
Enzymatic Conversion of Different Qualities of Refined Softwood Hemicellulose Recovered from Spent Sulfite Liquor

 , , , and
, , , and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Characterization of Acetyl-Galactoglucomannan from Spent Sulfite Liquor
2.1.1. Compositional Analysis
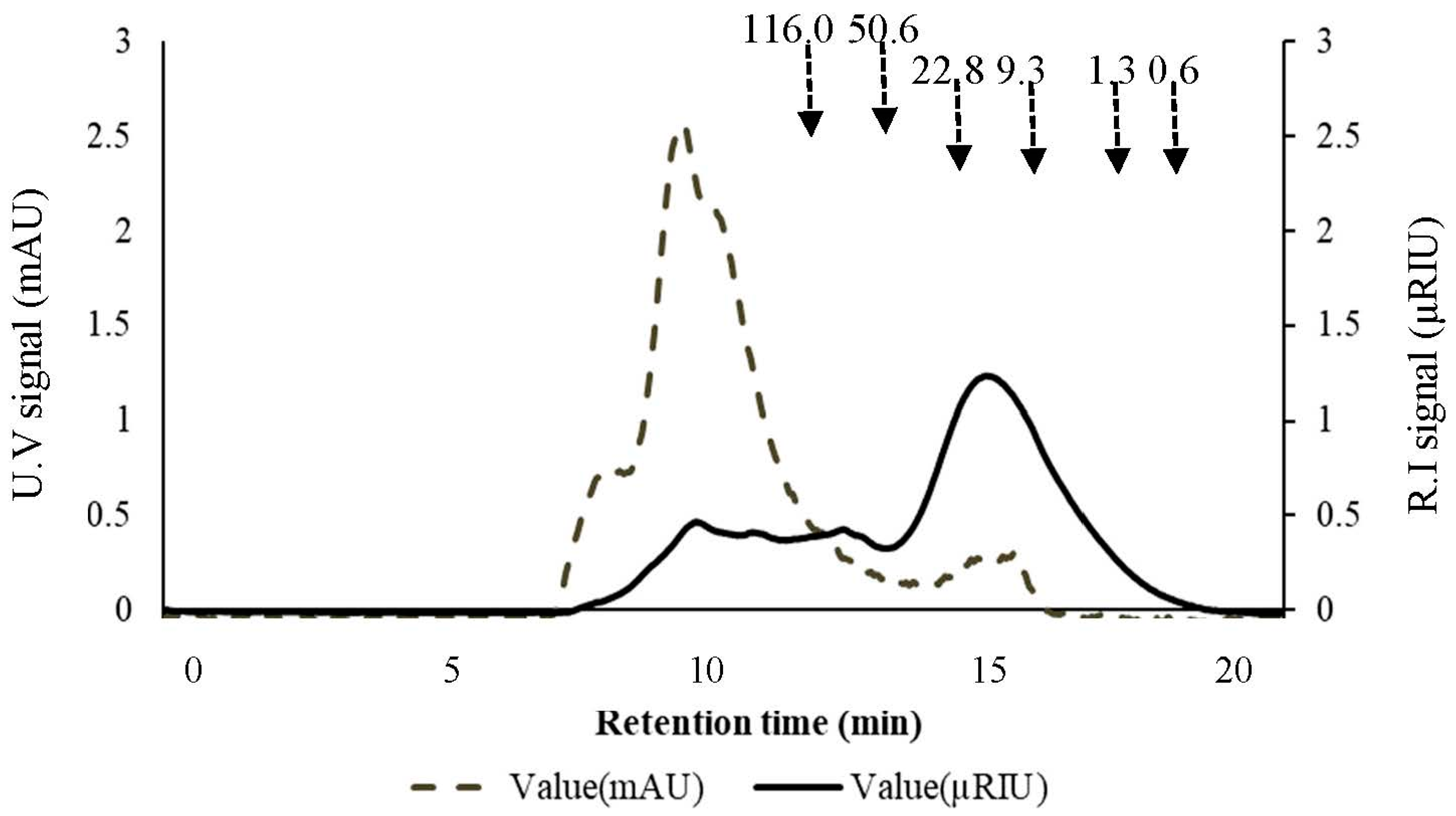
2.1.2. Apparent Molecular Weight of Acetyl-Galactoglucomannan in the Preparations
2.2. Selection of Commercial Enzyme Cocktails
2.2.1. Determining the Hydrolytic Efficiency of Commercial Enzyme Cocktails
2.2.2. Enzyme Activities in Commercial Enzyme Cocktails and Enzyme Stability
2.3. Effect of Purification on Hydrolysis of Acetylgalactoglucomannan from Spent Sulfite Liquor
2.3.1. Hydrolysis of Different Acetyl-Galactoglucomannan Preparations Using TrMan5A and Viscozyme L
2.3.2. Analysis of Major Hydrolysis Products Using High-Performance Anion-Exchange Chromatography with Pulsed Amperometric Detection (HPAEC-PAD)
2.3.3. Degree of Conversion of the SSL-AcGGM Preparations
2.4. Hydrolysis of Acetyl-Galactoglucomannan at Different Enzyme Loadings
2.5. Acetyl-Glactoglucomannan Biorefining and Future Prospectives
3. Materials and Methods
3.1. Materials
3.2. Purification of Acetyl Galactoglucomannan from Spent Sulfite Liquor (SSL-AcGGM)
3.2.1. Membrane Filtration
3.2.2. Antisolvent Precipitation
3.2.3. Adsorption
3.3. Characterization of the Material
3.3.1. Analysis of Acid-Insoluble Solids and Total Carbohydrate Content
3.3.2. Estimation of Lignin Content
3.3.3. Lyophilization and Resuspension of AcGGM Preparations
3.3.4. Acetyl Content Determination
3.3.5. Size Exclusion Chromatography
3.4. Enzymatic Conversion
3.4.1. Hydrolytic Efficiency of Commercial Enzyme Cocktails and TrMan5A
3.4.2. Enzyme Activity and Stability
3.4.3. Effect of Purification on Hydrolysis of Acetyl-Galactoglucomannan
3.4.4. Effect of Protein Loading on Enzymatic Hydrolysis of Acetyl-Galactoglucomannan
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Perea-Moreno, M.A.; Samerón-Manzano, E.; Perea-Moreno, A.J. Biomass as renewable energy: Worldwide research trends. Sustainability 2019, 11, 863. [Google Scholar] [CrossRef] [Green Version]
- Cherubini, F. The biorefinery concept: Using biomass instead of oil for producing energy and chemicals. Energy Convers. Manag. 2010, 51, 1412–1421. [Google Scholar] [CrossRef]
- Galbe, M.; Wallberg, O. Pretreatment for biorefineries: A review of common methods for efficient utilisation of lignocellulosic materials. Biotechnol. Biofuels 2019, 12, 294. [Google Scholar] [CrossRef] [Green Version]
- Johnson, M.A.; Hart, P.W. Integrating Biorefinery into an Operating Kraft Mill. BioResources 2016, 11, 10677–10710. [Google Scholar] [CrossRef] [Green Version]
- Soottitantawat, A.; Methaapanon, R.; Klaysom, C. 15-Key unit operations in biorefinery processes. In A-Z of Biorefinery; Thongchul, N., Kokossis, A., Assabumrungrat, S., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2022; pp. 523–555. ISBN 9780128192481. [Google Scholar] [CrossRef]
