
Taurocholic Acid and Glycocholic Acid Inhibit Inflammation and Activate Farnesoid X Receptor Expression in LPS-Stimulated Zebrafish and Macrophages
Abstract
:1. Introduction
2. Results
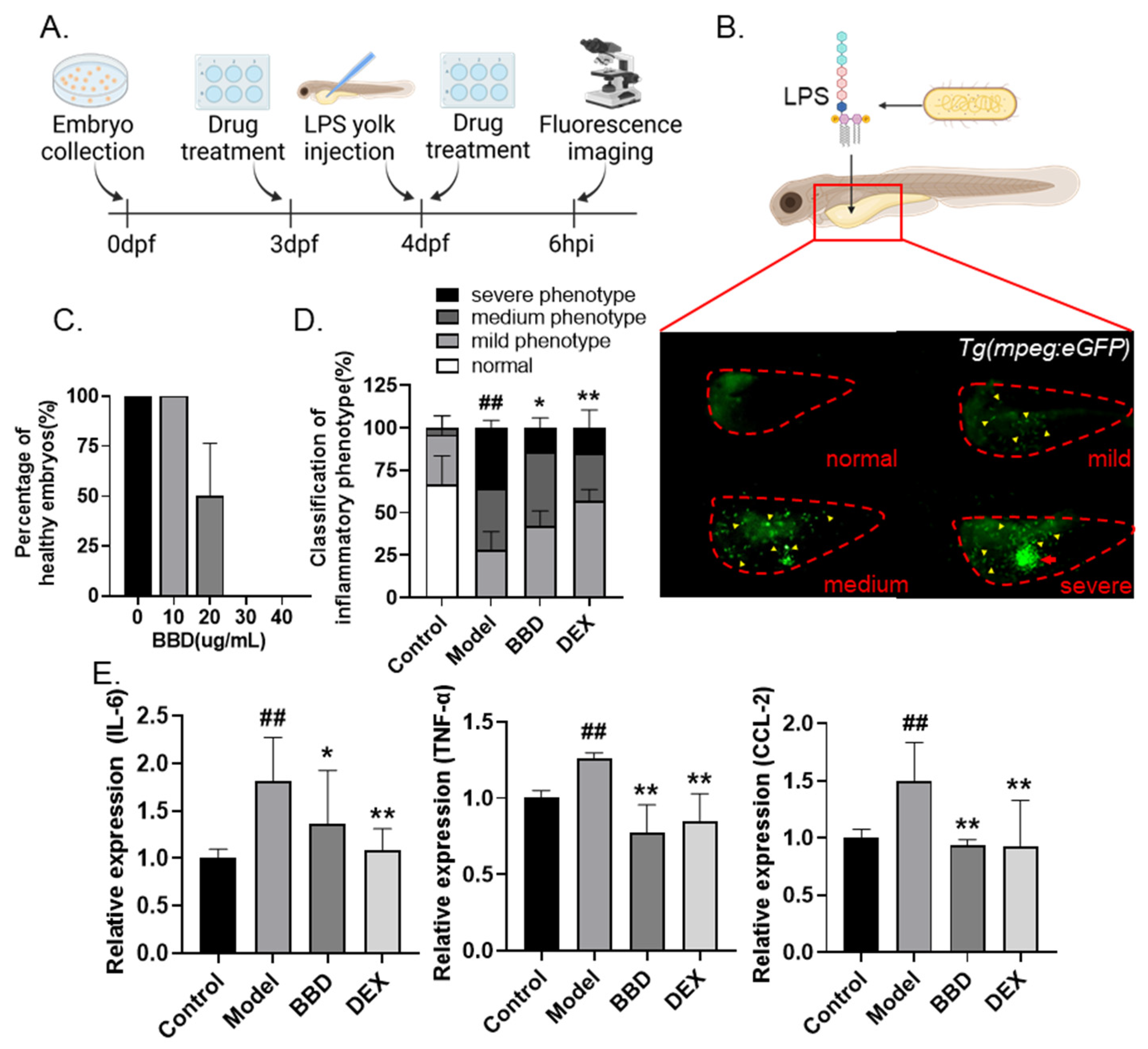
2.1. BBD Extract Attenuates LPS-Induced Zebrafish Inflammation
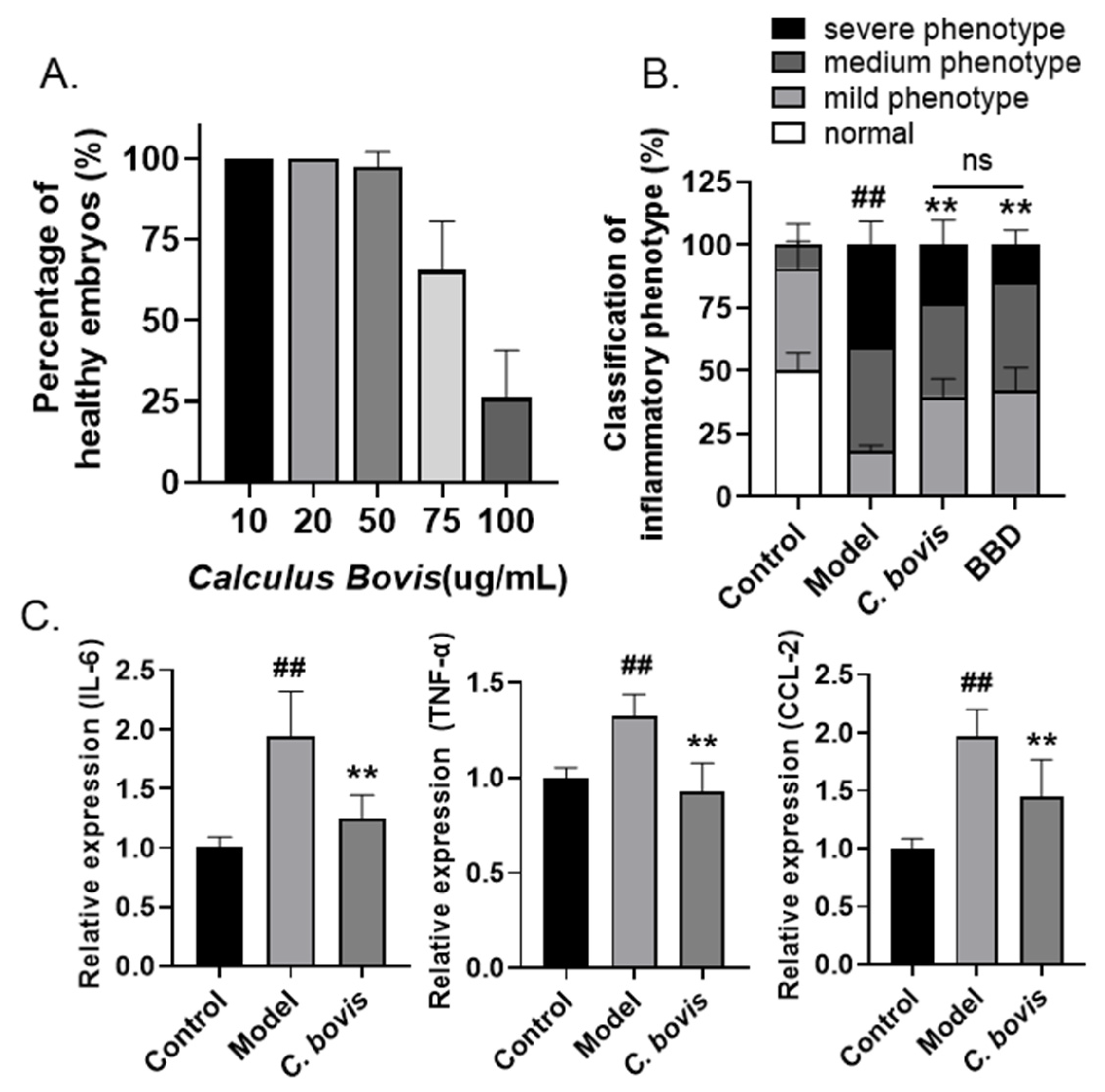
2.2. Calculus bovis Extract Attenuates LPS-Induced Zebrafish Yolk Sac Inflammation
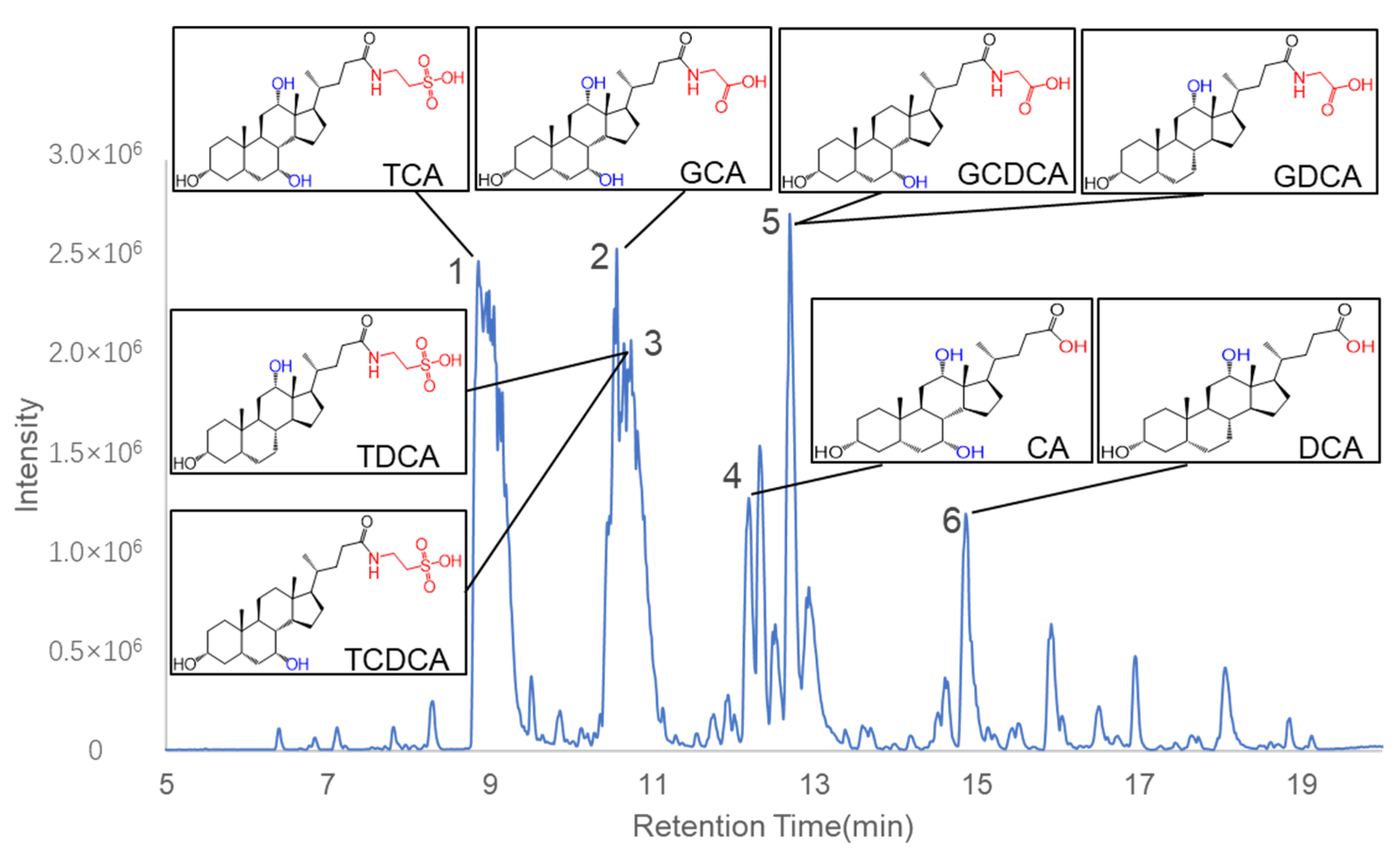
2.3. Identification of Chemical Constituents in C. bovis Extract
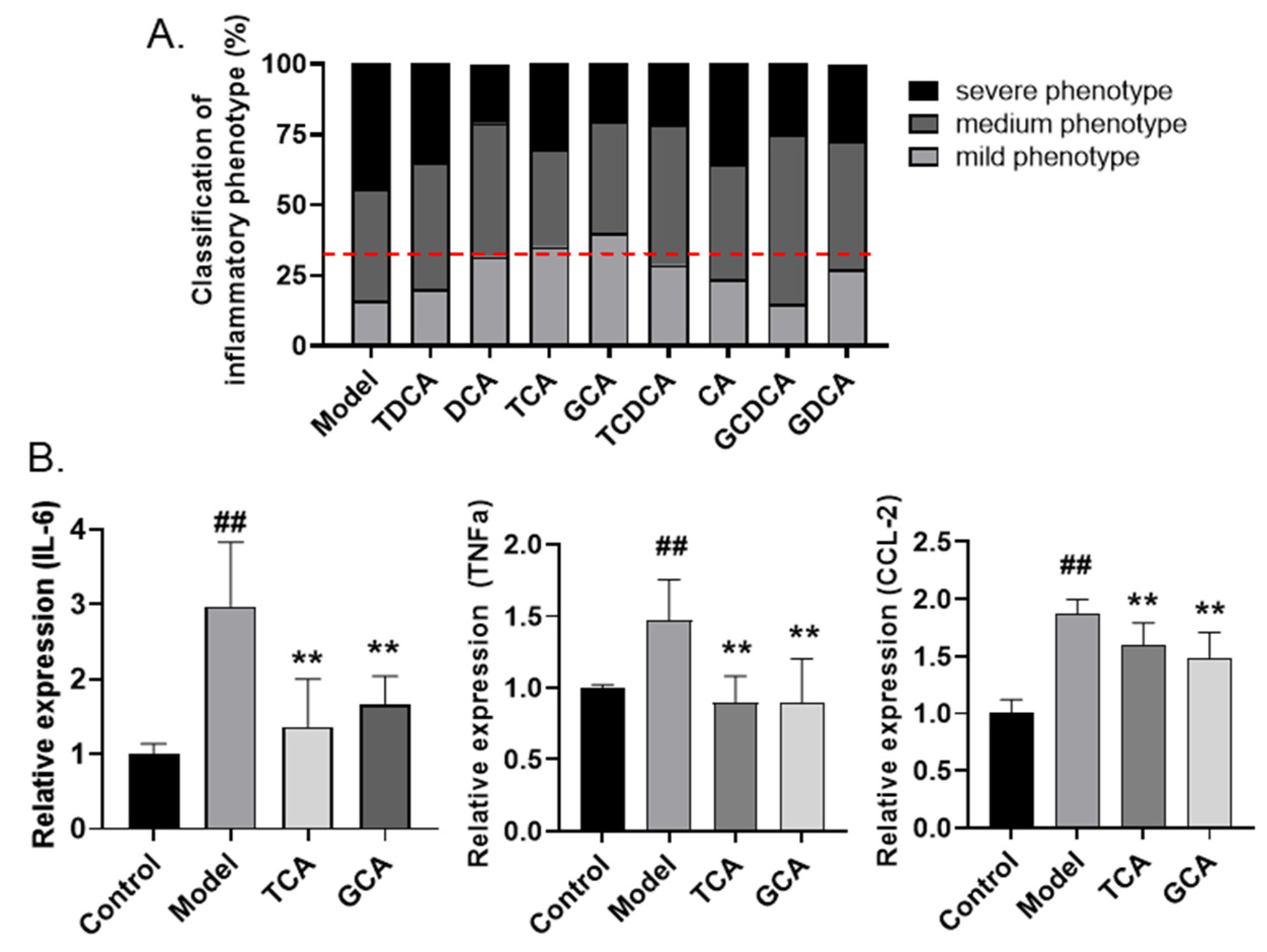
2.4. Screening of Anti-Inflammatory Bile Acids in Zebrafish Inflammatory Model
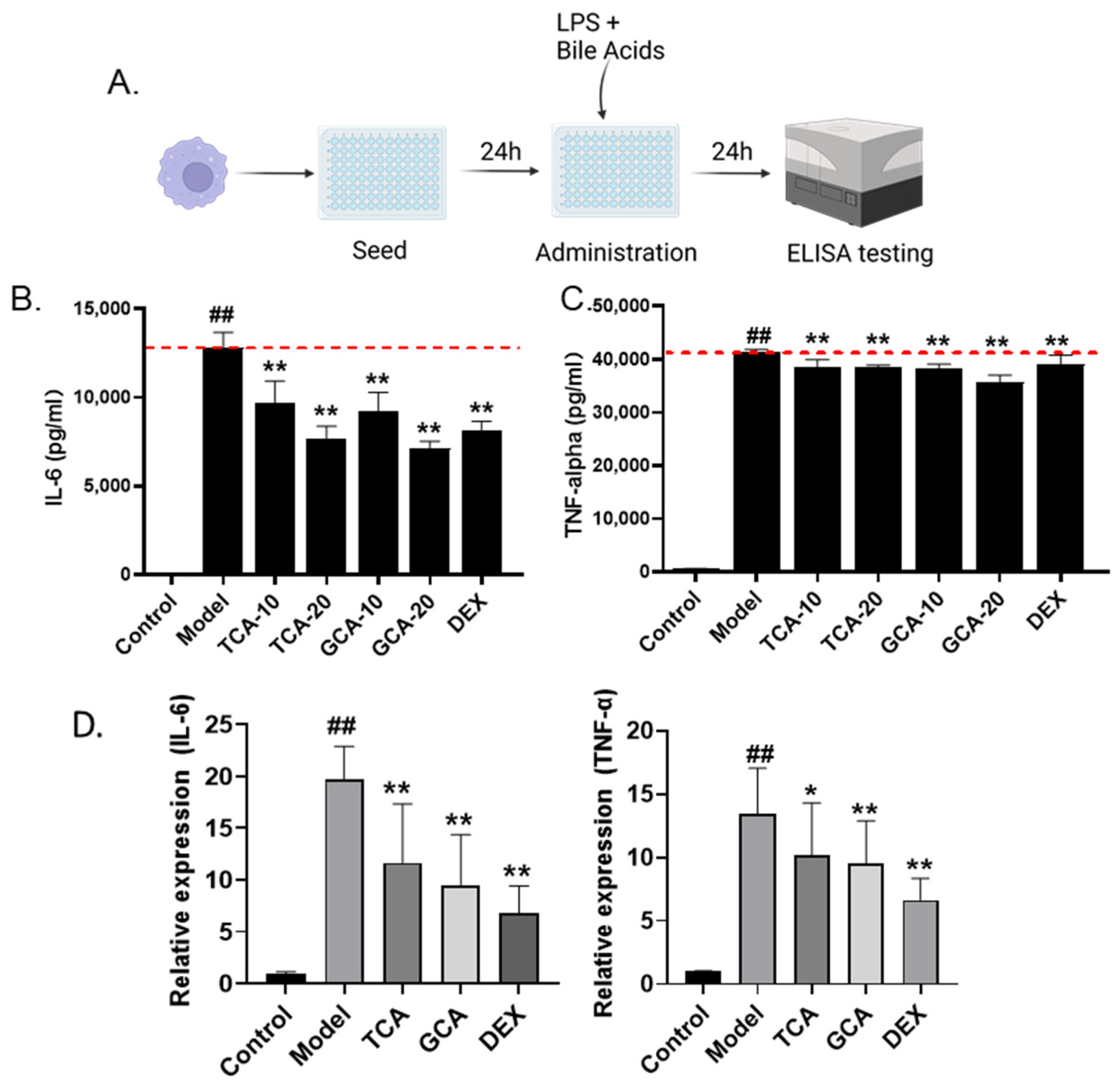
2.5. Validation of Anti-Inflammatory Bile Acids in LPS-Stimulated Macrophages
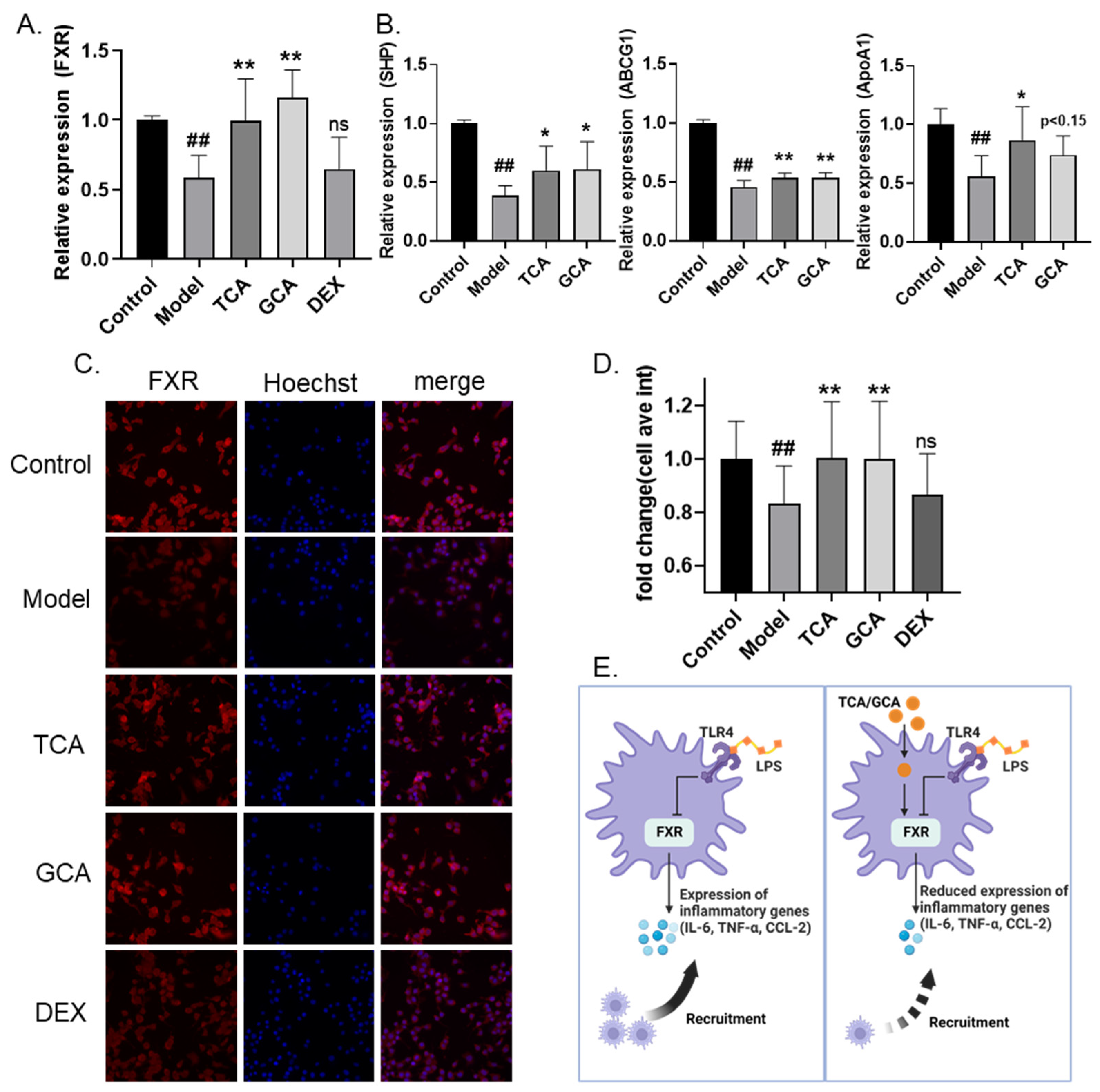
2.6. TCA and GCA Increase the Expression of Farnesoid X Receptor
3. Discussion
4. Materials and Methods
4.1. Sample Preparation
4.2. Zebrafish Husbandry and Animal Care Ethics
4.3. Zebrafish Acute Inflammation Model and Drug Administration
4.4. UPLC-QTOF-MS Analysis
4.5. Cell Culture and Drug Administration
4.6. qPCR
4.7. ELISA Assay
4.8. Immunofluorescence
4.9. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Vincent, J.-L.; Marshall, J.C.; A Ñamendys-Silva, S.A.; François, B.; Martin-Loeches, I.; Lipman, J.; Reinhart, K.; Antonelli, M.; Pickkers, P.; Njimi, H.; et al. Assessment of the worldwide burden of critical illness: The Intensive Care Over Nations (ICON) audit. Lancet Respir. Med. 2014, 2, 380–386. [Google Scholar] [CrossRef]
- Medzhitov, R. Origin and physiological roles of inflammation. Nature 2008, 454, 428–435. [Google Scholar] [CrossRef]
- Borish, L.C.; Steinke, J.W. 2. Cytokines and chemokines. J. Allergy Clin. Immunol. 2003, 111 (Suppl. 2), S460–S475. [Google Scholar] [CrossRef] [PubMed]
