
Yarrowia lipolytica as an Alternative and Valuable Source of Nutritional and Bioactive Compounds for Humans


1
Department of Molecular Biology, The John Paul II Catholic University of Lublin, Konstantynów Street 1I, 20-708 Lublin, Poland
2
Department of Pharmaceutical Microbiology, Medical University of Lublin, Chodźki Street 1, 20-093 Lublin, Poland
*
Author to whom correspondence should be addressed.
Molecules 2022, 27(7), 2300; https://doi.org/10.3390/molecules27072300
Submission received: 22 February 2022
/
Revised: 24 March 2022
/
Accepted: 28 March 2022
/
Published: 1 April 2022
(This article belongs to the Special Issue Microbial Natural Products: A Promising Source for Medical Application)
Abstract
:Yarrowia lipolytica, an oleagineous species of yeast, is a carrier of various important nutrients. The biomass of this yeast is an extensive source of protein, exogenous amino acids, bioavailable essenctial trace minerals, and lipid compounds as mainly unsaturated fatty acids. The biomass also contains B vitamins, including vitamin B12, and many other bioactive components. Therefore, Y. lipolytica biomass can be used in food supplements for humans as safe and nutritional additives for maintaining the homeostasis of the organism, including for vegans and vegetarians, athletes, people after recovery, and people at risk of B vitamin deficiencies.
1. Introduction
Yarrowia lipolytica is a well-known oleaginous yeast that has an ability to degrade wide range of hydrophobic substrates, effectively producing a huge number of valuable metabolic products such as protein and peptides, amino acids, trace minerals, vitamins, and carbohydrates or single-cell oil (SCO), predominantly as mono-unsaturated fatty acids (MUFAs) and saturated high-added-value lipids such as cocoa-butter equivalents [1,2,3,4,5,6,7,8,9,10,11,12,13,14]. For this reason, in recent years, it was the most studied unicellular fungus right after the conventional true yeast Saccharomyces cerevisiae [15,16]. The production of numerous metabolites by yeast cultured on agricultural, industrial, and food residues and wastes is perfect alternative way for the chemical and food industry. Moreover, this production helps to reduce environmental pollutions due to the conversion of waste and byproduct substrates to value-added products [3,8,9,17]. Y. lipolytica utilizing waste as a raw cassava (bioethanol industry by-product) reduces poisonous cyanide concentration to a safe level [18]. The yeast has relatively low nutritional requirements, showing very high growth potential, and its culture is independent of geographic and weather conditions and fresh water availability [6,19,20,21]. Additionally, the use of yeast biomass enriched in protein containing plenty of value compounds or its metabolites is easier accepted by society than other microorganisms due to its usefulness, i.e., in the production of fermented products [20].
Y. lipolytica strains are widespread in nature and can be easily isolated from various environments (i.e., water and soil) such as fat-rich substrates (i.e., oily wastes and sewage), hypersaline and marine sites, as well as contaminated soil at a car wash [22,23,24,25,26]. The yeast strains as a living cells also occur in wide ranges of food products with high amounts of fat and protein, e.g., dietary products such as cheese, especially blue cheese and cheddar cheese, milk, cream, kefir, yoghurt, butter, and margarine; as well as ripening meat products, e.g., various types of fermented sausages such as salami, and Spanish fermented sausages (i.e., chorizo); and various others, such as juice, cider, fruit, fruit concentrate, mayonnaise, seafood, raw plant materials, and wine [24,27,28,29]. Moreover, the yeast was isolated from a healthy human body. Therefore, Y. lipolytica is considered as normal human microbiota belonging to saprophytes in the adult respiratory system. It is also detected in breast tissue, some cutaneous nodules, and the digestive tract [30]. The natural occurrence of live yeast confirms their safety. The yeast is non-pathogenic for humans and animals. The live yeast cells in extreme cases can rarely cause an opportunistic fungemia exclusively in immunocompromised individuals with underlying diseases and those with catheters [24,27,31]. Thus, Y. lipolytica is considered as safe, and various production processes based on the yeast were classified as Generally Recognized as Safe (GRAS) by Food and Drug Administration (FDA) [27]. It is also classified as a Biosafety Level (BSL) 1 microorganism by the Public Health Service (Washington, DC, USA) as well as recognized as a “microorganism with a documented use in food” by the International Dairy Federation (IDF) and the European Food and Feed Cultures Association (EFFCA) [32].
In the 1950s in Great Britain, Y. lipolytica was used by British Petroleum Co. (BP) as a producer of single-cell protein (SCP) for animal feeding [27]. In 2010, throughout the European Union, dried and heat-killed biomass of Y. lipolytica cultivated in biofuel waste has been approved for animal nutrition [24]. Since then, a lot of clinical studies were carried out on Y. lipolytica biomass-fed animals such as fish species. e.g., Atlantic salmon [33,34], Nile tilapia [35], and Pacific red snapper [36]; birds, e.g., turkeys [37,38,39], piglets [40,41,42]; and ruminants, e.g., calves [34]. Dried and heat-killed yeast biomass work effectively as immunostimulants and animal growth promoters [35,37,39,43].
The growth of the human population leads to increased food production, especially protein-rich products. The need to provide food to so many people throughout the world creates a growing gap between suppliers and demand, mainly in developing countries [44,45]. Therefore, the search for new alternative ways of obtaining protein-enriched food is crucial to solving the increasing problem of food deficit. In the near future, the problem could be solved by the production of huge quantities of nutritional yeast biomass. At present, mainly S. cerevisiae biomass and extract are used for human nutrition but not commonly and in small numbers [19,46]. However, the first step to use the Y. lipolytica biomass as food has already been taken. In 2019, European Food and Safety Authority (EFSA) has allowed to use the biomass of Y. lipolytica cultured in waste substrate (i.e., biofuel waste) as a novel food in dietary supplements for the general human population above 3 years of age. The Y. lipolytica biomass was recognized as safe and nutritionally advantageous as shown in Figure 1. The proposed maximum daily portion is 3 g/day for children from 3 years up to 10 years of age and 6 g/day thereafter [24].
The aim of this review is to provide comprehensive information about nutritional metabolites, their genetic and enzymatic production machinery, and conditions for the utilization of wastes and by-products for the production of value compounds.
2. Characteristic of Yarrowia lipolytica
Y. lipolytica, formerly named Candida lipolytica, afterwards Endomycopsis lipolytica, and then as Saccharomycopsis lipolytica, just like S. cerevisiae, belongs to the kingdom of fungi, sub-kingdom Dikaryota, division Ascomycota, sub-division Saccharomycotina, phylum Ascomycetes, class Saccharomycetes (called Hemiascomycetes as well), and order Saccharomycetales [15,24,27,47,48]. Y. lipolytica belongs to the family Dipodascaceae and genus Yarrowia. The name was proposed by van der Walt and von Arx in 1980 in thanks for the identification of a new genus by David Yarrow [29,48]. The species name ‘lipolytica’ was given due to the ability of this yeast to hydrolyze fat substrates [29]. Y. lipolytica and Candida lipolytica are the teleomorth and anamorth (respectively) names of the same species [24]. Y. lipolytica belongs to the Yarrowia clade, a big group that includes many different species of yeasts from genera Candida and Yarrowia, such as C. alimentaria, C. bentonensis, C. galii, C. hispaniensis, C. hollandica, C. oslonensis, C. parafonii, C. pseudorhagii, Y. brassicae, Y. bubula, Y. deformants, Y. divulgata, Y. keelungensis, Y. phangngaensis, and Y. porcina and Y. yakushimensis, which were isolated from various biotic and abiotic environments (i.e., food, marine water, termite intestine) [49,50,51].
The genome sequence of Y. lipolytica shows that the yeast is distantly related to S. cerevisiae. The fundamental genetic mechanisms of Y. lipolytica are significantly different in comparison to S. cerevisiae. The genome of Y. lipolytica reveals an accretion of genes and protein families that are involved in hydrophobic substrate utilization, particularly fat-rich ones [52].
Y. lipolytica, as a unicellular form, multiplies vegetatively by polar budding, as in the conventional yeast form, and in sexual reproduction, forms asci which comprise 1–4 haploid ascospores [53]. Y. lipolytica, as a heterothallic microorganism, naturally occurs in two mating types, marked as MatA and MatB [29].
In contrast to S. cerevisiae, Y. lipolytica can grow as a dimorphic form, making it an ideal model for dimorphism studies. It can produce true filaments and presents pseudo-hyphae [27,29,54,55]. Y. lipolytica may form residual mycelium, well-developed mycelium, or does not form filaments at all. Not filamentous forms are usually oval and ellipsoidal. The creation of dimorphic forms depends primarily on adverse environmental conditions such as alteration in oxygen, pH, carbon, and nitrogen substrates [29,56]. Elongation into hyphae may be induced by replacing glucose with N-acetylglucosamine or by the addition of the serum to the medium [57]. Wild-type (wt) strains exhibit various colony morphologies that depend on culture conditions and genetic backgrounds. Colonies’ looks may vary from heavily convoluted and matte to smooth and glistening [29]. Moreover, Y. lipolytica is able to form a biofilm in the presence of different carbon sources, including alkanes [58]. The ability of Y. lipolytica to form biofilms may be important in natural environments, as well as when it grows in different wastes [15]. Wt Y. lipolytica utilizing various wastes or by-products (e.g., glycerol, alkanes, animal or vegetable fats, fatty esters, soap-stocks, pure free-fatty acids, petrol byproduct, and biofuel waste) can produce many bioavailable nutritional compounds [1,2,3,7,8,9,10,11,12,13,14]. Apart from glycerol and oily wastes, Y. lipolytica can degrade several hexoses, e.g., glucose, fructose, galactose, and mannose [59]. The yeast does not utilize sucrose and lactose [60]. However, one isolate from soil (Y. lipolytica B6) can degrade lactose [61]. The yeast also has the complete genetic pathway for xylose and arabinose utilization. Despite that, there are no studies to support the utilization of these substances by the yeast [62]. When glucose or glycerol are absent, Y. lipolytica can utilize ethanol, acetic, butyric, citric, lactic, malic, propionic, and succinic acids as carbon and energy sources [54,63,64]. Interestingly, lactic acid may be also used as a substrate when nitrogen is unavailable (i.e., free amino acids) in the culture medium [65]. The wt yeast does not synthesize carotenoid dyes and does not assimilate inositol and nitrates [20].
3. Culture Condition in Various Wastes for Nutritional Biomass Production
Y. lipolytica requires strictly aerobic condition to grow. Meeting this condition is actually the only difficulty in growing this yeast in an industrial scale. Y. lipolytica can be cultured at a wide range of temperatures from 2 °C to 32 °C and pH from 2.5 to 8.0 [56]. Most Yarrowia strains are incapable of growing above 32 °C, which supports the thesis about their safety of use for human nutrition [29]. It is considered that its optimal culture conditions are ambient temperatures between 28 °C and 30 °C and mild acidic pH about 5.5 [66]. However, as we showed in Table 1, depending on the waste substrate used, the optimal pH value changed. These pH values were in range from strong acidic (3.0) to mildly acidic (5.5). Very low pH may prevent the yeast culture from contamination with other microorganisms. These productions take a relatively short time, between 12 h and 120 h, depending on culture conditions and waste medium used.
Apart from temperature and pH, important parameters for the fermentation process are also the aeration rate, dissolved oxygen, or the moisture content of solid medium [78]. The choice of waste substrate and all culture conditions must be based on optimization studies, preferably using statistical techniques [9,68,79,80,81,82,83]. For the production of protein-enriched biomass, presented species (Table 1) were cultured both in liquid and on solid waste materials, mainly industrial substrates such as glycerol, biofuel waste, and solid tallow, as well as agricultural wastes such as rye straw, ray and oat brans, and soy bean hydrolysates. In case of substrates containing fiber, e.g., inulin or Jerusalem artichoke tubers, it was necessary to use recombinant strains (mutants) due to wt Y. lipolytica not being able to produce cellulases and hemicellulases; thus, it does not utilize fiber in agricultural or forestry residues [84]. In this regard, it is postulated to improve obtaining nutritional biomass by developing genetic engineering procedures for mass production [85]. However, there is a big problem due to the strict regulation on the use and culture of genetic modified organisms (GMO) in the human diet in the European Union [72]. Thus, for large-scale yeast biomass production at competitive prices constituting a feasible replacement for soybean and fishmeal protein in nutrition, including aquaculture [86], wt yeast culture development is strongly desired.
The capacity of growing wt strains in a wide-range of wastes contributes to the biodegradation of the wastes, especially alkanes, petroleum by-products, glycerol, biofuel waste, and plant or animal-waste fats [3,6,8,9,14,17]. Furthermore, liquid substrates are dispersed more easily with moderate agitation than solid media (i.e., tallow), which require strong agitation (i.e., 1200 rpm) for dispersal in the culture materials [75]. It is worth emphasizing that the oily waste biodegradation by the yeast is essential for environmental preservation, in line with the take–make–dispose concept [9,87,88,89]. Moreover, as for being environmentally friendly, Y. lipolytica tolerates heavy metal ions, including cadmium, cobalt, and nickel, as well as high salt concentrations up to 12% (v/v) [56]. Thus, it is involved in the detoxification of heavy metals and other pollutants [15].
Sometimes, during the optimization process, there is a need to add into the waste medium various nutrients such as nitrogen, phosphorus, and others to improve optimal yeast growth and value bioactive compound production by selected strain [90]. The presence of nitrogen in the medium has a fundamental impact on the metabolic pathways and the production of metabolites [65]. Nitrogen availability is particularly important for protein synthesis [91]. In turn, nitrogen deficiency in the medium is desirable for citric acid production or lipid accumulation as storage lipid bodies when glucose or glycerol are in the culture medium. However, there are Y. lipolytica strains which may accumulate lipids when nitrogen is available or just after being deprived of nitrogen. Then, when nitrogen-limited conditions were reached, the strains consumed the storage lipid bodies [92].