- Peng, X.; Ren, J.; Zhong, L.; Sun, R. Nanocomposite Films Based on Xylan-Rich Hemicelluloses and Cellulose Nanofibers with Enhanced Mechanical Properties. Biomacromolecules 2011, 12, 3321–3329. [Google Scholar] [CrossRef]
- Menon, V.; Rao, M. Trends in bioconversion of lignocellulose: Biofuels, platform chemicals & biorefinery concept. Prog. Energy Combust. Sci. 2012, 38, 522–550. [Google Scholar] [CrossRef]
- Ramesh, M.; Rajeshkumar, L.; Sasikala, G.; Balaji, D.; Saravanakumar, A.; Bhuvaneswari, V.; Bhoopathi, R. A critical review on wood-based polymer composites: Processing, properties and prospects. Polymers 2022, 14, 589. [Google Scholar] [CrossRef] [PubMed]
- Salmén, L. Wood morphology and properties from molecular perspectives. Ann. For. Sci. 2015, 72, 679–684. [Google Scholar] [CrossRef] [Green Version]
- Berglund, J.; Mikkelsen, A.; Flanagan, M.B.; Dhital, S.; Gaunitz, S.; Henriksson, G.; Lindstrom, M.E.; Yakubov, G.E.; Gidley, M.J.; Vilaplana, F. Wood hemicelluloses exert distinct biomechanical contributions to cellulose fibrillar networks. Nat. Commun. 2020, 11, 4692. [Google Scholar] [CrossRef] [PubMed]
- Alén, R. Pulp Mills and Wood-Based Biorefineries. In Industrial Biorefineries and White Biotechnology; Pandey, A., Höfer, R., Taherzadeh, M., Nampoothiri, M.K., Christian, L., Eds.; Elsevier B.V.: Amsterdam, The Netherlands, 2015; pp. 91–126. [Google Scholar] [CrossRef]
- Lundqvist, J.; Teleman, A.; Junel, L.; Zacchi, G.; Dahlman, O.; Tjerneld, F.; Stålbrand, H. Isolation and characterization of galactoglucomannan from spruce (Picea abies). Carbohydr. Polym. 2002, 8, 29–39. [Google Scholar] [CrossRef]
- Willför, S.; Sjöholm, R.; Laine, C.; Roslund, M.; Hemming, J.; Holmbom, B. Characterisation of water-soluble galactoglucomannans from Norway spruce wood and thermomechanical pulp. Carbohydr. Polym. 2003, 52, 175–187. [Google Scholar] [CrossRef]
- Mikkonen, K.S.; Heikkilä, M.I.; Helén, H.; Hyvönen, L.; Tenkanen, M. Spruce galactoglucomannan films show promising barrier properties. Carbohydr. Polym. 2010, 79, 1107–1112. [Google Scholar] [CrossRef]
- Thuvander, J.; Jönsson, A.S. Extraction of galactoglucomannan from thermomechanical pulp mill process water by microfiltration and ultrafiltration-Influence of microfiltration membrane pore size on ultrafiltration performance. Chem. Eng. Res. Des. 2016, 105, 171–176. [Google Scholar] [CrossRef]
- Al-Rudainy, B.; Galbe, M.; Schgerlof, H.; Wallberg, O. Antisolvent precipitation of hemicelluloses, lignosulfonates and their complexes from ultrafiltrated spent sulfite liquor. Holzforschung 2018, 72, 839–850. [Google Scholar] [CrossRef]
- Dax, D.; Eklund, P.; Hemming, J.; Sarfraz, J.; Backman, P.; Xu, C.; Willför, S. Amphiphilic spruce galactoglucomannan derivatives based on naturally-occurring fatty acids. BioResources 2013, 8, 3771–3790. [Google Scholar] [CrossRef]
