
The Involvement of l-Arginine-Nitric Oxide-cGMP-ATP-Sensitive K+ Channel Pathway in Antinociception of BBHC, a Novel Diarylpentanoid Analogue, in Mice Model

 , , , and
, , , and 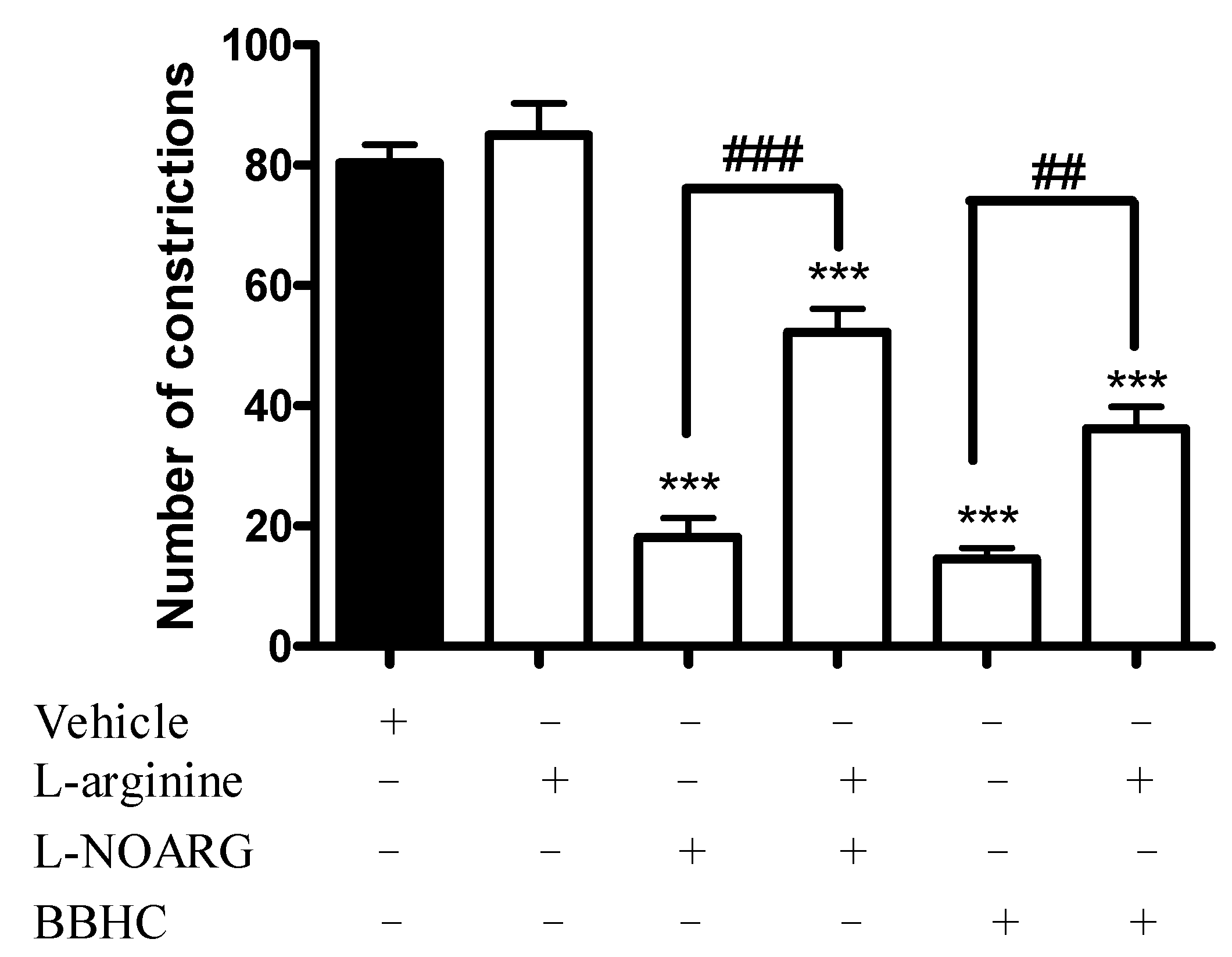{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Involvement of l-Arginine-Nitric Oxide
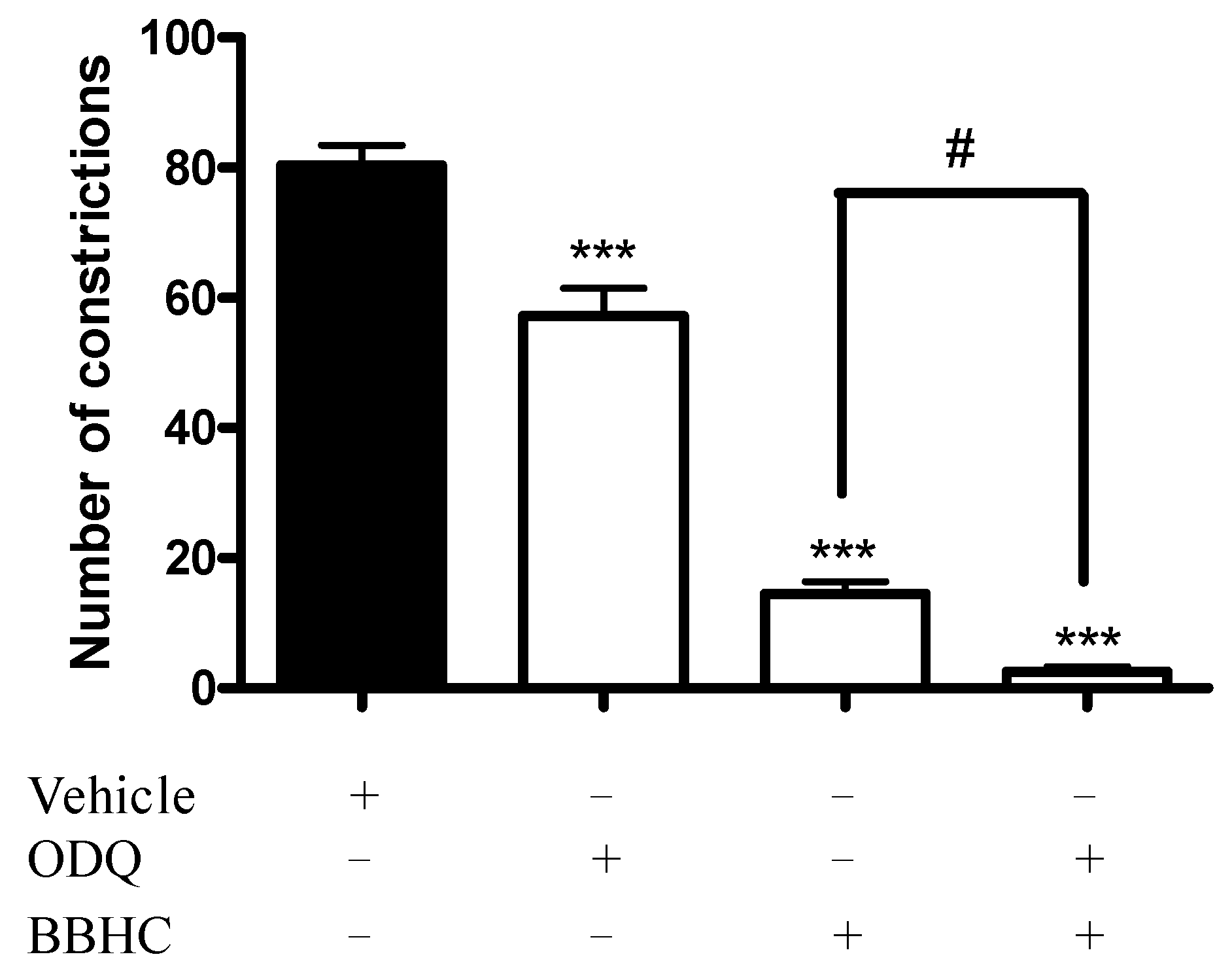
2.2. Involvement of Cyclic Guanosine Monophosphate
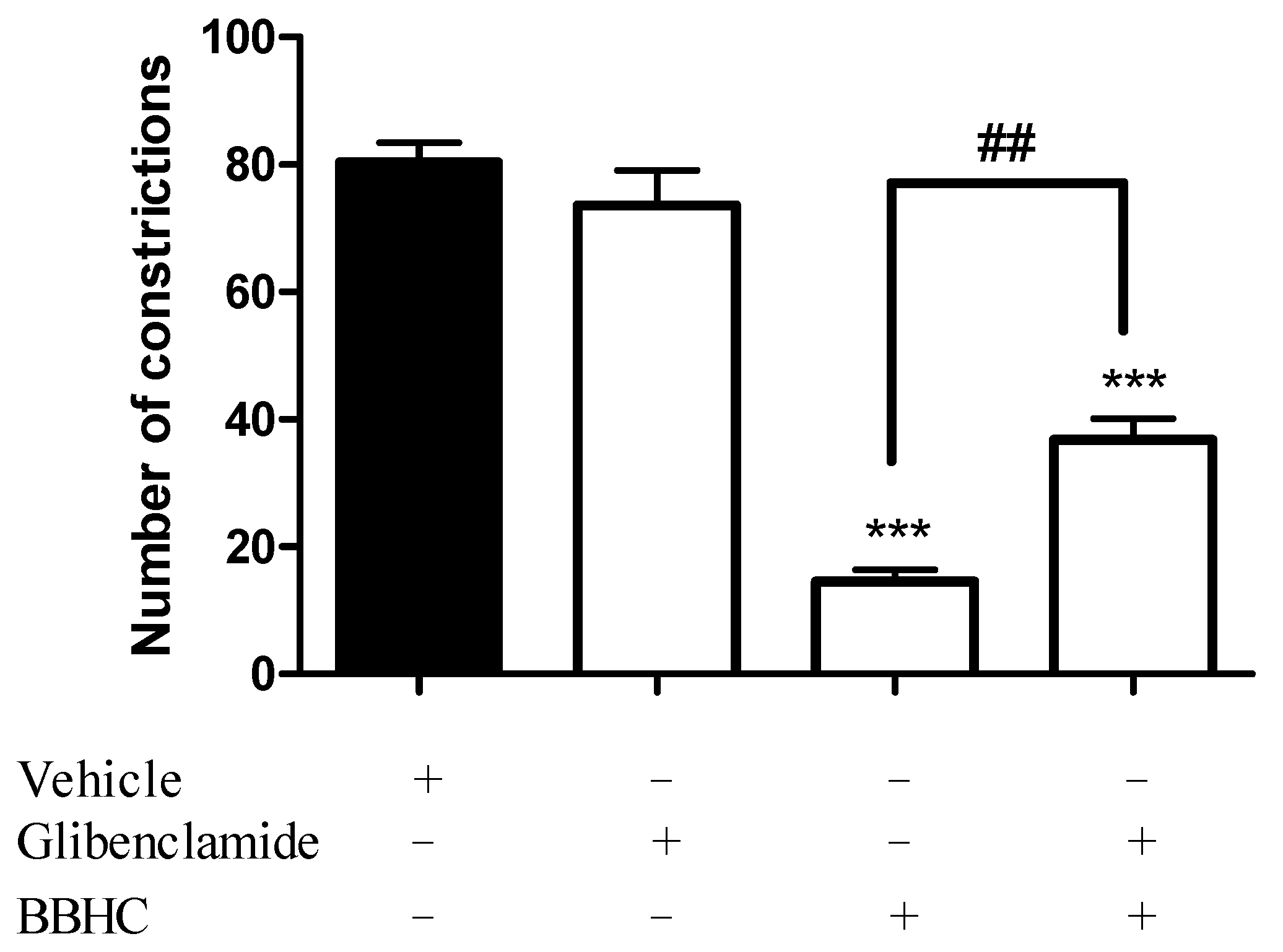
2.3. Involvement of ATP-Sensitive K+ Channel
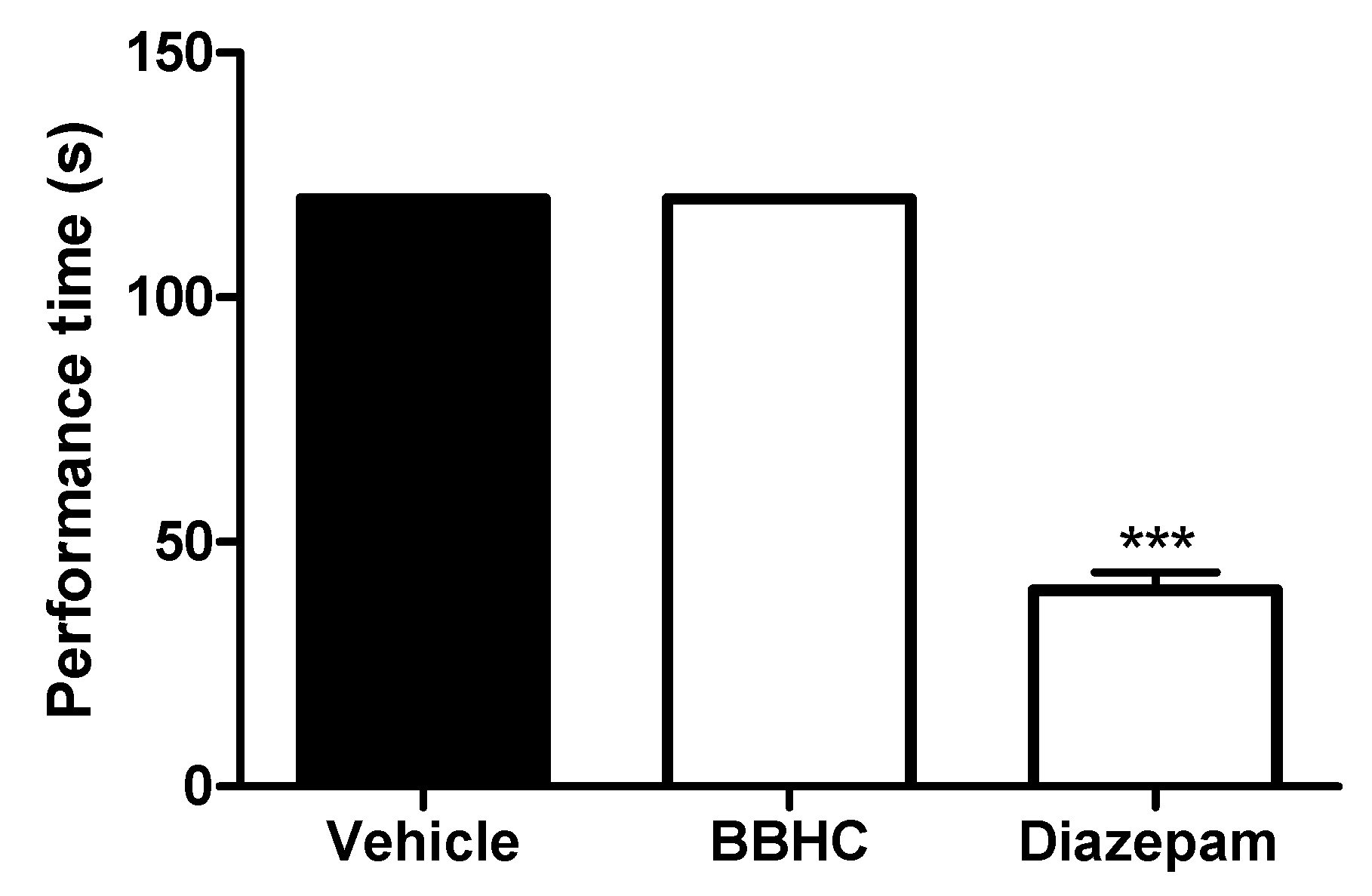
2.4. Rota-Rod Test
3. Discussion
4. Materials and Methods
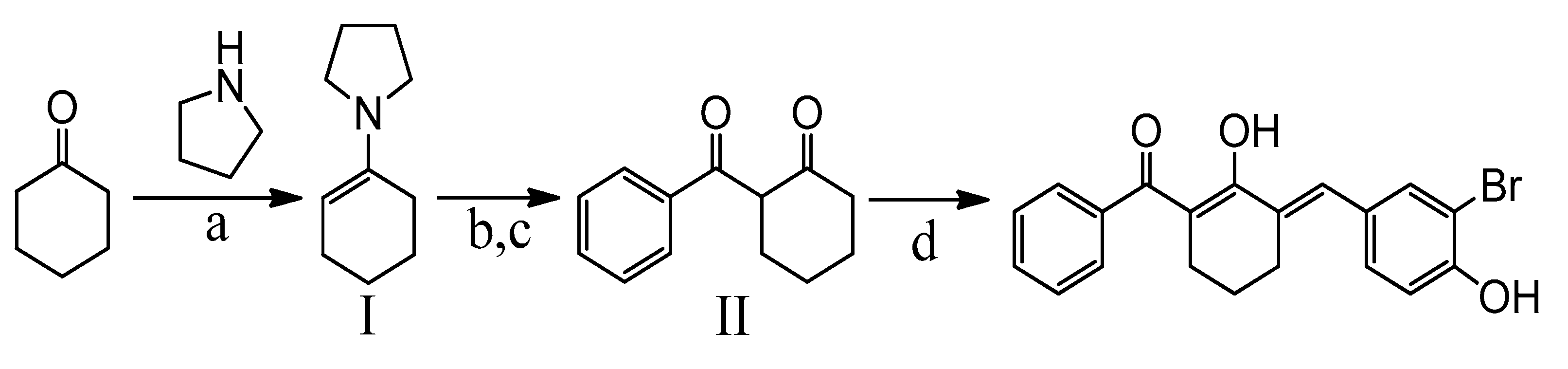
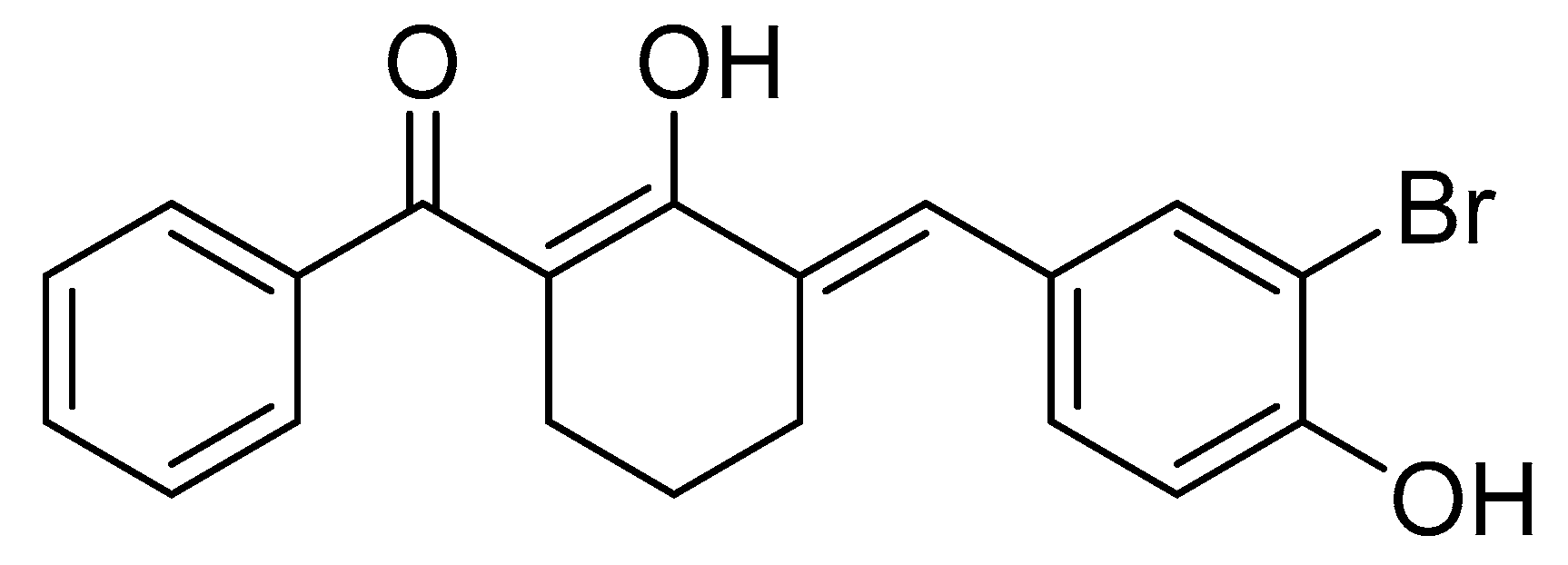
4.1. Synthesis of BBHC
4.1.1. Synthetic Scheme of BBHC
4.1.2. General Procedure for Synthesis of I and II
4.1.3. General Procedure for the Synthesis BBHC
4.2. Preparation of BBHC
4.3. Preparation of Drugs and Chemicals
4.4. Experimental Animals
4.5. Involvement of l-Arginine-Nitric Oxide
4.6. Involvement of Cyclic Guanosine Monophosphate
4.7. Involvement of an ATP-Sensitive K+ Channel
4.8. Rota-Rod Test
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Luo, Z.D.; Cizkova, D. The role of nitric oxide in nociception. Curr. Rev. Pain 2000, 4, 459–466. [Google Scholar] [CrossRef]
- Cury, Y.; Picolo, G.; Gutierrez, V.P.; Ferreira, S.H. Pain and analgesia: The dual effect of nitric oxide in the nociceptive system. Nitric Oxide 2011, 25, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Esplugues, J.V. NO as a signalling molecule in the nervous system. Br. J. Pharmacol. 2002, 135, 1079–1095. [Google Scholar] [CrossRef] [Green Version]
- Paradise, W.A.; Vesper, B.J.; Goel, A.; Waltonen, J.D.; Altman, K.W.; Haines, G.K.; Radosevich, J.A. Nitric oxide: Perspectives and emerging studies of a well known cytotoxin. Int. J. Mol. Sci. 2010, 11, 2715–2745. [Google Scholar] [CrossRef]
