

Medicarpin and Homopterocarpin Isolated from Canavalia lineata as Potent and Competitive Reversible Inhibitors of Human Monoamine Oxidase-B
 , ,
, ,  , and
, and
Abstract
:1. Introduction
2. Results and Discussion
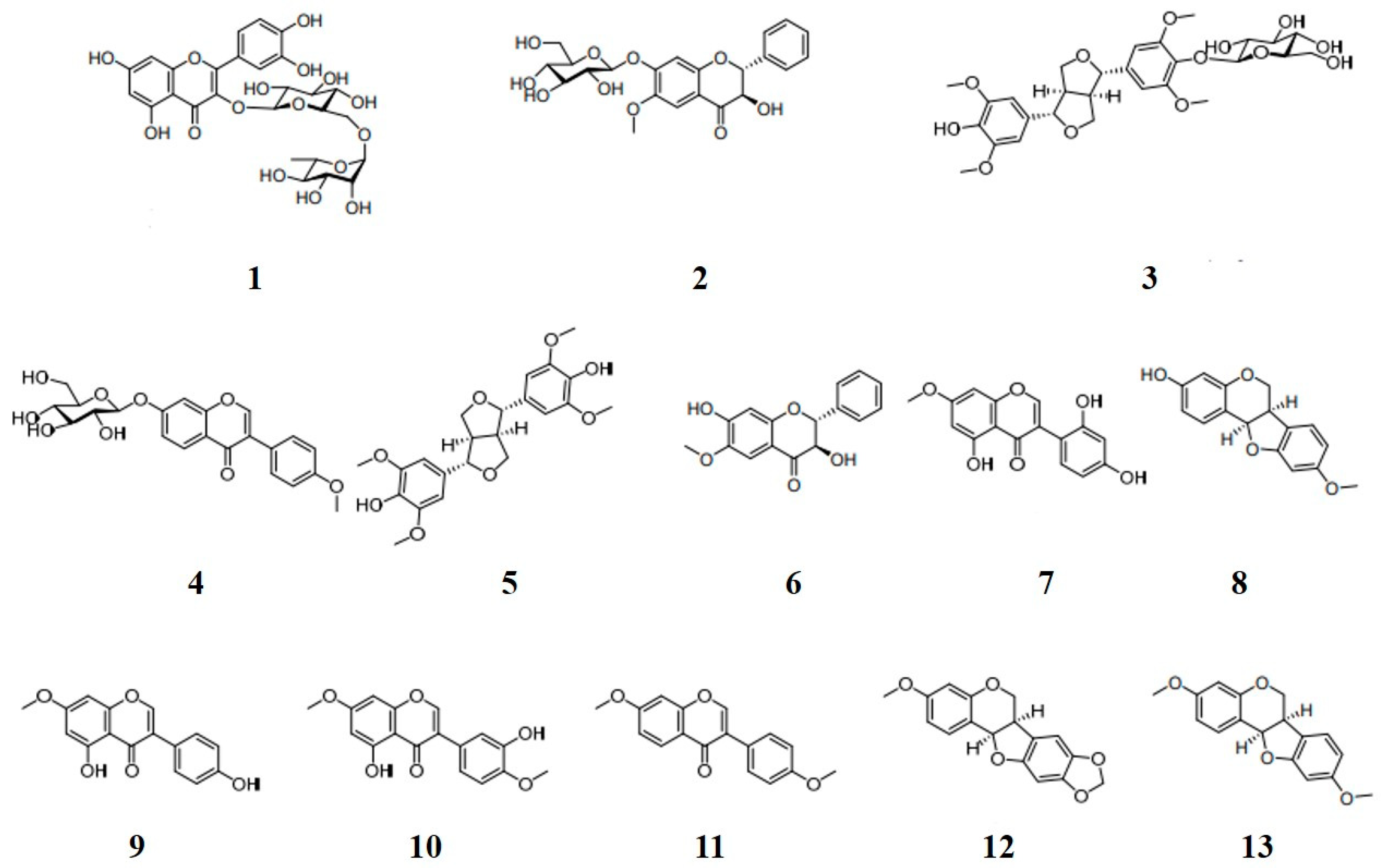
2.1. Preparation and Identification of Compounds 1–13
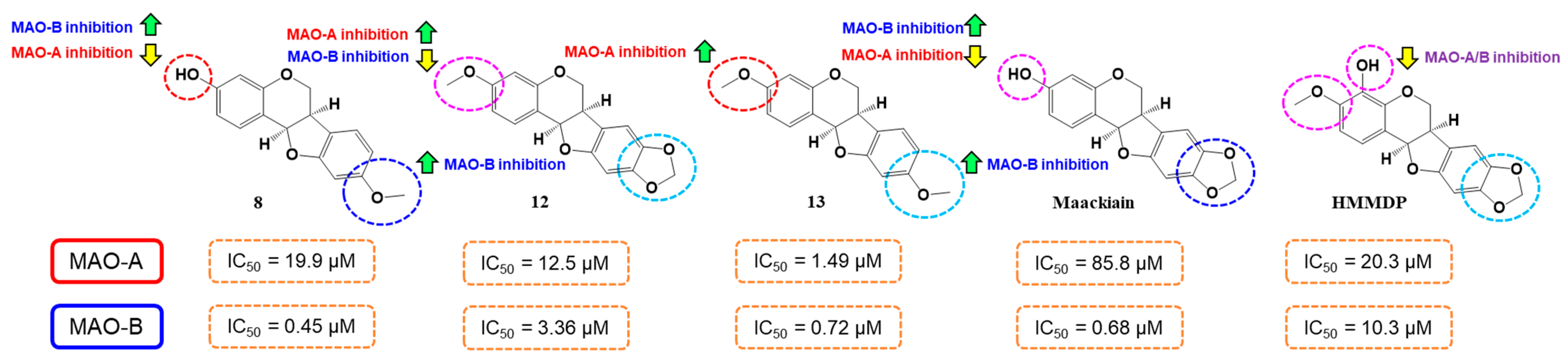
2.2. MAO Inhibitory Activities
2.3. Analysis of the Reversibility of hMAO-A and hMAO-B Inhibitions
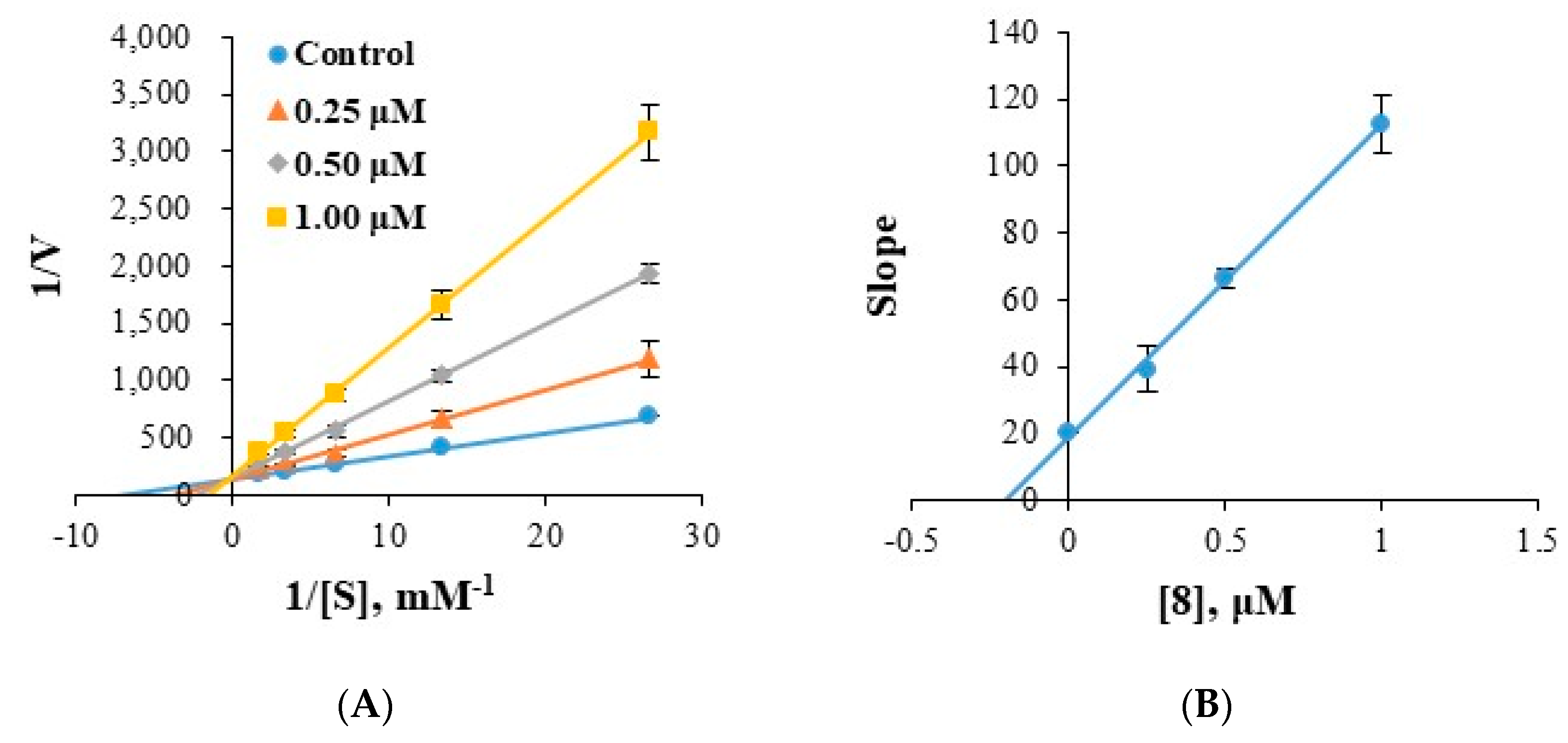
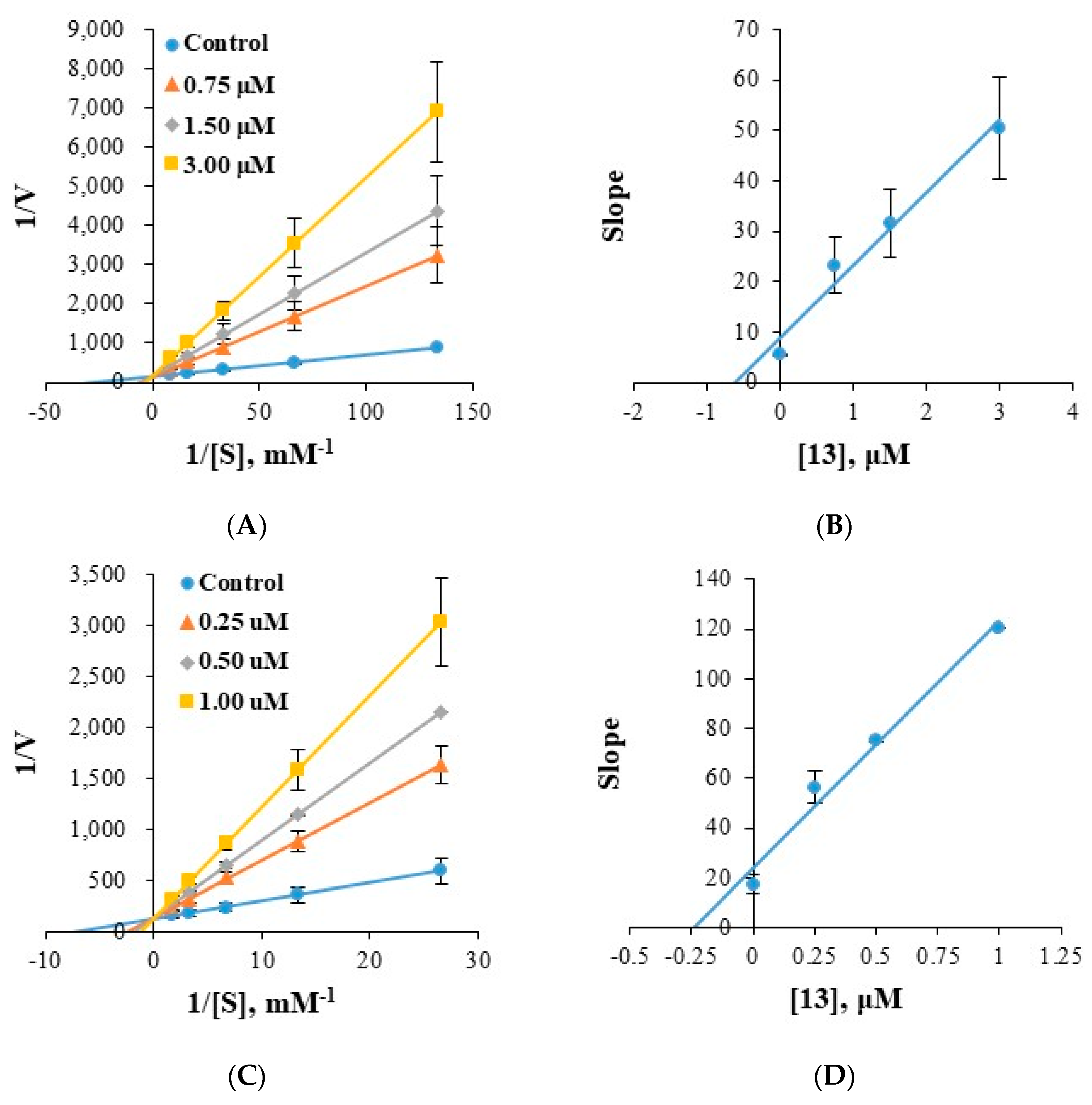
2.4. Analysis of Inhibitory Patterns
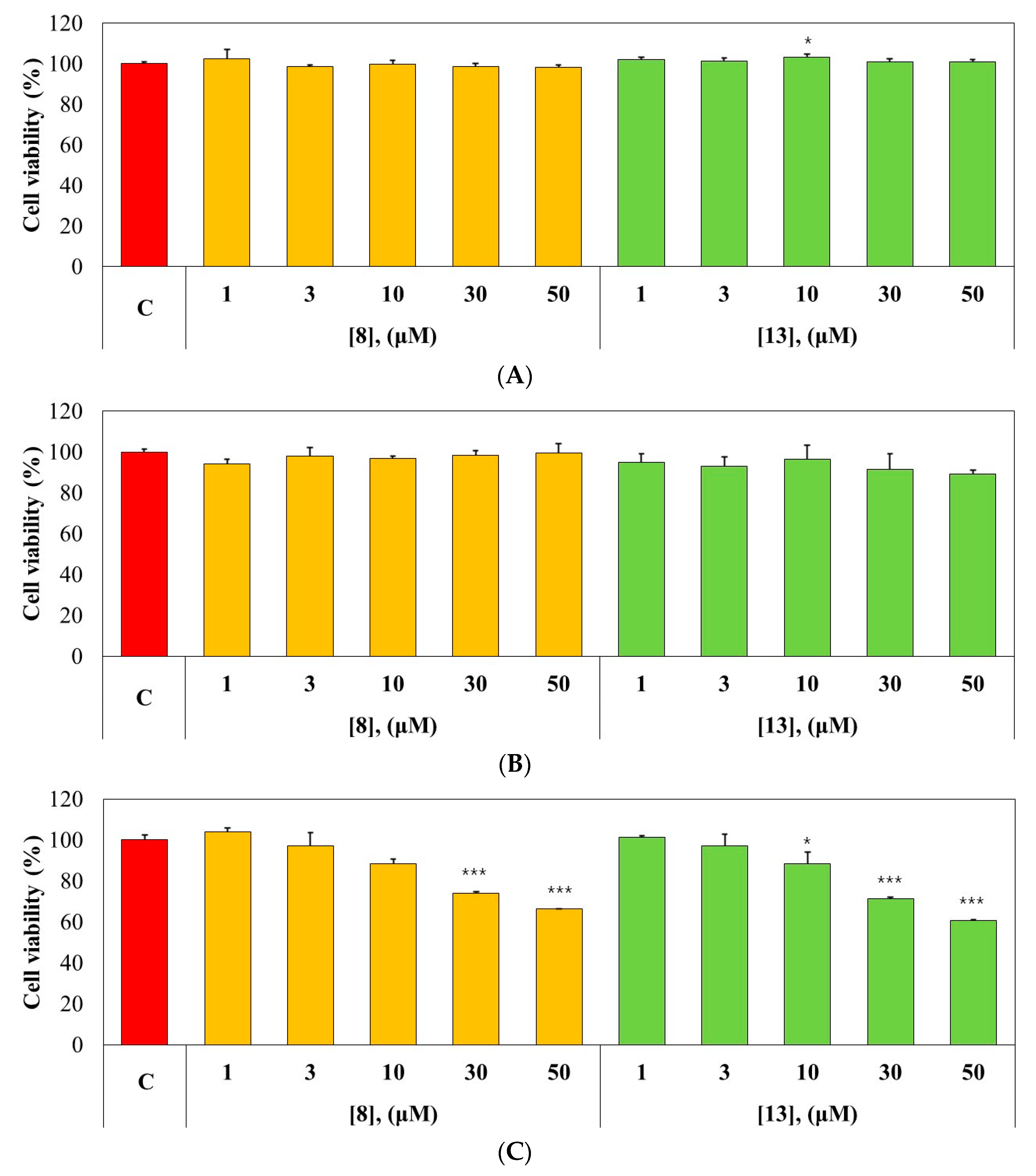
2.5. Cytotoxicity Test
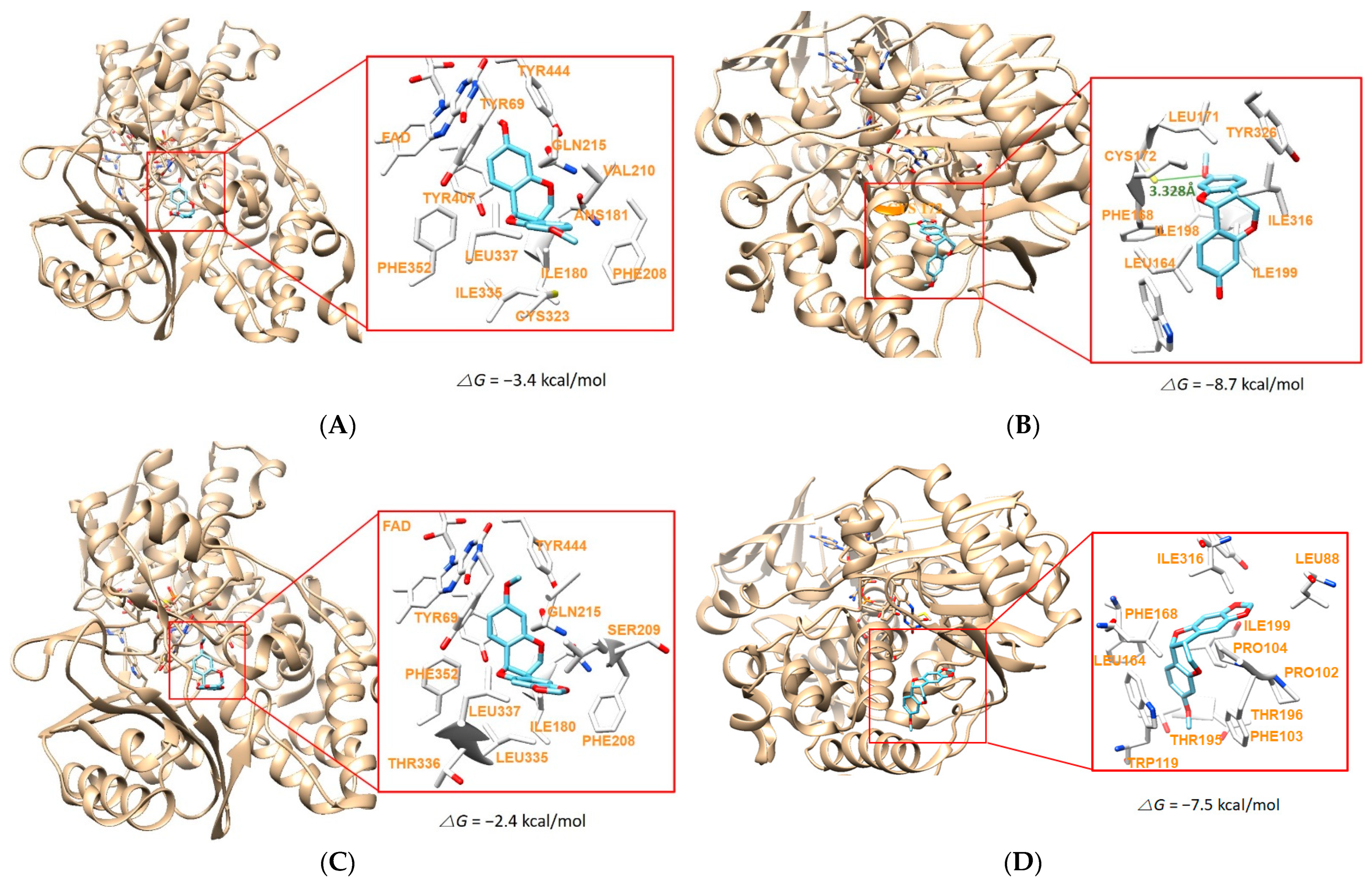
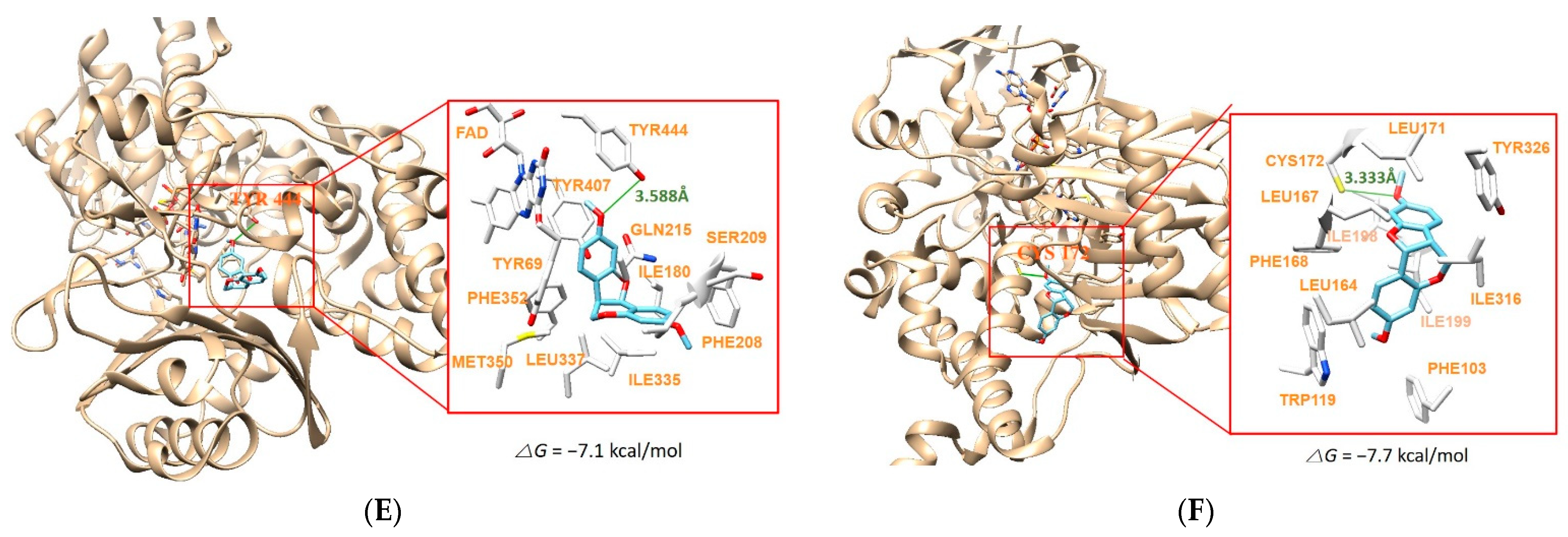
2.6. Molecular Docking Simulation
2.7. In Silico Pharmacokinetics of Medicarpin (8) and Homopterocarpin (13)
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Plant Materials and Preparation of Compounds 1–13
3.3. Chemicals and Enzymes
3.4. Enzyme Assays
3.5. Kinetics of Enzyme Inhibition
3.6. Analysis of Inhibitor Reversibility
3.7. Cytotoxicity
3.7.1. Reagents and Cell Lines
3.7.2. Cell Viability
3.8. Docking Simulations of the Compounds with MAO-A and MAO-B
3.9. Pharmacokinetic Analysis Using In Silico Method
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Sample Availability
References
- Barbosa, A.F.; Voos, M.C.; Chen, J.; Francato, D.C.V.; Souza, C.D.O.; Barbosa, E.R.; Chien, H.F.; Mansur, L.L. Cognitive or Cognitive-Motor Executive Function Tasks? Evaluating Verbal Fluency Measures in People with Parkinson’s Disease. BioMed Res. Int. 2017, 2017, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marino, B.; de Souza, L.R.; Sousa, K.; Ferreira, J.V.; Padilha, E.C.; da Silva, C.; Taft, C.A.; Hage-Melim, L. Parkinson’s disease: A review from pathophysiology to treatment. Mini Rev. Med. Chem. 2020, 20, 754–767. [Google Scholar] [CrossRef] [PubMed]
- Masato, A.; Plotegher, N.; Boassa, D.; Bubacco, L. Impaired dopamine metabolism in Parkinson’s disease pathogenesis. Mol. Neurodegener. 2019, 14, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khan, S.; Barve, K.H.; Kumar, M.S. Recent advancements in pathogenesis, diagnostics and treatment of Alzheimer’s disease. Curr. Neuropharmacol. 2020, 18, 1106–1125. [Google Scholar] [CrossRef] [PubMed]
- Cao, Z.; Song, Q.; Yu, G.; Liu, Z.; Cong, S.; Tan, Z.; Deng, Y. Novel 3-benzylidene/benzylphthalide mannich base derivatives as potential multifunctional agents for the treatment of Alzheimer’s disease. Bioorg. Med. Chem. 2021, 35, 116074. [Google Scholar] [CrossRef]