- Nathan, C.; Ding, A. Nonresolving Inflammation. Cell 2010, 140, 871–882. [Google Scholar] [CrossRef] [Green Version]
- Ziesmann, M.T.; Marshall, J.C. Multiple Organ Dysfunction: The Defining Syndrome of Sepsis. Surg. Infect. 2018, 19, 184–190. [Google Scholar] [CrossRef] [PubMed]
- van der Poll, T.; van de Veerdonk, F.L.; Scicluna, B.P.; Netea, M.G. The immunopathology of sepsis and potential therapeutic targets. Nat. Rev. Immunol. 2017, 17, 407–420. [Google Scholar] [CrossRef] [PubMed]
- van der Poll, T.; Shankar-Hari, M.; Wiersinga, W.J. The immunology of sepsis. Immunity 2021, 54, 2450–2464. [Google Scholar] [CrossRef] [PubMed]
- Hu, B.; Huang, S.; Yin, L. The cytokine storm and COVID-19. J. Med. Virol. 2021, 93, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Ye, Q.; Wang, B.; Mao, J. The pathogenesis and treatment of the ‘Cytokine Storm’ in COVID-19. J. Infect. 2020, 80, 607–613. [Google Scholar] [CrossRef]
- Bindu, S.; Mazumder, S.; Bandyopadhyay, U. Non-steroidal anti-inflammatory drugs (NSAIDs) and organ damage: A current perspective. Biochem. Pharmacol. 2020, 180, 114147. [Google Scholar] [CrossRef]
- Whitehouse, M.W. Anti-inflammatory glucocorticoid drugs: Reflections after 60 years. Inflammopharmacology 2010, 19, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Thomford, N.E.; Senthebane, D.A.; Rowe, A.; Munro, D.; Seele, P.; Maroyi, A.; Dzobo, K. Natural Products for Drug Discovery in the 21st Century: Innovations for Novel Drug Discovery. Int. J. Mol. Sci. 2018, 19, 1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atanasov, A.G.; Zotchev, S.B.; Dirsch, V.M.; Supuran, C.T. Natural products in drug discovery: Advances and opportunities. Nat. Rev. Drug Discov. 2021, 20, 200–216. