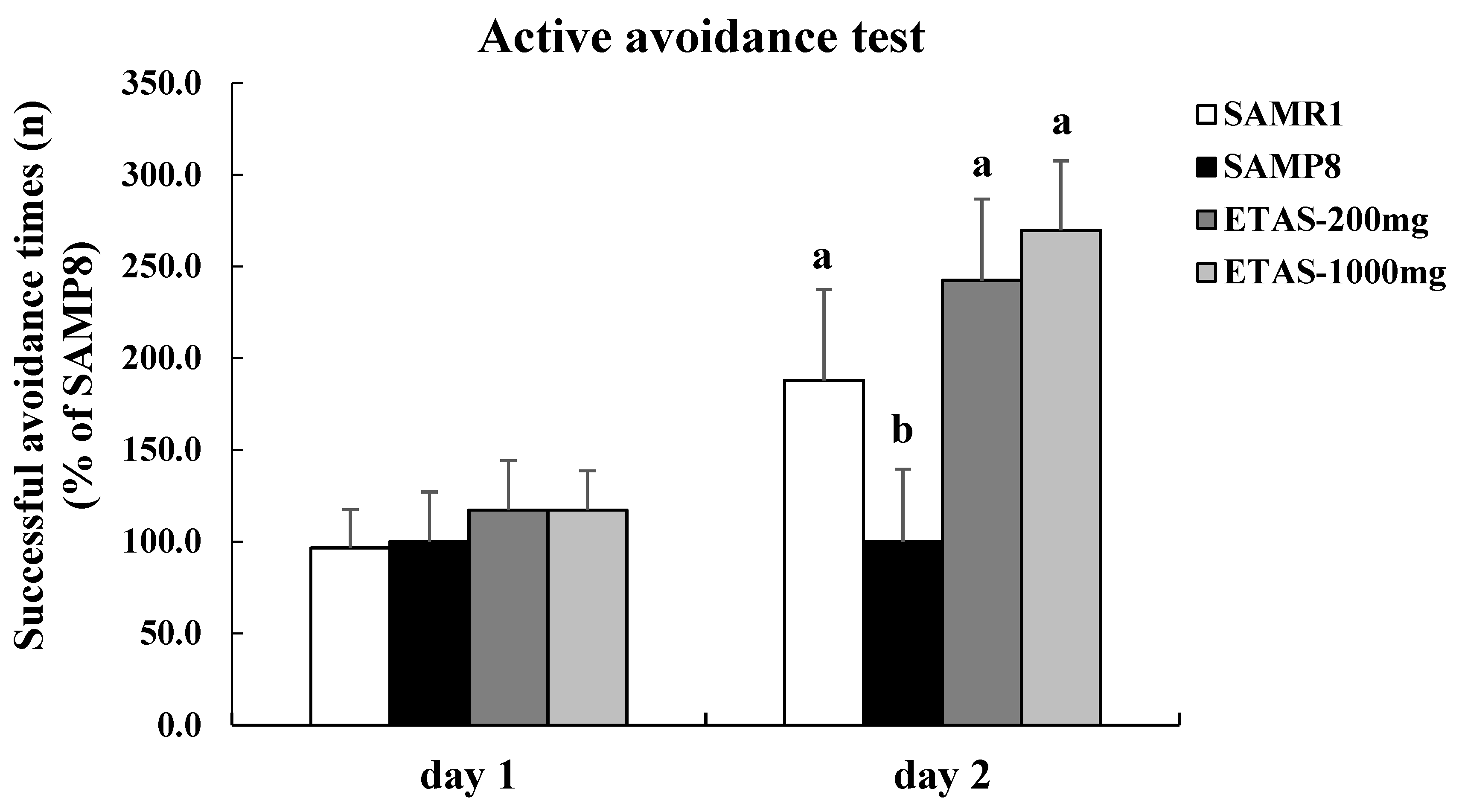
3.1. ETAS® Improved Cognitive Function
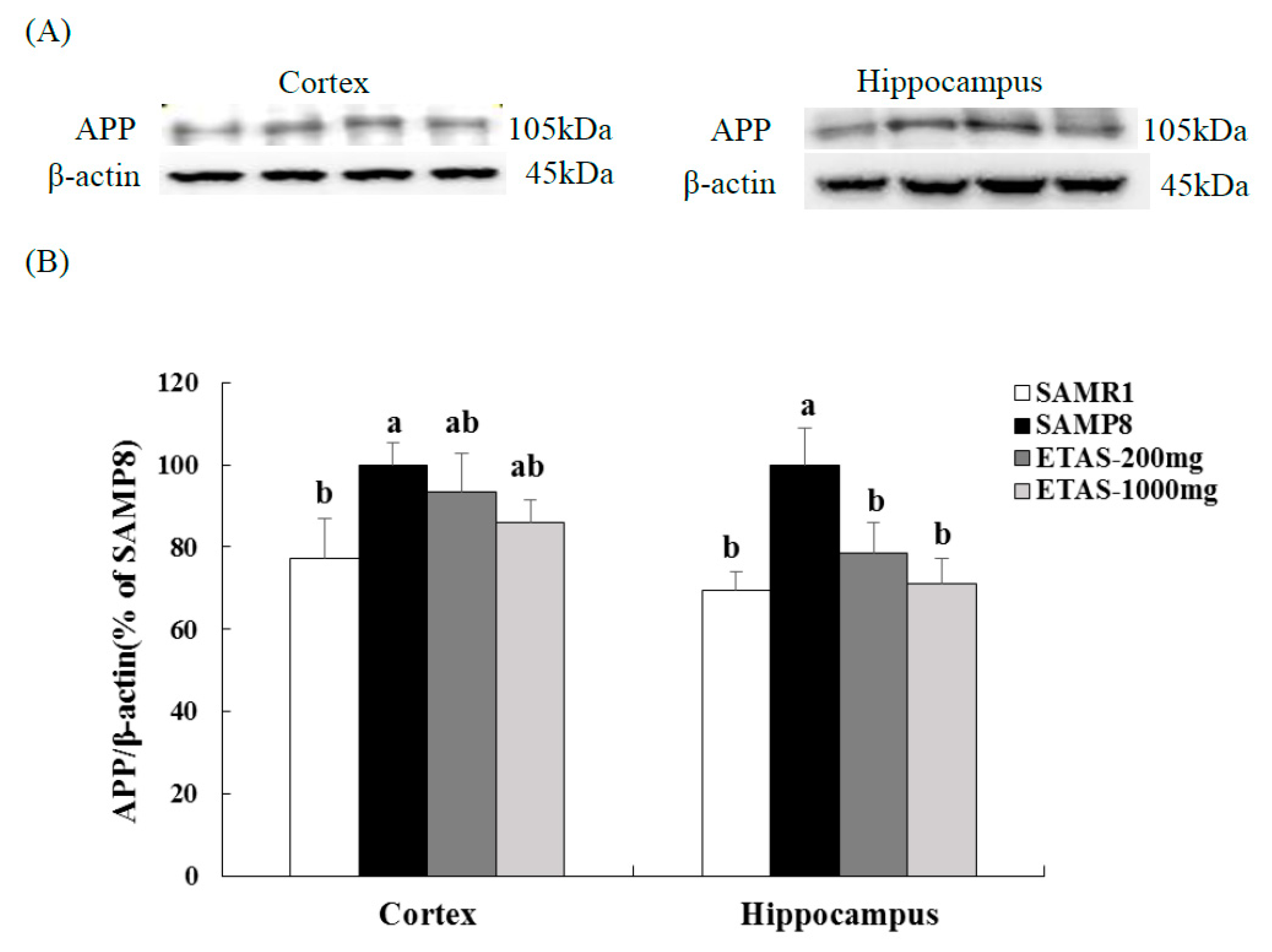
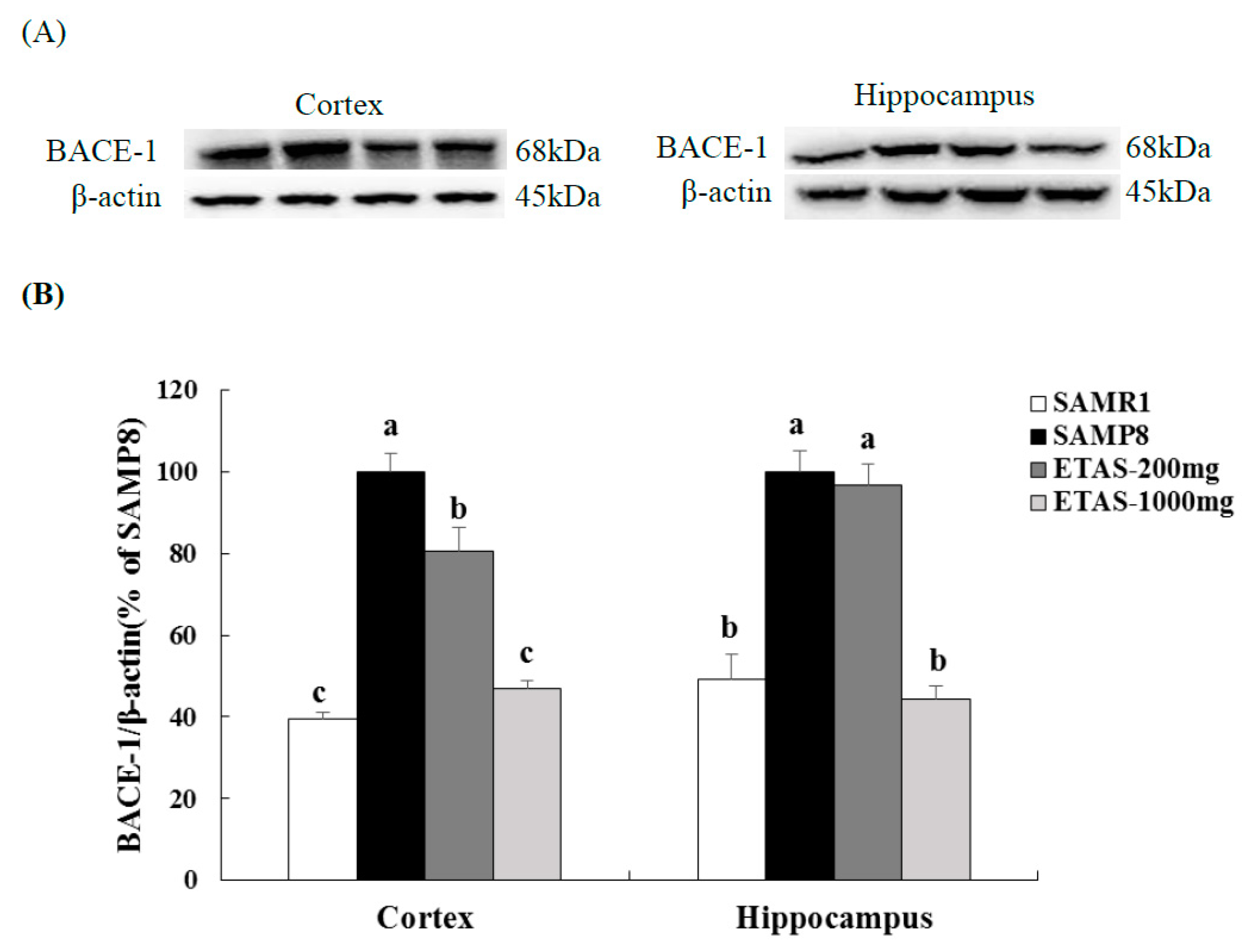
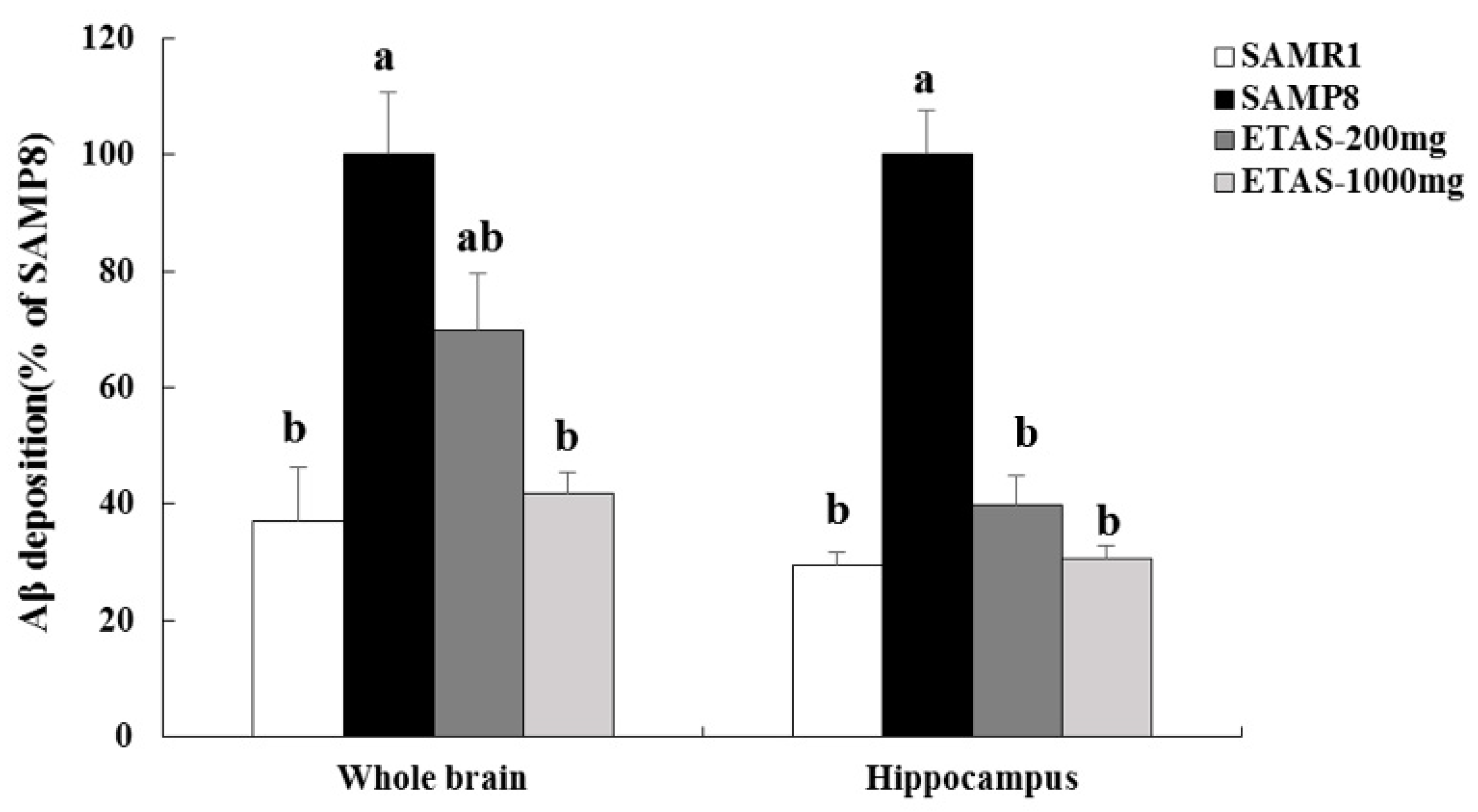
3.2. ETAS® Reduced the Expressions of APP, BACE-1 and Aβ Deposition
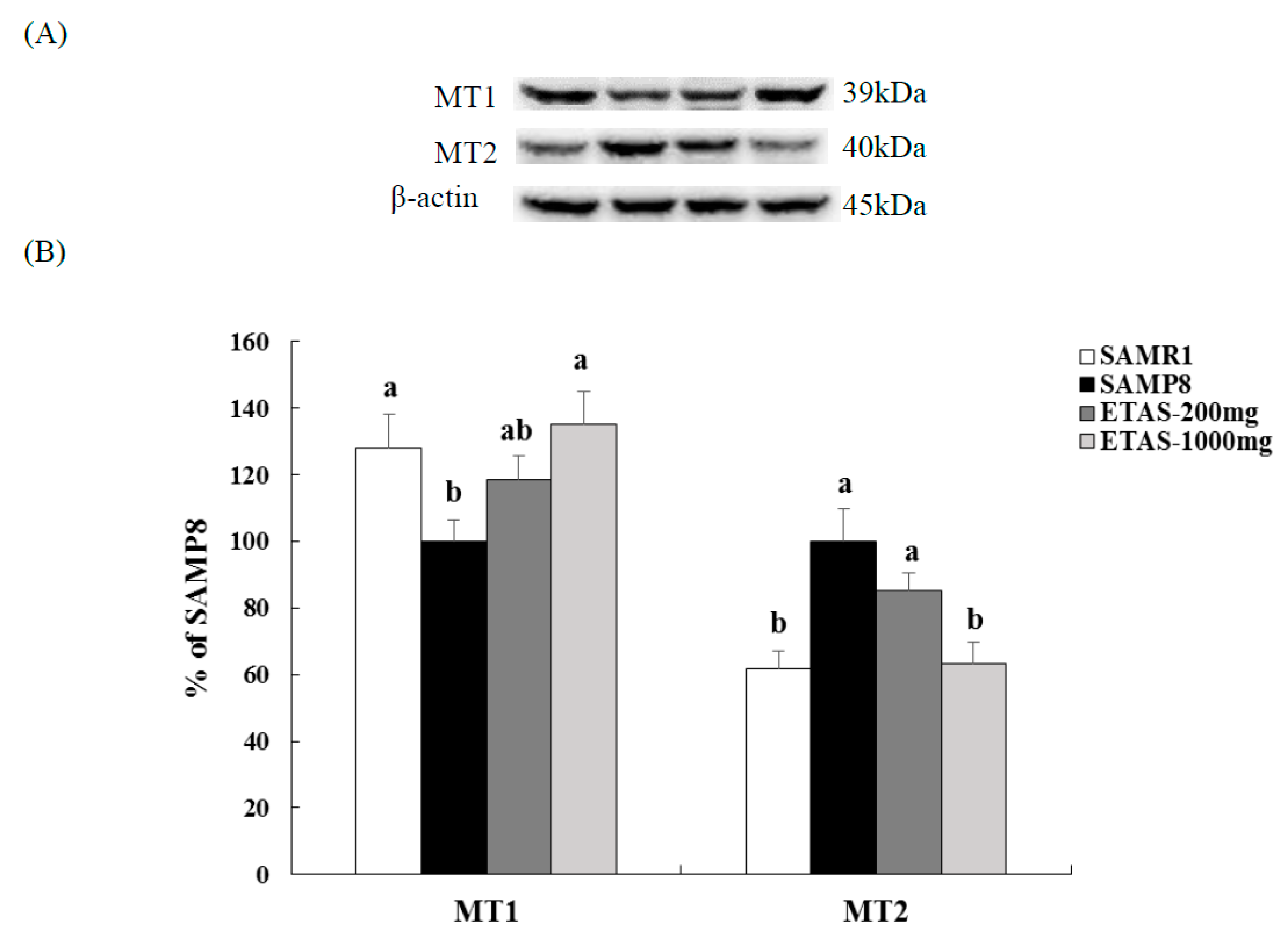
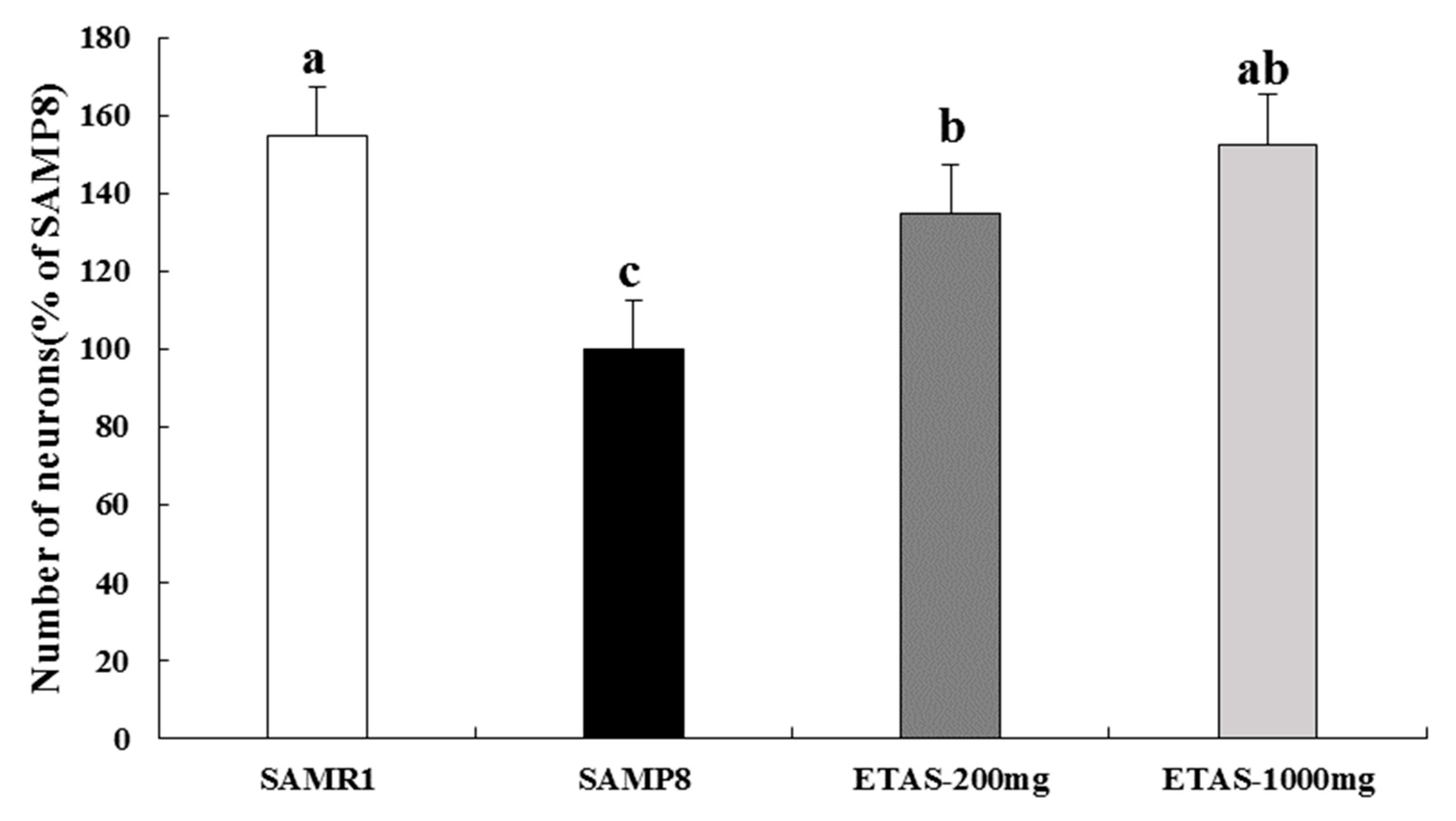
3.3. ETAS® Increased SCN Neuron and Improved the Circadian Rhythm Modulators
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Green, K.N.; Smith, I.F.; Laferla, F.M. Role of calcium in the pathogenesis of Alzheimer’s disease and transgenic models. Subcell. Biochem. 2007, 45, 507–521. [Google Scholar] [PubMed]
- Makhaeva, G.F.; Lushchekina, S.V.; Boltneva, N.P.; Sokolov, V.B.; Grigoriev, V.V.; Serebryakova, O.G.; Vikhareva, E.A.; Aksinenko, A.Y.; Barreto, G.E.; Aliev, G.; et al. Conjugates of γ-Carbolines and Phenothiazine as new selective inhibitors of butyrylcholinesterase and blockers of NMDA receptors for Alzheimer Diseasee. Sci. Rep. 2015, 5, 13164. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.H.; Bastianetto, S.; Mennicken, F.; Ma, W.; Kar, S. Amyloid beta peptide induces tau phosphorylation and loss of cholinergic neurons in rat primary septal cultures. Neuroscience 2002, 115, 201–211. [Google Scholar] [CrossRef]
- Turner, P.R.; O’Connor, K.; Tate, W.P.; Abraham, W.C. Roles of amyloid precursor protein and its fragments in regulating neural activity, plasticity and memory. Prog. Neurobiol. 2003, 70, 1–32. [Google Scholar] [CrossRef]
- Sanabria-Castro, A.; Alvarado-Echeverría, I.; Monge-Bonilla, C. Molecular pathogenesis of Alzheimer’s disease: An update. Ann. Neurosci. 2017, 24, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Song, W. The role of APP and BACE1 trafficking in APP processing and amyloid-β generation. Alzheimers Res. Ther. 2013, 5, 46. [Google Scholar] [CrossRef] [PubMed]
