3.1. Physicochemical Parameters
The physicochemical parameters of the investigated cambucis reflected accession diversity, as expected, since cambuci have not yet been selected in genetic breeding programs (
Table 1). The average fresh fruit mass was 57.0 g (CV: 26.2%). Small producers and fruit collectors from the Serra do Mar region employ an informal cambuci classification, where large fruits are considered as those over 68 g, medium, from 39 g to 68 g, and small, less than 39 g [
3]. In this study, fresh masses ranged from 36.3 to 88.3 g. Fruit height (h) was always smaller than their diameter (d), with and h/d ratio <1, indicating flat fruits in accordance with [
3,
15].
The average SSC of cambuci was 10.8 Brix, ranging from 7.8 to 14.0 Brix. These values are similar to the SSC observed in other chemical characterization studies in cambuci, which reported values between 7.3 to 13.3 Brix and 5.1 to 11.0 Brix [
3,
16]. TA values ranged from 1.1% to 4.3%, which is a large variation from that observed previously—1.3% to 2.9% (citric acid eq.) [
3].
For comparison, in a study evaluating the quality of passion fruits, 9.1%, 11.6% and 12.3% of soluble solids were reported for yellow, purple and orange fruits, respectively [
17]. Similarly, these same fruits contained total titratable acidity values of 2.1%, 2.8% and 2.2%. Less acidic cambucis exhibited TA similar to the average observed in oranges, of 1.0%, while high acidity cambucis are similar to acid limes, presenting TA between 5–6% [
18].
When the SSC/TA ratio is considered as a quality attribute, each fruit will present its own scale, based on sugar and acid contents, as well as other minority components. The SSC/TA ratio can be used as a discriminating quality criterion in fruit, as described previously for peaches and nectarines [
19]. When 39 peach and nectarine cultivars were evaluated, the variation ranged from 9.36 to 92.8 [
19]. For the cambucis assessed in the present study, the variation ranged between 3.1 and 8.0.
Usually, cambuci fruits present the following values of other physical and chemical properties: moisture content 86–90%, pH 2–3, fibers 0.1–0.2%, total lipids 0.02–0.05%, proteins 0.02–0.03%, carbohydrate 0.9 g Kg-1, ash content 0.02%, and mineral profile comprising nitrogen 0.01–0.03, phosphorus 0.01, potassium 0.1, calcium 0.001–0.005, magnesium 0.01, sulfur 0.01, boron 0.1, zinc 0.1, iron 0.2–0.3, manganese 0.02–0.1, copper 0.02–0.04, sodium 0.3–0.6 g kg
−1 DW [
3,
5,
20]. These parameters were previously reported presenting genetic variability values for fruit characters once, when different accessions were evaluated, the coefficient of variance was demonstrated as high levels (>20%), as Bianchini et al. show [