- Olson, N.; van der Vliet, A. Interactions between nitric oxide and hypoxia-inducible factor signaling pathways in inflammatory disease. Nitric Oxide 2011, 25, 125–137. [Google Scholar] [CrossRef] [Green Version]
- Miclescu, A.; Gordh, T. Nitric oxide and pain: ‘Something old, something new’. Acta Anaesthesiol. Scand. 2009, 53, 1107–1120. [Google Scholar] [CrossRef]
- Garthwaite, J.; Charles, S.L.; Chess-Williams, R. Endothelium-derived relaxing factor release on activation of NMDA receptors suggests role as intercellular messenger in the brain. Nature 1988, 336, 385–388. [Google Scholar] [CrossRef]
- Oess, S.; Icking, A.; Fulton, D.; Govers, R.; Muller-Esterl, W. Subcellular targeting and trafficking of nitric oxide synthases. Biochem. J. 2006, 396, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Gutierrez, V.P.; Zambelli, V.O.; Picolo, G.; Chacur, M.; Sampaio, S.C.; Brigatte, P.; Konno, K.; Cury, Y. The peripheral L-arginine-nitric oxide-cyclic GMP pathway and ATP-sensitive K+ channels are involved in the antinociceptive effect of crotalphine on neuropathic pain in rats. Behav. Pharmacol. 2012, 23, 14–24. [Google Scholar] [CrossRef]
- Zulazmi, N.A.; Gopalsamy, B.; Min, J.C.; Farouk, A.A.; Sulaiman, M.R.; Bharatham, B.H.; Perimal, E.K. Zerumbone Alleviates Neuropathic Pain through the Involvement of l-Arginine-Nitric Oxide-cGMP-K+ ATP Channel Pathways in Chronic Constriction Injury in Mice Model. Molecules 2017, 22, 555. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocana, M.; Cendan, C.M.; Cobos, E.J.; Entrena, J.M.; Baeyens, J.M. Potassium channels and pain: Present realities and future opportunities. Eur. J. Pharmacol. 2004, 500, 203–219. [Google Scholar] [CrossRef]
- Jain, N.K.; Patil, C.S.; Singh, A.; Kulkarni, S.K. Sildenafil, a phosphodiesterase-5 inhibitor, enhances the antinociceptive effect of morphine. Pharmacology 2003, 67, 150–156. [Google Scholar] [CrossRef]