Interestingly, the addition of selenium (NaHSeO3) from 1 to 6 mg/dm3 lowered the amount of protein in biomass of Y. lipolytica A-101 cultured in raw rapeseed oil twofold (from 56.4 to 24.7%, respectively). In turn, the addition of chromium (CrCl3 × 6H2O) from 5 to 80 mg/dm3 into the same medium caused a decrease in protein content from 39.4 to 20%, respectively [71]. The addition of 4 mg/dm3 selenium (IV) and 40 mg/dm3 chromium (III) in the culture medium was the most effective at simultaneously producing biomass rich in protein and high concentrations of these minerals. These results were confirmed by Juszczyk et al. [70]. These studies showed [70,71] that regardless of the carbon source, Y. lipolytica is characterized by a good accumulation of selenium and chromium in the cell.
The search for the best bioproduct producer should also be based on optimization methods. One of the most effective methods to derive new highly productive strains is an adaptive laboratory evolution (ALE) strategy. Daskalaki et al. [93], using ALE, obtained the strain that accumulated 30% more fats after 77 generations than the starting Y. lipolytica strain.
In case of yeast biomass production for human consumption, a process step should be added for the reduction or removal of nucleic acid to a safe level of 1% because higher RNA concentrations when consumed by humans can be toxic [94]. The biomass of most yeast contains low amounts of nucleic acid in ranges from 5% to 8%. It is lower than other microorganisms contain, e.g., bacteria contain 8 to 15% [95]. For example, the biomass of Y. lipolytica cultivated in biofuel waste contained from 6.4% of RNA in an initial lag phase, through 5.8% in the end of log phase to 0.4% at the end of stationary phase [6]. It was observed earlier that during stationary phase microbial cells have the lowest content of nucleic acids, which is partially caused by reduction of RNA quantity [96]. Hence, one of the most effective methods of nucleic acid reduction is simple elongation of Y. lipolytica culture time up to the final stage of the stationary phase while endogenous nucleases (ribonucleases) are activated [6]. Endogenous nucleases are also activated by thermal shock (60–90 °C) [97]. Other less recommendable methods are the following: chemical treatment with NaOH or 10% NaCl, the addition of ribonucleases to the cultivation process, or the use of these nucleases as immobilized enzymes [97,98,99] and two-step treatment with a chemical combination of N-lauroyl sarcosine and NH4OH, which reduced RNA to below 2% [100].
To obtain edible, digestible and bioavailable Y. lipolytica cells, they should be destroyed by drying at high temperatures, resulting in dried and heat-killed biomass adequate for human consumption [6,24]. Among cell disintegration methods (e.g., crushing, crumbling, grinding, pressure homogenization, or ultrasonification), drying at high temperature is one of the most effective methods of preventing contamination and extending the shelf life of feed or food [101]. The moisture content of dried Yarrowia powder was lowered below 6% [6,24]. A moisture level below 8% is considered as safe for long shelf life of products and helps to achieve proper texture and stability of yeast biomass as food ingredients [102]. Additionally, drying at high temperatures (e.g., 165–175 °C) kills cells, disintegrates the cell wall, and contributes to the release of the yeast cells’ contents. Therefore, drying significantly improves the quality of yeast biomass [46,85]. A heat-killing step of yeast biomass production results in the absence of viable Y. lipolytica in the powder [103,104]. Dried and heat-killed Y. lipolytica biomass obtained after the culture in biofuel waste is an amorphous hygroscopic beige-colored powder, as shown in Figure 2, with a slight yeast odor [5,40].
4. Y. lipolytica as a Producer of Protein Biomass
4.1. Protein Concentration
Y. lipolytica cultivated in varied oily substrates produces plenty of nutritional compounds; therefore, it was given a name nutritional yeast. As shown in Table 1, Y. lipolytica is an excellent producer of protein such as SCP. The Y. lipolytica biomass contains abundant protein in range between 30.5% and 56.4%, with an average protein value of 43%. It is considered that the preferable protein concentration is just above 40% [105]. EFSA and FAO recommend an intake of 50 g of protein in daily human diet for adults [106,107]. Hence, 100 g of Y. lipolytica powder contains almost 100% of the recommended daily portion. Moreover, protein quantity in Y. lipolytica powder is comparable to that of S. cerevisiae biomass and is similar or even higher than in traditional sources such as meat and soybean and higher than milk protein [19,46,72]. Furthermore, the protein efficiency ratio standardized for casein (PER) of yeast protein is comparable with the PER of meat and soybean [72,108]. The digestibility coefficients of Y. lipolytica protein are very high and are in the range of 72.3% and 77.2% [70,72,74] and are comparable to that of S. cerevisiae (73.7%) [72]. Because Y. lipolytica has a comparable chemical composition to that of S. cerevisiae, it may to have similar functions in human and animal organisms [38]. Moreover, the digestibility coefficients of protein biomass for both yeast species are similar in the feeding of different animals such as rats and piglets [109]. In turn, the digestibility of the ether extract of Y. lipolytica grown on industrial glycerol was 57.3%, as shown in Table 2, while, for S. cerevisiae, the ether extract was absent entirely [72].
The protein biomass of Y. lipolytica cultivated in industrial glycerol or biofuel waste contains good portion of dietary fiber up to 30.7% of total dry matter (Table 2) [24,72,103,104]. Y. lipolytica fiber is mainly composed of β-glucans, complex polysaccharides that form part of the cell walls of yeasts and cereals [24,103,104,111]. In the Western world, human diets are scarce in fiber. Recently, Barber et al. [112] analyzed many studies from which they summarized that dietary fiber intake is associated with overall metabolic health (through key pathways that include insulin sensitivity) and a variety of other pathological conditions including cardiovascular disease, colonic health, gut motility, and colorectal carcinoma risk. Dietary fiber intake is also correlated with mortality. The gut microflora works as a significant mediator of the pro-health effects of dietary fiber, including the regulation of appetite, metabolic processes, and chronic inflammatory pathways; hence, β-glucans are considered as prebiotic.
4.2. Amino Acid Content in Y. lipolytica Protein Biomass
The protein nutritional value is determined by proper amino acid composition, predominately of eight essential amino acids which the human body is incapable of synthesizing [97]. As shown in Table 3, Y. lipolytica powder contains a complete set of amino acids, making this biomass a valuable source of complete protein [6,67,70,72,113,114,115,116]. It is worth emphasizing that Y. lipolytica biomass enriched in good-quality protein is obtained in a much shorter time and much easier and cheaper, with a low environmental footprint and an environmentally friendly process that utilizes various wastes, which traditional protein sources such as plants and animals do not.
The number of amino acids depends on the medium used. The lowest amino acid levels were obtained when the yeast was grown on rye straw and rye or oat brans [67]. However, the use of fatty substrates such as crude or industrial glycerol, or biofuel waste enhanced the number of amino acids resulting in much higher quantities, yielding them in the upper limit (Table 3) [6,68,72]. The essential amino acid profile in the protein of Y. lipolytica when grown in these fatty substrates (i.e., glycerol and biofuel waste) is comparable to or higher than those required by the FAO for daily human diet (Table 3). It is very important because protein deficiency may occur even when one of the essential amino acids is missing [117]. Moreover, the protein of the yeast cultured in the fatty materials such as glycerol and biofuel waste contain high quantities of lysine, of which there is too little in wheats, including cereals [118].
4.3. Genetic and Enzymatic Machinery of Protein Biomass Production
It is known that Y. lipolytica growing in oily substrates accumulates huge amounts of lipids intracellularly as storage lipid bodies (more than 40%), referred to as single-cell oils (SCOs) [1,119,120]. However, the production de novo and the accumulation ex novo of SCOs depends on the type of strains and the culture substrates, and the two processes can occur simultaneously [121]. Wt strains, cultured in sugar medium, produce lipids less efficiently, reaching 4–20% of intracellular lipids in dry weight (DW), or more efficiently when they are cultivated in hydrophobic materials [8,10,56,121,122]. Production referred to as de novo occurs when the yeast grows in sugar medium and similarly metabolized substrates, i.e., glycerol. SCOs are synthetized during a secondary metabolism, which occurs especially when nitrogen is depleted or a high carbon/nitrogen (C/N) ratio in the medium exists [8,10]. Nitrogen starvation in the environment starts a cascade of metabolic reactions, leading to a rapid reduction in intracellular AMP, which causes mitochondrial AMP-activated isocitrate dehydrogenase (ICDH) inhibition. ICDH inhibition is decisive in signaling the first step of the lipogenesis process due to the disturbance of the Krebs cycle, inducing the strong accumulation of intramitochondrial citric acid, which is excreted to the cytoplasm instead of malate. Then, in the cytoplasm, citric acid is hydrolyzed to acetyl-CoA and oxaloacetate by ATP citrate lyase (acyltransferases, ACL). It is noteworthy that the transcription of genes for ACL and ICDH is observed in both oleaginous and non-oleaginous conditions [1,3]. Moreover, for even more effective lipid accumulation by Y. lipolytica, apart from nitrogen starvation, magnesium starvation was also favored with comparison to the presence of only one of the two (from 13.6 to 38.5%). Although, for effective lipid production, it is sufficient to provide nitrogen starvation or high C/N ration [1,56]. However, it is detected that the yeast has already produced lipids in the first step of its growth when both nitrogen and crude glycerol as carbon and energy source in the medium were presented [92,123]. Furthermore, simple pH control could also increase lipid accumulation [124]. In turn, ex novo lipid production as the primary anabolic cell process occurs when the yeast is cultured in fat-rich substrates. When Y. lipolytica is grown in fatty materials, it is proven that the yeast rapidly incorporates unsaturated fatty acids, i.e., oleic acid for proper growth and organic acid production. However, saturated fatty acids (SFAs), e.g., stearic acid, are slowly incorporated and used for proper growth, as well as SCO production [10]. SCOs are primarily composed as triacylglycerols (TAGs, called neutral fats, triglycerides TGs, or triacyl glycerides) with low levels of free fatty acids, neutral lipids, polar fractions, and sterols. Among the accumulated fatty acids, oleic, linoleic, and palmitic acids are mostly represented [66,125,126]. Moreover, the modulation of growth parameters and substrates resulted in obtaining tailor-made SCOs [8,10]. It is observed that Y. lipolytica cultured in various oils or oily wastes produced SCOs abundant in oleic acid. In turn, the yeast cultured in SFAs, e.g., stearin, generated SCOs rich in SFAs. Furthermore, when the yeast is grown in a mixture of hydrolyzed rapeseed oil and stearin in proportion 50:50, its lipids were composed similarly to cocoa butter [10].
However, it is proven that ex novo lipid production is independent of nitrogen starvation. Utilizing accumulated SCOs, fat-free metabolites are produced [10,56]. Hence, the protein biomass of Y. lipolytica grown in agricultural (i.e., rye and oat straw or brans) or industrial wastes (i.e., industrial or raw glycerol) contains low concentrations of total cellular lipids (8.2–20%, respectively), with a high spectrum of unsaturated fatty acids (65–90% of total lipids). Among the unsaturated fatty acids, it is dominated by MUFAs (57.6%), represented by oleic acid (34–59.2%), and in low concentrations of poly-unsaturated fatty acids (PUFAs) (34%), represented by linoleic acid (11.9–27%). Protein biomass has low amount of SFAs (8.3–34.9% of total lipids) [67,72]. The low content of total lipids in protein biomass is the result of using accumulated lipids in storage bodies as source of carbon and energy for the yeast cell growth when the extracellular carbon source is depleted and protein or cellular carbohydrates biosynthesis. Thus, the accumulation of lipids and protein or carbohydrates production are competitive processes [3,93].
The degradation of hydrophobic materials (i.e., alkanes, fatty acids, and oily wastes) by Y. lipolytica is a complicated multistep process in which a number of genes encoding protein family enzymes responsible for the fat utilization are involved [29]. Y. lipolytica possesses more than 25 genes encoding lipases and estreases, among them, LIP2, LIP7, and LIP8 are specific for fatty acid chains of lengths C18, C6, and C10, respectively. Extracellularly released lipase (Lip2) has a high specificity towards saturated triglyceride-tricaprylin (C8:0) and TGs containing MUFA triolein (C18:1), as well as C12, C14, and C16 methyl esters [127,128,129]. Y. lipolytica also produces intracellular lipases (Lip1, Lip3, and Lip6) and cell-bounded Lip7 and Lip8 [127,128]. The latter lipases prefer p-nitrophenyl C8–C12 esters and p-nitrophenyl C8–C10 ones [129]. Additionally, the yeast has six POX genes which are characteristic only for oleaginous ones. Among them, POX2 and POX3 code long and short specific acyl-CoA oxidase enzymes, respectively, which directly allows the yeast to utilize fat-rich substrates by complete oxidation using intermediates of β-oxidation [59,130,131,132]. Interestingly, a transfer of the one gene encoding acetyl-CoA oxidase from Y. lipolytica to S. cerevisiae enabled S. cerevisiae to grow in fat-rich medium [133]. Furthermore, a knockout of the all-POX genes in Y. lipolytica causes the inability of this yeast to degrade storage lipids, leading to an over-accumulation of fats in yeast cells [59,134]. Moreover, Y. lipolytica possesses 12 genes from the CYP52 family for cytochrome P450. Some of them code specific enzymes for alkanes or fatty acid hydroxylation [29,52,135].
When the yeast is cultivated in fatty substrates, at first, the hydrophobic cell wall of Y. lipolytica secrets emulsifiers, surfactants, and extracellular lipases that hydrolyze TAGs to fatty acids and glycerol [25]. During the growth on fatty-acid-containing media, extensive proliferation of peroxisomes and intense accumulation of lipids as storage lipid bodies occur. In turn, when yeast grow on a fatty acid-poor or fat-fee carbon source (i.e., sugar medium), peroximal structures cannot be identified. Peroxisomes play the sole role in β-oxidation in the fatty acid degradation process causing the formation of acetyl-CoA in the peroxisome [136,137]. The SCOs accumulation lasts until extracellular carbon sources exist. Furthermore, extracellular lipase content decreases with depletion of the oily substrate in the stationary growth phase [75]. However, the addition of oily materials (i.e., waste cooking oil) into the culture medium induces extracellular lipase production by Y. lipolytica, but in fat-free cultures, it does not [9,138].