- Behl, T.; Kaur, D.; Sehgal, A.; Singh, S.; Sharma, N.; Zengin, G.; Andronie-Cioara, F.L.; Toma, M.M.; Bungau, S.; Bumbu, A.G. Role of monoamine oxidase activity in Alzheimer’s disease: An insight into the therapeutic potential of inhibitors. Molecules 2021, 26, 3724. [Google Scholar] [CrossRef]
- Al Mamun, A.; Uddin, M.S. KDS2010: A potent highly selective and reversible MAO-B inhibitor for Alzheimer’s disease. Comb. Chem. High. Throughput. Screen. 2020, 23, 836–841. [Google Scholar] [CrossRef]
- Hampel, H.; Vassar, R.; De Strooper, B.; Hardy, J.; Willem, M.; Singh, N.; Zhou, J.; Yan, R.; Vanmechelen, E.; De Vos, A.; et al. The β-secretase BACE1 in Alzheimer’s disease. Biol. Psychiatry 2021, 89, 745–756. [Google Scholar] [CrossRef]
- Ali, S.; Asad, M.; Maity, S.; Zada, W.; Rizvanov, A.A.; Iqbal, J.; Babak, B.; Hussain, I. Fluoro-benzimidazole derivatives to cure Alzheimer’s disease: In-silico studies, synthesis, structure-activity relationship and in vivo evaluation for β secretase enzyme inhibition. Bioorg. Chem. 2019, 88, 102936. [Google Scholar] [CrossRef]
- Ramsay, R.R.; Albreht, A. Kinetics, mechanism, and inhibition of monoamine oxidase. J. Neural Transm. 2018, 125, 1659–1683. [Google Scholar] [CrossRef]
- Özdemir, Z.; Alagöz, M.A.; Bahçecioğlu, Ö.F.; Gök, S. Monoamine oxidase-B (MAO-B) inhibitors in the treatment of Alzheimer’s and Parkinson’s disease. Curr. Med. Chem. 2021, 28, 6045–6065. [Google Scholar] [CrossRef] [PubMed]
- Finberg, J.P.; Youdim, M.B. Selective MAO A and B inhibitors: Their mechanism of action and pharmacology. Neuropharmacology 1983, 22, 441–446. [Google Scholar] [CrossRef] [PubMed]
- Schedin-Weiss, S.; Inoue, M.; Hromadkova, L.; Teranishi, Y.; Yamamoto, N.G.; Wiehager, B.; Bogdanovic, N.; Winblad, B.; Sandebring-Matton, A.; Frykman, S.; et al. Monoamine oxidase B is elevated in Alzheimer disease neurons, is associated with γ-secretase and regulates neuronal amyloid β-peptide levels. Alzheimers Res. Ther. 2017, 9, 57. [Google Scholar] [CrossRef] [Green Version]
- Youdim, M.B.H. Monoamine oxidase inhibitors, and iron chelators in depressive illness and neurodegenerative diseases. J. Neural Transm. 2018, 125, 1719–1733. [Google Scholar] [CrossRef] [PubMed]
- Carradori, S.; D’Ascenzio, M.; Chimenti, P.; Secci, D.; Bolasco, A. Selective MAO-B inhibitors: A lesson from natural products. Mol. Divers. 2014, 18, 219–243. [Google Scholar] [CrossRef]
- Tripathi, R.K.P.; Ayyannan, S.R. Monoamine oxidase-B inhibitors as potential neurotherapeutic agents: An overview and update. Med. Res. Rev. 2019, 39, 1603–1706. [Google Scholar] [CrossRef]
- Guglielmi, P.; Carradori, S.; Ammazzalorso, A.; Secci, D. Novel approaches to the discovery of selective human monoamine oxidase-B inhibitors: Is there room for improvement? Expert Opin. Drug Discov. 2019, 14, 995–1035. [Google Scholar] [CrossRef]