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.-J.; Xu, Y.; Peng, W.; Liu, Y.-J.; Zhang, J.-M.; Li, J.-S.; Sun, T.; Wang, P. Calculus bovis: A review of the traditional usages, origin, chemistry, pharmacological activities and toxicology. J. Ethnopharmacol. 2020, 254, 112649. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.-F.; Sheng, H.-D.; Qian, J.; Wang, Y. The Chinese medicine babaodan suppresses LPS-induced sepsis by inhibiting NLRP3-mediated inflammasome activation. J. Ethnopharmacol. 2022, 292, 115205. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, Z.; Du, K.; Liang, M.; Zhu, X.; Yu, Z.; Chen, R.; Qin, L.; Li, Y.; Zheng, Y. Babaodan inhibits cell growth by inducing autophagy through the PI3K/AKT/mTOR pathway and enhances antitumor effects of cisplatin in NSCLC cells. Am. J. Transl. Res. 2019, 11, 5272–5283. [Google Scholar] [PubMed]
- Zhao, J.; Lan, W.; Peng, J.; Guan, B.; Liu, J.; Zhang, M.; Zhan, Z.; Lin, J. Babao Dan Reverses Multiple-Drug Resistance in Gastric Cancer Cells via Triggering Apoptosis and Autophagy and Inhibiting PI3K/AKT/mTOR Signaling. Evid.-Based Complement. Altern. Med. 2021, 2021, 5631942. [Google Scholar] [CrossRef] [PubMed]
- Sheng, D.; Zhao, S.; Gao, L.; Zheng, H.; Liu, W.; Hou, J.; Jin, Y.; Ye, F.; Zhao, Q.; Li, R.; et al. BabaoDan attenuates high-fat diet-induced non-alcoholic fatty liver disease via activation of AMPK signaling. Cell Biosci. 2019, 9, 77. [Google Scholar] [CrossRef]
- Lu, L.; Wu, C.; Lu, B.-J.; Xie, D.; Wang, Z.; Azami, N.L.B.; An, Y.-T.; Wang, H.-J.; Ye, G.; Sun, M.-Y. BabaoDan cures hepatic encephalopathy by decreasing ammonia levels and alleviating inflammation in rats. J. Ethnopharmacol. 2019, 249, 112301. [Google Scholar] [CrossRef]