- Van Erum, J.; Van Dam, D.; De Deyn, P.P. Sleep and Alzheimer’s disease: A pivotal role for the suprachiasmatic nucleus. Sleep Med. Rev. 2018, 40, 17–27. [Google Scholar] [CrossRef]
- Wu, Y.H.; Zhou, J.N.; Van Heerikhuize, J.; Jockers, R.; Swaab, D.F. Decreased MT1 melatonin receptor expression in the suprachiasmatic nucleus in aging and Alzheimer’s disease. Neurobiol. Aging. 2007, 28, 1239–1247. [Google Scholar] [CrossRef]
- Dubocovich, M.L.; Rivera-Bermudez, M.A.; Gerdin, M.J.; Masana, M.I. Molecular pharmacology, regulation and function of mammalian melatonin receptors. Front. Biosci. 2003, 8, d1093–d1108. [Google Scholar] [CrossRef]
- Reppert, S.M.; Weaver, D.R.; Ebisawa, T. Cloning and characterization of a mammalian melatonin receptor that mediates reproductive and circadian responses. Neuron 1994, 13, 1177–1185. [Google Scholar] [CrossRef]
- Reppert, S.M.; Godson, C.; Mahle, C.D.; Weaver, D.R.; Slaugenhaupt, S.A.; Gusella, J.F. Molecular characterization of a second melatonin receptor expressed in human retina and brain: The Mel1b melatonin receptor. Proc. Natl. Acad. Sci. USA 1995, 92, 8734–8738. [Google Scholar] [CrossRef] [PubMed]
- Sun, T.; Powers, J.R.; Tang, J. Enzyme-catalyzed change of antioxidants content and antioxidant activity of Asparagus juice. J. Agric. Food Chem. 2007, 55, 56–60. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Liu, Y.; Zhao, J.; Zhang, W.; Pang, X. Saponins extracted from by-product of Asparagus officinalis L. suppress tumour cell migration and invasion through targeting Rho GTPase signalling pathway. J. Sci. Food Agric. 2013, 93, 1492–1498. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Zhang, W.; Zhu, X.; Zhao, D.; Wang, K.; Wang, R.; Qu, W. The aqueous extract of Asparagus officinalis L. by-product exerts hypoglycaemic activity in streptozotocin-induced diabetic rats. J. Sci. Food Agric. 2011, 91, 2095–2099. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Zhang, W.; Zhao, J.; Wang, J.; Qu, W. Hypolipidaemic and hepatoprotective effects of ethanolic and aqueous extracts from Asparagus officinalis L. by-products in mice fed a high-fat diet. J. Sci. Food Agric. 2010, 90, 1129–1135. [Google Scholar] [CrossRef] [PubMed]
