
Oilseed Rape Cultivars Show Diversity of Root Morphologies with the Potential for Better Capture of Nitrogen

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. In Vitro Culture and 2D Root Morphology Analysis
2.3. Hydroponic Culture, Physiological Characterization, and Nitrate Uptake Assays
2.4. Soil Culture and Micro-Scale Computed Tomography Imaging
2.5. Statistical Treatment
3. Results
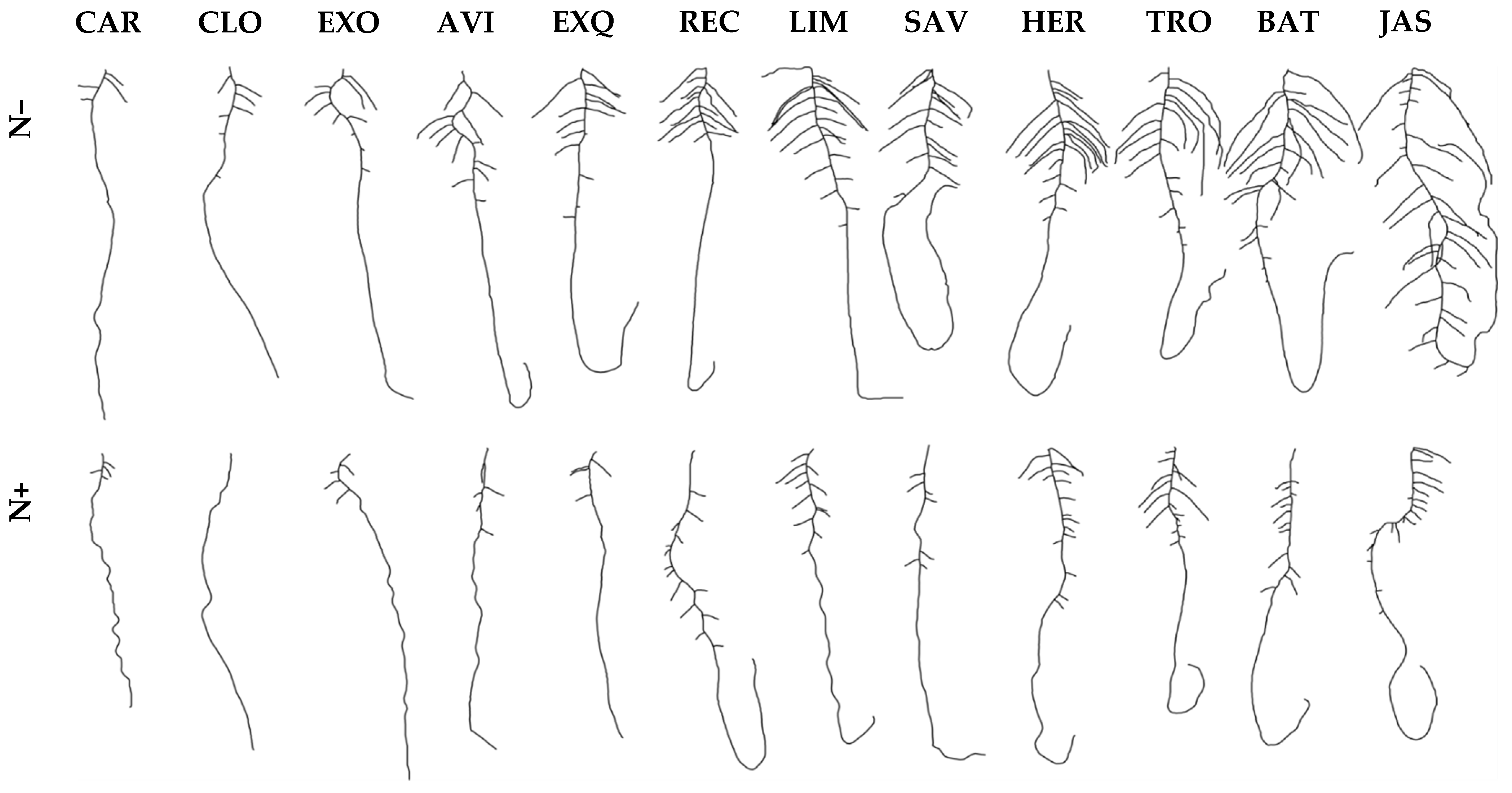
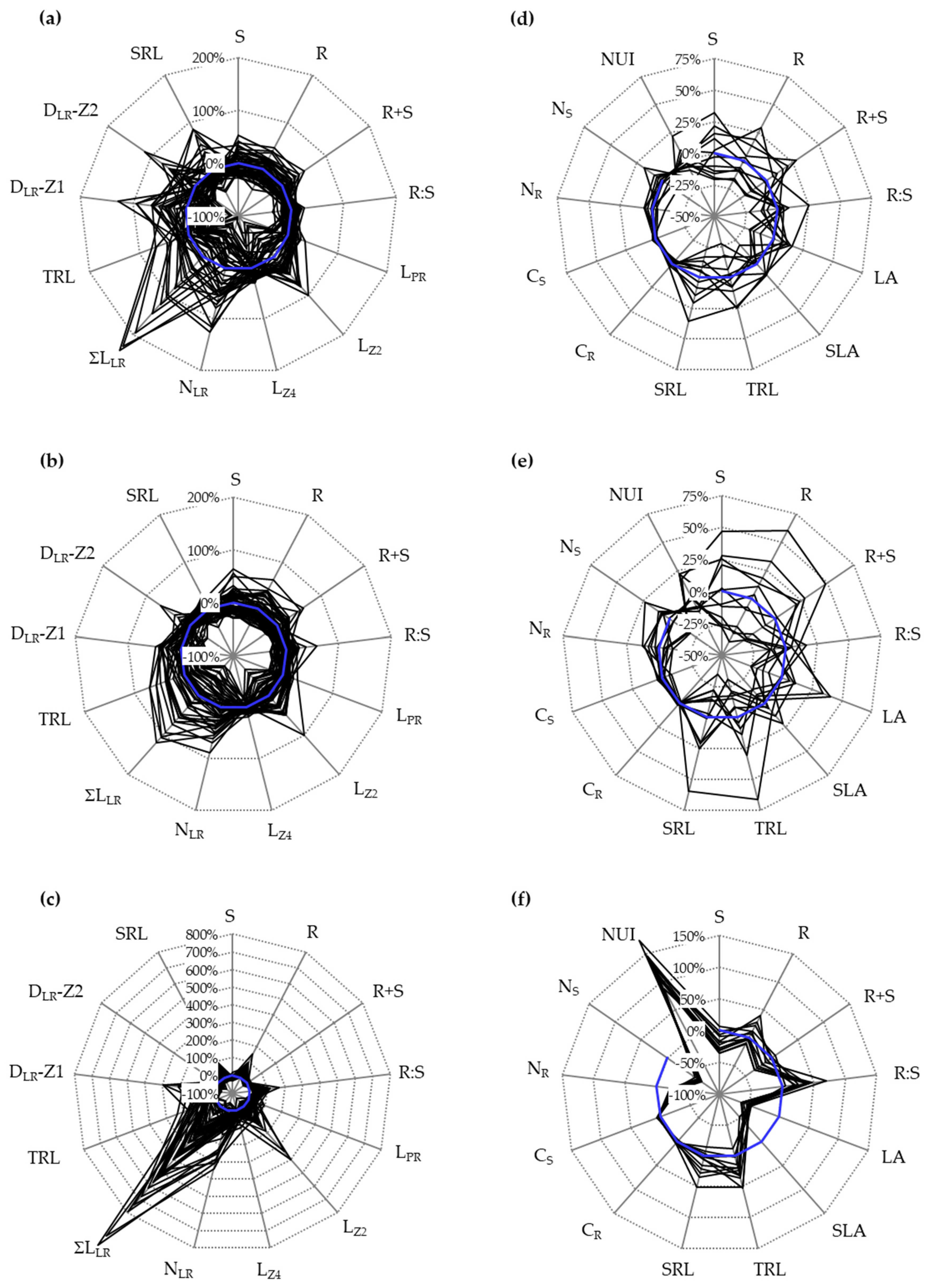