21]. However, there is no report of the complete metabolite profile as a parameter of chemical changes to the point the variability genetic of cambuci fruits as describe in the next section.
3.2. NMR Spectra Metabolite Assignment
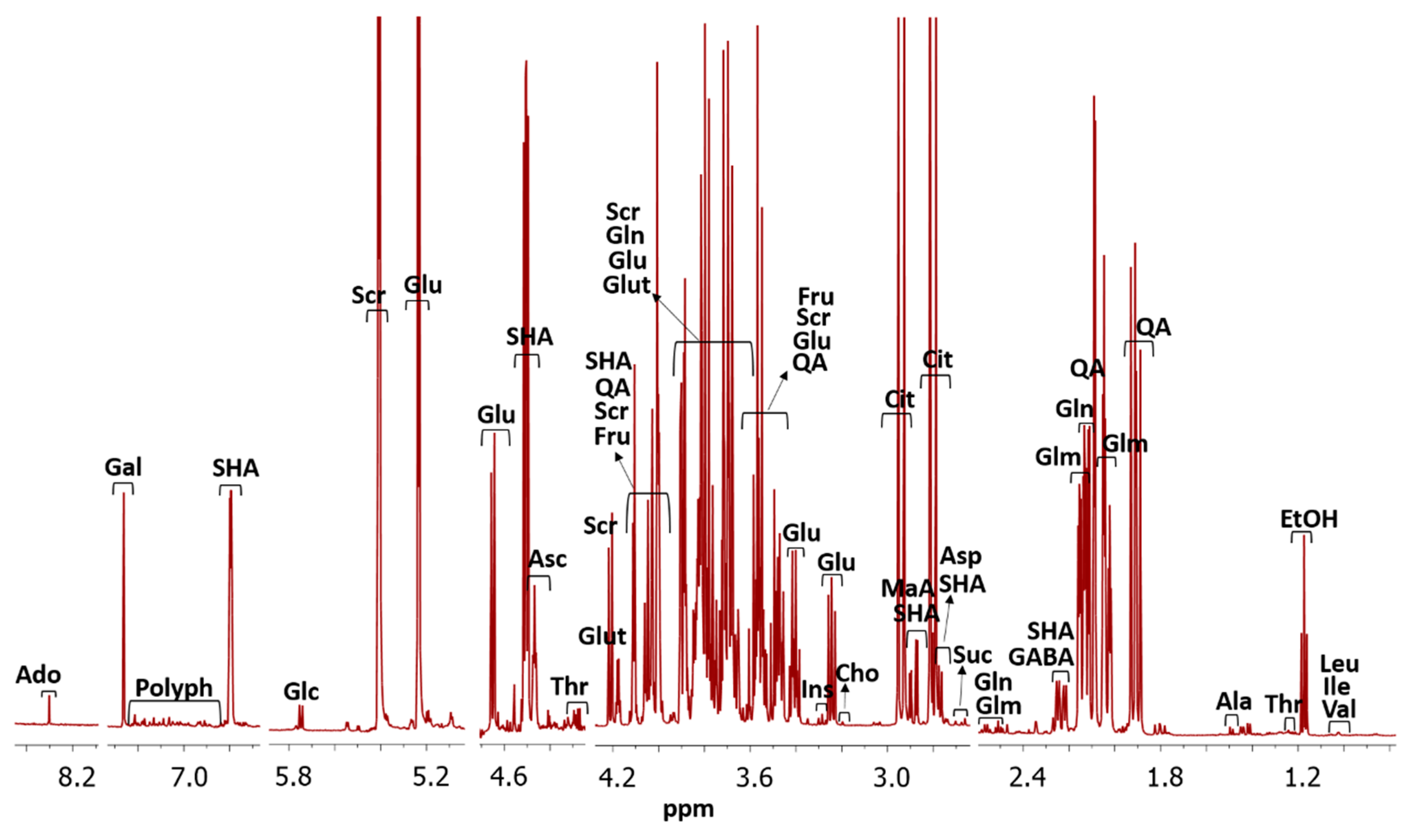
A typical cambuci NMR spectrum of 0.9–8.5 ppm is presented in
Figure 1. Twenty-five metabolites (adenosine-like substances, alanine, ascorbic acid, aspartic acid, choline, citric acid, ethanol, fructose, gamma-aminobutyric acid, gallic acid, glucose-6-phosphate, glutamic acid, glutamine, glucose, glutathione, inositol, isoleucine, leucine, malic acid, quinic acid, sucrose, shikimic acid, succinic acid, threonine and valine) were assigned. This number of compounds, as well as the compounds identified, are in agreement with the literature found for NMR analysis in fruits. For the postharvest quality of peaches and plums, 19 compounds have already been reported, and for tomatoes, 21 compounds [
22]. For banana and jujube quality, the number of compounds identified were 11 and 22, respectively [
6,
23].
The characteristic polyphenol region containing numerous signals is also exhibited, which may be associated to the presence of proanthocyanidins, such as ellagitannins. These characteristic polyphenol signals have also been reported for plums and peaches [
22]. The compound identification was possible due to the comparison of deconvoluted signals of
1H NMR spectrum of a sample with database, and confirmation with the interpretation of
1H-
1H COSY NMR,
1H-
13C HSQC NMR spectrum, and
1H JRES NMR spectrum as showed in
Figure S1 (Supplementary Materials).
The NMR analysis in the present study was possible once careful sample preparation with pH control of the extracts and mathematical alignment of peaks was employed to prevent some compounds which are susceptible to interactions from affecting their chemical shifts and to keep the reproducibility. Additionally, the resolution was guaranteed by the carefully adjusted parameters such as calibrated pulse at 90° and the respective power to suppress water residual, correspond receiver gain (intensity), and the number of points.
For quantification, the spectra showed the metabolite amounts after the signal integration and the software table filled using the
Eretic2 method with metabolites and sample information (
Figure S1, Supplementary Materials). The amounts of metabolites found will be discussed in the next section. When classified by accession (or plant), the spectra showed few changes between themselves (
Figure S2, Supplementary Materials), also better explored in the next section.