- Sakurai, T.; Ito, T.; Wakame, K.; Kitadate, K.; Arai, T.; Ogasawara, J.; Kizaki, T.; Sato, S.; Ishibashi, Y.; Fujiwara, T.; et al. Enzyme-treated Asparagus officinalis extract shows neuroprotective effects and attenuates cognitive impairment in senescence-accelerated mice. Nat. Prod. Commun. 2014, 9, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Hwang, J.H.; Lee, I.T.; Jeng, K.C.; Wang, M.F.; Hou, R.C.; Wu, S.M.; Chan, Y.C. Spirulina prevents memory dysfunction, reduces oxidative stress damage and augments antioxidant activity in senescence-accelerated mice. J. Nutr. Sci. Vitaminol. Tokyo 2011, 57, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Engelberth, R.C.; Silva, K.D.; Azevedo, C.V.; Gavioli, E.C.; dos Santos, J.R.; Soares, J.G.; Nascimento Junior, E.S.; Cavalcante, J.C.; Costa, M.S.; Cavalcante, J.S. Morphological changes in the suprachiasmatic nucleus of aging female marmosets (Callithrix jacchus). Biomed Res. Int. 2014, 2014, 243825. [Google Scholar] [CrossRef] [PubMed]
- Chan, Y.C.; Lee, I.T.; Wang, M.F.; Yeh, W.C.; Liang, B.C. Tempeh attenuates cognitive deficit, antioxidant imbalance, and amyloid β of senescence-accelerated mice by modulating Nrf2 expression via MAPK pathway. J. Funct. Foods 2018, 50, 112–119. [Google Scholar] [CrossRef]
- Takeda, T.; Hosokawa, M.; Higuchi, K. Senescence-accelerated mouse (SAM): A novel murine model of accelerated senescence. J. Am. Geriatr. Soc. 1991, 39, 911–919. [Google Scholar] [CrossRef]
- Flood, J.F.; Morley, J.E. Age-related changes in footshock avoidance acquisition and retention in senescence accelerated mouse (SAM). Neurobiol. Aging 1993, 14, 153–157. [Google Scholar] [CrossRef]
- Flood, J.F.; Morley, J.E. Learning and memory in SAMP8 mouse. Neurosci. Biobehav Rev. 1998, 72, 1–20. [Google Scholar] [CrossRef]