Since carbon sources in the culture medium are exhausted, the yeast have to utilize their own storage lipids as a carbon and energy source to survive and maintain vital functions, using intracellular lipases to hydrolase SCOs, then consequently decreasing fat content in the yeast cells and increasing the production of protein and other fat-free metabolites by shifting the metabolism from SCOs production to fat-free metabolites biosynthesis [1,3,10,56,75,93,139,140]. This process leads to obtaining de-fatted Y. lipolytica biomass containing half of its weight as protein (up to 56.4%) with all the essential amino acids in significant amounts. It has been proven that with the increase in protein in the yeast biomass, there is a significant degradation of the storage lipid bodies dependent on the availability of nitrogen and magnesium in the culture medium [1,93]. Moreover, it was observed that the enhancement of the state of aeration and agitation of the culture medium increased the activity of short-chain acyl-CoA oxidases. Consequently, the carbon flow was driven to the synthesis of acetyl-CoA and then to formation of de-fatted protein biomass rather than storage lipophilic biomass [141]. Of note, both extracellular and intracellular lipases show their activity in a wide spectrum of temperatures (4–55 °C), depending on the used strain [127,129,141]. However, the maximum activity of the lipases was found at the temperatures between 30 °C and 40 °C and at pH 5.0, while a significant lowering in their stability was observed at pH values over 6.5 [9,142]. Moreover, the most efficient production of protein biomass with many bioactive compounds was also noted under similar conditions: temperatures of 30 °C and pH 5.0 [6,68]. Therefore, the manipulation of culture conditions (i.e., agitation and aeration), oily medium compounds (i.e., addition of nitrogen and magnesium), and the depletion of the external carbon source collectively contribute to production of the de-fatted protein-rich biomass.
4.4. Concentration of Trace Minerals and Vitamins in Y. lipolytica Protein Biomass
Y. lipolytica protein biomass is a good source of macro-elements such as calcium, copper, phosphorus, potassium, and zinc and micro-elements such as chromium, iodine, and selenium, as shown in Table 4. Those elements are in bioavailable organic forms found in the biomass [19,24,38,46,69,70,71,72].
In a similar way to S. cerevisiae, Y. lipolytica is capable of assimilating minerals from the culture medium and incorporating them into its cell organic structures [40,66,69]. For example, chromium as a trace element is essential for the proper metabolism of glucose, insulin, fatty acids, proteins, and muscle growth. It is suspected that its adequate supply may reduce the fat content of pig carcasses. Minerals in yeast protein biomass, such as chromium or selenium, behave like chelates in the animal’s body. As a result of feeding organic forms with micronutrients, their absorption and use is better, which is of great physiological and ecological importance and reduces environmental contamination [69]. After a three-step sequential extraction (i.e., fractionation) and a simulated in vitro gastrointestinal digestion, it has been detected that in Y. lipolytica protein biomass, chromium occurred in the water-soluble part (50.2% of total Cr) and as polysaccharide-bound fraction of chromium (43.9% of total Cr) and protein-bound fraction (4.3% of total Cr). Moreover, the whole amount of Cr was trivalent (Cr (III)). The toxic form of Cr (IV) was not detected [104]. For human nutrition, according to EFSA recommendation [104], the maximum total Y. lipolytica chromium content of the product should be 23 μg/g, with the micro-nutrient present as Cr (III). The target population is general population from 3 years of age upwards, with maximum proposed use levels of 2 g/day for children from 3 to 9 years of age and 4 g/day thereafter. EFSA considered that chromium-enriched biomass of Y. lipolytica is safe under proposed condition of use.
In case of selenium from Y. lipolytica protein biomass, according to EFSA recommendation [104], the maximum total Se content in the products for children from 3 to 9 years of age is 40 μg Se/day and 200 μg Se/day thereafter. Se in the protein biomass is primarily presented as bioavailable organic selenium compounds: L-selenomethionine and L-selenocysteine, with low amounts in inorganic compounds as selenate and selenite [143]. It is worth emphasizing that the Se compounds provided by Y. lipolytica protein biomass are the same and as safe as those present in other dietary sources [103]. An advantageous feature of Y. lipolytica protein biomass is that it has low concentrations of sodium [40,69].
The protein biomass of the yeast is also a good source of vitamins, especially B-complex group. As shown in Table 5, protein biomass of Y. lipolytica cultured in industrial glycerol or biofuel waste contains a good amount of B vitamins, such as thiamine (vitamin B1), riboflavin (vitamin B2), pyridoxine (vitamin B6), folic acid (vitamin B9), and cyanocobalamin (vitamin B12) [4,5].
However, biotin (vitamin B7) may be produced in insufficient quantity (mean 20 μg biotin/100 g DW) by Y. lipolytica when cultivated in biofuel waste [5]. The yeast is able to synthesize the vitamins de novo or accumulate them ex novo (e.g., vitamin B12) from the aqueous environment in a similar manner to animal cells [4,5,70].
The bioavailability of dietary B vitamin-enriched Y. lipolytica biomass is very high [144,145]. These vitamins are water-soluble, and a surplus of them is generally excreted in urine [146]. Therefore, their doses must be much higher than the nutrient-recommended values (NRVs) [110] and, simultaneously, more consistent than that of the lipid-soluble vitamins. Interestingly, in developed countries, diets are impoverished of B vitamins [146]. The quantities of B vitamins in 100 g of the Y. lipolytica protein biomass completely cover the required NRVs for vitamin B1, B2, B6, B9, and B12. Hence, biomass enriched in B vitamins should be included in thediet for people who avoid eating meat and other animal products, particularly vegans and vegetarians, or those whose diet is poor in animal products. Deficiency of B vitamins, especially vitamin B12, leads to serious health problems [146,147,148,149,150].
5. Added-Value Compounds for Industrial Application
Y. lipolytica is considered as a ‘workhorse’ of a wide range of biotechnological production [54]. When the yeast is grown in various wastes and by-products, it is used as a producer of fatty acids [3,55,151]. It also produces lactones used as “fruity” aroma. Among them, the most interesting one is γ-decalactone, which is produced by the yeast when cultured in ricinoleic acid [152].
The yeast growing in various substrates under different conditions also produced some organic acids such as citric acid and isocitric acids [153,154,155,156,157,158,159,160,161,162,163,164], α-ketoglutaric acid (KGA) and pyruvic acid (PA) [27,56,165,166,167,168], and kynurenic acid (KYNA) [169]. Citric acid is commonly used as a flavoring agent, acidifier, antioxidant, and preservative in the food and pharmaceutical industries [140,170]. Of note, in wt strains, citric and isocitric acids are usually co-produced. However, the co-production of these acids is frequently not desired due to difficulties in their separation and receiving pure compounds. Therefore, some researchers tried to modulate citric acid/isocitric acid ratio either via the modulation of culture parameters such as substrate composition, micronutrients or pH, or mutant creation [167,171,172,173,174]. Similarly, KGA and PA are co-produced and realized in specific culture conditions, such as thiamine deficiency, low pH, and substrate degradation by glycolysis, impairing Krebs’s cycle. By contrast, when the yeast cultured in oily substrates (e.g., rapeseed oil), PA is not accumulated [168]. Furthermore, the selection of wt strains or the alteration of culture conditions were intensively assessed for obtaining KGA without PA [165,167]. KGA and PA are important in food, pharmaceutical, fodder, and other industries [175,176]. Y. lipolytica cultivated on honey also produces a tryptophan metabolite, KYNA, which exhibits pro-healthy properties due to its antioxidant, anti-inflammatory, and anticonvulsant action. KYNA also can protect the human brain and an inhibit the proliferation of colon cancer cells (i.e., adenocarcinoma HT-29 cells) [169].
Y. lipolytica is able to produce some polyols such as erythritol and mannitol commonly used as food additives for their flavor enhancer, sweetener, and humectant features [177,178,179,180]. The yeast polyols are produced in conditions such as high C/N ratio, high sugars or similarly metabolized compounds, low pH (3.0–3.5), and low oxygen [181]. In the conditions depending on strain used, it mainly produced mannitol in amount of max 30 g/L (around 80–88% of total polyols) [92,182]. However, when the yeast was cultured in glycerol, increasing pH to 5.5 for 72 h caused a shift in the yeast cell metabolism towards citric acid production (up to 40 g/L). Moreover, there are isolated strains which can produce both citric acid and polyols [182]. However, Y. lipolytica recombinant (MK1) produced up to 225 g of erythritol/L from glycerol [183]. Mutants generally can produce more polyols than wt strains [173,183,184,185,186], but recombinants have weaker adaptive properties to environmental changes in the culture medium than wt strains. When NaCl content was changed in the culture medium, the wt isolate produced higher mannitol concentration than the mutant strain (14 g/L and 4 g/L, respectively) [179,181,187]. Interestingly, it was noted that the produced polyols were re-utilized completely by the yeast as energy sources for growth needs when glycerol was exhausted [188].
Y. lipolytica is a graceful model to genetically modify to improve its metabolic productivity, increase biomass yields, and/or expand the substrates to be utilized [56,62,179,189]. Metabolic engineering of Y. lipolytica to produce bioactive compounds seems to be a good choice, especially when the yeast is employed to utilize common forestry and agricultural wastes such as cellulose, inulin, starch, or xylan, unused by wt strains [77,190], and/or to produce novel compounds that are not produced by the wt yeast. Engineered Y. lipolytica strains produce several heterologous valuable metabolites such as carotenoids [191,192]; terpenes, e.g., limonene [193]; polyketides with various biological activities; and aromatic amino-acid-derived molecules [194,195].
Y. lipolytica also has an operational secretory system with which it releases huge quantities of enzymes, such as lipases [196] and proteases [197], and heterologous proteins [27,198,199,200,201,202] for industrial applications. Among the more than 100 recombinant proteins produced by engineered Y. lipolytica [202], there are several pharmaceutically important human-compatible bio-compounds secreted by ‘humanized’ strains, including interferon alpha 2b [203], epidermal growth factor [204], N-glycoproteins [205,206], blood coagulation factor XIIIa [207], proinsulin and insulinotropin [208], cytochrome P450 [209], estrogen receptor α [210], and anaphylatoxin C5a that possesses both spasmogenic and leukocyte-related properties [211].
6. Safety of Y. lipolytica as a Valuable Source of Bioactive Compounds for Food
Dried and killed Y. lipolytica protein biomass is recognized as safe for human and animal nutrition in accordance with the current food and nutrition safety law. The protein biomass of Y. lipolytica cultivated in wastes such as industrial glycerol or biofuel waste did not contain excessive quantities of toxic minerals such as arsenic, cadmium, lead, and mercury. The contents of heavy metals were in low levels and did not exceed the standards [24,72,103,104]. The other parameters, including the levels of pathogens and total amount of contaminated microorganisms (i.e., mesophilic bacteria, yeast, and mold) and pesticides such as organochlorinated and organophosphate pesticides, pyrethroids, and other pesticides, were also below their limits of quantification [24,103,104].
It is worth emphasizing that Y. lipolytica is naturally occurring in foods and that it is not known to cause allergic reactions in humans [24,103]. Moreover, Y. lipolytica is not among the yeast species which have been noted to elicit allergic reactions in humans [212]. EFSA considered that given the protein content of the Y. lipolytica protein biomass (45–55 g protein/100 g DW), allergic reaction to the biomass cannot be excluded. However, the risk of allergic reactions is low [24,103,104].
Additionally, it is recognized that Y. lipolytica protein biomass is gluten-free. It is proven that the nutritional yeast products, i.e., the protein biomass, contained gluten at below the 20 mg/kg threshold defined by the Codex Alimentarius, The Codex, or the Gluten-Free Certification Organization (GFCO) [213].
7. Conclusions
Y. lipolytica biomass is a valuable source of bioactive compounds such as protein with an appropriate spectrum of amino acids and low amounts of fats and sodium. The protein biomass is also rich in trace minerals; vitamins, especially B-group vitamins; and other valuable compounds. The nutritional protein biomass may be prepared as dietary supplements in the forms of capsules, tablets, and sachets or be used as food additive to improve nutritional quality food and feed. The use of the nutritional protein biomass is an excellent alternative for traditional nutrition sources such as plants and animals. Products from protein biomass are especially intended for people at risk of vitamin B deficiency who avoid animal products, including vegans and vegetarians, and those with low intake of animal foods, such as populations who do not consume animal products due to culture, conviction, poverty, or those with restrictive diet patterns, as well as athletes and people after recovery. These protein biomasses are obtained in a short time with the use of various wastes in a small land area; throughout the whole year and with a low environmental footprint; and independently from climatic and weather conditions and fresh water availability. Moreover, engineering Y. lipolytica creates new perspectives in terms of its use for the production of novel bioactive compounds, especially recombinant therapeutic proteins.