- Yamazumi, I.; Okuda, T.; Koga, Y. Involvement of potassium channels in spinal antinociceptions induced by fentanyl, clonidine and bethanechol in rats. Jpn. J. Pharmacol. 2001, 87, 268–276. [Google Scholar] [CrossRef] [Green Version]
- Ahmad, W.; Kumolosasi, E.; Jantan, I.; Bukhari, S.N.; Jasamai, M. Effects of novel diarylpentanoid analogues of curcumin on secretory phospholipase A2, cyclooxygenases, lipo-oxygenase, and microsomal prostaglandin E synthase-1. Chem. Biol. Drug Des. 2014, 83, 670–681. [Google Scholar] [CrossRef]
- Lee, K.H.; Ab Aziz, F.H.; Syahida, A.; Abas, F.; Shaari, K.; Israf, D.A.; Lajis, N.H. Synthesis and biological evaluation of curcumin-like diarylpentanoid analogues for anti-inflammatory, antioxidant and anti-tyrosinase activities. Eur. J. Med. Chem. 2009, 44, 3195–3200. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Li, X.; Chen, L.; Yang, S.; Wu, X.; Studer, E.; Gurley, E.; Hylemon, P.B.; Ye, F.; Li, Y.; et al. Synthesis and anti-inflammatory activities of mono-carbonyl analogues of curcumin. Bioorg. Med. Chem. Lett. 2008, 18, 1525–1529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, C.; Yang, J.; Wang, Y.; Liang, D.; Yang, X.; Li, X.; Wu, J.; Wu, X.; Yang, S.; Li, X.; et al. Synthesis of mono-carbonyl analogues of curcumin and their effects on inhibition of cytokine release in LPS-stimulated RAW 264.7 macrophages. Bioorg. Med. Chem. 2010, 18, 2388–2393. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Yang, S.; Zhou, H.; Shao, L.; Huang, K.; Xiao, J.; Huang, Z.; Li, X. Synthesis, crystal structure and anti-inflammatory properties of curcumin analogues. Eur. J. Med. Chem. 2009, 44, 915–919. [Google Scholar] [CrossRef]
- Anand, P.; Kunnumakkara, A.B.; Newman, R.A.; Aggarwal, B.B. Bioavailability of curcumin: Problems and promises. Mol. Pharm. 2007, 4, 807–818. [Google Scholar] [CrossRef]