- Weissgram, M.; Gstöttner, J.; Lorantfy, B.; Tenhaken, R.; Herwig, C.; Weber, H. Generation of PHB from Spent Sulfite Liquor Using Halophilic Microorganisms. Microorganisms 2015, 3, 268–289. [Google Scholar] [CrossRef] [Green Version]
- Ishii, J.; Okazaki, F.; Djohan, A.C.; Hara, K.Y.; Asai-Nakashima, N.; Teramura, H.; Andriani, A.; Tominaga, M.; Wakai, S.; Kahar, P.; et al. From mannan to bioethanol: Cell surface co-display of β-mannanase and β-mannosidase on yeast Saccharomyces cerevisiae. Biotechnol. Biofuels 2016, 9, 188. [Google Scholar] [CrossRef] [Green Version]
- Morrill, J.; Månberger, A.; Rosengren, A.; Naidjonoka, P.; von Freiesleben, P.; Krogh, K.B.R.M.; Bergquist, K.E.; Nylander, T.; Karlsson, E.N.; Adlercreutz, P.; et al. β-Mannanase-catalyzed synthesis of alkyl mannooligosides. Appl. Microbiol. Biotechnol. 2018, 102, 5149–5163. [Google Scholar] [CrossRef] [Green Version]
- Rosengren, A.; Butler, S.J.; Arcos-Hernandez, M.; Bergquist, K.E.; Jannasch, P.; Stålbrand, H. Enzymatic synthesis and polymerisation of β-mannosyl acrylates produced from renewable hemicellulosic glycans. Green Chem. 2019, 21, 2104–2118. [Google Scholar] [CrossRef] [Green Version]
- Butler, S.J.; Birgersson, S.; Wiemann, M.; Acros-Hernandez, M.; Stålbrand, H. Transglycosylation by β-mannanase TrMan5A variants and enzyme synergy for synthesis of allyl glycosides from galactomannan. Process Biochem. 2022, 112, 154–166. [Google Scholar] [CrossRef]
- Li, Y.; Liu, H.; Shi, Y.; Yan, Q.; You, X.; Jiang, Z. Preparation, characterization, and prebiotic activity of manno-oligosaccharides produced from cassia gum by a glycoside hydrolase family 134 β-mannanase. Food Chem. 2020, 309, 125709. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, A.; Wiemann, M.; Stålbrand, S. β-Mannanase BoMan26B from Bacteroides ovatus produces mannan-oligosaccharides with prebiotic potential from galactomannan and softwood β-mannans. LWT 2021, 151, 112215. [Google Scholar] [CrossRef]
- Al-Rudainy, B.; Galbe, M.; Wallberg, O. Influence of prefiltration on membrane performance during isolation of lignin-carbohydrate complexes from spent sulfite liquor. Sep. Purif. Technol. 2017, 187, 380–388. [Google Scholar] [CrossRef]
- Al-Rudainy, B.; Galbe, M.; Wallberg, O. From lab-scale to on-site pilot trials for the recovery of hemicellulose by ultrafiltration: Experimental and theoretical evaluations. Sep. Purif. Technol. 2020, 250, 117187. [Google Scholar] [CrossRef]
- Al-Rudainy, B.; Galbe, M.; Wallberg, O. Hemicellulose recovery from spent-sulfite-liquor: Lignin removal by adsorption to resins for improvement of the ultrafiltration process. Molecules 2020, 25, 3435. [Google Scholar] [CrossRef] [PubMed]
- Sipponen, H.M.; Rahikainen, J.; Leskinen, T.; Pihlajaniemi, V.; Mattinen, M.; Lange, H.; Crestini, C.; Osterberg, O. Structural changes of lignin in biorefinery pretreatments and consequences to enzyme-lignin interactions. Nord. Pulp Pap. Res. J. 2017, 32, 550–571. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Y.; Jiang, B.; Chen, H.; Wu, W.; Wu, S.; Jin, Y.; Xiao, H. Recent advances in understanding the effects of lignin structural characteristics on enzymatic hydrolysis. Biotechnol. Biofuels 2021, 14, 205. [Google Scholar] [CrossRef]