- Qian, J.; Xu, H.; Lv, D.; Liu, W.; Chen, E.; Zhou, Y.; Wang, Y.; Ying, K.; Fan, X. Babaodan controls excessive immune responses and may represent a cytokine-targeted agent suitable for COVID-19 treatment. Biomed. Pharmacother. 2021, 139, 111586. [Google Scholar] [CrossRef]
- Zanandrea, R.; Bonan, C.D.; Campos, M.M. Zebrafish as a model for inflammation and drug discovery. Drug Discov. Today 2020, 25, 2201–2211. [Google Scholar] [CrossRef] [PubMed]
- Ellett, F.; Pase, L.; Hayman, J.W.; Andrianopoulos, A.; Lieschke, G.J. mpeg1 promoter transgenes direct macrophage-lineage expression in zebrafish. Blood 2011, 117, e49–e56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, C.; Flores, M.V.; Storm, T.; Crosier, K.; Crosier, P. The zebrafish lysozyme C promoter drives myeloid-specific expression in transgenic fish. BMC Dev. Biol. 2007, 7, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Xie, R.; Qian, Q.; Yan, J.; Wang, H.; Wang, X. Triclosan induced zebrafish immunotoxicity by targeting miR-19a and its gene socs3b to activate IL-6/STAT3 signaling pathway. Sci. Total Environ. 2022, 815, 152916. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Chen, J.; Zhang, X.; Wang, Y.; Wang, S.; Zhao, L.; Wang, Y. Identification of anti-inflammatory compounds from Zhongjing formulae by knowledge mining and high-content screening in a zebrafish model of inflammatory bowel diseases. Chin. Med. 2021, 16, 42. [Google Scholar] [CrossRef]
- Zhao, L.; Liu, H.; Wang, Y.; Wang, S.; Xun, D.; Wang, Y.; Cheng, Y.; Zhang, B. Multimodal Identification by Transcriptomics and Multiscale Bioassays of Active Components in Xuanfeibaidu Formula to Suppress Macrophage-Mediated Immune Response. Engineering 2021. [Google Scholar] [CrossRef]