- Chavarria, D.; Fernandes, C.; Silva, V.; Silva, C.; Gil-Martins, E.; Soares, P.; Silva, T.; Silva, R.; Remião, F.; Oliveira, P.J.; et al. Design of novel monoamine oxidase-B inhibitors based on piperine scaffold: Structure-activity-toxicity, drug-likeness and efflux transport studies. Eur. J. Med. Chem. 2020, 185, 111770. [Google Scholar] [CrossRef]
- Subaraja, M.; Vanisree, A.J. The novel phytocomponent asiaticoside-D isolated from Centella asiatica exhibits monoamine oxidase-B inhibiting potential in the rotenone degenerated cerebral ganglions of Lumbricus terrestris. Phytomedicine 2019, 58, 152833. [Google Scholar] [CrossRef]
- Anand, P.; Singh, B. A review on cholinesterase inhibitors for Alzheimer’s disease. Arch. Pharm. Res. 2013, 36, 375–399. [Google Scholar] [CrossRef]
- Darvesh, S.; Hopkins, D.A.; Geula, C. Neurobiology of butyrylcholinesterase. Nature reviews. Neuroscience 2003, 4, 131–138. [Google Scholar] [PubMed]
- Mesulam, M.; Guillozet, A.; Shaw, P.; Quinn, B. Widely spread butyrylcholinesterase can hydrolyze acetylcholine in the normal and Alzheimer brain. Neurobiol. Dis. 2002, 9, 88–93. [Google Scholar] [CrossRef] [Green Version]
- Darvesh, S. Butyrylcholinesterase as a diagnostic and therapeutic target for Alzheimer’s disease. Curr. Alzheimer Res. 2006, 13, 1173–1177. [Google Scholar] [CrossRef]
- Moussa-Pacha, N.M.; Abdin, S.M.; Omar, H.A.; Alniss, H.; Al-Tel, T.H. BACE1 inhibitors: Current status and future directions in treating Alzheimer’s disease. Med. Res. Rev. 2020, 40, 339–384. [Google Scholar] [CrossRef] [PubMed]
- Sevigny, J.; Chiao, P.; Bussière, T.; Weinreb, P.H.; Williams, L.; Maier, M.; Dunstan, R.; Salloway, S.; Chen, T.; Ling, Y.; et al. The antibody aducanumab reduces Aβ plaques in Alzheimer’s disease. Nature 2016, 537, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, M.M.; Gabr, M.T. Multitarget therapeutic strategies for Alzheimer’s disease. Neural. Regen. Res. 2019, 14, 437–440. [Google Scholar]
- Chowdhury, S.; Kumar, S. Inhibition of BACE1, MAO-B, cholinesterase enzymes, and anti-amyloidogenic potential of selected natural phytoconstituents: Multi-target-directed ligand approach. J. Food Biochem. 2021, 45, e13571. [Google Scholar] [CrossRef]
- Ramsay, R.R.; Tipton, K.F. Assessment of enzyme inhibition: A review with examples from the development of monoamine oxidase and cholinesterase inhibitory drugs. Molecules 2017, 22, 1192. [Google Scholar] [CrossRef] [Green Version]