- Miyamoto, M. Characteristics of age-related behavioral changes in senescence accelerated mouse SAMP8 and SAMP10. Exp. Gerontol. 1997, 32, 139–148. [Google Scholar] [CrossRef]
- Ohta, A.; Hirano, T.; Yagi, H.; Tanaka, S.; Hosokawa, M.; Takeda, T. Behavioral characteristics of the SAMP8 strain in Sidman active avoidance task. Brain Res. 1989, 498, 195–198. [Google Scholar] [CrossRef]
- McAuley, J.D.; Miller, J.P.; Beck, E.; Nagy, Z.M.; Pang, K.C. Age-related disruptions in circadian timing: Evidence for “split” activity rhythms in the SAMP8. Neurobiol. Aging 2002, 23, 625–632. [Google Scholar] [CrossRef]
- Satlin, A.; Teicher, M.H.; Lieberman, H.R.; Baldessarini, R.J.; Volicer, L.; Rheaume, Y. Circadian locomotor activity rhythms in Alzheimer’s disease. Neuropsychopharmacology 1991, 5, 115–126. [Google Scholar]
- Van Gool, W.A.; Mirmiran, M. Aging and circadian rhythms. Prog. Brain Res. 1986, 70, 255–277. [Google Scholar]
- Wu, Y.H.; Swaab, D.F. The human pineal gland and melatonin in aging and Alzheimer’s disease. J. Pineal Res. 2005, 38, 145–152. [Google Scholar] [CrossRef]
- Schwartz, W.; Busis, N.; Tessa Hedley-Whyte, E. A discrete lesion of ventral hypothalamus and optic chiasm that disturbed the daily temperature rhythm. J. Neurol. 1986, 233, 1–4. [Google Scholar] [CrossRef]
- Swaab, D.F.; Fliers, E.; Partiman, T.S. The suprachiasmatic nucleus of the human brain in relation to sex, age and senile dementia. Brain Res. 1985, 342, 37–44. [Google Scholar] [CrossRef]
- Zhou, J.N.; Liu, R.Y.; van Heerikhuize, J.; Hofman, M.A.; Swaab, D.F. Alterations in the circadian rhythm of salivary melatonin begin during middle-age. J. Pineal Res. 2003, 34, 11–16. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.Y.; Zhou, J.N.; van Heerikhuize, J.; Hofman, M.A.; Swaab, D.F. Decreased melatonin levels in postmortem cerebrospinal fluid in relation to aging, Alzheimer’s disease, and apolipoprotein E-ε4/4 genotype. J. Clin. Endocrinol. Metabol. 1999, 84, 323–327. [Google Scholar]