Author Contributions
Conceptualization, M.E.J. and A.M.; methodology, M.E.J.; software, M.E.J.; validation—formal analysis, M.E.J.; investigation, resources, data curation, M.E.J.; writing—original draft preparation, M.E.J.; writing—review and editing, A.M.; visualization, M.E.J.; supervision, A.M.; project administration, A.M.; funding acquisition, A.M. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Bellou, S.; Triantaphyllidou, I.-E.; Aggeli, D.; Elazzazy, A.M.; Baeshen, M.N.; Aggelis, G. Microbial oils as food additives: Recent approaches for improving microbial oil production and its polyunsaturated fatty acid content. Curr. Opin. Biotechnol. 2016, 37, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Beopoulos, A.; Nicaud, J.-M.; Gaillardin, C. An overview of lipid metabolism in yeasts and its impact on biotechnological processes. Appl. Microbiol. Biotechnol. 2011, 90, 1193–1206. [Google Scholar] [CrossRef] [PubMed]
- Dourou, M.; Aggeli, D.; Papanikolaou, S.; Aggelis, G. Critical steps in carbon metabolism affecting lipid accumulation and their regulation in oleaginous microorganisms. Appl. Microbiol. Biotechnol. 2018, 102, 2509–2523. [Google Scholar] [CrossRef]
- Jach, M.E.; Masłyk, M.; Juda, M.; Sajnaga, E.; Malm, A. Vitamin B12-enriched Yarrowia lipolytica biomass obtained from biofuel waste. Waste Biomass Valorization 2020, 11, 1711–1716. [Google Scholar] [CrossRef] [Green Version]
- Jach, M.E.; Sajnaga, E.; Janeczko, M.; Juda, M.; Kochanowicz, E.; Baj, T.; Malm, A. Production of enriched in B vitamins biomass of Yarrowia lipolytica grown in biofuel waste. Saudi J. Biol. Sci. 2021, 28, 2925–2932. [Google Scholar] [CrossRef]
- Jach, M.E.; Sajnaga, E.; Świder, R.; Baier, A.; Mickowska, B.; Juda, M.; Chudzik-Rząd, B.; Szyszka, R.; Malm, A. Yarrowia lipolytica grown on biofuel waste as a source of single cell protein and essential amino acids for human diet. Saudi J. Med. Pharm. Sci. 2017, 3, 1344–1351. [Google Scholar]
- Liu, H.H.; Ji, X.J.; Huang, H. Biotechnological applications of Yarrowia lipolytica: Past, present and future. Biotechnol. Adv. 2015, 33, 1522–1546. [Google Scholar] [CrossRef]
- Lopes, M.; Gomes, A.S.; Silva, C.M.; Bel, I. Microbial lipids and added value metabolites production by Yarrowia lipolytica from pork lard. J. Biotechnol. 2018, 265, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Lopes, M.; Miranda, S.M.; Alves, J.M.; Pereira, A.S.; Belo, I. Waste cooking oils as feedstock for lipase and lipid-rich biomass production. Eur. J. Lipid Sci. Technol. 2019, 121, 1800188. [Google Scholar] [CrossRef] [Green Version]
- Papanikolaou, S.; Aggelis, G. Lipids of oleaginous yeasts. Part I: Biochemistry of single cell oil production. Eur. J. Lipid Sci. Technol. 2011, 113, 1031–1051. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Aggelis, G. Yarrowia lipolytica: A model microorganism used for the production of tailor-made lipids. Eur. J. Lipid Sci. Technol. 2010, 112, 639–654. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Aggelis, G.; Marc, I. Kinetic profile of the cellular lipid composition in an oleaginous Yarrowia lipolytica capable of producing a cocoa-butter substitute from industrial fats. Antonie Leeuwenhoek 2001, 80, 215–224. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Muniglia, L.; Chevalot, I.; Aggelis, G.; Marc, I. Accumulation of a cocoa-butter-like lipid by Yarrowia lipolytica cultivated on agro-industrial residues. Curr. Microbiol. 2003, 46, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Rywińska, A.; Juszczyk, P.; Wojtatowicz, M.; Robak, M.; Lazar, Z.; Tomaszewska, L.; Rymowicz, W. Glycerol as a promising substrate for Yarrowia lipolytica biotechnological applications. Biomass Bioenerg. 2013, 48, 148–166. [Google Scholar] [CrossRef]
- Bankar, A.V.; Kumar, A.R.; Zinjarde, S.S. Environmental and industrial applications of Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2009, 84, 847–865. [Google Scholar] [CrossRef]
- Jach, M.E.; Serefko, A.; Sajnaga, E.; Kozak, E.; Poleszak, E.; Malm, A. Dietary supplements based on the yeast biomass. Curr. Top. Nutraceutical Res. 2015, 13, 83–88. [Google Scholar]
- Katre, G.; Joshi, C.; Khot, M.; Zinjarde, S.; RaviKumar, A. Evaluation of single cell oil (SCO) from a tropical marine yeast Yarrowia lipolytica NCIM 3589 as a potential feedstock for biodiesel. AMB Expr. 2012, 2, 36. [Google Scholar] [CrossRef] [Green Version]
- Vuong, M.D.; Thanh, N.T.; Son, C.K.; Yves, W. Protein enrichment of cassava-based dried distiller’s grain by solid state fermentation using Trichoderma Harzianum and Yarrowia Lipolytica for feed ingredients. Waste Biomass Valorization 2021, 12, 3875–3888. [Google Scholar] [CrossRef]
- Jach, M.E.; Serefko, A. Nutritional yeast biomass: Characterization and application. In Diet, Microbiome and Health. Handbook of Food Bioengineering, 1st ed.; Grumezescu, A., Holban, A.M., Eds.; Academic Press: London, UK; Elsevier: San Diego, CA, USA; Cambridge, MA, USA, 2018; Volume 11, pp. 237–270. [Google Scholar]
- Kieliszek, M.; Kot, A.M.; Bzducha-Wróbel, A.; Błażejak, S.; Gientka, I.; Kurcz, A. Biotechnological use of Candida yeasts in the food industry: A review. Fungal Biol. Rev. 2017, 31, 185–198. [Google Scholar] [CrossRef]
- Santos, C.A.; Caldeira, M.L.; Lopes da Silva, T.; Novais, J.M.; Reis, A. Enhanced lipidic algae biomass production using gas transfer from a fermentative Rhodosporidium toruloides culture to an autotrophic Chlorella protothecoides culture. Bioresour. Technol. 2013, 138, 48–54. [Google Scholar] [CrossRef] [Green Version]
- Beopoulos, A.; Desfougeres, T.; Sabirova, J.; Zinjarde, S.; Neuvéglise, C.; Nicaud, J.-M. The Hydrocarbon-Degrading Oleaginous Yeast Yarrowia lipolytica. In Handbook of Hydrocarbon and Lipid Microbiology, 1st ed.; Timmis, K.N., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 1, pp. 2111–2121. [Google Scholar] [CrossRef]
- Devillers, H.; Brunel, F.; Polomska, X.; Sarilar, V.; Lazar, Z.; Robak, M.; Neuveglise, C. Draft genome sequence of Yarrowia lipolytica strain A-101 isolated from polluted soil in Poland. Genome Announc. 2016, 4, e01094-16. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Safety of Yarrowia lipolytica yeast biomass as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2019, 17, 5594. [Google Scholar] [CrossRef]
- Thevenieau, F.; Beopoulos, A.; Desfougeres, T.; Sabirova, J.; Albertin, K.; Zinjarde, S.; Nicaud, J.-M. Uptake and assimilation of hydrophobic substrates by the oleaginous yeast Yarrowia lipolytica. In Handbook of Hydrocarbon and Lipid Microbiology, 1st ed.; Timmis, K.M., Ed.; Springer: Berlin/Heidelberg, Germany, 2010; Volume 1, pp. 1513–1527. [Google Scholar]
- Wojtatowicz, M.; Rymowicz, W. Effect of inoculum on kinetics and yield of citric acid production on glucose media by Yarrowia lipolytica A-101. Acta Aliment. Pol. 1991, 41, 137–143. [Google Scholar]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; van Dijk, P.W.M.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef]
- Lanciotti, R.; Vannini, L.; Lopez, C.C.; Gobbetti, M.; Guerzoni, E. Evaluation of the ability of Yarrowia lipolytica to impart strain-dependent characteristics to cheese when used as a ripening adjunct. Int. J. Dairy Technol. 2005, 58, 89–99. [Google Scholar] [CrossRef]
- Nicaud, J.-M. Yarrowia lipolytica. Yeast 2012, 29, 409–418. [Google Scholar] [CrossRef]
- Irby, R.F.; Kandula, M.; Zadikany, R.; Sandin, R.; Greene, J. Yarrowia lipolytica as normal human flora: A case series of 24 patients with positive cultures and no attributable disease. Infect. Dis. Clin. Pract. 2014, 22, 207–209. [Google Scholar] [CrossRef]
- Zieniuk, B.; Fabiszewska, A. Yarrowia lipolytica: A beneficious yeast in biotechnology as a rare opportunistic fungal pathogen: A minireview. World J. Microbiol. Biotechnol. 2019, 35, 10. [Google Scholar] [CrossRef] [Green Version]
- Madzak, C. Yarrowia lipolytica strains and their biotechnological applications: How natural biodiversity and metabolic engineering could contribute to cell factories improvement. J. Fungi 2021, 7, 548. [Google Scholar] [CrossRef]
- Berge, G.M.; Hatlen, B.; Odom, J.M.; Ruyter, B. Physical treatment of high EPA Yarrowia lipolytica biomass increases the availability of n-3 highly unsaturated fatty acids when fed to Atlantic salmon. Aquac. Nutr. 2013, 19, 110–121. [Google Scholar] [CrossRef]
- Stefańska, B.; Komisarek, J.; Stanisławski, D.; Gąsiorek, M.; Kasprowicz-Potocka, M.; Frankiewicz, A.; Nowak, W. The effect of Yarrowia lipolytica culture on growth performance, ruminal fermentation and blood parameters of dairy calves. Anim. Feed Sci. Technol. 2018, 243, 72–79. [Google Scholar] [CrossRef]
- Neuls, L.; de Souza, V.J.; Romao, S.; Bitencourt, T.B.; Ramos, C.J.R.; Parra, J.E.G.; Cazarolli, L.H. Immunomodulatory effects of Yarrowia lipolytica as food additive in the diet of Nile tilapia. Fish Shellfish Immunol. 2021, 119, 272–279. [Google Scholar] [CrossRef]
- Alamillo, E.; Reyes-Becerril, M.; Cuesta, A.; Angulo, C. Marine yeast Yarrowia lipolytica improves the immune responses in Pacific red snapper (Lutjanus peru) leukocytes. Fish Shellfish Immunol. 2017, 70, 48–56. [Google Scholar] [CrossRef]
- Czech, A.; Merska, M.; Ognik, K. Blood immunological and biochemical indicators in turkey hens fed diets with a different content of the yeast Yarrowia lipolytica. Ann. Anim. Sci. 2014, 14, 935–946. [Google Scholar] [CrossRef] [Green Version]
- Merska, M.; Czech, A.; Ognik, K. The effect of yeast Yarrowia lipolytica on the antioxidant indices and macro-and microelements in blood plasma of turkey hens. Pol. J. Vet. Sci. 2015, 18, 709–714. [Google Scholar] [CrossRef] [Green Version]
- Merska-Kazanowska, M.; Czech, A.; Ognik, K. The effect of different doses of dried yeast Yarrowia lipolytica on production effects of turkey hens and hematological indicators of blood. Ann. Univ. Mariae Curie-Skłodowska Sect. EE Zootech. 2013, 31, 35–41. [Google Scholar]
- Czech, A.; Smolczyk, A.; Ognik, K.; Kiesz, M. Nutritional value of Yarrowia lipolytica yeast and its effect on growth performance indicators in piglets. Ann. Anim. Sci. 2016, 16, 1091–1100. [Google Scholar] [CrossRef] [Green Version]
- Czech, A.; Smolczyk, A.; Grela, E.R.; Kiesz, M. Effect of dietary supplementation with Yarrowia lipolytica or Saccharomyces cerevisiae yeast and probiotic additives on growth performance, basic nutrients digestibility and biochemical blood profile in piglets. J. Anim. Physiol. Anim. Nutr. 2018, 102, 1720–1730. [Google Scholar] [CrossRef]
- Czech, A.; Smolczyk, A.; Ognik, K.; Wlazło, Ł.; Nowakowicz-Dębek, B.; Kiesz, M. Effect of dietary supplementation with Yarrowia lipolytica or Saccharomyces cerevisiae yeast and probiotic additives on haematological parameters and the gut microbiota in piglets. Res. Vet. Sci. 2018, 119, 221–227. [Google Scholar] [CrossRef]
- Guardiola, F.A.; Esteban, M.Á.; Angulo, C. Yarrowia lipolytica, health benefits for animals. Appl. Microbiol. Biotechnol. 2021, 105, 7577–7592. [Google Scholar] [CrossRef]
- Moore, M.; Gould, P.; Keary, B.S. Global urbanization and impact on health. Int. J. Hyg. Environ. Health 2003, 206, 269–278. [Google Scholar] [CrossRef] [PubMed]
- United Nations. World Population Prospects 2019. Available online: https://www.un.org/development/desa/en/news/population/world-population-prospects-2019.html (accessed on 17 June 2019).