- Mathew, B.; Parambi, D.G.T.; Mathew, G.E.; Uddin, M.S.; Inasu, S.T.; Kim, H.; Marathakam, A.; Unnikrishnan, M.K.; Carradori, S. Emerging therapeutic potentials of dual-acting MAO and AChE inhibitors in Alzheimer’s and Parkinson’s diseases. Arch. Pharm. 2019, 352, e1900177. [Google Scholar] [CrossRef]
- Baek, S.C.; Park, M.H.; Ryu, H.W.; Lee, J.P.; Kang, M.-G.; Park, D.; Park, C.M.; Oh, S.-R.; Kim, H. Rhamnocitrin isolated from Prunus padus var. seoulensis: A potent and selective reversible inhibitor of human monoamine oxidase A. Bioorg. Chem. 2019, 83, 317–325. [Google Scholar]
- Oh, J.M.; Jang, H.-J.; Kim, W.J.; Kang, M.-G.; Baek, S.C.; Lee, J.P.; Park, D.; Oh, S.-R.; Kim, H. Calycosin and 8-O-methylretusin isolated from Maackia amurensis as potent and selective reversible inhibitors of human monoamine oxidase-B. Int. J. Biol. Macromol. 2020, 151, 441–448. [Google Scholar] [CrossRef]
- Oh, J.M.; Jang, H.J.; Kang, M.G.; Song, S.; Kim, D.Y.; Kim, J.H.; Noh, J.I.; Park, J.E.; Park, D.; Yee, S.T.; et al. Acetylcholinesterase and monoamine oxidase-B inhibitory activities by ellagic acid derivatives isolated from Castanopsis cuspidata var. sieboldii. Sci. Rep. 2021, 11, 13953. [Google Scholar] [CrossRef] [PubMed]
- Bu, H.-J.; Lee, H.-J.; Yoo, E.-S.; Jung, D.-S.; Riu, K.-Z.; Lee, S.-J. Antioxidant effects and inhibitory effect on NO synthesis by extracts of Canavalia lineata. Korean J. Pharmacogn. 2004, 35, 338–345. [Google Scholar]
- Hong, S.-J.; Kwon, O.-K.; Hwang, D.; Goo, S.H.; Kim, D.-Y.; Kim, M.H.; Kim, S.-Y.; Jang, H.-J.; Oh, S.-R. Anti-inflammatory activity of Cajanin, an isoflavonoid derivative isolated from Canavalia lineata pods. Int. J. Mol. Sci. 2022, 23, 9492. [Google Scholar] [CrossRef] [PubMed]
- Terada, S.; Fujimura, S.; Kino, S.; Kimoto, E. Purification and characterization of three proteinase inhibitors from Canavalia lineata seeds. Biosci. Biotechnol. Biochem. 1994, 58, 371–375. [Google Scholar] [CrossRef]
- Lee, H.W.; Ryu, H.W.; Kang, M.-G.; Park, D.; Oh, S.-R.; Kim, H. Potent selective monoamine oxidase B inhibition by maackiain, a pterocarpan from the roots of Sophora flavescens. Bioorg. Med. Chem. Lett. 2016, 26, 4714–4719. [Google Scholar] [CrossRef]
- Lee, H.W.; Ryu, H.W.; Baek, S.C.; Kang, M.-G.; Park, D.; Han, H.-Y.; An, J.H.; Oh, S.-R.; Kim, H. Potent inhibitions of monoamine oxidase A and B by acacetin and its 7-O-(6-O-malonylglucoside) derivative from Agastache rugosa. Int. J. Biol. Macromol. 2017, 104, 547–553. [Google Scholar] [CrossRef]
- Chaurasiya, N.D.; Zhao, J.; Pandey, P.; Doerksen, R.J.; Muhammad, I.; Tekwani, B.L. Selective inhibition of human monoamine oxidase B by acacetin 7-methyl ether isolated from Turnera diffusa (Damiana). Molecules 2019, 24, 810. [Google Scholar] [CrossRef] [Green Version]