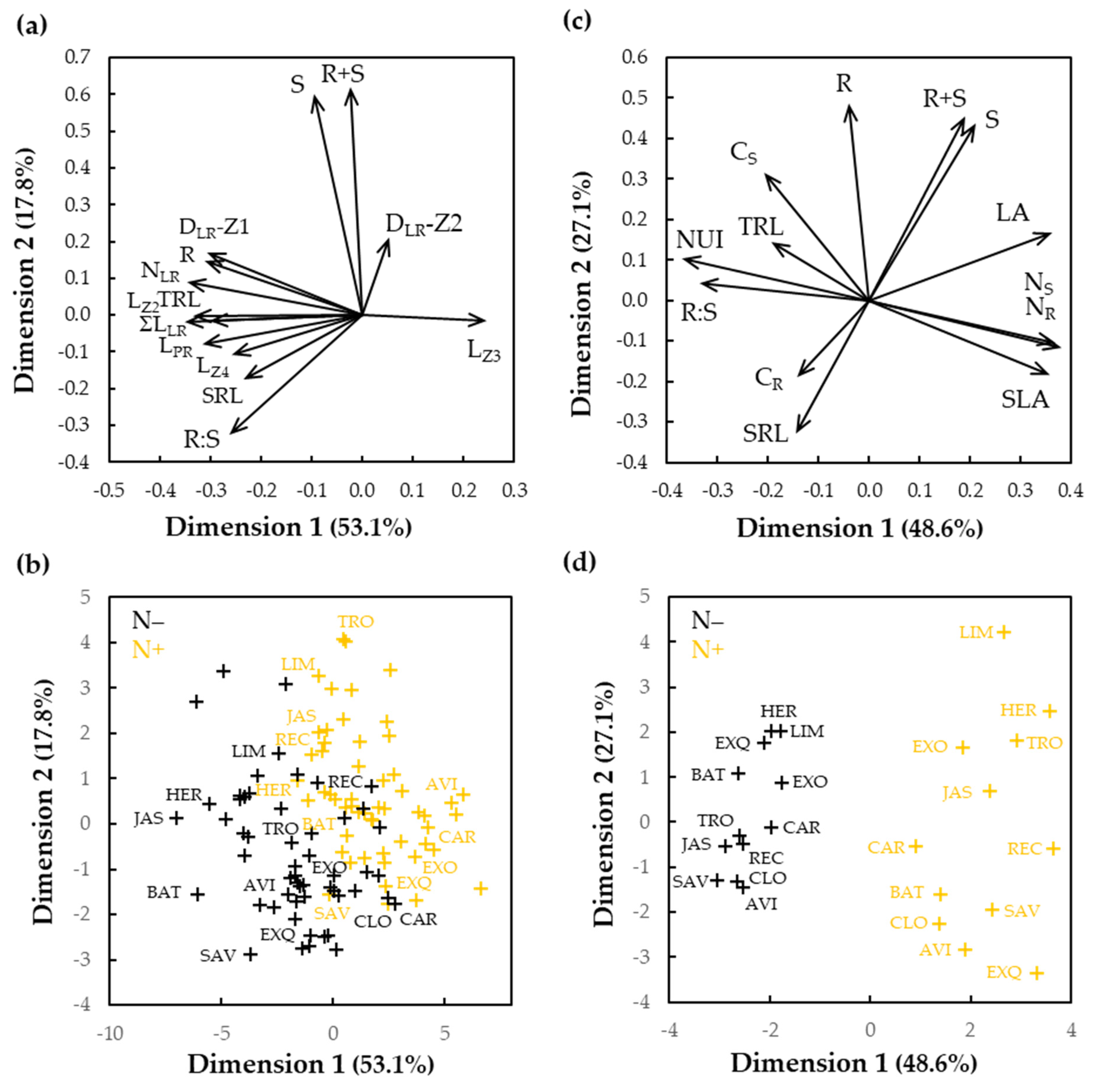
3.1. Variation for Biomass Production and Root Morphologies in Seedlings Cultivated In Vitro
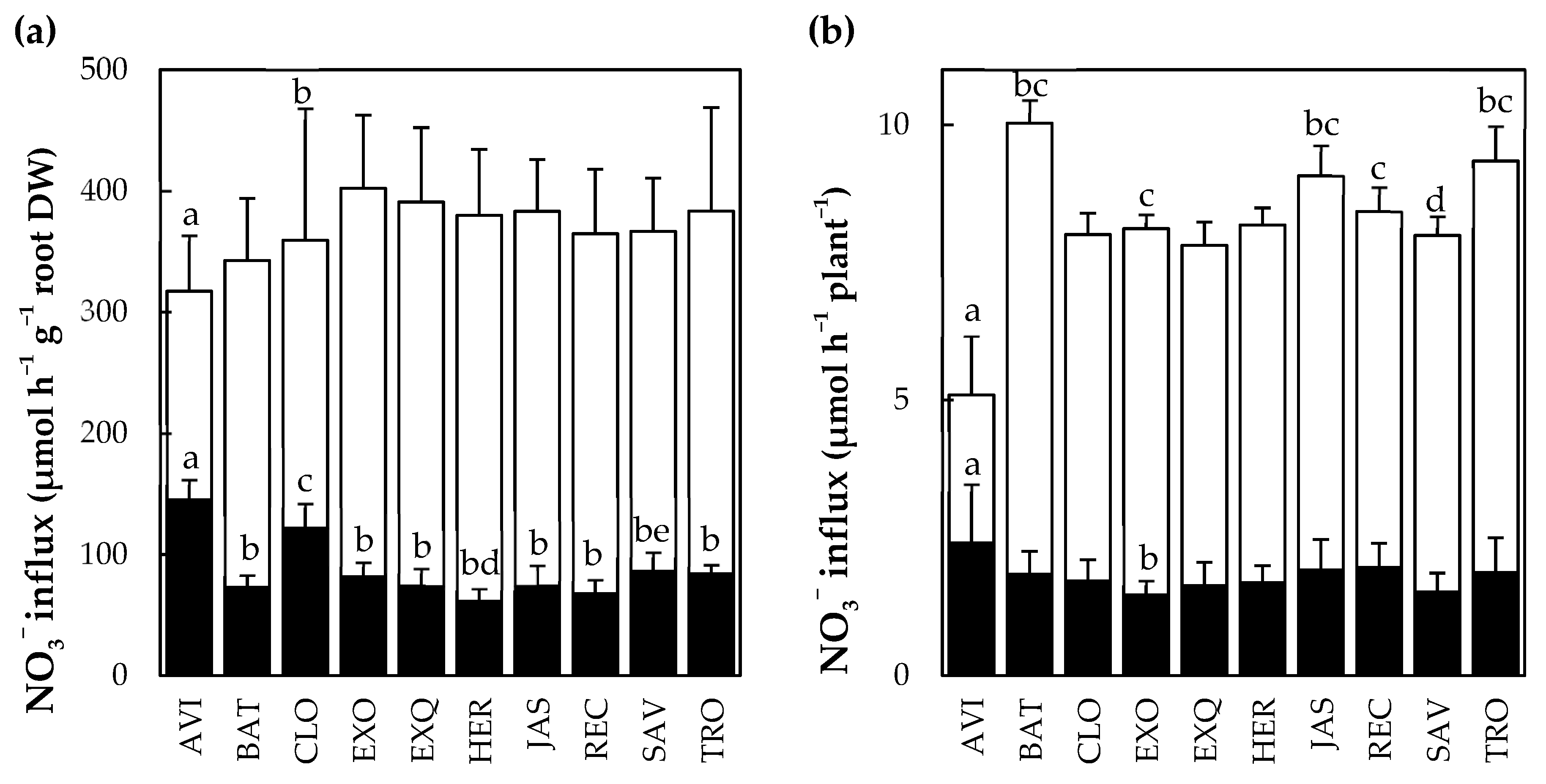
3.2. Characterization of Selected Cultivars in Hydroponics
3.3. In-Soil Root Phenotyping
4. Discussion
- (i)
- The in vitro culture system on vertical agar plate in sterile conditions, is a reliable approach for germinating seeds and observing two-dimensional root morphology. We have adapted the system, which is conveniently used for Arabidopsis model [41,49], to the more vigorous growth of oilseed rape (Figure 1a). That method is low cost but handling agar plates and analyzing scanned images are two laborious tasks. Despite, some algorithms could be used to speed up the extraction of root morphological traits from image data [50]. Finally, agar plate for culturing sprouts constitutes one first step towards further characterization of later developmental stages.