- Jach, M.E.; Serefko, A.; Ziaja, M.; Kieliszek, M. Yeast protein as an easily accessible food source. Metabolites 2022, 12, 63. [Google Scholar] [CrossRef] [PubMed]
- Barth, G.; Gaillardin, C. Physiology and genetics of the dimorphic fungus Yarrowia lipolytica. FEMS Microbiol. Rev. 1997, 19, 219–237. [Google Scholar] [CrossRef] [PubMed]
- Yarrow, D. Four new combinations in yeasts. Antonie Leeuwenhoek 1972, 38, 357–360. [Google Scholar] [CrossRef]
- Kurtzman, C.P. New species and a new combination in the Hyphopichia and Yarrowia yeast clades. Antonie Leeuwenhoek 2005, 88, 121–130. [Google Scholar] [CrossRef]
- Michely, S.; Gaillardin, C.; Nicaud, J.-M.; Neuveglise, C. Comparative physiology of oleaginous species from the Yarrowia clade. PLoS ONE 2013, 8, e63356. [Google Scholar] [CrossRef]
- Rakicka-Pustułka, M.; Miedzianka, J.; Jama, D.; Kawalec, S.; Liman, K.; Janek, T.; Skaradziński, G.; Rymowicz, W.; Lazar, Z. High value-added products derived from crude glycerol via microbial fermentation using Yarrowia clade yeast. Microb. Cell Factories 2021, 20, 195. [Google Scholar] [CrossRef]
- Thevenieau, F.; Gaillardin, C.; Nicaud, J.-M. Applications of the non-conventional yeast Yarrowia lipolytica. In Diversity and Potential Biotechnological Applications of Yeasts; Kunze, G., Satyanarayana, T., Eds.; Elsevier: Amsterdam, The Netherlands, 2009; Volume 1, pp. 589–613. [Google Scholar]
- Amata, I.A. Yeast a single cell protein: Characteristics and metabolism. Int. J. Appl. Biol. Pharm. Technol. 2013, 4, 158–170. [Google Scholar]
- Coelho, M.A.Z.; Amaral, P.; Belo, I. Yarrowia lipolytica: An industrial workhorse. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Mendez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; pp. 930–944. [Google Scholar]
- Nandy, S.K.; Srivastava, R.K. A review on sustainable yeaste biotechnological processes and applications. Microbiol. Res. 2018, 207, 83–90. [Google Scholar] [CrossRef]
- Gottardi, D.; Siroli, L.; Vannini, L.; Patrignani, F.; Lanciotti, R. Recovery and valorization of agri-food wastes and by-products using the non-conventional yeast Yarrowia lipolytica. Trends Food Sci. Technol. 2021, 115, 74–86. [Google Scholar] [CrossRef]
- Pérez-Campo, F.; Domínguez, A. Factors Affecting the Morphogenetic Switch in Yarrowia lipolytica. Curr. Microbiol. 2001, 43, 429–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusane, D.H.; Nancharaiah, Y.V.; Venugopalan, V.P.; Kumar, A.R.; Zinjarde, S.S. Biofilm formation by biotechnologically important tropical marine yeast isolate, Yarrowia lipolytica NCIM 3589. Water Sci. Technol. 2008, 58, 1221–1229. [Google Scholar] [CrossRef] [PubMed]
- Ledesma-Amaro, R.; Nicaud, J.-M. Yarrowia lipolytica as a biotechnological chassis to produce usual and unusual fatty acids. Prog. Lipid Res. 2016, 61, 40–50. [Google Scholar] [CrossRef]
- Kurtzman, C.P. Phylogeny of the ascomycetous yeasts and the renaming of Pichia anomala to Wickerhamomyces anomalus. Antonie Leeuwenhoek 2011, 99, 13–23. [Google Scholar] [CrossRef]
- Taskin, M.; Saghafi, A.; Aydogan, M.N.; Arslan, N.P. Microbial lipid production by cold-adapted oleaginous yeast Yarrowia lipolytica B9 in non-sterile whey medium. Biofuels Bioprod. Biorefin. 2015, 9, 595–605. [Google Scholar] [CrossRef]
- Spagnuolo, M.; Shabbir Hussain, M.; Gambill, L.; Blenner, M. Alternative Substrate Metabolism in Yarrowia lipolytica. Front. Microbiol. 2018, 25, 1077. [Google Scholar] [CrossRef]
- Gao, R.; Li, Z.; Zhou, X.; Cheng, S.; Zheng, L. Oleaginous yeast Yarrowia lipolytica culture with synthetic and food waste-derived volatile fatty acids for lipid production. Biotechnol. Biofuels 2017, 10, 247. [Google Scholar] [CrossRef]
- Rodrigues, G.; Pais, C. The influence of acetic and other weak carboxylic acids on growth and cellular death of the yeast Yarrowia lipolytica. Food Technol. Biotechnol. 2000, 12, 579–1006. [Google Scholar]
- Mansour, S.; Beckerich, J.M.; Bonnarme, P. Lactate and amino acid catabolism in the cheese-ripening yeast Yarrowia lipolytica. Appl. Environ. Microbiol. 2008, 74, 6505–6512. [Google Scholar] [CrossRef] [Green Version]
- Patsios, S.I.; Dedousi, A.; Sossidou, E.N.; Zdragas, A. Sustainable animal feed protein through the cultivation of Yarrowia lipolytica on agro-industrial wastes and by-products. Sustainability 2020, 12, 1398. [Google Scholar] [CrossRef] [Green Version]
- Drzymała, K.; Mirończuk, A.M.; Pietrzak, W.; Dobrowolski, A. Rye and oat agricultural wastes as substrate candidates for biomass production of the non-conventional yeast Yarrowia lipolytica. Sustainability 2020, 12, 7704. [Google Scholar] [CrossRef]
- Jach, M.E.; Baj, T.; Juda, M.; Świder, R.; Mickowska, B.; Malm, A. Statistical evaluation of growth parameters in biofuel waste as a culture medium for improved production of single cell protein and amino acids by Yarrowia lipolytica. AMB Expr. 2020, 10, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Woźnica, A.; Czech, A. Utilization of glycerol and yeast biomass produced from glycerol in the animal nutrition. Post Nauk. Roln. 2010, 62, 136–140. [Google Scholar]
- Juszczyk, P.; Smolczyk, A.; Gil, Z.; Rywińska, A.; Rymowicz, W. Produkcja drożdży paszowych Yarrowia lipolytica wzbogaconych w aminokwasy selenowe i witaminę B12. Inż. Ap. Chem. 2015, 54, 165–166. [Google Scholar]
- Musiał, I.; Juszczyk, P.; Rymowicz, W.; Kinal, S. Production of selenium and chromium enriched Yarrowia lipolytica fodder yeast. Acta Sci. Pol. 2005, 4, 55–64. [Google Scholar]
- Michalik, B.; Biel, W.; Lubowicki, R.; Jacyno, E. Chemical composition and biological value of proteins of the yeast Yarrowia lipolytica growing on industrial glycerol. Can. J. Anim. Sci. 2014, 94, 99–104. [Google Scholar] [CrossRef] [Green Version]
- Chi, Z.; Wang, F.; Wang, L.; Li, J.; Wang, X. Selection of Yarrowia lipolytica strains with high protein content from yeasts isolated from different marine environments. J. Ocean Univ. China 2007, 6, 360–364. [Google Scholar] [CrossRef]
- Juszczyk, P.; Tomaszewska, L.; Kira, A.; Rymowicz, W. Biomass production by novel strain of Yarrowia lipolytica using raw glicerol, derived from biodiesel production. Bioresour. Technol. 2013, 137, 124. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chevalot, I.; Galiotou-Panayotou, M.; Komaitis, M. Industrial derivative of tallow: A promising renewable substrate for microbial lipid, single-cell protein and lipase production by Yarrowia lipolytica. Electron. J. Biotechnol. 2007, 10, 425–435. [Google Scholar] [CrossRef] [Green Version]
- Zhao, M.-X.; Chi, Z.; Chi, Z.-M.; Madzak, C. The simultaneous production of single-cell protein and a recombinant antibacterial peptide by expression of an antibacterial peptide gene in Yarrowia lipolytica. Process Biochem. 2013, 48, 212–217. [Google Scholar] [CrossRef]
- Cui, W.; Wang, Q.; Zhang, F.; Zhang, S.-C.; Chi, Z.-M.; Madzak, C. Direct conversion of inulin into single cell protein by the engineered Yarrowia lipolytica carrying inulinase gene. Process Biochem. 2011, 46, 1442–1448. [Google Scholar] [CrossRef]
- Mondal, A.K.; Sengupta, S.; Bhowal, J.; Bhattacharya, D.K. Utilization of fruit wastes in producing single cell protein. Int. J. Environ. Sci. 2012, 1, 430–438. [Google Scholar]
- Hezarjaribi, M.; Ardestani, F.; Ghorbani, R.H. Single cell protein production by Saccharomyces cerevisiae using an optimized culture medium composition in a batch submerged bioprocess. Appl. Biochem. Biotechnol. 2016, 179, 1336–1345. [Google Scholar] [CrossRef] [PubMed]
- Yunus, F.-N.; Nadeem, M.; Rashid, F. Single-cell protein production through microbial conversion of lignocellulosic residue (wheat bran) for animal feed. J. Inst. Brew. 2015, 121, 553–557. [Google Scholar] [CrossRef] [Green Version]
- Yunus, F.-N.; Nadeem, M.; Rashid, F. Enhancement of protein contents of rice bran for animal feed by solid state fermentation. Biomed. Lett. 2015, 1, 31–36. [Google Scholar]
- Polpass, A.J.; Kunjukrishnan, K.S.; Solomon, R.D.J. Formulation and statistical optimization of culture medium for improved production of antimicrobial compound by Streptomyces sp. JAJ06. Int. J. Microbiol. 2013, 2013, 526260. [Google Scholar] [CrossRef] [Green Version]
- Shahabadi, S.M.S.; Reyhani, A. Optimization of operating conditions in ultrafiltration process for produced water treatment via the full factorial design methodology. Sep. Purif. Technol. 2014, 132, 50–61. [Google Scholar] [CrossRef]
- Wang, W.; Wei, H.; Alahuhta, M.; Chen, X.; Hyman, D.; Johnson, D.K.; Zhang, M.; Himmel, M.E. Heterologous expression of xylanase enzymes in lipogenic yeast Yarrowia lipolytica. PLoS ONE 2014, 9, e111443. [Google Scholar] [CrossRef] [Green Version]
- Adedayo, M.R.; Ajiboye, E.A.; Akintunde, J.K.; Odaibo, A. single cell proteins: As nutritional enhncer. Adv. Appl. Sci. Res. 2011, 2, 396–409. [Google Scholar]
- Agboola, J.O.; Øverland, M.; Skrede, A.; Hansen, J.Ø. Yeast as major protein-rich ingredient in aquafeeds: A review of the implications for aquaculture production. Rev. Aquac. 2021, 13, 949–970. [Google Scholar] [CrossRef]
- Saygün, A.; Sahin-Yesilcubuk, N.; Aran, N. Effects of different oil sources and residues on biomass and metabolite production by Yarrowia lipolytica YB 423-12. J. Am. Oil Chem. Soc. 2014, 91, 1521–1530. [Google Scholar] [CrossRef]
- Tzirita, M.; Papanikolaou, S.; Chatzifragkou, A.; Quilty, B. Waste fat biodegradation and biomodification by Yarrowia lipolytica and a bacterial consortium composed of Bacillus spp. and Pseudomonas putida. Eng. Life Sci. 2018, 18, 932–942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vasiliadou, I.; Bellou, S.; Daskalaki, A.; Tomaszewska-Hetman, L.; Chatzikotoula, C.; Kompoti, B.; Papanikolaou, S.; Vayenas, D.; Pavlou, S.; Aggelis, G. Biomodification of fats and oils and scenarios of adding value on renewable fatty materials through microbial fermentations: Modelling and trials with Yarrowia lipolytica. J. Clean. Prod. 2018, 200, 1111–1129. [Google Scholar] [CrossRef]
- Ukaegbu-Obi, K.M. Single cell protein: A resort to global protein challenge and waste management. J. Microbiol. Microb. Technol. 2016, 1, 5. [Google Scholar]
- Srividya, A.R.; Vishnuvarthan, V.J.; Murugappan, M.; Dahake, P.G. Single cell protein- A review. Int. J. Pharm. Res. Sch. 2013, 2, 472–485. [Google Scholar]
- Filippousi, R.; Antoniou, D.; Tryfinopoulou, P.; Nisiotou, A.A.; Nychas, G.-J.; Koutinas, A.A.; Papanikolaou, S. Isolation, identification and screening of yeasts towards their ability to assimilate biodiesel-derived crude glycerol: Microbial production of polyols, endopolysaccharides and lipid. J. Appl. Microbiol. 2019, 127, 1080–1100. [Google Scholar] [CrossRef]
- Daskalaki, A.; Perdikouli, N.; Aggeli, D.; Aggelis, G. Laboratory evolution strategies for improving lipid accumulation in Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2019, 103, 8585–8596. [Google Scholar] [CrossRef]
- Kurbanoglu, E.B. Production of single cell protein from ram horn hydrolysate. Turk. J. Biol. 2001, 25, 371–377. [Google Scholar] [CrossRef]
- Gao, Y.; Li, D.; Liu, Y. Production of single cell protein from soy molasses using Candida tropicalis. Ann. Microbiol. 2012, 62, 1165–1172. [Google Scholar] [CrossRef]
- Øverland, M.; Karlsson, A.; Mydland, L.T.; Romarheim, O.H.; Skrede, A. Evaluation of Candida utilis, Kluyveromyces marxianus and Saccharomyces cerevisiae yeasts as protein sources in diets for Atlantic salmon (Salmon salar). Aquaculture 2013, 402–403, 1–7. [Google Scholar] [CrossRef]
- Ritala, A.; Häkkinen, S.T.; Toivari, M.; Wiebe, M.G. Single cell protein -state-of-the-art, industrial landscape and patents 2001–2016. Front. Microbiol. 2017, 8, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Hames, E.E.; Demir, T. Microbial ribonucleases (RNases): Production and application potential. World J. Microbiol. Biotechnol. 2015, 31, 1853–1862. [Google Scholar] [CrossRef] [PubMed]
- Knight, N.; Roberts, G.; Shelton, D. The thermal stability of QuornTM pieces. Int. J. Food Sci. Technol. 2001, 36, 47–52. [Google Scholar] [CrossRef]
- Yadav, J.S.S.; Yan, S.; Ajila, C.M.; Bezawada, J.; Tyagi, R.D.; Surampalli, R.Y. Food-grade single-cell protein production, characterization and ultrafiltration recovery of residual fermented whey proteins from whey. Food Bioprod. Process. 2016, 99, 156–165. [Google Scholar] [CrossRef]
- Ibarz, A.; Barbosa-Cánovas, G.V. Introduction to Unit Operations: Fundamental Concepts. In Introduction to Food Process Engineering. Food Preservation Technology Series, 1st ed.; Ibarz, A., Barbosa-Cánovas, G.V., Eds.; Taylor and Francis Group CRC Press: Boca Raton, FL, USA, 2014; Volume 1, pp. 4–12. [Google Scholar]
- Baudelaire, E.D. Grinding for food powder production. In Handbook of Food Powders Processes and Properties, 1st ed.; Bhandari, B., Bansal, N., Zhang, M., Schuck, P., Eds.; Woodhead Publishing: Cambridge, UK, 2013; Volume 1, pp. 132–149. [Google Scholar]