- Singh, R.S.; Das, U.; Dimmock, J.R.; Alcorn, J. A general HPLC-UV method for the quantitative determination of curcumin analogues containing the 1,5-diaryl-3-oxo-1,4-pentadienyl pharmacophore in rat biomatrices. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2010, 878, 2796–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Zhao, C.; He, W.; Wang, Z.; Fang, Q.; Xiao, B.; Liu, Z.; Liang, G.; Yang, S. Synthesis, characterization, and catalytic property of nanosized MgO flakes with different shapes. J. Alloys Compd. 2014, 590, 373–379. [Google Scholar] [CrossRef]
- Leong, S.W.; Faudzi, S.M.; Abas, F.; Aluwi, M.F.; Rullah, K.; Wai, L.K.; Bahari, M.N.; Ahmad, S.; Tham, C.L.; Shaari, K.; et al. Synthesis and sar study of diarylpentanoid analogues as new anti-inflammatory agents. Molecules 2014, 19, 16058–16081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leong, S.W.; Mohd Faudzi, S.M.; Abas, F.; Mohd Aluwi, M.F.; Rullah, K.; Lam, K.W.; Abdul Bahari, M.N.; Ahmad, S.; Tham, C.L.; Shaari, K.; et al. Nitric oxide inhibitory activity and antioxidant evaluations of 2-benzoyl-6-benzylidenecyclohexanone analogs, a novel series of curcuminoid and diarylpentanoid derivatives. Bioorg. Med. Chem. Lett. 2015, 25, 3330–3337. [Google Scholar] [CrossRef]
- Anbar, M.; Gratt, B.M. Role of nitric oxide in the physiopathology of pain. J. Pain Symptom Manag. 1997, 14, 225–254. [Google Scholar] [CrossRef]
- Perimal, E.K.; Akhtar, M.N.; Mohamad, A.S.; Khalid, M.H.; Ming, O.H.; Khalid, S.; Tatt, L.M.; Kamaldin, M.N.; Zakaria, Z.A.; Israf, D.A.; et al. Zerumbone-induced antinociception: Involvement of the L-arginine-nitric oxide-cGMP -PKC-K+ ATP channel pathways. Basic Clin. Pharmacol. Toxicol. 2011, 108, 155–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boger, R.H.; Bode-Boger, S.M. The clinical pharmacology of L-arginine. Annu. Rev. Pharmacol. Toxicol. 2001, 41, 79–99. [Google Scholar] [CrossRef] [Green Version]
- Hughes, S.R.; Williams, T.J.; Brain, S.D. Evidence that endogenous nitric oxide modulates oedema formation induced by substance P. Eur. J. Pharmacol. 1990, 191, 481–484. [Google Scholar] [CrossRef]