- Hunt, A.E.; Al-Ghoul, W.M.; Gillette, M.U.; Dubocovich, M.L. Activation of MT(2) melatonin receptors in rat suprachiasmatic nucleus phase advances the circadian clock. Am. J. Physiol. Cell Physiol. 2001, 280, C110–C118. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Ni, C.; Li, Z.; Yang, N.; Zhou, Y.; Rong, X.; Qian, M.; Chui, D.; Guo, X. Prophylactic melatonin attenuates isoflurane-induced cognitive impairment in aged rats through hippocampal melatonin receptor 2-cAMP response element binding signalling. Basic Clin. Pharmacol. Toxicol. 2017, 120, 219–226. [Google Scholar] [CrossRef]
- Asai, M.; Ikeda, M.; Akiyama, M.; Oshima, I.; Shibata, S. Administration of melatonin in drinking water promotes the phase advance of light-dark cycle in senescence-accelerated mice, SAMR1 but not SAMP8. Brain Res. 2000, 876, 220–224. [Google Scholar] [CrossRef]
- Caballero, B.; Vega-Naredo, I.; Sierra, V.; Huidobro-Fernández, C.; Soria-Valles, C.; De Gonzalo-Calvo, D.; Tolivia, D.; Gutierrez-Cuesta, J.; Pallas, M.; Camins, A.; et al. Favorable effects of a prolonged treatment with melatonin on the level of oxidative damage and neurodegeneration in senescence-accelerated mice. J. Pineal Res. 2008, 45, 302–311. [Google Scholar] [CrossRef]
- Selkoe, D.J. Translating cell biology into therapeutic advances in Alzheimer’s disease. Nature 1999, 399, A23–A31. [Google Scholar] [CrossRef]
- Cai, H.; Wang, Y.; McCarthy, D.; Wen, H.; Borchelt, D.R.; Price, D.L.; Wong, P.C. BACE1 is the major beta-secretase for generation of Abeta peptides by neurons. Nat. Neurosci. 2001, 4, 233–234. [Google Scholar] [CrossRef]
- Laird, F.M.; Cai, H.; Savonenko, A.V.; Farah, M.H.; He, K.; Melnikova, T.; Wen, H.; Chiang, H.C.; Xu, G.; Koliatsos, V.E.; et al. BACE1, a major determinant of selective vulnerability of the brain to amyloid-beta amyloidogenesis, is essential for cognitive, emotional, and synaptic functions. J. Neurosci. 2005, 25, 11693–11709. [Google Scholar] [CrossRef]