- Yang, L.; Zhou, X.; Huang, W.; Fang, Q.; Hu, J.; Yu, L.; Ma, N.; Zhang, W. Protective Effect of Phillyrin on Lethal LPS-Induced Neutrophil Inflammation in Zebrafish. Cell. Physiol. Biochem. 2017, 43, 2074–2087. [Google Scholar] [CrossRef]
- Kong, W.-J.; Xing, X.-Y.; Xiao, X.-H.; Wang, J.-B.; Zhao, Y.-L.; Yang, M.-H. Multi-component analysis of bile acids in natural Calculus bovis and its substitutes by ultrasound-assisted solid–liquid extraction and UPLC-ELSD. Anal. 2012, 137, 5845–5853. [Google Scholar] [CrossRef]
- Liu, W.-X.; Cheng, X.-L.; Guo, X.-H.; Hu, X.-R.; Wei, F.; Ma, S.-C. Identification of Calculus Bovis and its mixed varieties by ultra-high-performance liquid chromatography/quadrupole time-of-flight mass spectrometry (UHPLC-Q/TOF-MS) combined with the principal component analysis (PCA) method. J. Pharm. Biomed. Anal. 2019, 179, 112979. [Google Scholar] [CrossRef]
- Chávez-Talavera, O.; Tailleux, A.; Lefebvre, P.; Staels, B. Bile Acid Control of Metabolism and Inflammation in Obesity, Type 2 Diabetes, Dyslipidemia, and Nonalcoholic Fatty Liver Disease. Gastroenterology 2017, 152, 1679–1694.e3. [Google Scholar] [CrossRef]
- Anderson, K.M.; Gayer, C.P. The Pathophysiology of Farnesoid X Receptor (FXR) in the GI Tract: Inflammation, Barrier Function and Innate Immunity. Cells 2021, 10, 3206. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.; Zhang, C.L.; Cai, H.J.; Liu, D. Research progress in pharmacological effects of Calculus Bovis and Calculus Bovis cultivated in vitro. China Pharm. 2014, 17, 1396–1399. [Google Scholar]
- Huang, J.; Wu, J.; Sun, Y.; Gong, B.; Liu, D. Taurine inhibits inflammatory pain in rats and its mechanism. Chin. J. Mod. Drug Appl. 2016, 10, 283–284. [Google Scholar]