3.3. Metabolite Amounts
Under appropriate quantitative NMR (qNMR) conditions, NMR spectra can provide direct quantitative information since the signal intensity of each resonance in the NMR spectrum is directly proportional to the number of equivalent nuclei responsible for that signal, or in other words, is directly proportional to the molar amount of the detected isotope [
24,
25]. Thus, with NMR it is possible to determine absolute concentrations, relying on the use of one internal, external, or electronic standard [
25,
26,
27]. Using the one-dimensional (1D)
1H NMR spectroscopy as the conventional way to perform quantitative NMR analysis [
28], we used the external 2 mM sucrose. Both sample and standard spectrum were measured under a set of appropriate conditions to obtain accurate results in quantitative analysis. As quantitative conditions depend on the targeted accuracy (trueness and precision), for a maximum error of 1%, a relaxation delay (delay before the excitation) was included that was equal to at least 5 times the longitudinal relaxation time (T1) for a 90° pulse, an acquisition time longer than 3 times the transverse relaxation time (T2), and a sufficient signal to noise (at least 50:1) [
24,
27]. Careful processing of NMR spectra was also required to extract accurate peak areas.
Sucrose, citric acid, and glutathione represented the main cambuci pulp constituents, comprising 61% of the total amount of the polar metabolites in this fruit (
Table 2,
Table 3 and
Table 4).
Total sugar content (sucrose, glucose and fructose) represented 40% of all pulp metabolites. A balance between sugars is characteristic of each fruit species and decisively affects the quality perception experienced by consumers. Sucrose was the predominant sugar, although accession comparisons indicate a sucrose content variation of up to 3.9-fold (
Table 2). Sucrose, fructose and glucose determine fruit sweetness, where fructose is perceived as sweeter, followed by sucrose and glucose [
29]. In ‘Puket’ pineapples, the total sugar contents reach 13.2 g 100 g fw, with the sucrose amount being four times higher than that of glucose and fructose [
30]. Strawberries containing 0.2–2.2 g 100 g
−1 of sucrose, 1.7–2.5 g 100 g
−1 of glucose and 1.6–2.9 g 100 g
−1 of fructose [
31] indicate that sweetness varies among cultivars.
Glucose-6-phosphate, the precursor of most glucose-implicated pathways, was present in cambuci from 8.7 mg 100 g
−1 to 51.2 mg 100 g
−1 (
Table 2). The operation of this pathway during the postharvest stage of the fruit is involved with the available energy [
32,
33,
34]. For the cambucis evaluated, the quantification of glucose-6-phosphate was not directly related to the measured levels for the most common sugars in fleshy fruit: sucrose, fructose and glucose. Concerning related compounds, adenosine ribonucleoside and the sugar alcohol inositol were detected in lower amounts. These compounds have been reported in quality NMR analyses in minor amounts for several fruits, such as melons and peaches [
35,
36].
Ethanol content ranged from 3.5 mg 100 g
−1 to 11.3 mg 100 g
−1, indicating ongoing fermentation processes (
Table 2). The presence of this compound was expected, since cambuci is a highly perishable fruit with a short postharvest life, indicating fermentation even before being detached from the plant. Ethanol results from the reduction of acetaldehyde (glycolysis result) during the anaerobic respiration process, responsible for the emission of volatile compounds described as strongly alcoholic, perceived in overripe fruits. High ethanol levels in newly harvested cambucis suggest that the control of this compound may be essential for the successful postharvest conservation of this fruit. Currently, cambucis are mostly consumed frozen. However, using conservation techniques that enable
in natura consumption are promising, such as refrigeration and controlled and modified atmospheres.