- Rahikainen, J.; Mikander, S.; Marjamaa, K.; Tamminen, T.; Lappas, A.; Viikari, L.; Kruus, K. Inhibition of enzymatic hydrolysis by residual lignins from softwood-study of enzyme binding and inactivation on lignin-rich surface. Biotechnol. Bioeng. 2011, 108, 2823–2834. [Google Scholar] [CrossRef]
- Vermaas, J.V.; Petridis, L.; Qi, X.; Schulz, R.; Lindner, B.; Smith, J.C. Mechanism of lignin inhibition of enzymatic biomass deconstruction. Biotechnol. Biofuels 2015, 8, 217. [Google Scholar] [CrossRef] [Green Version]
- Zheng, W.; Lan, T.; Li, H.; Yue, G.; Zhou, H. Exploring why sodium lignosulfonate influenced enzymatic hydrolysis efficiency of cellulose from the perspective of substrate–enzyme adsorption. Biotechnol. Biofuels 2020, 13, 19. [Google Scholar] [CrossRef] [Green Version]
- Yarbrough, J.M.; Mittal, A.; Mansfield, E.; Taylor, L.E.; Hobdey, S.E.; Sammond, D.W.; Bomble, Y.J.; Crowley, M.F.; Decker, S.R.; Himmel, M.E.; et al. New perspective on glycoside hydrolase binding to lignin from pretreated corn stover. Biotechnol. Biofuels 2015, 8, 214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, S.; Huang, Y.; Sun, R.; Tu, M. The strong association of condensed phenolic moieties in isolated lignins with their inhibition of enzymatic hydrolysis. Green Chem. 2016, 18, 4276–4286. [Google Scholar] [CrossRef]
- Malgas, S.; Van Dyk, S.J.; Abboo, S.; Pletschke, B.I. The inhibitory effects of various substrate pre-treatment by-products and wash liquors on mannanolytic enzymes. J. Mol. Catal. B Enzym. 2016, 123, 132–140. [Google Scholar] [CrossRef]
- Avila-Gaxiola, J.; Velarde-Escobar, O.J.; Millan-Almaraz, J.R.; Ramos-Brito, F.; Atondo-Rubio, G.; Yee-Rendon, C.; Avila-Gaxiola, E. Treatments to improve obtention of reducing sugars from agave leaves powder. Ind. Crops Prod. 2018, 112, 577–583. [Google Scholar] [CrossRef]
- López-Gutiérrez, I.; Razo-Flores, E.; Méndez-Acosta, H.O.; Amaya-Delgado, L.; Alatriste-Mondragón, F. Optimization by response surface methodology of the enzymatic hydrolysis of non-pretreated agave bagasse with binary mixtures of commercial enzymatic preparations. Biomass Convers. Biorefinery 2021, 11, 2923–2935. [Google Scholar] [CrossRef]
- Krawczyk, H.; Jönsson, A.S. Separation of dispersed substances and galactoglucomannan in thermomechanical pulp process water by microfiltration. Sep. Purif. Technol. 2011, 79, 43–49. [Google Scholar] [CrossRef]
- Al-Rudainy, B.; Galbe, M.; Lipnizki, F.; Wallberg, O. Galactoglucomannan recovery with hydrophilic and hydrophobic membranes: Process performance and cost estimations. Membranes 2019, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersson, A.; Persson, T.; Zacchi, G.; Stålbrand, H.; Jönsson, A.S. Comparison of diafiltration and size-exclusion chromatography to recover hemicelluloses from process water from thermomechanical pulping of spruce. Appl. Biochem. Biotechnol. 2007, 137, 971–983. [Google Scholar] [CrossRef]