- Ialenti, A.; Ianaro, A.; Moncada, S.; Di Rosa, M. Modulation of acute inflammation by endogenous nitric oxide. Eur. J. Pharmacol. 1992, 211, 177–182. [Google Scholar] [CrossRef]
- Pfeiffer, S.; Leopold, E.; Schmidt, K.; Brunner, F.; Mayer, B. Inhibition of nitric oxide synthesis by NG-nitro-L-arginine methyl ester (L-NAME): Requirement for bioactivation to the free acid, NG-nitro-L-arginine. Br. J. Pharmacol. 1996, 118, 1433–1440. [Google Scholar] [CrossRef] [PubMed]
- MacMicking, J.; Xie, Q.W.; Nathan, C. Nitric oxide and macrophage function. Annu. Rev. Immunol. 1997, 15, 323–350. [Google Scholar] [CrossRef]
- Meller, S.T.; Gebhart, G.F. Nitric oxide (NO) and nociceptive processing in the spinal cord. Pain 1993, 52, 127–136. [Google Scholar] [CrossRef]
- Lam, H.H.; Hanley, D.F.; Trapp, B.D.; Saito, S.; Raja, S.; Dawson, T.M.; Yamaguchi, H. Induction of spinal cord neuronal nitric oxide synthase (NOS) after formalin injection in the rat hind paw. Neurosci. Lett. 1996, 210, 201–204. [Google Scholar] [CrossRef]
- Kubes, P.; Suzuki, M.; Granger, D.N. Nitric oxide: An endogenous modulator of leukocyte adhesion. Proc. Natl. Acad. Sci. USA 1991, 88, 4651–4655. [Google Scholar] [CrossRef] [Green Version]
- Abacioglu, N.; Tunctan, B.; Akbulut, E.; Cakici, I. Participation of the components of L-arginine/nitric oxide/cGMP cascade by chemically-induced abdominal constriction in the mouse. Life Sci. 2000, 67, 1127–1137. [Google Scholar] [CrossRef]
- Aley, K.O.; McCarter, G.; Levine, J.D. Nitric oxide signaling in pain and nociceptor sensitization in the rat. J. Neurosci. 1998, 18, 7008–7014. [Google Scholar] [CrossRef] [Green Version]
- Haley, J.E.; Dickenson, A.H.; Schachter, M. Electrophysiological evidence for a role of nitric oxide in prolonged chemical nociception in the rat. Neuropharmacology 1992, 31, 251–258. [Google Scholar] [CrossRef]