- (ii)
- The hydroponic culture in containers filled with nutrient solution (Figure 1b), is suitable for mineral nutrition studies. That culture system without substrate gives easy access to clean root organs, for physiological and molecular characterization data [51,52]. While plant biometrics (e.g., biomass and total root surface) can easily be monitored over time, no such detailed analysis of root morphological traits can be provided.
- (iii)
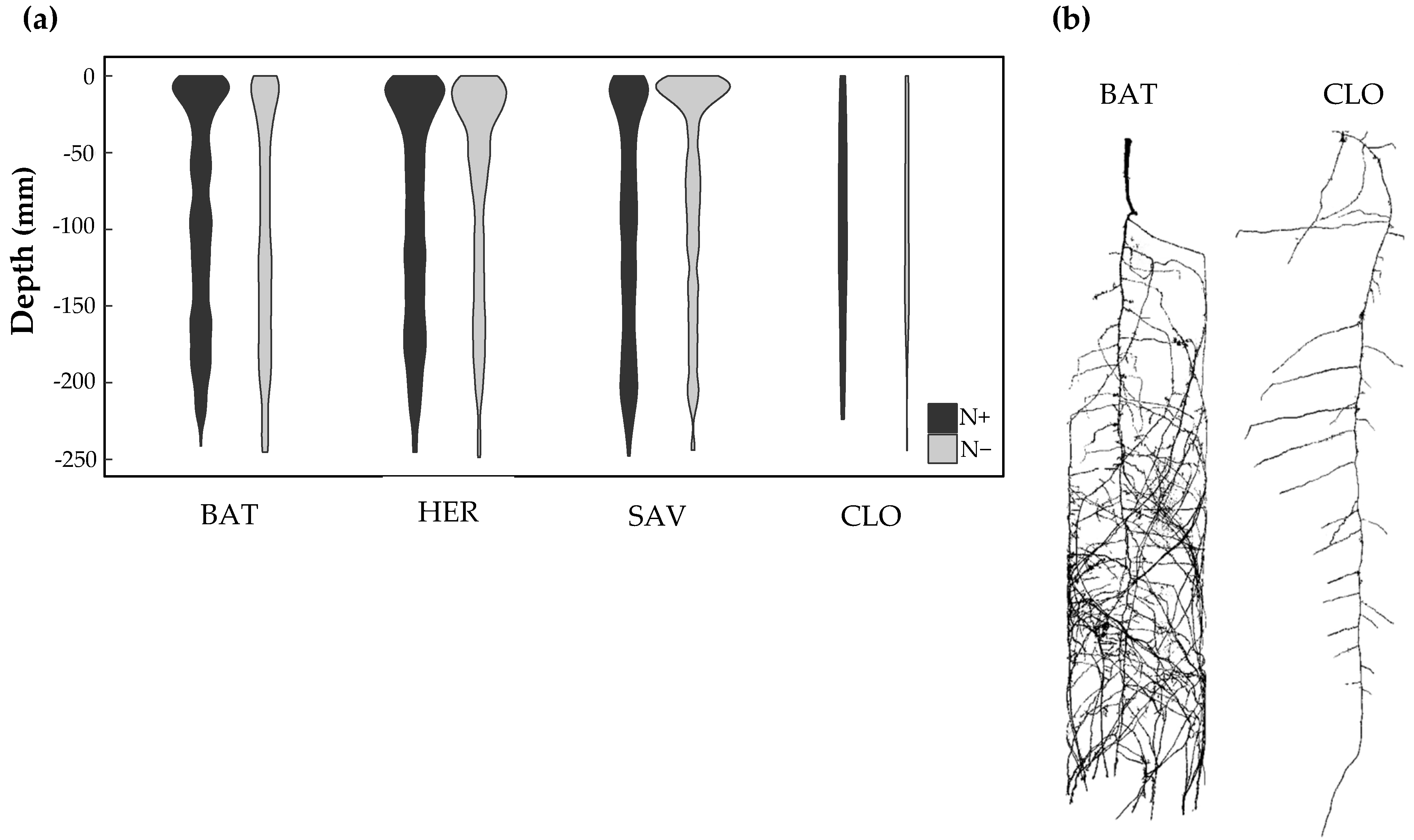
- The technique based on X-ray computed tomography allows to visualize the three-dimensional structure of the root system [48,53,54]. We recovered the segmentation of the main root and first-order lateral roots of four oilseed rape cultivars in a column filled with soil (Figure 1c). That experimental setting is low throughput, as image capture and analysis are time-consuming. The technique relies on discriminating differences in X-ray attenuation between roots and soil matrix. In this case, distinguishing between water-filled pore space and fine roots quickly became a challenge. Hence, the technique only provides a qualitative assessment of the root system in a small size pipe. Nonetheless, this may constitute a final laboratory phase prior to field phenomics [48]. For a small number of cultivars, the root phenotype at germination during in vitro culture (Figure 2) was indicative of the root system deployment in the soil (Figure 6). However, these observations should be treated with caution, as studies in other crops (e.g., wheat) are reporting a lack of correlation between the root systems at a young developmental stage in the laboratory environment and at a reproductive stage in the field [55].