- Li, X.; Xu, Y.; Zhang, C.; Deng, L.; Chang, M.; Yu, Z.; Liu, D. Protective Effect of Calculus Bovis Sativus on Dextran Sulphate Sodium-Induced Ulcerative Colitis in Mice. Evid.-Based Complement. Altern. Med. 2015, 2015, 469506. [Google Scholar] [CrossRef] [Green Version]
- Zhang, N.R. Current research in Calculus Bovis. Acta Chin. Med. Pharmacol. 1994, 21, 44–48. [Google Scholar]
- Thomas, C.; Pellicciari, R.; Pruzanski, M.; Auwerx, J.; Schoonjans, K. Targeting bile-acid signalling for metabolic diseases. Nat. Rev. Drug Discov. 2008, 7, 678–693. [Google Scholar] [CrossRef]
- Chiang, J.Y. Bile acid metabolism and signaling. Compr. Physiol. 2013, 3, 1191–1212. [Google Scholar]
- Makishima, M.; Okamoto, A.Y.; Repa, J.J.; Tu, H.; Learned, R.M.; Luk, A.; Hull, M.V.; Lustig, K.D.; Mangelsdorf, D.J.; Shan, B. Identification of a Nuclear Receptor for Bile Acids. Science 1999, 284, 1362–1365. [Google Scholar]
- Lefebvre, P.; Cariou, B.; Lien, F.; Kuipers, F.; Staels, B. Role of bile acids and bile acid receptors in metabolic regulation. Physiol. Rev. 2009, 89, 147–191. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, S.; Meng, X.; Li, Y.; Sun, L.; Jiang, L.; Xuan, H.; Chen, X. Bile Acids Elevated in Chronic Periaortitis Could Activate Farnesoid-X-Receptor to Suppress IL-6 Production by Macrophages. Front. Immunol. 2021, 12, 632864. [Google Scholar] [CrossRef]
- Liu, H.; Wang, J.; Ding, Y.; Shi, X.; Ren, H. Antibiotic pretreatment attenuates liver ischemia–reperfusion injury by Farnesoid X receptor activation. Cell Death Dis. 2022, 13, 484. [Google Scholar] [CrossRef]
- Takahashi, S.; Tanaka, N.; Golla, S.; Fukami, T.; Krausz, K.W.; Polunas, M.A.; Weig, B.C.; Masuo, Y.; Xie, C.; Jiang, C.; et al. Editor’s Highlight: Farnesoid X Receptor Protects Against Low-Dose Carbon Tetrachloride-Induced Liver Injury Through the Taurocholate-JNK Pathway. Toxicol. Sci. 2017, 