- Larit, F.; Elokely, K.M.; Chaurasiya, N.D.; Benyahia, S.; Nael, M.A.; León, F.; Abu-Darwish, M.S.; Efferth, T.; Wang, Y.-H.; Belouahem-Abed, D.; et al. Inhibition of human monoamine oxidase A and B by flavonoids isolated from two Algerian medicinal plants. Phytomedicine 2018, 40, 27–36. [Google Scholar] [CrossRef]
- Li, D.; Cai, C.; Liao, Y.; Wu, Q.; Ke, H.; Guo, P.; Wang, Q.; Ding, B.; Fang, J.; Fang, S. Systems pharmacology approach uncovers the therapeutic mechanism of medicarpin against scopolamine-induced memory loss. Phytomedicine 2021, 91, 153662. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, D.M.; Cho, Y.J.; Hyun, J.W.; Ahn, M.J. Medicarpin increases antioxidant genes by inducing NRF2 transcriptional level in HeLa cells. Antioxidants 2022, 11, 421. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Yang, R.; Yan, F.; Jin, Y.; Liu, X.; Wang, T. Medicarpin protects cerebral microvascular endothelial cells against oxygen-glucose deprivation/reoxygenation-induced injury via the PI3K/Akt/FoxO pathway: A study of network pharmacology analysis and experimental validation. Neurochem. Res. 2022, 47, 347–357. [Google Scholar] [CrossRef] [PubMed]
- Gatouillat, G.; Magid, A.A.; Bertin, E.; El Btaouri, H.; Morjani, H.; Lavaud, C.; Madoulet, C. Medicarpin and millepurpan, two flavonoids isolated from Medicago sativa, induce apoptosis and overcome multidrug resistance in leukemia P388 cells. Phytomedicine 2015, 22, 1186–1194. [Google Scholar] [CrossRef] [PubMed]
- Militão, G.C.; Dantas, I.N.; Pessoa, C.; Falcão, M.J.; Silveira, E.R.; Lima, M.A.; Curi, R.; Lima, T.; Moraes, M.O.; Costa-Lotufo, L.V. Induction of apoptosis by pterocarpans from Platymiscium floribundum in HL-60 human leukemia cells. Life Sci. 2006, 78, 2409–2417. [Google Scholar] [CrossRef]
- Akinmoladun, A.C.; Olaleye, M.T.; Komolafe, K.; Adetuyi, A.O.; Akindahunsi, A.A. Effect of homopterocarpin, an isoflavonoid from Pterocarpus erinaceus, on indices of liver injury and oxidative stress in acetaminophen-provoked hepatotoxicity. J. Basic Clin. Physiol. Pharmacol. 2015, 26, 555–562. [Google Scholar] [CrossRef]
- Mathew, B.; Baek, S.C.; Thomas Parambi, D.G.; Lee, J.P.; Mathew, G.E.; Jayanthi, S.; Vinod, D.; Rapheal, C.; Devikrishna, V.; Kondarath, S.S.; et al. Potent and highly selective dual-targeting monoamine oxidase-B inhibitors: Fluorinated chalcones of morpholine versus imidazole. Arch. Pharm. 2019, 352, e1800309. [Google Scholar] [CrossRef]
- Mathew, G.E.; Oh, J.M.; Mohan, K.; Kumudhavalli, M.V.; Jayanthi, S.; Kim, H. Inhibitions of monoamine oxidases and acetylcholinesterase by 1-mehtyl, 5-phenyl substituted thiosemicarbazones: Synthesis, biochemical, and computational investigations. Process Biochem. 2020, 99, 246–253. [Google Scholar] [CrossRef]