- Naidjonoka, P.; Hernandez, M.A.; Pálsson, G.K.; Heinrich, F.; Stålbrand, H.; Nylander, T. On the interaction of softwood hemicellulose with cellulose surfaces in relation to molecular structure and physicochemical properties of hemicellulose. Soft Matter 2020, 16, 7063–7076. [Google Scholar] [CrossRef]
- Stalbrand, H.; Siika-aho, M.; Tenkanen, M.; Viikari, L. Purification and characterization of two β-mannanases from Trichoderma reesei. J. Biotechnol. 1993, 29, 229–242. [Google Scholar] [CrossRef]
- Vu, H.P.; Nguyen, L.N.; Vu, M.T.; Johir, M.A.H.; McLaughlan, R.; Nghiem, L.D. A comprehensive review on the framework to valorise lignocellulosic biomass as biorefinery feedstocks. Sci. Total Environ. 2020, 743, 140630. [Google Scholar] [CrossRef] [PubMed]
- Kumar, B.; Bhardwaj, N.; Agrawal, K.; Chaturvedi, V.; Verma, P. Current perspective on pretreatment technologies using lignocellulosic biomass: An emerging biorefinery concept. Fuel Process. Technol. 2020, 199, 106244. [Google Scholar] [CrossRef]
- Islam, M.K.; Wang, H.; Rehman, S.; Dong, C.; Hsu, H.Y.; Lin, C.S.K.; Leu, S.Y. Sustainability metrics of pretreatment processes in a waste-derived lignocellulosic biomass biorefinery. Bioresour. Technol. 2020, 298, 122558. [Google Scholar] [CrossRef]
- Von Freiesleben, P.; Spodsberg, N.; Stenbæk, A.; Stålbrand, H.; Krogh, K.B.R.M.; Meyer, A.S. Boosting of enzymatic softwood saccharification by fungal GH5 and GH26 endomannanases. Biotechnol. Biofuels 2018, 11, 194. [Google Scholar] [CrossRef] [PubMed]
- Tenkanen, M.; Makkonen, M.; Perttula, M.; Viikari, L.; Teleman, A. Action of Trichoderma reesei mannanase on galactoglucomannan in pine kraft pulp. J. Biotechnol. 1997, 57, 191–204. [Google Scholar] [CrossRef]
- Dias, F.M.V.; Vincent, F.; Pell, G.; Prates, J.A.M.; Centeno, M.S.J.; Tailford, L.E.; Ferreira, L.M.A.; Fontes, C.M.G.A.; Davies, G.J.; Gilbert, H.J. Insights into the molecular determinants of substrate specificity in glycoside hydrolase family 5 revealed by the crystal structure and kinetics of Cellvibrio mixtus mannosidase 5A. J. Biol. Chem. 2004, 279, 25517–25526. [Google Scholar] [CrossRef] [Green Version]
- Meyer, A.S.; Dam, B.P.; Lærke, H.N. Enzymatic solubilization of a pectinaceous dietary fiber fraction from potato pulp: Optimization of the fiber extraction process. Biochem. Eng. J. 2009, 43, 106–112. [Google Scholar] [CrossRef]
- Setati, M.E.; Ademark, P.; Van Zyl, W.H.; Hahn-Hägerdal, B.; Stålbrand, H. Expression of the Aspergillus aculeatus endo-β-1,4-mannanase encoding gene (man 1) in Saccharomyces cerevisiae and characterization of the recombinant enzyme. Protein Expr. Purif. 2001, 21, 105–114. [Google Scholar] [CrossRef]
- Agrawal, R.; Gaur, R.; Mathur, A.; Kumar, R.; Gupta, R.P.; Tuli, D.K.; Satlewal, A. Improved saccharification of pilot-scale acid pretreated wheat straw by exploiting the synergistic behavior of lignocellulose degrading enzymes. RSC Adv. 2015, 5, 71462–71471. [Google Scholar] [CrossRef]