- Tamagno, E.; Guglielmotto, M.; Aragno, M.; Borghi, R.; Autelli, R.; Giliberto, L.; Muraca, G.; Danni, O.; Zhu, X.; Smith, M.A.; et al. Oxidative stress activates a positive feedback between the gamma- and beta-secretase cleavages of the beta-amyloid precursor protein. J. Neurochem. 2008, 104, 683–695. [Google Scholar]
- Kumar, V.B.; Farr, S.A.; Flood, J.F.; Kamlesh, V.; Franko, M.; Banks, W.A.; Morley, J.E. Site-directed antisense oligonucleotide decreases the expression of amyloid precursor protein and reverses deficits in learning and memory in aged SAMP8 mice. Peptides 2000, 21, 1769–1775. [Google Scholar] [CrossRef]
- Che, H.; Zhou, M.; Zhang, T.; Zhang, L.; Ding, L.; Yanagita, T.; Xu, J.; Xue, C.; Wang, Y. Comparative study of the effects of phosphatidylcholine rich in DHA and EPA on Alzheimer’s disease and the possible mechanisms in CHO-APP/PS1 cells and SAMP8 mice. Food Funct. 2018, 9, 643–654. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Wu, D.; Wang, J.; Li, F.; Lu, L.; Gao, Y.; Zhong, Z. Effects of Panax notoginseng saponin on α, β, and γ secretase involved in Aβ deposition in SAMP8 mice. Neuroreport 2014, 25, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Yang, J.; Liu, C.; Xie, J.; Qiu, S.; Yang, X.; Wu, C. Pseudoginsenoside-F11 alleviates cognitive deficits and Alzheimer’s disease-type pathologies in SAMP8 mice. Pharmacol. Res. 2019, 139, 512–523. [Google Scholar] [CrossRef] [PubMed]
- Makris, D.P.; Rossiter, J.T. Domestic processing of onion bulbs (Allium cepa) and asparagus spears (Asparagus officinalis), effect on flavonol content and antioxidant status. J. Agric. Food Chem. 2001, 49, 3216–3222. [Google Scholar] [CrossRef] [PubMed]
- Deli, J.; Matus, Z.; Tóth, G. Carotenoid composition in the fruits of Asparagus officinalis. J. Agric. Food Chem. 2000, 48, 2793–2796. [Google Scholar] [CrossRef] [PubMed]