Citric acid and shikimic acid were the most important organic acids in cambuci, representing 32% of all metabolites (
Table 3). The citric acid content of cambucis is comparable to several citrus fruits, such as orange (560–980 mg 100 g
−1) and grapefruit (1190–2100 mg 100 g
−1), and is significantly higher than in apples (30–50 mg 100 g
−1) [
37]. Typical shikimic acid values in fruits may range from 0.3 mg 100 g
−1 in apples and 84.4 mg 100 g
−1 in jostaberries [
37]. Nevertheless, reported cambuci values were higher than in these fruits, evidencing their peculiar composition. Furthermore, quinic acid is usually more abundant in fruits than shikimic acid, although this has not been confirmed for cambuci [
37]. Accession 9 contained higher malic acid rates, while accession 6 contained the lowest amount among all evaluated accessions. Malic acid, which is derived from succinic acid, leads to a tart flavor. Quinic acid exhibited the lowest coefficient of variation (15%) in all cambuci accessions. Kiwi is considered a quinic acid-rich fruit, with reference values ranging from 400 to 1300 mg 100 g
−1. The richest quinic acid cambuci accession contained 466.9 mg 100 g
−1 (accession 4) (
Table 3). Quinic, shikimic and gallic acid are known as polyphenol precursors. The mean amounts observed for polyphenols, quinic acid, shikimic acid, and gallic acid were 412.1 mg 100 g
−1, 373.3 mg 100 g
−1, 584.5 mg 100 g
−1, and 4.1 mg 100 g
−1, respectively (
Table 3 and
Table 5). Quinic acid together with gallic acid is involved in the early stage of tannin biosynthesis, which is associated with the typical astringent taste of cambucis [
38]. Tannins are polyphenols belonging to the class of proanthocyanidins that occur in fruits and vegetables. The biosynthesis of tannins results in ellagitannin production, one of the major compounds of the phenolic class for this fruit, considered beneficial for human health, with a protective action against cardiovascular diseases and obesity [
2].
Accession 6 and 8 contained the highest amounts of polyphenols, while accession 3 contained the lowest (
Table 3). A paper assessing phenolic cambuci compounds indicated a potentially beneficial action concerning glucose intolerance attenuation and adipose tissue inflammation induced by a high-fat, high-sucrose diet [
2]. Another study demonstrated polyphenols present in cambuci as important therapeutic actions that improve complications associated with obesity [
39]. In fact, fruits from the botanical family Myrtaceae represent a rich source of secondary metabolites, which in nature are intrinsically involved in plant defense, especially biologically active polyphenols [
40,
41]. Alongside the shikimic acid contents reported herein, these findings point to a balance between polyphenols and shikimic acid (SHA) synthesis, revealing that this organic acid is probably directly involved in polyphenol synthesis. The SHA pathway in plants is responsible for generating numerous secondary metabolites [
42]. It plays an important role as an intermediary for the biosynthesis of aromatic amino acids (tryptophan, tyrosine, and phenylalanine) and has extensive biotechnological applications, with analgesic, antioxidant, anticoagulant, anti-inflammatory, antithrombotic, neuroprotective effects and as material for the synthesis of antivirals [
43,
44,
45,
46,
47].
Ascorbic acid (vitamin C), a natural antioxidant that prevents the action of free radicals, is an important organic acid in cambuci, averaging 47 mg 100 g
−1, ranging from 26 mg 100 g
−1 to 103 mg 100 g
−1 (
Table 3). These values are in agreement with previously reported data, where cambuci ascorbic acid contents ranging from 31.12 to 139.38 mg 100 g
1 were described [
3]. Therefore, cambuci is an excellent ascorbic acid source when compared to sweet orange (18.9–22.2 mg 100 g
−1), mandarins (16.2–31.6 mg 100 g
−1) and lemons (39.6–42.3 mg 100 g
−1) [
48]. The recommended daily dose (RDA) of vitamin C is of 75 (women) −90 (men) mg day
−1, sufficient to meet cellular needs and reduce human health risks, such as cardiovascular and neurodegenerative diseases, cancer and stroke [
49]. This finding could be explored in the food, cosmetics, and pharmaceutical industry as foods with increased vitamin C, cosmetics rich in natural vitamin C and complex vitamins including vitamin C could have a potential international market.
Distinguishing sugar, organic acid and phenolic compound levels can contribute to the quality differentiation of fruits produced from different plants and aid in future efforts to breed cambucis. This type of differentiation has been previously reported for other cultivars, such as apples [
50].
Amino acids’ relevance in plants arises from their role as central regulators of plant growth and responses to environmental signals, along with being agents that influence human food nutritional quality [
51]. Eleven amino acids and related compounds were detected in cambuci (
Table 4), where glutathione was the most noteworthy, representing 10% of all metabolites. Glutathione is a water-soluble antioxidant molecule consisting of three amino acids, glutamic acid, cysteine and glycine (a tripeptide). Glutamic acid amounts ranked second, always at least 2.5-fold lower than glutathione. Glutathione and glutamic acid were highly and positively correlated, with an index of 0.96. Glutamine was also positively correlated with glutamic acid, with an index of 0.94. Glutamic acid is the major amino acid of ripe fruits [
52]. Glutathione, glutamic acid and glutamine displayed the lowest coefficients of variation among all detected amino acids.