- EFSA. Safety of selenium-enriched biomass of Yarrowia lipolytica as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2020, 18, 5992. [Google Scholar]
- EFSA. Safety of chromium-enriched biomass of Yarrowia lipolytica as a novel food pursuant to Regulation (EU) 2015/2283. EFSA J. 2020, 18, 6005. [Google Scholar]
- Cheslo, F. Small animal nutrition. In Mosby’s Comprehensive Review for Veterinary Technicians, 3rd ed.; Tighe, M.M., Brown, M., Eds.; Mosby Elsevier: St. Louis, MO, USA, 2008; Volume 1, pp. 243–262. [Google Scholar]
- Millward, D.J. Amino acid scoring patterns for protein quality assessment. Br. J. Nutr. 2012, 108, S31–S43. [Google Scholar] [CrossRef] [Green Version]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific opinion on the safety of ‘yeast beta-glucans’ as a novel food ingredient. EFSA Panel on Dietetic Products. Nutrition and Allergies (NDA). EFSA J. 2011, 9, 2137. [Google Scholar] [CrossRef]
- Bacha, U.; Nasir, M.; Khalique, A.; Anjum, A.A.; Jabbar, M.A. Comparative assessment of various agro-industrial wastes for Saccharomyces cerevisiae biomass production and its quality evaluation as single cell protein. J. Anim. Plant Sci. 2011, 21, 844–849. [Google Scholar]
- Eggum, B.O. Protein and energy digestibility. In The Rat as a Model for Man and Pig in Nutritional and Physiological Studies; Jungvid, H., Forshel, L.P., Eggum, B.O., Eds.; Gramineer AB: Copenhagen, Denmark; AB AnalyCen/National Institute of Animal Science: Copenhagen, Denmark, 1990; pp. 77–86. [Google Scholar]
- European Parliament and Council. Regulation (EU) No 1169/2011 of the European Parliament and of the Council of 25 October 2011 on the provision of food information to costumers. OJEU 2011, 304, 18–63. [Google Scholar]
- Esteban, P.F.; Vazquez de Aldana, C.R.; del Rej, F. Cloning and characterization of 1,3-beta-glucanase-encoding genes from non-conventional yeasts. Yeast 1999, 15, 1631–1644. [Google Scholar] [CrossRef]
- Barber, T.M.; Kabisch, S.; Weickert, M.O. The Health Benefits of Dietary Fibre. Nutrients 2020, 12, 3209. [Google Scholar] [CrossRef] [PubMed]
- Attia, Y.A.; Al-Harthi, M.A.; Korish, M.A.; Shiboob, M.H. Protein and amino acid content in four brands of commercial table eggs in retail markets in relation to human requirements. Animals 2020, 10, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ismail, M.; Mariod, A.; Pin, S.S. Effects of preparation methods on protein and amino acid contents of various eggs available in Malaysian local markets. Acta Sci. Pol. Technol. Aliment. 2013, 12, 21–31. [Google Scholar] [PubMed]
- Khan, M.S.; Ali, E.; Ali, S.; Khan, W.M.; Sajjad, M.A.; Hussain, F. Assessment of essential amino acids in wheat proteins: A case study. J. Biodivers. Environ. Sci. 2014, 4, 185–189. [Google Scholar]
- Landi, N.; Ragucci, S.; Di Maro, A. Amino acid composition of milk from cow, sheep and goat raised in Ailano and Valle Agricola, two localities of ‘Alto Casertano’ (Campania Region). Foods 2021, 10, 2431. [Google Scholar] [CrossRef]
- Bhutta, Z.A.; Sadiq, K. Protein Deficiency. In Encyclopedia of Human Nutrition, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2013; pp. 111–115. [Google Scholar] [CrossRef]
- Biel, W.; Kazimierska, K.; Bashutska, U. Nutritional value of wheat, triticale, barley and oat grains. Acta Sci. Pol. Zootech. 2020, 19, 19–28. [Google Scholar] [CrossRef]
- Zhao, L.; Li, B.; Xiong, D.; Zhang, H.; Tang, X.; Zhang, H.; Song, Y.; Yang, S. Cocoa-butter-equivalent production from Yarrowia lipolytica by optimization of fermentation technology. Am. J. Biochem. Biotechnol. 2016, 12, 196–205. [Google Scholar] [CrossRef] [Green Version]
- Kantar, S.E.; Khelfa, A.; Vorobiev, E.; Koubaa, M. Strategies for increasing lipid accumulation and recovery from Y. lipolytica: A review. OCL 2021, 28, 51. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Galiotou-Panayotou, M.; Chevalot, I.; Komaitis, M.; Marc, I.; Aggelis, G. Influence of glucose and saturated free-fatty acid mixtures on citric acid and lipid production by Yarrowia lipolytica. Curr. Microbiol. 2006, 52, 134–142. [Google Scholar] [CrossRef]
- Darvishi, F.; Nahvi, I.; Zarkesh-Esfahani, H.; Momenbeik, F. Effect of plant oils upon lipase and citric acid production in Yarrowia lipolytica yeast. J. Biomed. Biotechnol. 2009, 2009, 562943. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diamantopoulou, P.; Filippousi, R.; Antoniou, D.; Varfi, E.; Xenopoulos, E.; Sarris, D.; Papanikolaou, S. Production of added-value microbial metabolites during growth of yeast strains on media composed of biodiesel-derived crude glycerol and glycerol/xylose blends. FEMS Microbiol. Lett. 2020, 367, fnaa063. [Google Scholar] [CrossRef] [PubMed]
- Kuttiraja, M.; Dhouha, A.; Tyagi, R.D. Harnessing the Effect of pH on lipid production in batch cultures of Yarrowia lipolytica SKY7. Appl. Biochem. Biotechnol. 2018, 184, 1332–1346. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.L.; Chi, Z.; Wang, G.Y.; Wang, Z.P.; Li, Y.; Chi, Z.M. Yeast killer toxins, molecular mechanisms of their action and their applications. Crit. Rev. Biotechnol. 2015, 35, 222–234. [Google Scholar] [CrossRef] [PubMed]
- Rakicka, M.; Lazar, Z.; Dulermo, T.; Fickers, P.; Nicaud, J.M. Lipid production by the oleaginous yeast Yarrowia lipolytica using industrial by-products under different culture conditions. Biotechnol. Biofuels 2015, 8, 104. [Google Scholar] [CrossRef] [Green Version]
- Brígida, A.I.S.; Amaral, P.F.F.; Coelho, M.A.Z.; Gonçalves, L.R.B. Lipase from Yarrowia lipolytica: Production, characterization and application as an industrial biocatalyst. J. Mol. Catal. B 2014, 101, 148–158. [Google Scholar] [CrossRef]
- Kumari, A.; Gupta, R. Purification and Biochemical Characterization of a novel magnesium dependent lipase from Trichosporon asahii MSR 54 and its application in biodiesel production. Asian J. Biotechnol. 2012, 4, 70–82. [Google Scholar] [CrossRef] [Green Version]
- Fickers, P.; Marty, A.; Nicaud, J.M. The lipases from Yarrowia lipolytica: Genetics, production, regulation, biochemical characterization and biotechnological applications. Biotechnol. Adv. 2011, 29, 632–644. [Google Scholar] [CrossRef]
- Dellomonaco, C.; Rivera, C.; Campbell, P.; Gonzalez, R. Engineered respire-fermentative metabolism for the production of biofuels and biochemical from fatty acid-rich feedstock. Appl. Environ. Microbiol. 2010, 76, 5067–5078. [Google Scholar] [CrossRef] [Green Version]
- Haddouche, R.; Delessert, S.; Sabirova, J.; Neuveglise, C.; Poirier, Y.; Nicaud, J.-M. Roles of multiple acyl-CoA oxidases in the routing of carbon flow towards β-oxidation and polyhydroxyalkanoate biosynthesis in Yarrowia lipolytica. FEMS Yeast Res. 2010, 10, 917–927. [Google Scholar] [CrossRef] [Green Version]
- Strijbis, K.; Vaz, F.M.; Distel, B. Enzymology of the carnitine biosynthesis pathway. Life 2010, 62, 357–362. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Daviet, L.; Schalk, M.; Siewers, V.; Nielsen, J. Establishing a platform cell factory through engineering of yeast acetyl-CoA metabolism. Metab. Eng. 2013, 15, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Dulermo, R.; Gamboa-Melendez, H.; Ledesma-Amaro, R.; Thevenieau, F.; Nicaud, J.-M. Unraveling fatty acid transport and activation mechanisms in Yarrowia lipolytica. Biochim. Biophys. Acta 2015, 1851, 1202–1217. [Google Scholar] [CrossRef] [PubMed]
- Mauersberger, S. Cytochromes P450 of the alkane-utilising yeast. In Yarrowia lipolytica. Microbiology Monographs, 1st ed.; Barth, G., Ed.; Springer: Berlin, Heidelberg, Germany, 2013; Volume 25, pp. 227–262. [Google Scholar] [CrossRef]
- Swiegers, J.H.; Dippenaar, N.; Pretorius, I.S.; Bauer, F.F. Carnitine-dependent metabolic activities in Saccharomyces cerevisiae: Three carnitine acetyltransferases are essential in a carnitine-dependent strain. Yeast 2001, 18, 585–595. [Google Scholar] [CrossRef]
- Rousta, N.; Ferreira, J.A.; Taherzadeh, M.J. Production of L-carnitine-enriched edible filamentous fungal biomass through submerged cultivation. Bioengineered 2021, 12, 358–368. [Google Scholar] [CrossRef]
- Dominguez, A.; Deive, F.J.; Angeles Sanroman, M.; Longo, M.A. Biodegradation and utilization of waste cooking oil by Yarrowia lipolytica CECT 1240. Eur. J. Lipid Sci. Technol. 2010, 112, 1200–1208. [Google Scholar] [CrossRef] [Green Version]
- Carsanba, E.; Papanikolaou, S.; Erten, H. Production of oils and fats by oleaginous microorganisms with an emphasis given to the potential of the nonconventional yeast Yarrowia lipolytica. Crit. Rev. Biotechnol. 2018, 38, 1230–1243. [Google Scholar] [CrossRef]
- Kerkhoven, E.J.; Pomraning, K.R.; Baker, S.E.; Nielsen, J. Regulation of amino-acid metabolism controls flux to lipid accumulation in Yarrowia lipolytica. NPJ Syst. Biol. Appl. 2016, 2, 16005. [Google Scholar] [CrossRef] [Green Version]
- Parfene, G.; Hornicar, V.B.; Bahrim, G.; Vannini, L.; Gottarrdi, D.; Guerzoni, M.E. Lipolytic activity of lipases from different strains of Yarrowia lipolytica in hydrolysed vegetable fats at low temperature and water activity. Rom. Biotechnol. Lett. 2011, 16, 46–52. [Google Scholar]
- Wang, G.; Liu, Z.; Xu, Z.; Yan, Y. Aromatic amino acid mutagenesis at the substrate binding pocket of Yarrowia lipolytica lipase Lip2 affects its activity and thermostability. Sci. World J. 2014, 2014, 382581. [Google Scholar] [CrossRef] [Green Version]
- EFSA. Scientific Opinion on Dietary Reference Values for selenium. EFSA J. 2014, 12, 3846. [Google Scholar] [CrossRef]
- Izah, S.C.; Enaregha, E.B.; Epidi, J.O. Vitamin content of Saccharomyces cerevisiae biomass cultured in cassava wastewater. MOJ Toxicol. 2019, 4, 42–45. [Google Scholar]
- Paalme, T.; Kevvai, K.; Vilbaste, A.; Hälvin, K.; Nisamedtinov, I. Uptake and accumulation of B-group vitamers in Saccharomyces cerevisiae in ethanol-stat fed-batch culture. World J. Microbiol. Biotechnol. 2014, 30, 2351–2359. [Google Scholar] [CrossRef]
- Kennedy, D.O. B vitamins and the brain: Mechanisms, dose and efficacy—A review. Nutrients 2016, 8, 68. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Devi, A.; Rush, E.; Harper, M.; Venn, B. Vitamin B12 status of various ethnic group living in New Zeland: An analysis of the adult nutrition survey 2008/2009. Nutrients 2018, 10, 181. [Google Scholar] [CrossRef] [Green Version]
- O’Leary, F.; Samman, S. Vitamin B12 in Health and Disease. Nutrients 2010, 2, 299–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Green, R. Vitamin B12 deficiency from the perspective of a practicing hematologist. Blood 2017, 129, 2603–2611. [Google Scholar] [CrossRef] [Green Version]
- Naik, S.; Mahalle, N.; Bhide, V. Identification of vitamin B12 deficiency in vegetarian Indians. Br. J. Nutr. 2018, 15, 629–635. [Google Scholar] [CrossRef] [Green Version]
- Dobrowolski, A.; Drzymala, K.; Rzechonek, D.A.; Mitula, P.; Mironczuk, A.M. Lipid production from waste materials in seawater-based medium by the yeast Yarrowia lipolytica. Front. Microbiol. 2019, 10, 547. [Google Scholar] [CrossRef] [Green Version]
- Braga, A.; Belo, I. Biotechnological production of γ-decalactone, a peach like aroma, by Yarrowia lipolytica. World J. Microbiol. Biotechnol. 2016, 32, 169. [Google Scholar] [CrossRef] [Green Version]
- Arslan, N.P.; Aydogan, M.N.; Taskin, M. Citric acid production from partly deproteinized whey under non-sterile culture conditions using immobilized cells of lactose-positive and cold-adapted Yarrowia lipolytica B9. J. Biotechnol. 2016, 231, 32–39. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, E.; Charreau, H.; Cerrutti, P.; Foresti, M.L. Yarrowia lipolytica: A model yeast for citric acid production. FEMS Yeast Res. 2017, 17, fox084. [Google Scholar] [CrossRef] [PubMed]
- Cavallo, E.; Nobile, M.; Cerrutti, P.; Foresti, M.L. Exploring the production of citric acid with Yarrowia lipolytica using corn wet milling products as alternative low-cost fermentation media. Biochem. Eng. J. 2020, 155, 107463. [Google Scholar] [CrossRef]
- Dobrowolski, A.; Mikuła, P.; Rymowicz, W.; Mirończuk, A.M. Efficient conversion of crude glycerol from various industrial wastes into single cell oil by yeast Yarrowia lipolytica. Bioresour. Technol. 2016, 207, 237–243. [Google Scholar] [CrossRef]
- Ferreira, I.M.P.L.V.O.; Pinho, O.; Vieira, E.; Tavarela, J.G. Brewer’s Saccharomyces yeast biomass: Characteristics and potential applications. Trends Food Sci. Technol. 2010, 21, 77–84. [Google Scholar] [CrossRef]
- Gonçalves, F.A.G.; Colen, G.; Takahashi, J.A. Yarrowia lipolytica and its multiple applications in the biotechnological industry. Sci. World J. 2014, 2014, 1–14. [Google Scholar] [CrossRef] [Green Version]
- Kumar, L.R.; Sravan, K.; Yellapu, K.S.; Yan, S.; Tyagi, R.D.; Drogui, P. Elucidating the effect of impurities present in different crude glycerol sources on lipid and citric acid production by Yarrowia lipolytica SKY7. J. Chem. Technol. Biotechnol. 2020, 96, 227–240. [Google Scholar] [CrossRef]