- Ishola, I.O.; Jacinta, A.A.; Adeyemi, O.O. Cortico-hippocampal memory enhancing activity of hesperetin on scopolamine-induced amnesia in mice: Role of antioxidant defense system, cholinergic neurotransmission and expression of BDNF. Metab. Brain Dis. 2019, 34, 979–989. [Google Scholar] [CrossRef] [PubMed]
- Zaplatic, E.; Bule, M.; Shah, S.Z.A.; Uddin, M.S.; Niaz, K. Molecular mechanisms underlying protective role of quercetin in attenuating Alzheimer’s disease. Life Sci. 2019, 224, 109–119. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.; Gan, D.; Fan, C.; Wen, C.; Li, A.; Li, Q.; Zhao, J.; Wang, Z.; Zhu, L.; Lu, D. The secretion from neural stem cells pretreated with lycopene protects against tert-butyl hydroperoxide-induced neuron oxidative damage. Oxid. Med. Cell. Longev. 2018, 2018, 5490218. [Google Scholar] [CrossRef]
- Lakey-Beitia, J.; Doens, D.; Jagadeesh Kumar, D.; Murillo, E.; Fernandez, P.L.; Rao, K.S.; Durant-Archibold, A.A. Anti-amyloid aggregation activity of novel carotenoids: Implications for Alzheimer’s drug discovery. Clin. Interv. Aging 2017, 12, 815–822. [Google Scholar] [CrossRef]
- Ito, T.; Maeda, T.; Goto, K.; Miura, T.; Wakame, K.; Nishioka, H.; Sato, A. Enzyme-treated asparagus extract promotes expression of heat shock protein and exerts antistress effects. J. Food Sci. 2014, 79, H413–H419. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Goto, K.; Takanari, J.; Miura, T.; Wakame, K.; Ishioka, H.; Tanaka, A.; Nishihira, J. Effects of enzyme treated asparagus extract on heat shock protein 70, stress indices, and sleep in healthy adult men. J. Nutr. Sci. Vitaminol. 2014, 60, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Takanari, J.; Nakahigashi, J.; Sato, A.; Waki, H.; Miyazaki, S.; Uebaba, K.; Hisajima, T. Effect of enzyme-treated asparagus extract (ETAS) on psychological stress in healthy individuals. J. Nutr. Sci. Vitaminol. Tokyo 2016, 62, 198–205. [Google Scholar] [CrossRef] [PubMed]