Other amino acids and related compounds exhibiting a high positive correlation (>0.9) comprised gamma-aminobutyric acid (GABA) vs. isoleucine, GABA vs. valine, GABA vs. leucine, GABA vs. asparagine, isoleucine vs. valine, isoleucine versus leucine, isoleucine vs. asparagine, valine vs. leucine, and valine vs. asparagine. Amino acids in fruits are not only linked to protein constitution, but also contribute to organoleptic fruit qualities. For example, in tomatoes, the taste described as “umami” is directly associated to glutamic acid levels [
53]. Amino acids are also the precursors of several volatile organic compounds, such as 2- and 3-methyl butanal, 2- and 3-methyl butanol, phenylacetaldehyde, 2-phenylethanol and methyl salicylate. [
5].
The sugar and organic acid ratio (S/OA) ranged between 0.5 and 2.1, indicating both sweet and acidic fruits. The S/OA ratio was also correlated to the SSC/TA ratio, in which 0.5 (S/OA) corresponded to 3 (SSC/TA) and 2.1 (S/OA) to 8 (SSC/TA) (
Table 5). The use of the S/OA ratio to replace the SSC/TA ratio is here presented as a new method to measure the quality attribute. Thus, NMR quantification of sugars and organic acids can be used to establish or replace the optimal values of the SSC/TA ratio of an unknown fruit such as cambuci, predicting the sweetness of the fruit, and guiding genetic breeding programs of the fruit.
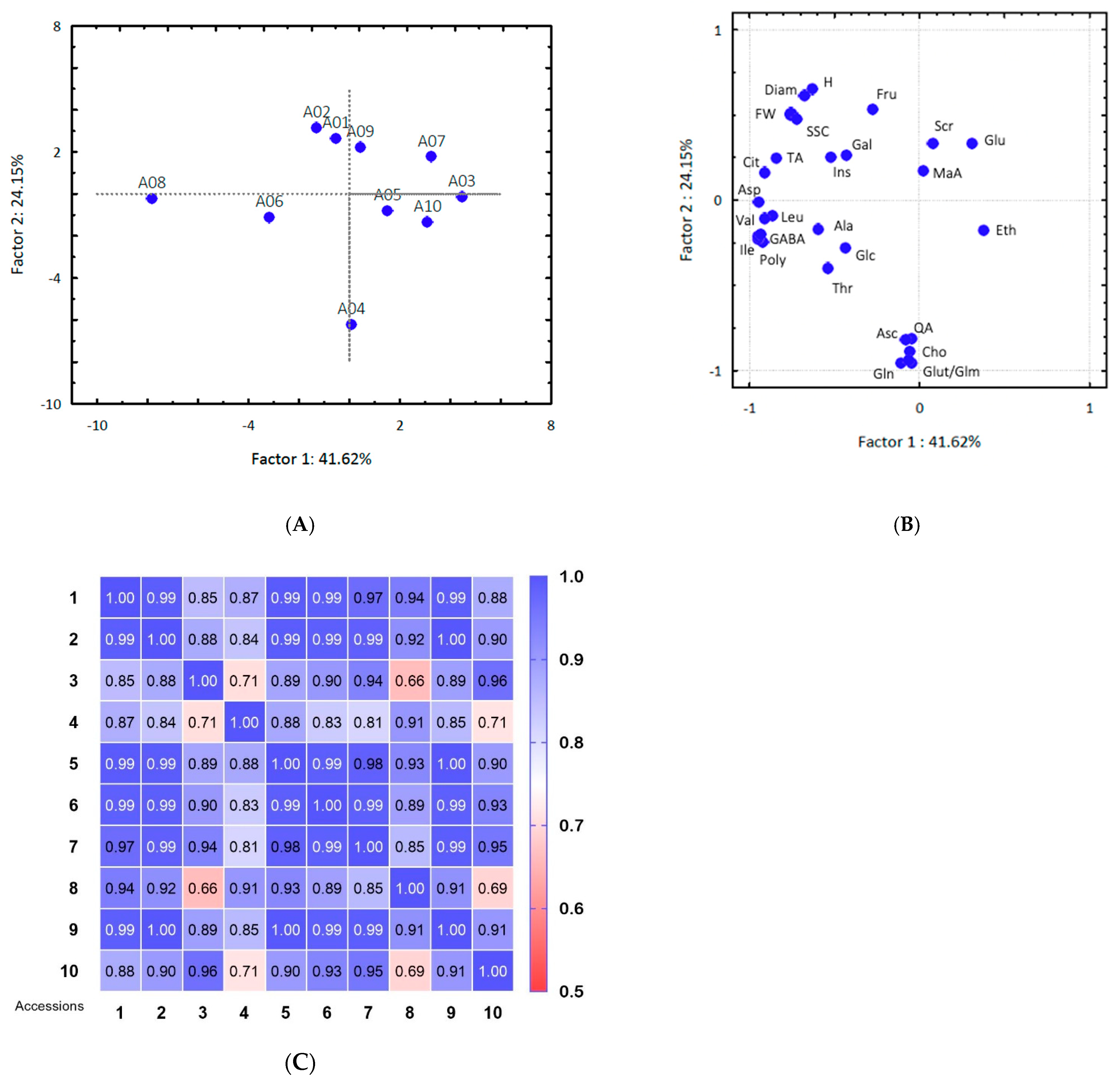
Furthermore, a lower sugar content was detected in accession 4, and lower organic acid and polyphenols, in accession 3. Accessions 3, 4, and 6 were obtained from the same city, under similar environmental conditions, which emphasizes possible genetic differences among producing plants.
Accessions 3 and 4 stood out for their visible separation tendency while accessions 01 and 02, from Paraibuna-SP, showed high similarity (
Figure 2A). Nevertheless, similarities and differences in fruit pulp profiles were observed between fruits harvested in the same municipality and between municipalities for the remaining accessions. Ethanol was relevant to distinguish accessions 3 from 4 (
Figure 2B), gallic acid to distinguish accession 6 from 3 and 4.
Therefore, the fruits could not be clearly separated by their municipality of origin. A range of 66% to 100% similarity in fruit pulp constituents was found by correlating parameters reported in this study (
Figure 2C).
3.4. Metabolic Pathway of Common Metabolites
The metabolic pathway analysis indicated a total of 36 pathways (
Table 6). The top 12 metabolic pathway of common cambuci metabolites described according to impact factor are displayed in