- Levinson, W.E.; Kurtzman, C.P.; Kuo, T.M. Characterization of Yarrowia lipolytica and related species for citric acid production from glycerol. Enzym. Microb. Technol. 2007, 41, 292–295. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Chevalot, I.; Komaitis, M.; Marc, I.; Aggelis, G. Single cell oil production by Yarrowia lipolytica growing on an industrial derivative of animal fat in batch cultures. Appl. Microbiol. Biotechnol. 2002, 58, 308–312. [Google Scholar] [CrossRef]
- Makri, A.; Fakas, S.; Aggelis, G. Metabolic activities of biotechnological interest in Yarrowia lipolytica grown on glycerol in repeated batch cultures. Bioresour. Technol. 2010, 101, 2351–2358. [Google Scholar] [CrossRef]
- Rakicka, M.; Wolniak, J.; Lazar, Z.; Rymowicz, W. Production of high titer of citric acid from inulin. BMC Biotechnol. 2019, 19, 11. [Google Scholar] [CrossRef] [PubMed]
- Sabra, W.; Bommareddy, R.R.; Maheshwari, G.; Papanikolaou, S.; Zeng, A.-P. Substrates and oxygen dependent citric acid production by Yarrowia lipolytica: Insights through transcriptome and fluxome analyses. Microb. Cell Factories 2017, 16, 78. [Google Scholar] [CrossRef] [PubMed]
- Cybulski, K.; Tomaszewska-Hetman, L.; Rakicka, M.; Juszczyk, P.; Rywinska, A. Production of pyruvic acid from glycerol by Yarrowia lipolytica. Folia Microbiol. 2019, 64, 809–820. [Google Scholar] [CrossRef] [PubMed]
- Lei, Q.; Zeng, W.; Zhou, J.; Du, G. Efficient separation of α-ketoglutarate from Yarrowia lipolytica WSH-Z06 culture broth by converting pyruvate to l-tyrosine. Bioresour. Technol. 2019, 292, 121897. [Google Scholar] [CrossRef] [PubMed]
- Morgunov, I.G.; Kamzolova, S.V.; Lunina, J.N. The citric acid production from raw glycerol by Yarrowia lipolytica yeast and its regulation. Appl. Microbiol. Biotechnol. 2013, 97, 7387–7397. [Google Scholar] [CrossRef]
- Rywińska, A.; Tomaszewska-Hetman, L.; Rakicka-Pustułka, M.; Juszczyk, P.; Rymowicz, W. Alpha-Ketoglutaric acid production from a mixture of glycerol and rapeseed oil by Yarrowia lipolytica using different substrate feeding strategies. Sustainability 2020, 12, 6109. [Google Scholar] [CrossRef]
- Wróbel-Kwiatkowska, M.; Turski, W.; Juszczyk, P.; Kita, A.; Rymowicz, W. Improved production of kynurenic acid by Yarrowia lipolytica in media containing different honeys. Sustainability 2020, 12, 9424. [Google Scholar] [CrossRef]
- Fickers, P.; Cheng, H.; Lin, C.S.K. Sugar alcohols and organic acids synthesis in Yarrowia lipolytica: Where are we? Microorganisms 2020, 8, 574. [Google Scholar] [CrossRef]
- Fu, G.Y.; Lu, Y.; Chi, Z.; Liu, G.L.; Zhao, S.F.; Jiang, H.; Chi, Z.M. Cloning and characterization of a pyruvate carboxylase gene from penicillium rubens and overexpression of the genein the Yeast Yarrowia lipolytica for enhanced citric acid production. Mar. Biotechnol. 2016, 18, 1–14. [Google Scholar] [CrossRef]
- Kamzolova, S.V.; Morgunov, I.G. Optimization of medium composition and fermentation conditions for α-ketoglutaric acid production from biodiesel waste by Yarrowia lipolytica. Appl. Microbiol. Biotechnol. 2020, 104, 7979–7989. [Google Scholar] [CrossRef]
- Rzechonek, D.A.; Dobrowolski, A.; Rymowicz, W.; Mirończuk, A.M. Aseptic production of citric and isocitric acid from crude glycerol by genetically modified Yarrowia lipolytica. Bioresour. Technol. 2019, 271, 340–344. [Google Scholar] [CrossRef] [PubMed]
- Yuzbasheva, E.Y.; Scarcia, P.; Yuzbashev, T.V.; Messina, E.; Kosikhina, I.M.; Palmieri, L.; Shutov, A.V.; Taratynova, M.O.; Amaro, R.L.; Palmieri, F.; et al. Engineering Yarrowia lipolytica for the selective and high-level production of isocitric acid through manipulation of mitochondrial dicarboxylate-tricarboxylate carriers. Metab. Eng. 2021, 65, 156–166. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Bowman, R.S. Retention of inorganic oxyanions by organo-kaolinite. Water Res. 2001, 35, 3771–3776. [Google Scholar] [CrossRef]
- Otto, C.; Yovkova, V.; Barth, G. Overproduction and secretion of α-ketoglutaric acid by microorganisms. Appl. Microbiol. Biotechnol. 2011, 92, 689–695. [Google Scholar] [CrossRef]
- Grembecka, M. Natural sweeteners in a human diet. Rocz. Państwowego Zakładu Hig. 2015, 66, 195–202. [Google Scholar]
- Mirończuk, A.M.; Biegalska, A.; Zugaj, K.; Rzechonek, D.A.; Dobrowolski, A. A role of a newly identified isomerase from Yarrowia lipolytica in erythritol catabolism. Front. Microbiol. 2018, 9, 1122. [Google Scholar] [CrossRef]
- Rakicka, M.; Biegalska, A.; Rymowicz, W.; Dobrowolski, A.; Mironczuk, A.M. Polyol production from waste materials by genetically modified Yarrowia lipolytica. Bioresour. Technol. 2017, 243, 393–399. [Google Scholar] [CrossRef]
- Rzechonek, D.A.; Dobrowolski, A.; Rymowicz, W.; Mirończuk, A.M. Recent advances in biological production of erythritol. Crit. Rev. Biotechnol. 2018, 38, 620–633. [Google Scholar] [CrossRef]
- Papanikolaou, S.; Kampisopoulou, E.; Blanchard, F.; Rondags, E.; Gardeli, C.; Koutinas, A.A.; Chevalot, I.; Aggelis, G. Production of secondary metabolites through glycerol fermentation under carbon-excess conditions by the yeasts Yarrowia lipolytica and Rhodosporidium toruloides. Eur. J. Lipid Sci. Technol. 2017, 119, 1600507. [Google Scholar] [CrossRef]
- Egermeier, M.; Russmayer, H.; Sauer, M.; Marx, H. Metabolic flexibility of Yarrowia lipolytica growing on glycerol. Front. Microbiol. 2017, 8, 49. [Google Scholar] [CrossRef] [Green Version]
- Mirończuk, A.M.; Rakicka, M.; Biegalska, A.; Rymowicz, W.; Dobrowolski, A. A two-stage fermentation process of erythritol production by yeast Y. lipolytica from molasses and glycerol. Bioresour. Technol. 2015, 198, 445–455. [Google Scholar] [CrossRef] [PubMed]
- Rywinska, A.; Bak, M.; Rakicka, M.; Tomaszewska, L.; Boruczkowski, T.; Lazar, Z.; Musial, I.; Rymowicz, W. Selection of the UV mutants of Yarrowia lipolytica yeast for erythritol biosynthesis from glycerol. Acta Sci. Pol. 2012, 11, 23–37. [Google Scholar]
- Tomaszewska, L.; Rywińska, A.; Gładkowski, W. Production of erythritol and mannitol by Yarrowia lipolytica yeast in media containing glycerol. J. Ind. Microbiol. Biotechnol. 2012, 39, 1333–1343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szczepańczyk, M.; Rzechonek, D.A.; Dobrowolski, A.; Mirończuk, A.M. The overexpression of YALI0B07117g results in enhanced erythritol synthesis from glycerol by the yeast Yarrowia lipolytica. Molecules 2021, 26, 7549. [Google Scholar] [CrossRef]
- Mirończuk, A.M.; Furgała, J.; Rakicka, M.; Rymowicz, W. Enhanced production of erythritol by Yarrowia lipolytica on glycerol in repeated batch cultures. J. Ind. Microbiol. Biotechnol. 2014, 41, 57–64. [Google Scholar] [CrossRef] [Green Version]
- Andre, A.; Chatzifragkou, A.; Diamantopoulou, P.; Sarris, D.; Philippoussis, A.; Galiotou-Panayotou, M.; Komaitis, M.; Papanikolaou, S. Biotechnological conversions of bio-diesel-derived crude glycerol by Yarrowia lipolytica strains. Eng. Life Sci. 2009, 9, 468–478. [Google Scholar] [CrossRef]
- Larroude, M.; Trabelsi, H.; Nicaud, J.M.; Rossignol, T. A set of Yarrowia lipolytica CRISPR/Cas9 vectors for exploiting wild-type strain diversity. Biotechnol. Lett. 2020, 42, 773–785. [Google Scholar] [CrossRef] [Green Version]
- Celińska, E.; Nicaud, J.M.; Białas, W. Hydrolytic secretome engineering in Yarrowia lipolytica for consolidated bioprocessing on polysaccharide resources: Review on starch, cellulose, xylan, and inulin. Appl. Microbiol. Biotechnol. 2021, 105, 975–989. [Google Scholar] [CrossRef]
- Bruder, S.; Melcher, F.A.; Zoll, T.; Hackenschmidt, S.; Kabisch, J. Evaluation of a Yarrowia lipolytica strain collection for its lipid and carotenoid production capabilities. Eur. J. Lipid Sci. Technol. 2020, 122, 1900172. [Google Scholar] [CrossRef]
- Worland, A.M.; Czajka, J.J.; Xing, Y.; Harper, W.F.; Moore, A.; Xiao, Z.; Han, Z.; Wang, Y.; Su, W.W.; Tang, Y.J. Analysis of Yarrowia lipolytica growth, catabolism, and terpenoid biosynthesis during utilization of lipid-derived feedstock. Metab. Eng. Commun. 2020, 11, e00130. [Google Scholar] [CrossRef]
- Pang, Y.; Zhao, Y.; Li, S.; Zhao, Y.; Li, J.; Hu, Z.; Zhang, C.; Xiao, D.; Yu, A. Engineering the oleaginous yeast Yarrowia lipolytica to produce limonene from waste cooking oil. Biotechnol. Biofuels 2019, 12, 1–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, K.K.; Alper, H.S. Yarrowia lipolytica: More than an oleaginous workhorse. Appl. Microbiol. Biotechnol. 2019, 103, 9251–9262. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.M.; Miller, K.K.; Nguyen, A.; Alper, H.S. Engineering 4-coumaroyl- CoA derived polyketide production in Yarrowia lipolytica through a β-oxidation mediated strategy. Metab. Eng. 2020, 57, 174–181. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Han, B.; Gui, X.; Wang, G.; Xu, L.; Yan, Y.; Madzak, C.; Pan, D.; Wang, Y.; Zha, G.; et al. Engineering Yarrowia lipolytica to simultaneously produce lipase and single cell protein from agro-industrial wastes for feed. Sci. Rep. 2018, 8, 758. [Google Scholar] [CrossRef] [Green Version]
- Madzak, C.; Tréton, B.; Blanchin-Roland, S. Strong hybrid promoters and integrative expression/secretion vectors for quasi-constitutive expression of heterologous proteins in the yeast Yarrowia lipolytica. J. Mol. Microbiol. Biotechnol. 2000, 2, 207–216. [Google Scholar] [PubMed]
- Vandermies, M.; Fickers, P. Bioreactor-scale strategies for the production of recombinant protein in the yeast Yarrowia lipolytica. Microorganisms 2019, 7, 40. [Google Scholar] [CrossRef] [Green Version]
- Madzak, C. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol. 2015, 99, 4559–4577. [Google Scholar] [CrossRef]
- Madhavan, A.; Arun, K.B.; Sindhu, R.; Krishnamoorthy, J.; Reshmy, R.; Sirohi, R.; Pugazhendi, A.; Awasthi, M.K.; Szakacs, G.; Binod, P. Customized yeast cell factories for biopharmaceuticals: From cell engineering to process scale up. Microb. Cell Factories 2021, 20, 124. [Google Scholar] [CrossRef]
- Jiao, X.; Gu, Y.; Zhou, P.; Yu, H.; Ye, L. Recent advances in construction and regulation of yeast cell factories. World J. Microbiol. Biotechnol. 2022, 38, 57. [Google Scholar] [CrossRef]
- Madzak, C.; Beckerich, J.M. Heterologous protein expression and secretion in Yarrowia lipolytica. In Yarrowia lipolytica: Biotechnological Applications, Microbiology Monographs; Barth, G., Ed.; Springer: Heidelberg, Germany, 2013; Volume 25, pp. 1–76. [Google Scholar]
- Gasmi, N.; Ayed, A.; Ammar, B.B.; Zrigui, R.; Nicaud, J.-M.; Kallel, H. Development of a cultivation process for the enhancement of human interferon alpha 2b production in the oleaginous yeast, Yarrowia lipolytica. Microb. Cell Factories 2011, 10, 90–100. [Google Scholar] [CrossRef] [Green Version]
- Hamsa, P.V.; Kachroo, P.; Chattoo, B.B. Production and secretion of biologically active human epidermal growth factor in Yarrowia lipolytica. Curr. Genet. 1998, 33, 231–237. [Google Scholar] [CrossRef] [PubMed]
- Kang, H.A.; Kim, J.Y.; Oh, D.B.; Park, J.N.; Song, Y. A Novel Ylmpo1 Gene Derived from Yarrowia lipolytica and a Process for Preparing a Glycoprotein Not Being Mannosylphosphorylated by Using a Mutated Yarrowia lipolytica in Which Ylmpo1 Gene Is Disrupted. WO Patent WO2008136564, 13 November 2008. [Google Scholar]
- Callewaert, N.L.M.; Vervecken, W.; Marcel de Pourcq, K.J.; Geysens, S.C.J.; Guerfal, M. Glycosylation of Molecules. WO Patent WO2008120107, 9 October 2008. [Google Scholar]
- Tharaud, C.; Ribet, A.M.; Costes, C.; Gaillardin, C. Secretion of human blood coagulation factor XIIIa by the yeast Yarrowia lipolytica. Gene 1992, 121, 111–119. [Google Scholar] [CrossRef]
- James, L.; Strick, C. Multiple Integrative Vectors and Yarrowia lipolytica Transformants. U.S. Patent US5786212A, 28 July 1998. [Google Scholar]
- Nthangeni, M.B.; Urban, P.; Pompon, D.; Smit, M.S.; Nicaud, J.-M. The use of Yarrowia lipolytica for the expression of human cytochrome P450 CYP1A1. Yeast 2004, 21, 583–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cho, E.M.; Lee, H.S.; Eom, C.Y.; Ohta, A. Construction of high sensitive detection system for endocrine disruptors with yeast n-alkane-assimilating Yarrowia lipolytica. J. Microbiol. Biotechnol. 2010, 20, 1563–1570. [Google Scholar] [CrossRef]
- Davidow, L.S.; Franke, A.E.; De Zeeuw, J.R. Expression and Secretion of Heterologous Proteins by Yarrowia lipolytica Transformants. EP Patent EP0220864B1, 3 March 1993. [Google Scholar]
- Simon-Nobbe, B.; Denk, U.; Pöll, V.; Rid, R.; Breitenbach, M. The spectrum of fungal allergy. Int. Arch. Allergy Immunol. 2008, 145, 58–86. [Google Scholar] [CrossRef] [Green Version]
- Allred, L.K.; Nye-Wood, M.G.; Colgrave, M.L. Analysis of gluten in dried yeast and yeast-containing products. Foods 2020, 9, 1790. [Google Scholar] [CrossRef]
Figure 1.