- Malmberg, A.B.; Yaksh, T.L. Spinal nitric oxide synthesis inhibition blocks NMDA-induced thermal hyperalgesia and produces antinociception in the formalin test in rats. Pain 1993, 54, 291–300. [Google Scholar] [CrossRef]
- Chen, Y.; Boettger, M.K.; Reif, A.; Schmitt, A.; Uceyler, N.; Sommer, C. Nitric oxide synthase modulates CFA-induced thermal hyperalgesia through cytokine regulation in mice. Mol. Pain 2010, 6, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawabata, A.; Manabe, S.; Manabe, Y.; Takagi, H. Effect of topical administration of L-arginine on formalin-induced nociception in the mouse: A dual role of peripherally formed NO in pain modulation. Br. J. Pharmacol. 1994, 112, 547–550. [Google Scholar] [CrossRef] [Green Version]
- de Moura, R.S.; Rios, A.A.; Santos, E.J.; Nascimento, A.B.; de Castro Resende, A.; Neto, M.L.; de Oliveira, L.F.; Mendes Ribeiro, A.C.; Tano, T. Role of the NO-cGMP pathway in the systemic antinociceptive effect of clonidine in rats and mice. Pharmacol. Biochem. Behav. 2004, 78, 247–253. [Google Scholar] [CrossRef]
- Ferreira, J.; Santos, A.R.; Calixto, J.B. The role of systemic, spinal and supraspinal L-arginine-nitric oxide-cGMP pathway in thermal hyperalgesia caused by intrathecal injection of glutamate in mice. Neuropharmacology 1999, 38, 835–842. [Google Scholar] [CrossRef]
- Lazaro-Ibanez, G.G.; Torres-Lopez, J.E.; Granados-Soto, V. Participation of the nitric oxide-cyclic GMP-ATP-sensitive K+ channel pathway in the antinociceptive action of ketorolac. Eur. J. Pharmacol. 2001, 426, 39–44. [Google Scholar] [CrossRef]
- Ortiz, M.I.; Granados-Soto, V.; Castaneda-Hernandez, G. The NO-cGMP-K+ channel pathway participates in the antinociceptive effect of diclofenac, but not of indomethacin. Pharmacol. Biochem. Behav. 2003, 76, 187–195. [Google Scholar] [CrossRef]