- Bombeck, P.L.; Khatri, V.; Meddeb-Mouelhi, F.; Montplaisir, D.; Richel, A.; Beauregard, M. Predicting the most appropriate wood biomass for selected industrial applications: Comparison of wood, pulping, and enzymatic treatments using fluorescent-tagged carbohydrate-binding modules. Biotechnol. Biofuels 2017, 10, 293. [Google Scholar] [CrossRef] [Green Version]
- Pawar, P.M.A.; Koutaniemi, S.; Tenkanen, M.; Mellerowicz, E.J. Acetylation of woody lignocellulose: Significance and regulation. Front. Plant Sci. 2013, 4, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tenkanen, M.; Puls, J.; Rättö, M.; Viikari, L. Enzymatic deacetylation of galactoglucomannans. Appl. Microbiol. Biotechnol. 1993, 39, 159–165. [Google Scholar] [CrossRef]
- Arnling Bååth, J.; Martínez-Abad, A.; Berglund, J.; Larsbrink, J.; Vilaplana, F.; Olsson, L. Mannanase hydrolysis of spruce galactoglucomannan focusing on the influence of acetylation on enzymatic mannan degradation. Biotechnol. Biofuels 2018, 11, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Börcsök, Z.; Pásztory, Z. The role of lignin in wood working processes using elevated temperatures: An abbreviated literature survey. Eur. J. Wood Wood Prod. 2021, 79, 511–526. [Google Scholar] [CrossRef]
- Zhu, J.Y.; Pan, X.J.; Wang, G.S.; Gleisner, R. Sulfite pretreatment (SPORL) for robust enzymatic saccharification of spruce and red pine. Bioresour. Technol. 2009, 100, 2411–2418. [Google Scholar] [CrossRef]
- Wang, Z.; Zhu, J.Y.; Fu, Y.; Qin, M.; Shao, Z.; Jiang, J.; Yang, F. Lignosulfonate-mediated cellulase adsorption: Enhanced enzymatic saccharification of lignocellulose through weakening nonproductive binding to lignin. Biotechnol. Biofuels 2013, 6, 156. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, M.; Pu, Y.; Ragauskas, A.J.; Klett, A.S.; Thies, M.; Zheng, Y. Inhibitory effects of lignin on enzymatic hydrolysis: The role of lignin chemistry and molecular weight. Renew. Energy 2018, 123, 664–674. [Google Scholar] [CrossRef]
- Pareek, N.; Gillgren, T.; Jonsson, L. Adsorption of proteins involved in hydrolysis of lignocellulose on lignins and hemicelluloses. Bioresour. Technol. 2013, 148, 70–77. [Google Scholar] [CrossRef] [Green Version]
- Ruwoldt, J. A Critical Review of the Physicochemical Properties of Lignosulfonates: Chemical Structure and Behavior in Aqueous Solution, at Surfaces and Interfaces. Surfaces 2020, 3, 622–648. [Google Scholar] [CrossRef]
- Palonen, H.; Tjerneld, F.; Zacchi, G.; Tenkanen, M. Adsorption of Trichoderma reesei CBH I and EG II and their catalytic domains on steam pretreated softwood and isolated lignin. J. Biotechnol. 2004, 107, 65–72. [Google Scholar] [CrossRef]