Composition of Yarrowia lipolytica biomass.

Figure 2.
The dried powder of Y. lipolytica.


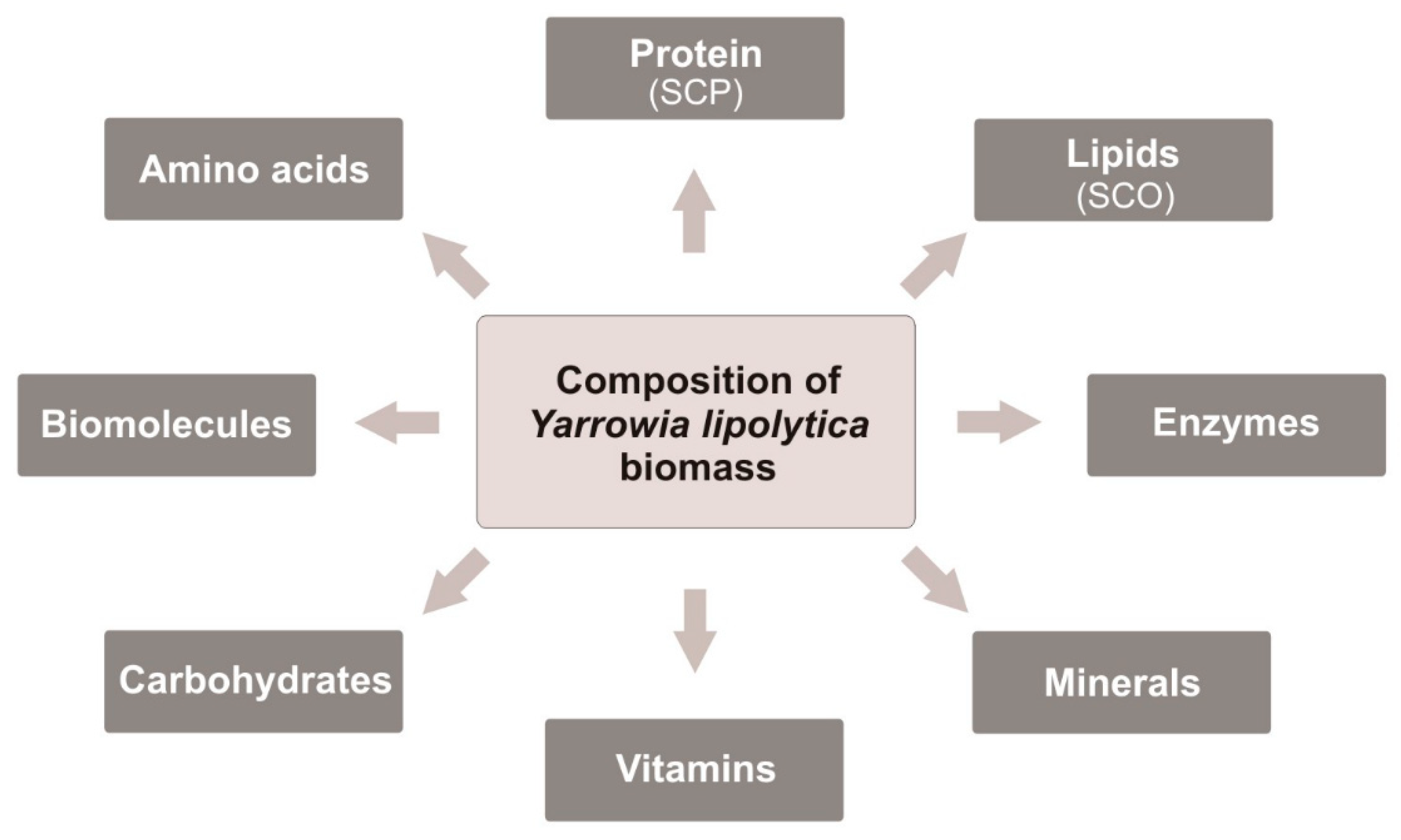{kind=link}
{kind=link}
Table 1.
Reports of Yarrowia lipolytica strains cultured in specific wastes, culture conditions, and protein content.
Table 1.
Reports of Yarrowia lipolytica strains cultured in specific wastes, culture conditions, and protein content.
| Yarrowia lipolytica Strains | Waste Substrates | Culture Conditions | Protein Content | Reference |
|---|---|---|---|---|
| wt A-101 | Rye straw | Shake-flask, 28 °C, pH 3.5, with shaking at 150 rpm, 120 h | 30.5% | [67] |
| Biofuel waste | Bioreactor 100 L, 30 °C, pH 5.0, 40% oxygen agitation, 12 h | 40–50% | [6,68] | |
| Rye brans | Shake-flask, 28 °C, pH 3.5, with shaking at 150 rpm, 120 h | 44.5% | [67] | |
| Oat brans | Shake-flask, 28 °C, pH 3.5, at shaking 150 rpm, 120 h | 44.5% | [67] | |
| Industrial raw oil waste with glycerol (88%) | Nd | 45% | [69] | |
| Waste glycerol (78%) with degumming from rapeseed oil | Biostat B-Plus 5 L with mechanical agitation and bubble bioreactor 2000 L, 30 °C, pH 3.5–3.8 | 46.4–50.3% | [70] | |
| Raw rapeseed oil | AK 210 bioreactor, 30 °C, pH 3.5, with shaking at 750 rpm, 1 vvn oxygenation, 12 h | 56.4% | [71] | |
| Nd | Industrial glycerol | AK 210 bioreactor, 28 °C, pH 5.5, 1.2 L/ min of aeration, 48 h | 46.7% | [72] |
| wt SWJ-1b | Soy bean cake hydrolysate with 20 g/L of glucose | 28 °C, pH 5.5 | 41–47.6% | [73] |
| wt S6 | pure and raw glycerol (25 g/L) | Bioreactor, 30 °C, pH 3.5 | 45% | [74] |
| wt S11 | pure and raw glycerol (25 g/L) | Bioreactor, 30 °C, pH 3.5 | 50.1% | [74] |
| wt ACA-DC 50109 | Industrial derivative of tallow (stearin 100%) | Bioreactor, 28 °C, pH 6.0, 650 rpm agitation, 0.3 vvm of aeration, 24 h | Nd | [75] |
| Recombinant 29a | Hydrolysate of soy bean meal with ammonium sulfate | Shake-flask 250 mL, 28 °C, at shaking 170 rpm, 120 h | 46.1% | [76] |
| Recombinant | Inulin | Bioreactor, 72 h | 47.5% | [77] |
| meal of Jerusalem artichoke tuber (8.0%) | 2–1 bioreactor, 80 h | 53.7 |
Nd—not disclosed; wt—wild type.
Table 2.
Nutritional analysis of Yarrowia lipolytica protein powder (dry weight).
| Parameter | Amount | Requirements for Adults [110] |
|---|---|---|
| Dietary fiber (g/100 g) | 20.0–30.7 | - |
| Ether extract (g/100 g) | 57.3 | - |
| Nitrogen-free extract (%) | 96.2 | - |
| Saccharides (g/100 g) | 0–2.0 | 90 g |
| Total ash (%) | 9.0–15.0 | - |
| Dry matter content (%) | 86.2–98.5 | - |
Table 3.
Amino acids concentration in protein of Yarrowia lipolytica powder (dry weight).
| Parameter | Amount | FAO Requirements for Adults |
|---|---|---|
| Protein (g/100 g DW) | 20–56.4 | 50 |
| Amino acid | mg/1 g protein | |
| Alanine | 17.8–80.0 | - |
| Arginine | 12.7–48.0 | - |
| Asparagine | 31.8–88.0 | - |
| Cysteine | 3.5–11.0 | - |
| Glycine | 14.4–46.0 | - |
| Glutamine | 40.2–120.0 | - |
| Histidine * | 8.5–26.0 | 15 |
| Isoleucine * | 13.9–44.0 | 30 |
| Leucine * | 33.8–71.2 | 59 |
| Lysine * | 18.3–70.0 | 45 |
| Methionine * | 4.7–14.4 | - |
| ΣSAA | 8.2–25.4 | 22 |
| Phenylalanine * | 13.8–69.0 | - |
| Proline | 13.0–42.0 | - |
| Serine | 15.6–44.7 | - |
| Tryptophan * | 3.5–47.0 | - |
| Tyrosine | 8.1–110.0 | - |
| ΣAAA | 25.4–197.0 | 38 |
| Threonine * | 18.9–50.0 | 23 |
| Valine * | 17.7–53.3 | 39 |
| ΣEAA | 133.1–413.9 | - |
DW—dry weight; *—essential amino acid; ΣAAA—sum of aromatic amino acids: phenylalanine, tyrosine and tryptophan; ΣEAA—sum of essential amino acids; FAO—Food and Agricultural Organization; ΣSAA—sum of sulfur amino acids: cysteine and methionine.
Table 4.
Trace mineral content in Yarrowia lipolytica protein powder (dry weight).
| Parameter | Amount | Requirements for Adults [110] |
|---|---|---|
| Calcium (mg/100 g) | 154–426 | 800 mg |
| Cupper (mg/100 g) | 0.65–0.68 | 1 mg |
| Chromium (mg/100 g) | 0.94–9.19 | 40 μg |
| Iodine (mg/100 g) | 41 | 150 μg |
| Iron (mg/100 g) | 10.99–12.42 | 14 mg |
| Magnesium (mg/100 g) | 182–204 | 375 mg |
| Manganese (mg/100 g) | 0.25–1.82 | 2 mg |
| Molybdenum (μg/100 g) | 38 | 50 μg |
| Phosphorus (mg/100 g) | 340–528 | 700 mg |
| Potassium (g/100 g) | 1.34–2.51 | 2 g |
| Selenium (mg/100 g) | 2.48–37.7 | 55 μg |
| Sulfur (g/100 g) | 0.35–0.62 | - |
| Zinc (mg/100 g) | 5.79–8.25 | 10 mg |
| Sodium content (g/100 g) | 1.03–1.98 | 6 g |
Table 5.
Vitamin concentrations in Yarrowia lipolytica protein powder (dry weight).
| Vitamin | Amount | Requirements for Adults [110] |
|---|---|---|
| Vitamin B1 (mg/100 g) | 0.68–11.5 | 1.1 mg |
| Vitamin B2 (mg/100 g) | 1.1–6.9 | 1.4 mg |
| Vitamin B6 (mg/100 g) | 2.53–6.50 | 1.4 mg |
| Vitamin B7 (μg/100 g) | 9.32–62.0 | 50 μg |
| Vitamin B9 (μg/100 g) | 184–330 | 200 μg |
| Vitamin B12 (μg/100 g) | 5.2–11.2 | 2.5 μg |
| Vitamin E (mg/100 g) | 0.60–0.75 | 12 mg |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jach, M.E.; Malm, A. Yarrowia lipolytica as an Alternative and Valuable Source of Nutritional and Bioactive Compounds for Humans. Molecules 2022, 27, 2300. https://doi.org/10.3390/molecules27072300
AMA Style
Jach ME, Malm A. Yarrowia lipolytica as an Alternative and Valuable Source of Nutritional and Bioactive Compounds for Humans. Molecules. 2022; 27(7):2300. https://doi.org/10.3390/molecules27072300
Chicago/Turabian StyleJach, Monika Elżbieta, and Anna Malm. 2022. "Yarrowia lipolytica as an Alternative and Valuable Source of Nutritional and Bioactive Compounds for Humans" Molecules 27, no. 7: 2300. https://doi.org/10.3390/molecules27072300