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Huang, T.; Ju, X.; Yang, H. Nitrate leaching in a winter wheat-summer maize rotation on a calcareous soil as affected by nitrogen and straw management. Sci. Rep. 2017, 8, 42247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mateo-Marìn, N.; Quìlez, D.; Isla, R. Utility of stabilized nitrogen fertilizers to reduce nitrate leaching under optimal management practices. J. Plant Nutr. Soil Sci. 2020, 183, 567–578. [Google Scholar] [CrossRef]
- Velthof, G.L.; Lesschen, J.P.; Webb, J.; Pietrzak, S.; Miatkowski, Z.; Pinto, M.; Kros, J.; Oenema, O. The impact of the Nitrates Directive on nitrogen emissions from agriculture in the EU-27 during 2000–2008. Sci. Total. Environ. 2014, 468–469, 1225–1233. [Google Scholar] [CrossRef]
- He, T.; Yuan, J.; Luo, J.; Wang, W.; Fan, J.; Liu, D.; Ding, W. Organic fertilizers have divergent effects on soil N2O emissions. Biol. Fert. Soils 2019, 55, 685–699. [Google Scholar] [CrossRef]
- Sharma, L.K.; Bali, S.K. A review of methods to improve nitrogen use efficiency in agriculture. Sustainability 2018, 10, 51. [Google Scholar] [CrossRef] [Green Version]
- Moll, R.H.; Kamprath, E.J.; Jackson, W.A. Analysis and interpretation of factors which contribute to efficiency to nitrogen utilization. Agron. J. 1982, 74, 562–564. [Google Scholar] [CrossRef]
- Bouchet, A.-S.; Laperche, A.; Bissuel-Belaygue, C.; Baron, C.; Morice, J.; Rousseau-Gueutin, M.; Dheu, J.-E.; George, P.; Pinochet, X.; Foubert, T.; et al. Genetic basis of nitrogen use efficiency and yield stability across environments in winter rapeseed. BMC Genet. 2016, 17, 131. [Google Scholar] [CrossRef] [Green Version]
- Smith, S.; De Smet, I. Root system architecture: Insights from Arabidopsis and cereal crops. Phil. Trans. R. Soc. B 2012, 367, 1441–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnett, T.; Conn, V.; Kaiser, B.N. Root based approaches to improve nitrogen use efficiency in plants. Plant Cell Environ. 2009, 32, 1272–1283. [Google Scholar] [CrossRef]
- Rosolem, C.A.; Ritz, K.; Cantarella, H.; Galdos, M.V.; Hawkesford, M.J.; Whalley, W.R.; Mooney, S.J. Enhanced plant rooting and crop system management for improving N Use Efficiency. Adv. Agron. 2017, 146, 205–239. [Google Scholar]
- Sylvester-Bradley, R.; Kindred, D.R. Analysing nitrogen responses of cereals to prioritize routes to the improvement of nitrogen use efficiency. J. Exp. Bot. 2009, 60, 1939–1951. [Google Scholar] [CrossRef] [Green Version]
- Han, M.; Okamoto, M.; Beatty, P.H.; Rothstein, S.J.; Good, A.G. The genetics of nitrogen use efficiency in crop plants. Annu. Rev. Genet. 2015, 49, 269–289. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dun, X.; Shi, J.; Wang, X.; Liu, G.; Wang, H. Genetic dissection of root morphological traits related to nitrogen use efficiency in Brassica napus L. under two contrasting nitrogen conditions. Front. Plant Sci. 2017, 8, 1709. [Google Scholar] [CrossRef] [Green Version]
- Girondé, A.; Etienne, P.; Trouverie, J.; Bouchereau, A.; Le Cahérec, F.; Leport, L.; Orsel, M.; Niogret, M.-F.; Nesi, N.; Carole, D.; et al. The contrasting N management of two oilseed rape genotypes reveals the mechanisms of proteolysis associated with leaf N remobilization and the respective contributions of leaves and stems to N storage and remobilization during seed filling. BMC Plant Biol. 2015, 15, 59. [Google Scholar] [CrossRef] [Green Version]
- Koelin-Findeklee, F.; Becker, M.A.; van der Graaff, E.; Roitsch, T.; Horst, W.J. Differences between winter oilseed rape (Brassica napus L.) cultivars in nitrogen starvation-induced leaf senescence are governed by leaf-inherent rather than root-derived signals. J. Exp. Bot. 2015, 66, 3669–3681. [Google Scholar] [CrossRef] [Green Version]
- Han, Y.; Liao, J.; Yu, Y.; Song, H.; Rong, N.; Guan, C.; Lepo, J.E.; Ismail, A.M.; Zhang, Z. Exogenous abscisic acid promotes the nitrogen use efficiency of Brassica napus by increasing nitrogen remobilization in the leaves. J. Plant Nutr. 2017, 40, 18. [Google Scholar] [CrossRef]
- Williams, S.T.; Vail, S.; Arcand, M.M. Nitrogen Use Efficiency in parent vs. hybrid canola under varying nitrogen availabilities. Plants 2021, 10, 2364. [Google Scholar] [CrossRef] [PubMed]
- Louvieaux, J.; Leclercq, A.; Haelterman, L.; Hermans, C. In-field observation of root growth and nitrogen uptake efficiency of winter oilseed rape. Agronomy 2020, 10, 105. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Carrasquer, V.; Laperche, A.; Bissuel-Bélaygue, C.; Chelle, M.; Richard-Molard, C. Nitrogen Uptake Efficiency mediated by fine root growth early determines temporal and genotypic variations in Nitrogen Use Efficiency of Winter Oilseed Rape. Front. Plant Sci. 2021, 12, 641459. [Google Scholar] [CrossRef] [PubMed]