- Sachs, D.; Cunha, F.Q.; Ferreira, S.H. Peripheral analgesic blockade of hypernociception: Activation of arginine/NO/cGMP/protein kinase G/ATP-sensitive K+ channel pathway. Proc. Natl. Acad. Sci. USA 2004, 101, 3680–3685. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, G.A.; Sachs, D.; Cunha, F.Q.; Vale, M.L.; Lotufo, C.M.; Ferreira, S.H.; Ribeiro, R.A. Peripheral antinociceptive effect of pertussis toxin: Activation of the arginine/NO/cGMP/PKG/ ATP-sensitive K channel pathway. Eur. J. Neurosci. 2006, 24, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Pacheco, A.; Araiza-Saldana, C.I.; Granados-Soto, V.; Mixcoatl-Zecuatl, T. Possible participation of the nitric oxide-cyclic GMP-protein kinase G-K+ channels pathway in the peripheral antinociception of melatonin. Eur. J. Pharmacol. 2008, 596, 70–76. [Google Scholar] [CrossRef]
- Ortiz, M.I.; Medina-Tato, D.A.; Sarmiento-Heredia, D.; Palma-Martinez, J.; Granados-Soto, V. Possible activation of the NO-cyclic GMP-protein kinase G-K+ channels pathway by gabapentin on the formalin test. Pharmacol. Biochem. Behav. 2006, 83, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Alves, D.; Duarte, I. Involvement of ATP-sensitive K+ channels in the peripheral antinociceptive effect induced by dipyrone. Eur. J. Pharmacol. 2002, 444, 47–52. [Google Scholar] [CrossRef]
- Jesse, C.R.; Savegnago, L.; Nogueira, C.W. Role of nitric oxide/cyclic GMP/K+ channel pathways in the antinociceptive effect caused by 2,3-bis(mesitylseleno)propenol. Life Sci. 2007, 81, 1694–1702. [Google Scholar] [CrossRef]
- Amoroso, S.; Schmid-Antomarchi, H.; Fosset, M.; Lazdunski, M. Glucose, sulfonylureas, and neurotransmitter release: Role of ATP-sensitive K+ channels. Science 1990, 247, 852–854. [Google Scholar] [CrossRef]
- Mohamad, A.S.; Akhtar, M.N.; Khalivulla, S.I.; Perimal, E.K.; Khalid, M.H.; Ong, H.M.; Zareen, S.; Akira, A.; Israf, D.A.; Lajis, N.; et al. Possible participation of nitric oxide/cyclic guanosine monophosphate/protein kinase C/ATP-sensitive K+ channels pathway in the systemic antinociception of flavokawin B. Basic Clin. Pharmacol. Toxicol. 2011, 108, 400–405. [Google Scholar] [CrossRef] [PubMed]