- Ramsay, R.R.; Majekova, M.; Medina, M.; Valoti, M. Key targets for multi-target ligands designed to combat neurodegeneration. Front. Neurosci. 2016, 10, 375. [Google Scholar] [CrossRef] [Green Version]
- Oh, J.M.; Kang, M.-G.; Hong, A.; Park, J.-E.; Kim, S.H.; Lee, J.P.; Baek, S.C.; Park, D.; Nam, S.-J.; Cho, M.-L.; et al. Potent and selective inhibition of human monoamine oxidase-B by 4-dimethylaminochalcone and selected chalcone derivatives. Int. J. Biol. Macromol. 2019, 137, 426–432. [Google Scholar] [CrossRef]
- Flockhart, D.A. Drug Interactions: Cytochrome P450 Drug Interaction Table. Indiana University School of Medicine, 2007. Available online: http://medicine.iupui.edu/flockhart/table.htm (accessed on 22 September 2008).
- Jang, H.-J.; Kim, W.J.; Lee, S.U.; Kim, M.-O.; Park, M.H.; Song, S.; Kim, D.-Y.; Lee, S.M.; Yuk, H.J.; Lee, D.Y.; et al. Optimization of chiisanoside and chiisanogenin isolation from Eleutherococcus sessiliflorus (Rupr. & Maxim.) leaves for industrial application: A pilot study. Ind. Crops Prod. 2022, 185, 115099. [Google Scholar]
- Baek, S.C.; Lee, H.W.; Ryu, H.W.; Kang, M.-G.; Park, D.; Kim, S.H.; Cho, M.-L.; Oh, S.-R.; Kim, H. Selective inhibition of monoamine oxidase A by hispidol. Bioorg. Med. Chem. Lett. 2018, 28, 584–588. [Google Scholar] [CrossRef] [PubMed]
- Baek, S.C.; Choi, B.; Nam, S.-J.; Kim, H. Inhibition of monoamine oxidase A and B by demethoxycurcumin and bisdemethoxycurcumin. J. Appl. Biol. Chem. 2018, 61, 187–190. [Google Scholar] [CrossRef]
- Baek, S.C.; Ryu, H.W.; Kang, M.-G.; Lee, H.; Park, D.; Cho, M.-L.; Oh, S.-R.; Kim, H. Selective inhibition of monoamine oxidase A by chelerythrine, an isoquinoline alkaloid. Bioorg. Med. Chem. Lett. 2018, 28, 2403–2407. [Google Scholar] [CrossRef] [PubMed]
- Ellman, G.L.; Courtney, K.D.; Andres, V.; Feather-Stone, R.M. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem. Pharmacol. 1961, 7, 88–95. [Google Scholar] [CrossRef]
- Lee, J.P.; Kang, M.-G.; Lee, J.Y.; Oh, J.M.; Baek, S.C.; Leem, H.H.; Park, D.; Cho, M.-L.; Kim, H. Potent inhibition of acetylcholinesterase by sargachromanol I from Sargassum siliquastrum and by selected natural compounds. Bioorg. Chem. 2019, 89, 103043. [Google Scholar] [CrossRef] [PubMed]
- Jeong, G.S.; Kang, M.-G.; Lee, J.Y.; Lee, S.R.; Park, D.; Cho, M.; Kim, H. Inhibition of butyrylcholinesterase and human monoamine oxidase-B by the coumarin glycyrol and liquiritigenin isolated from Glycyrrhiza uralensis. Molecules 2020, 25, 3896. [Google Scholar] [CrossRef]