- Sabini, E.; Schubert, H.; Murshudov, G.; Wilson, K.S.; Siika-Aho, M.; Penttila, M. The three-dimensional structure of a Trichoderma reesei β-mannanase from glycoside hydrolase family 5. Acta Crystallogr. Sect. D Biol. Crystallogr. 2000, 56, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Min, D.Y.; Li, Q.; Chiang, V.; Jameel, H.; Chang, H.M.; Lucia, L. The influence of lignin-carbohydrate complexes on the cellulase-mediated saccharification I: Transgenic black cottonwood (western balsam poplar, California poplar) P. trichocarpa including the xylan down-regulated and the lignin down-regulated lines. Fuel 2014, 119, 207–213. [Google Scholar] [CrossRef]
- Min, D.Y.; Yang, C.; Chiang, V.; Jameel, H.; Chang, H.M. The influence of lignin-carbohydrate complexes on the cellulase-mediated saccharification II: Transgenic hybrid poplars (Populus nigra L. and Populus maximowiczii A.). Fuel 2014, 116, 56–62. [Google Scholar] [CrossRef]
- Zhao, Y.; Shakeel, U.; Saif Ur Rehman, M.; Li, H.; Xu, X.; Xu, J. Lignin-carbohydrate complexes (LCCs) and its role in biorefinery. J. Clean. Prod. 2020, 253, 120076. [Google Scholar] [CrossRef]
- Rahikainen, J.L.; Moilanen, U.; Nurmi-Rantala, S.; Lappas, A.; Koivula, A.; Viikari, L.; Kruus, K. Effect of temperature on lignin-derived inhibition studied with three structurally different cellobiohydrolases. Bioresour. Technol. 2013, 146, 118–125. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Pu, Y.; Ragauskas, A.J. Current Understanding of the Correlation of Lignin Structure with Biomass Recalcitrance. Front. Chem. 2016, 4, 45. [Google Scholar] [CrossRef] [Green Version]
- Rosales-Calderon, O.; Arantes, V. A review on commercial-scale high-value products that can be produced alongside cellulosic ethanol. Biotechnol. Biofuels 2019, 12, 240. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, H.; Hasunuma, T.; Ogino, C.; Kondo, A. Bioprocessing of bio-based chemicals produced from lignocellulosic feedstocks. Curr. Opin. Biotechnol. 2016, 42, 30–39. [Google Scholar] [CrossRef] [PubMed]
- Xavier, A.M.R.B.; Correia, M.F.; Pereira, S.R.; Evtuguin, D.V. Second-generation bioethanol from eucalypt sulphite spent liquor. Bioresour. Technol. 2010, 101, 2755–2761. [Google Scholar] [CrossRef]
- Aro, T.; Fatehi, P. Production and Application of Lignosulfonates and Sulfonated Lignin. ChemSusChem 2017, 10, 1861–1877. [Google Scholar] [CrossRef]
- Hägglund, P.; Eriksson, T.; Collén, A.; Nerinckx, W.; Claeyssens, M.; Stålbrand, H. A cellulose-binding module of the Trichoderma reesei β-mannanase Man5A increases the mannan-hydrolysis of complex substrates. J. Biotechnol. 2003, 101, 37–48. [Google Scholar] [CrossRef]
- Gonzalez-garcia, S.; Hospido, A.; Agnemo, R.; Svensson, P.; Selling, E.; Moreira, T.M.; Feijoo, G. Environmental Life Cycle Assessment of a Swedish Dissolving Pulp Mill Integrated Biorefinery. J. Ind. Ecol. 2011, 15, 568–583. [Google Scholar] [CrossRef]
- Sluiter, A.; Hames, B.; Ruiz, R.; Scarlata, C.; Sluiter, J.; Templeton, D.; Crocker, D. Determination of Structural Carbohydrates and Lignin in Biomass. Laboratory Analytical Procedures (LAP); Technical Report NREL/TP-510-42618; National Renewable Energy Laboratory: Golden, CO, USA, 2012.