- Khalid, M.H.; Akhtar, M.N.; Mohamad, A.S.; Perimal, E.K.; Akira, A.; Israf, D.A.; Lajis, N.; Sulaiman, M.R. Antinociceptive effect of the essential oil of Zingiber zerumbet in mice: Possible mechanisms. J. Ethnopharmacol. 2011, 137, 345–351. [Google Scholar] [CrossRef] [PubMed]
- Kamarudin, N.; Hisamuddin, N.; Ong, H.M.; Ahmad Azmi, A.F.; Leong, S.W.; Abas, F.; Sulaiman, M.R.; Shaik Mossadeq, W.M. Analgesic Effect of 5-(3,4-Dihydroxyphenyl)-3-hydroxy-1-(2-hydroxyphenyl)penta-2,4-dien-1-one in Experimental Animal Models of Nociception. Molecules 2018, 23, 2099. [Google Scholar] [CrossRef] [PubMed] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ong, H.M.; Ahmad Azmi, A.F.; Leong, S.W.; Abas, F.; Perimal, E.K.; Farouk, A.A.O.; Israf, D.A.; Sulaiman, M.R. The Involvement of l-Arginine-Nitric Oxide-cGMP-ATP-Sensitive K+ Channel Pathway in Antinociception of BBHC, a Novel Diarylpentanoid Analogue, in Mice Model. Molecules 2021, 26, 7431. https://doi.org/10.3390/molecules26247431
Ong HM, Ahmad Azmi AF, Leong SW, Abas F, Perimal EK, Farouk AAO, Israf DA, Sulaiman MR. The Involvement of l-Arginine-Nitric Oxide-cGMP-ATP-Sensitive K+ Channel Pathway in Antinociception of BBHC, a Novel Diarylpentanoid Analogue, in Mice Model. Molecules. 2021; 26(24):7431. https://doi.org/10.3390/molecules26247431
Chicago/Turabian StyleOng, Hui Ming, Ahmad Farhan Ahmad Azmi, Sze Wei Leong, Faridah Abas, Enoch Kumar Perimal, Ahmad Akira Omar Farouk, Daud Ahmad Israf, and Mohd Roslan Sulaiman. 2021. "The Involvement of l-Arginine-Nitric Oxide-cGMP-ATP-Sensitive K+ Channel Pathway in Antinociception of BBHC, a Novel Diarylpentanoid Analogue, in Mice Model" Molecules 26, no. 24: 7431. https://doi.org/10.3390/molecules26247431