- Noh, J.-I.; Mun, S.-K.; Lim, E.H.; Kim, H.; Chang, D.-J.; Hur, J.-S.; Yee, T. Induction of apoptosis in MDA-MB-231 cells treated with the methanol extract of lichen Physconia hokkaidensis. J. Fungi. 2021, 7, 188. [Google Scholar] [CrossRef]
- Park, J.E.; Mun, S.-K.; Yee, S.-T.; Kim, H. Evaluation of inhibitory activities of Sophora flavescens and Angelica gigas Nakai root extracts against monoamine oxidases, cholinesterases, and β-Secretase. Processes 2022, 10, 880. [Google Scholar] [CrossRef]
- Trott, O.; Olson, A.J. Auto Dock Vina: Improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar]
- Mills, J.E.; Dean, P.M. Three-dimensional hydrogen-bond geometry and probability information from a crystal survey. J. Comput. Aided Mol. Des. 1996, 10, 607–622. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera--a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heo, J.H.; Eom, B.H.; Ryu, H.W.; Kang, M.G.; Park, J.E.; Kim, D.Y.; Kim, J.H.; Park, D.; Oh, S.R.; Kim, H. Acetylcholinesterase and butyrylcholinesterase inhibitory activities of khellactone coumarin derivatives isolated from Peucedanum japonicum Thurnberg. Sci. Rep. 2020, 10, 21695. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Extract/Fraction | Residual Activity at 25 µg/mL (%) | ||||
|---|---|---|---|---|---|
| hMAO-A | hMAO-B | AChE | BChE | BACE1 | |
| MeOH | 76.1 ± 3.07 | 53.5 ± 8.60 | 78.2 ± 3.01 | 98.9 ± 0.50 | 76.1 ± 3.07 |
| EtOAc | 60.9 ± 1.54 | 41.4 ± 2.86 | 64.9 ± 0.75 | 99.6 ± 0.50 | 60.9 ± 1.54 |
| BuOH | 90.8 ± 0.77 | 85.9 ± 1.43 | 72.9 ± 5.27 | 97.9 ± 1.00 | 90.8 ± 0.77 |
| Compound | Residual Activity at 10 µM (%) | IC50 (µM) | SI b | |||||
|---|---|---|---|---|---|---|---|---|
| hMAO-A | hMAO-B | AChE | BChE | BACE1 | hMAO-A | hMAO-B | ||
| 1 | 96.79 ± 1.52 | 75.22 ± 6.76 | 92.4 ± 7.16 | 99.2 ± 0.88 | 82.56 ± 3.77 | >40 | >40 | - |
| 2 | 98.28 ± 2.01 | 69.57 ± 2.46 | 84.8 ± 0.00 | 97.5 ± 0.42 | 163.67 ± 2.96 | >40 | >40 | - |
| 3 | 80.00 ± 2.02 | 93.63 ± 3.47 | 95.22 ± 0.08 | 98.91 ± 1.54 | 76.68 ± 4.92 | >40 | >40 | - |
| 4 | 98.28± 1.64 | 63.48 ± 1.23 | 86.1 ± 1.79 | 96.7 ± 1.68 | 85.90 ± 0.08 | >40 | 28.5 ± 1.55 | >1.40 |
| 5 | 80.36 ± 6.57 | 63.24 ± 6.24 | 84.49 ± 0.75 | 99.41 ± 0.83 | 81.70 ± 4.01 | >40 | 24.32 ± 2.20 | >1.64 |
| 6 | 94.83 ± 1.90 | 94.64 ± 4.21 | 84.3 ± 1.31 | 91.8 ± 1.35 | 95.72 ± 0.86 | >40 | >40 | - |
| 7 | 56.90 ± 0.92 | 31.55 ± 4.21 | 70.8 ± 7.20 | 86.9 ± 2.11 | 83.25 ± 0.45 | 13.1 ± 0.23 | 5.18 ± 0.11 | 2.53 |
| 8 | 64.18 ± 0.53 | 4.17 ± 0.84 | 80.6 ± 2.62 | 98.4 ± 1.00 | 111.42 ± 0.52 | 19.9 ± 0.85 | 0.45 ± 0.032 | 44.2 |
| 9 | 24.75 ± 1.22 | 30.81 ± 0.82 | 83.2 ± 4.76 | 93.0 ± 0.65 | 87.73 ± 0.59 | 2.49 ± 0.32 | 5.18 ± 0.059 | 0.48 |