- Fletcher, R.S.; Mullen, J.L.; Heiliger, A.; McKay, J.K. QTL analysis of root morphology, flowering time, and yield reveals trade-offs in response to drought in Brassica napus. J. Exp. Bot. 2015, 66, 245–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.L.; Alcock, T.D.; Graham, N.S.; Hayden, R.; Matterson, S.; Wilson, L.; Young, S.D.; Dupuy, L.X.; White, P.J.; Hammond, J.P.; et al. Root morphology and seed and leaf ionomic traits in a Brassica napus L. diversity panel show wide phenotypic variation and are characteristic of crop habit. BMC Plant Biol. 2016, 16, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thomas, C.L.; Graham, N.S.; Hayden, R.; Meacham, M.C.; Neugebauer, K.; Nightingale, M.; Dupuy, L.X.; Hammond, J.P.; White, P.J.; Broadley, M.R. High-throughput phenotyping (HTP) identifies seedling root traits linked to variation in seed yield and nutrient capture in field-grown oilseed rape (Brassica napus L.). Ann. Bot. 2016, 118, 655–665. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Thomas, C.L.; Xiang, J.; Long, Y.; Wang, X.; Zou, J.; Luo, Z.; Ding, G.; Cai, H.; Graham, N.S.; et al. QTL meta-analysis of root traits in Brassica napus under contrasting phosphorus supply in two growth systems. Sci. Rep. 2016, 6, 33113. [Google Scholar] [CrossRef] [PubMed]
- Sun, C.; Yu, J.; Hu, D. Nitrate: A crucial signal during lateral roots development. Front. Plant Sci. 2017, 8, 485. [Google Scholar] [CrossRef] [PubMed]
- Muller, B.; Guédon, Y.; Passot, S.; Lobet, G.; Nacry, P.; Pagès, L.; Wissuwa, M.; Draye, X. Lateral roots: Random diversity in adversity. Trends Plant Sci. 2019, 24, 810–825. [Google Scholar] [CrossRef]
- Jia, Z.; von Wirén, N. Signaling pathways underlying nitrogen-dependent changes in root system architecture: From model to crop species. J. Exp. Bot. 2020, 71, 4393–4404. [Google Scholar] [CrossRef]
- Meier, M.; Liu, Y.; Lay-Pruitt, K.S.; Takahashi, H.; von Wirén, N. Auxin-mediated root branching is determined by the form of available nitrogen. Nat. Plants 2020, 6, 1136–1145. [Google Scholar] [CrossRef]
- Cai, Q.; Ji, C.; Yan, Z.; Jiang, X.; Fang, J. Anatomical responses of leaf and stem of Arabidopsis thaliana to nitrogen and phosphorus addition. J. Plant Res. 2017, 130, 1035–1045. [Google Scholar] [CrossRef]
- Li, J.; Song, X.; Kong, X.; Wang, J.; Sun, W.; Zuo, K. Natural variation of Arabidopsis thaliana root architecture in response to nitrate availability. J. Plant Nutr. 2019, 42, 723–736. [Google Scholar] [CrossRef]
- Meyer, R.C.; Gryczka, C.; Neitsch, C.; Müller, M.; Bräutigam, A.; Schlereth, A.; Schön, H.; Weigelt-Fischer, K.; Altmann, T. Genetic diversity for nitrogen use efficiency in Arabidopsis thaliana. Planta 2019, 250, 41–57. [Google Scholar] [CrossRef] [Green Version]
- Yan, Z.; Eziz, A.; Tian, D.; Li, X.; Hou, X.; Peng, H.; Han, W.; Guo, Y.; Fang, J. Biomass allocation in response to nitrogen and phosphorus availability: Insight from experimental manipulations of Arabidopsis thaliana. Front. Plant Sci. 2019, 10, 598. [Google Scholar] [CrossRef] [PubMed]
- Yue, K.; Fornara, D.A.; Li, W.; Ni, X.; Peng, Y.; Liao, S.; Tan, S.; Wang, D.; Wu, F.; Yang, Y. Nitrogen addition affects plant biomass allocation but not allometric relationships among different organs across the globe. J. Plant Ecol. 2021, 14, 361–371. [Google Scholar] [CrossRef]
- McGrail, R.K.; Van Sanford, D.A.; McNear, D.H., Jr. Trait-based root phenotyping as a necessary tool for crop selection and improvement. Agronomy 2020, 10, 1328. [Google Scholar] [CrossRef]
- Khan, M.A.E.; Gemenet, D.C.; Villordon, A. Root system architecture and abiotic stress tolerance: Current knowledge in root and tuber crops. Front. Plant Sci. 2016, 7, 1584. [Google Scholar] [CrossRef] [Green Version]
- Louvieaux, J.; De Gernier, H.; Hermans, C. Exploiting genetic variability of root morphology as a lever to improve nitrogen use efficiency in oilseed rape. In Engineering Nitrogen Utilization in Crop Plants; Springer: Berlin/Heidelberg, Germany, 2018; pp. 185–201. [Google Scholar]
- Ulas, A.; Erley, G.S.A.; Kamh, M.; Wiesler, F.; Horst, W.J. Root-growth characteristics contributing to genotypic variation in nitrogen efficiency of oilseed rape. J. Plant Nutr. Soil Sci. 2012, 175, 489–498. [Google Scholar] [CrossRef]
- Guo, X.; Ma, B.; McLaughlin, N.B.; Wu, X.; Chen, B.; Gao, Y. Nitrogen utilisation-efficient oilseed rape (Brassica napus) genotypes exhibit stronger growth attributes from flowering stage onwards. Funct. Plant Biol. 2021, 48, 755–765. [Google Scholar] [CrossRef] [PubMed]