158, 334–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.H.; Park, M.J.; Kim, K.W.; Lee, H.C.; Choi, Y.H.; Cheong, J. The orphan nuclear receptor SHP is involved in monocytic differentiation, and its expression is increased by c-Jun. J. Leukoc. Biol. 2004, 76, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Magee, N.; Zou, A.; Ghosh, P.; Ahamed, F.; Delker, D.; Zhang, Y. Disruption of hepatic small heterodimer partner induces dissociation of steatosis and inflammation in experimental nonalcoholic steatohepatitis. J. Biol. Chem. 2020, 295, 994–1008. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Xie, S.; Chi, Z.; Zhang, J.; Liu, Y.; Zhang, L.; Zheng, M.; Zhang, X.; Xia, D.; Ke, Y.; et al. Bile Acids Control Inflammation and Metabolic Disorder through Inhibition of NLRP3 Inflammasome. Immunity 2016, 45, 802–816. [Google Scholar] [CrossRef] [Green Version]
- Yang, T.; Shu, T.; Liu, G.; Mei, H.; Zhu, X.; Huang, X.; Zhang, L.; Jiang, Z. Quantitative profiling of 19 bile acids in rat plasma, liver, bile and different intestinal section contents to investigate bile acid homeostasis and the application of temporal variation of endogenous bile acids. J. Steroid Biochem. Mol. Biol. 2017, 172, 69–78. [Google Scholar] [CrossRef] [PubMed]
- Yan, N.; Yan, T.; Xia, Y.; Hao, H.; Wang, G.; Gonzalez, F.J. The pathophysiological function of non-gastrointestinal farnesoid X receptor. Pharmacol. Ther. 2021, 226, 107867. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Chen, J.; Hollister, K.; Sowers, L.C.; Forman, B.M. Endogenous Bile Acids Are Ligands for the Nuclear Receptor FXR/BAR. Mol. Cell 1999, 3, 543–553. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NO. | RT (min) | Molecular Formula | MW | Error (ppm) | Identified Compound | Fragment Ion |