- Jacobs, A.; Lundqvist, J.; Stålbrand, H.; Tjerneld, F.; Dahlman, O. Characterization of water-soluble hemicelluloses from spruce and aspen employing SEC/MALDI mass spectroscopy. Carbohydr. Res. 2002, 337, 711–717. [Google Scholar] [CrossRef]
- Immerzeel, P.; Falck, P.; Galbe, M.; Adlercreutz, P.; Nordberg, E.; Stålbrand, H. Extraction of water-soluble xylan from wheat bran and utilization of enzymatically produced xylooligosaccharides by Lactobacillus, Bifidobacterium and Weissella spp. LWT 2014, 56, 321–327. [Google Scholar] [CrossRef]
- Berglund, J.; Azhar, S.; Lawoko, M.; Lindström, M.; Vilaplana, F.; Wohlert, J.; Henriksson, G. The structure of galactoglucomannan impacts the degradation under alkaline conditions. Cellulose 2019, 26, 2155–2175. [Google Scholar] [CrossRef]
- Bagenholm, V.; Reddy, S.K.; Bouraoui, H.; Morrill, J.; Kulcinskaja, E.; Bahr, C.M.; Aurelius, O.; Rogers, T.; Xiao, Y.; Logan, D.T.; et al. Galactomannan catabolism conferred by a polysaccharide utilization locus of Bacteroides ovatus: Enzyme synergy and crystal structure of a β-mannanase. J. Biol. Chem. 2017, 292, 229–243. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| All Values in (mg/g) | UFR | AC | AD |
|---|---|---|---|
| Ash | 36.81 | 11.56 | 2.56 |
| Total lignin | 192.16 | 25.44 | 2.65 |
| Klason lignin | 5.27 | 0.88 | 1.57 |
| Arabinan | 0.75 | 0.41 | 0.35 |
| Galactan | 9.62 | 8.92 | 8.04 |
| Glucan | 8.75 | 7.43 | 6.69 |
| Xylan | 2.89 | 2.45 | 2.19 |
| Mannan | 25.52 | 23.59 | 21.56 |
| Arabinose | 0.32 | 0.046 | 0.034 |
| Galactose | 0.09 | 0.015 | 0.002 |
| Glucose | 0.12 | 0.011 | 0.003 |
| Xylose | 0.11 | 0.003 | 0.019 |
| Mannose | 0.11 | 0.017 | 0.011 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bhattacharya, A.; Butler, S.; Al-Rudainy, B.; Wallberg, O.; Stålbrand, H. Enzymatic Conversion of Different Qualities of Refined Softwood Hemicellulose Recovered from Spent Sulfite Liquor. Molecules 2022, 27, 3207. https://doi.org/10.3390/molecules27103207
Bhattacharya A, Butler S, Al-Rudainy B, Wallberg O, Stålbrand H. Enzymatic Conversion of Different Qualities of Refined Softwood Hemicellulose Recovered from Spent Sulfite Liquor. Molecules. 2022; 27(10):3207. https://doi.org/10.3390/molecules27103207
Chicago/Turabian StyleBhattacharya, Abhishek, Samuel Butler, Basel Al-Rudainy, Ola Wallberg, and Henrik Stålbrand. 2022. "Enzymatic Conversion of Different Qualities of Refined Softwood Hemicellulose Recovered from Spent Sulfite Liquor" Molecules 27, no. 10: 3207. https://doi.org/10.3390/molecules27103207