- Berry, P.M.; Spink, J.; Foulkes, M.J.; White, P.J. The physiological basis of genotypic differences in nitrogen use efficiency in oilseed rape (Brassica napus L.). Field Crops Res. 2010, 119, 365–373. [Google Scholar] [CrossRef]
- Lynch, P.J. Root phenes that reduce the metabolic costs of soil exploration: Opportunities for 21st century agriculture. Plant Cell Environ. 2015, 38, 1775–1784. [Google Scholar] [CrossRef] [PubMed]
- Hermans, C.; Porco, S.; Verbruggen, N.; Bush, D. Chitinase-like protein CTL1 plays a role in the root system plasticity in response to multiple environmental signals. Plant Physiol. 2010, 152, 904–917. [Google Scholar] [CrossRef] [Green Version]
- De Pessemier, J.; Chardon, F.; Juraniec, M.; Delaplace, P.; Hermans, C. Natural variation of the root morphological response to nitrate supply in Arabidopsis thaliana. Mech. Develop. 2013, 130, 45–53. [Google Scholar] [CrossRef]
- Pound, M.P.; French, A.P.; Atkinson, J.A.; Wells, D.M.; Bennett, M.J.; Pridmore, T. RootNav: Navigating images of complex root architectures. Plant Physiol. 2013, 162, 1802–1814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hermans, C.; Vuylsteke, M.; Coppens, F.; Craciun, A.; Inzé, D.; Verbruggen, N. The early transcriptomic changes induced by magnesium deficiency in Arabidopsis thaliana reveal the alteration of circadian clock genes expression in roots and the triggering of ABA-responsive genes. New Phytol. 2010, 187, 119–131. [Google Scholar] [CrossRef]
- Hermans, C.; Vuylsteke, M.; Coppens, F.; Cristescu, S.; Harren, F.J.M.; Inzé, D.; Verbruggen, N. System analysis of the responses to long term magnesium deficiency and restoration in Arabidopsis thaliana. New Phytol. 2010, 187, 132–144. [Google Scholar] [CrossRef]
- Malagoli, P.; Laineé, P.; Deunff, E.; Rossato, L.; Ney, B.; Ourry, A. Modeling nitrogen uptake in oilseed rape cv Capitol during a growth cycle using influx kinetics of root nitrate transport systems and field experimental data. Plant Physiol. 2004, 134, 388–400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wasson, A.P.; Richards, R.A.; Chatrath, R.; Misra, S.C.; Prasad, S.V.; Rebetzke, G.J.; Kirkegaard, J.A.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Voss-Fels, K.P.; Qian, L.; Parra-Londono, S.; Uptmoor, R.; Frisch, M.; Keeble-Gagnère, G.; Appels, R.; Snowdon, R.J. Linkage drag constrains the roots of modern wheat. Plant Cell Environ. 2017, 40, 717–725. [Google Scholar] [CrossRef]
- Louvieaux, J.; Spanoghe, M.; Hermans, C. Root morphological traits of seedlings are predictors of seed yield and quality in winter oilseed rape hybrid cultivars. Front. Plant. Sci. 2021, 11, 568009. [Google Scholar] [CrossRef]
- Hu, Y.; Omary, M.; Hu, Y.; Doron, O.; Hoermayer, L.; Chen, Q.; Megides, O.; Chekli, O.; Ding, Z.; Friml, J.; et al. Cell kinetics of auxin transport and activity in Arabidopsis root growth and skewing. Nat. Commun. 2021, 12, 1657. [Google Scholar] [CrossRef]
- Dupuy, L.X.; Wright, G.; Thompson, J.A.; Taylor, A.; Dekeyser, S.; White, C.P.; Thomas, W.T.B.; Nightingale, M.; Hammond, J.P.; Graham, N.S.; et al. Accelerating root system phenotyping of seedlings through a computer-assisted processing pipeline. Plant Methods 2017, 13, 57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Z.; Ma, J.; Lei, P.; Wang, Q.; Feng, X.; Xu, H. Poly-γ-glutamic acid induces system tolerance to drought stress by promoting abscisic acid accumulation in Brassica napus L. Sci. Rep. 2020, 14, 252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Murad, M.A.; Razi, K.; Benjamin, L.K.; Lee, J.H.; Kim, T.H.; Muneer, S. Ethylene regulates sulfur acquisition by regulating the expression of sulfate transporter genes in oilseed rape. Physiol. Plant. 2021, 171, 533–545. [Google Scholar] [CrossRef] [PubMed]
- Mairhofer, S.; Pridmore, T.; Johnson, J.; Wells, D.M.; Bennett, M.J.; Mooney, S.J.; Sturrock, C.J. X-ray Computed Tomography of crop plant root systems grown in soil. Curr. Protoc. Plant Biol. 2017, 2, 270–286. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, J.; Kirchgessner, N.; Colombi, T.; Walter, A. Rapid phenotyping of crop root systems in undisturbed field soils using X-ray computed tomography. Plant Methods 2015, 11, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watt, M.; Moosavi, S.; Cunningham, S.C.; Kirkegaard, J.A.; Rebetzke, G.J.; Richards, R.A. A rapid, controlled-environment seedling root screen for wheat correlates well with rooting depths at vegetative, but not reproductive, stages at two field sites. Ann. Bot. 2013, 112, 447–455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leijten, W.; Koes, R.; Roobeek, I.; Frugis, G. Translating flowering time from Arabidopsis thaliana to Brassicaceae and Asteraceae crop species. Plants 2018, 7, 111. [Google Scholar] [CrossRef] [Green Version]