- Nishizawa, M.; Kano, M.; Okuyama, T.; Okumura, T.; Ikeya, Y. Anti-inflammatory effects of enzyme-treated asparagus extract and its constituents in hepatocytes. FFHD 2016, 6, 91–109. [Google Scholar]
- Shirato, K.; Takanari, J.; Sakurai, T.; Ogasawara, J.; Imaizumi, K.; Ohno, H. Enzyme-treated asparagus extract prevents’ hydrogen peroxide-induced pro-Inflammatory responses by suppressing p65 nuclear translocation in skin L929 fibroblasts. Nat. Prod. Commun. 2016, 11, 1883–1888. [Google Scholar] [CrossRef] [PubMed]
- Heneka, M.T.; Carson, M.J.; EI Khoury, J.; Landreth, G.E.; Brosseron, F.; Feinstein, D.L.; Jacobs, A.H.; Wyss-Coray, T.; Vitorica, J.; Ransohoff, R.M.; et al. Neuroinflammation in Alzheimer’s disease. Lancet Neurol. 2015, 14, 388–405. [Google Scholar] [CrossRef]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: The role of inflammation in Alzheimer disease. Nat. Rev. Neurosci. 2015, 16, 358–372. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chan, Y.-C.; Wu, C.-S.; Wu, T.-C.; Lin, Y.-H.; Chang, S.-J. A Standardized Extract of Asparagus officinalis Stem (ETAS®) Ameliorates Cognitive Impairment, Inhibits Amyloid β Deposition via BACE-1 and Normalizes Circadian Rhythm Signaling via MT1 and MT2. Nutrients 2019, 11, 1631. https://doi.org/10.3390/nu11071631
Chan Y-C, Wu C-S, Wu T-C, Lin Y-H, Chang S-J. A Standardized Extract of Asparagus officinalis Stem (ETAS®) Ameliorates Cognitive Impairment, Inhibits Amyloid β Deposition via BACE-1 and Normalizes Circadian Rhythm Signaling via MT1 and MT2. Nutrients. 2019; 11(7):1631. https://doi.org/10.3390/nu11071631
Chicago/Turabian StyleChan, Yin-Ching, Ci-Sian Wu, Tsai-Chen Wu, Yu-Hsuan Lin, and Sue-Joan Chang. 2019. "A Standardized Extract of Asparagus officinalis Stem (ETAS®) Ameliorates Cognitive Impairment, Inhibits Amyloid β Deposition via BACE-1 and Normalizes Circadian Rhythm Signaling via MT1 and MT2" Nutrients 11, no. 7: 1631. https://doi.org/10.3390/nu11071631