|---|---|---|---|---|---|---|
| 1 | 8.797 | C26H45NO7S | 515.29113 | 5.3 | Taurocholic acid | 514.2844[M-H]− |
| 2 | 10.504 | C26H43NO6 | 465.30849 | 2.7 | Glycocholic acid | 464.3030[M-H]− 446.2935[M-H-H2O]− 420.3130[M-H-CO2]− 402.3013[M-H-H2CO3]− |
| 3 | 10.962 | C26H45NO6S | 499.29621 | 4 | Tauroursodeoxycholic acid/Taurochenodeoxycholic acid/Taurodeoxycholic acid/Taurohyodeoxycholic acid | 498.2915[M-H]− |
| 4 | 12.133 | C24H40O5 | 408.28703 | −0.5 | Cholic acid | 407.2801[M-H]− 389.2695[M-H-H2O]− 345.2795[M-H-H2CO3]− |
| 5 | 12.312 | C26H43NO5 | 449.31358 | 4.8 | Glycoursodeoxycholic acid/Glycodeoxycholic acid/Glycohyodeoxycholic acid/Glycochenodeoxycholic acid | 448.3090[M-H]− 404.3194[M-H-CO2]− 386.3077[M-H-H2CO3]− |
| 6 | 14.808 | C24H40O4 | 392.29211 | −1 | Deoxycholic acid/Ursodeoxycholic acid/Chenodeoxycholic acid/Hyodeoxycholic Acid | 391.2850[M-H]− 347.2952[M-HMCO2]− 327.2699[M-H-H2CO3]− |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ge, X.; Huang, S.; Ren, C.; Zhao, L. Taurocholic Acid and Glycocholic Acid Inhibit Inflammation and Activate Farnesoid X Receptor Expression in LPS-Stimulated Zebrafish and Macrophages. Molecules 2023, 28, 2005. https://doi.org/10.3390/molecules28052005
Ge X, Huang S, Ren C, Zhao L. Taurocholic Acid and Glycocholic Acid Inhibit Inflammation and Activate Farnesoid X Receptor Expression in LPS-Stimulated Zebrafish and Macrophages. Molecules. 2023; 28(5):2005. https://doi.org/10.3390/molecules28052005
Chicago/Turabian StyleGe, Xutao, Shaoze Huang, Can Ren, and Lu Zhao. 2023. "Taurocholic Acid and Glycocholic Acid Inhibit Inflammation and Activate Farnesoid X Receptor Expression in LPS-Stimulated Zebrafish and Macrophages" Molecules 28, no. 5: 2005. https://doi.org/10.3390/molecules28052005