- Stephenson, P.; Stacey, N.; Brüser, M.; Pullen, N.; Ilyas, M.; O’Neill, C.; Wells, R.; Østergaard, L. The power of model-to-crop translation illustrated by reducing seed loss from pod shatter in oilseed rape. Plant Reprod. 2019, 32, 331–340. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Zeng, R.; Liao, H. Improving crop nutrient efficiency through root architecture modifications. J. Integr. Plant Biol. 2016, 58, 193–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorup-Kristensen, K.; Kirkegaard, J. Root system-based limits to agricultural productivity and efficiency: The farming systems context. Ann. Bot. 2016, 118, 573–592. [Google Scholar] [CrossRef]
- Pierret, A.; Maeght, J.; Clément, C.; Montoroi, J.; Hartmann, C.; Gonkhamdee, S. Understanding deep roots and their functions in ecosystems: An advocacy for more unconventional research. Ann. Bot. 2016, 118, 621–635. [Google Scholar] [CrossRef] [Green Version]
- Van der Bom, F.J.T.; Williams, A.; Bell, M.J. Root architecture for improved resource capture: Trade-offs in complex environments. J. Exp. Bot. 2020, 71, 5752–5763. [Google Scholar] [CrossRef]
- Gu, M.G.; Hu, X.; Wang, T.; Xu, G. Modulation of plant root traits by nitrogen and phosphate: Transporters, long-distance signaling proteins and peptides, and potential artificial traps. Breed Sci. 2021, 71, 62–75. [Google Scholar] [CrossRef] [PubMed]
- Ohkubo, Y.H.; Tanaka, M.; Tabata, R.; Ogawa-Ohnishi, M.; Matsubayashi, Y. Shoot-to-root mobile polypeptides involved in systemic regulation of nitrogen acquisition. Nat. Plants 2017, 3, 17029. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biomass Production | |
| R | Root biomass |
| S | Shoot biomass |
| R + S | Total biomass |
| R:S | Root to shoot biomass ratio |
| Root Morphology | |
| LPR | Length of primary root; LPR = LZ2 + LZ3 + LZ4 |
| LZ2 | Length of primary root zone 2, between the first and last lateral roots; = 0 if NLR = 0 or NLR = 1 |
| LZ3 | Length of primary root zone 3, between the hypocotyl junction and the first lateral root; = LPR if NLR ≤ 1 |
| LZ4 | Length of primary root zone 4, between the last lateral root and the primary root tip; = 0 if NLR ≤ 1 |
| NLR | Number of lateral roots > 1 mm |
| ∑LLR | Sum of lateral root lengths |
| TRL | Total root length; = LPR + ∑LLR |
| DLR-Z1 | Density of lateral roots in zone 1; = NLR/LPR |
| DLR-Z2 | Density of lateral roots in zone 2; = (NLR-1)/LZ2, not defined if NLR ≤ 1 |
| SRL | Specific root length; = (LPR + ∑LLR)/R |
| Shoot Morphology | |
| LA | Leaf area |
| SLA | Specific leaf area; = LA/S |
| Carbon and Nitrogen Analyses | |
| CR | Carbon concentration in root tissues |
| CS | Carbon concentration in shoot tissues |
| NR | Nitrogen concentration in root tissues |
| NS | Nitrogen concentration in shoot tissues |
| NUI | Nitrogen utilization index; = S/((S × NS) + (R × NR)) |
| Nitrate Uptake Assay | |
| 15NQR | 15N amount in root tissues |
| 15NQS | 15N amount in shoot tissues |
| HATS/LATS plan−1 | |
| HATS/LATS mediated 15N influx rate per hour and per plant; = (15NQR + 15NQS)/5 × 60 | |
| HATS/LATS root biomass−1 | |
| HATS/LATS mediated 15N influx rate per hour and per root biomass; = (15NQR + 15NQS)/5 × 60 × R | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kupcsik, L.; Chiodi, C.; Moturu, T.R.; De Gernier, H.; Haelterman, L.; Louvieaux, J.; Tillard, P.; Sturrock, C.J.; Bennett, M.; Nacry, P.; et al. Oilseed Rape Cultivars Show Diversity of Root Morphologies with the Potential for Better Capture of Nitrogen. Nitrogen 2021, 2, 491-505. https://doi.org/10.3390/nitrogen2040033
Kupcsik L, Chiodi C, Moturu TR, De Gernier H, Haelterman L, Louvieaux J, Tillard P, Sturrock CJ, Bennett M, Nacry P, et al. Oilseed Rape Cultivars Show Diversity of Root Morphologies with the Potential for Better Capture of Nitrogen. Nitrogen. 2021; 2(4):491-505. https://doi.org/10.3390/nitrogen2040033
Chicago/Turabian StyleKupcsik, László, Claudia Chiodi, Taraka Ramji Moturu, Hugues De Gernier, Loïc Haelterman, Julien Louvieaux, Pascal Tillard, Craig J. Sturrock, Malcolm Bennett, Philippe Nacry, and et al. 2021. "Oilseed Rape Cultivars Show Diversity of Root Morphologies with the Potential for Better Capture of Nitrogen" Nitrogen 2, no. 4: 491-505. https://doi.org/10.3390/nitrogen2040033