| 10 | 41.02 ± 1.09 | 16.86 ± 2.47 | 78.4 ± 3.40 | 98.1 ± 1.98 | 78.86 ± 0.56 | 8.05 ± 0.10 | 3.47 ± 0.055 | 2.32 |
| 11 | 49.15 ± 2.25 | 62.02 ± 1.09 | 82.7 ± 5.44 | 98.7 ± 0.66 | 91.96 ± 0.59 | 9.80 ± 0.27 | 13.7 ± 0.10 | 0.72 |
| 12 | 60.34 ± 0.90 | 13.01 ± 0.72 | 84.9 ± 0.61 | 93.6 ± 2.11 | 102.36 ± 0.40 | 12.5 ± 0.33 | 3.36 ± 0.090 | 3.72 |
| 13 | 11.59 ± 2.60 | 4.05 ± 1.91 | 81.0 ± 2.44 | 88.5 ± 1.96 | 101.48 ± 0.01 | 1.49 ± 0.021 | 0.72 ± 0.028 | 2.07 |
| Toloxatone | - | - | - | 1.080 ± 0.025 | - | |||
| Lazabemide | - | - | - | - | 0.110 ± 0.016 | |||
| Clorgyline | - | - | - | 0.007 ± 0.001 | - | |||
| Pargyline | - | - | - | - | 0.140 ± 0.006 | |||
| Tacrine * | 0.270 ± 0.019 | 0.060 ± 0.0022 | - | - | - | |||
| Donepezil * | 0.010 ± 0.002 | 0.180 ± 0.0038 | - | - | - | |||
| Quercetin * | - | - | 13.420 ± 0.035 | - | - | |||
| Inhibitor IV * | 0.440 ± 0.064 | |||||||
| Compound | Docking Energy (kcal/mol) | H-Bond Predicted | ||
|---|---|---|---|---|
| hMAO-A | hMAO-B | hMAO-A | hMAO-B | |
| 8 | −3.4 | −8.7 | Cys172 (3.328 Å) | |
| 12 | −2.4 | −7.5 | ||
| 13 | −7.1 | −7.7 | Tyr444 (3.588 Å) | Cys172 (3.333 Å) |
| Compound | GI Absorption | BBB Permeant | P-gp Substrate | Inhibitor | Log Kp | ||||
|---|---|---|---|---|---|---|---|---|---|
| CYP1A2 | CYP2C19 | CYP2C9 | CYP2D6 | CYP3A4 | Skin Permeation (cm/s) | ||||
| 8 | High | Y | Y | Y | Y | N | Y | Y | −5.98 |
| 13 | High | Y | Y | Y | Y | N | Y | Y | −5.84 |
| Compound | Mw (g/mol) | cLog P | HBD | HBA | TPSA (Å2) | RB | Lipinski Violations |
|---|---|---|---|---|---|---|---|
| 8 | 270.28 | 2.53 | 1 | 4 | 47.92 | 1 | 0 |
| 13 | 284.31 | 2.91 | 0 | 4 | 36.92 | 2 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oh, J.M.; Jang, H.-J.; Kang, M.-G.; Mun, S.-K.; Park, D.; Hong, S.-J.; Kim, M.H.; Kim, S.-Y.; Yee, S.-T.; Kim, H. Medicarpin and Homopterocarpin Isolated from Canavalia lineata as Potent and Competitive Reversible Inhibitors of Human Monoamine Oxidase-B. Molecules 2023, 28, 258. https://doi.org/10.3390/molecules28010258
Oh JM, Jang H-J, Kang M-G, Mun S-K, Park D, Hong S-J, Kim MH, Kim S-Y, Yee S-T, Kim H. Medicarpin and Homopterocarpin Isolated from Canavalia lineata as Potent and Competitive Reversible Inhibitors of Human Monoamine Oxidase-B. Molecules. 2023; 28(1):258. https://doi.org/10.3390/molecules28010258
Chicago/Turabian StyleOh, Jong Min, Hyun-Jae Jang, Myung-Gyun Kang, Seul-Ki Mun, Daeui Park, Su-Jin Hong, Min Ha Kim, Soo-Young Kim, Sung-Tae Yee, and Hoon Kim. 2023. "Medicarpin and Homopterocarpin Isolated from Canavalia lineata as Potent and Competitive Reversible Inhibitors of Human Monoamine Oxidase-B" Molecules 28, no. 1: 258. https://doi.org/10.3390/molecules28010258