Figure 3.
When assessing the top 12 relevant pathways, only seven exhibited a pathway impact value higher than 0.1, as follows: (1) alanine, aspartate and glutamate metabolism, (2) starch and sucrose metabolism, (3) glutathione metabolism, (4) citrate cycle (TCA cycle), (5) butanoate metabolism, (6) glycine, serine and threonine metabolism, and (7) inositol phosphate metabolism, with impact values of 0.77, 0.53, 0.40, 0.16, 0.14, 0.12, and 0.10, respectively. Based on both their p values and impact values, this relevance can be further restricted to five metabolisms, namely (1), (2), (4), (5), and (6), with significant p values <0.05 (−log p > 3.15).
The alanine, aspartate and glutamate metabolism is the most relevant for cambucis, associated to their fundamental involvement as a central molecule of the amino acid metabolism in higher plants [
54] (
Table 6,
Figure 3). Amino acids are involved in reactions in the primary and secondary metabolism of plants including those processes linked to ripening that directly deal with fruit quality once it has been harvested [
55]. Amino acids are essential agents in physiological and biochemical reactions during maturation and senescence, having functions in the carbon and nitrogen cycle and playing vital roles during signaling processes in plant stress response [
55]. Regarding fruit metabolism, it was described in figs under refrigeration that the behavior of glutamic acid and aspartic acid levels presents a positive correlation with increased soluble solids, glucose, fructose and the loss of fresh weight [
56]. This relation was only observed with aspartic acid in cambucis, where accession 6 that presented the greatest amount of soluble solids, sucrose, glucose and fructose also had aspartic acid amounts higher than the mean (
Table 4).
Sucrose,
d-glucose and
d-fructose are the main starch and sucrose metabolism components (
Table 6,
Figure 2). The balance between sugars is dependent on fruit genotypes, as noted for pomegranates, where variability in eight detected sugars was significantly high in different accessions [
57]. This difference relates closely to the activity of the enzymes involved in the production of sugars, and therefore the sugar content of the fruit alters in line with the activity level of each one. The evaluated cambucis are fruits from wild accessions, which did not go through breeding, and therefore the sugar profile is related to what is found directly in nature. In this way, their sugars play roles essentially related to the survival of the species as sensing metabolism, fruit development, facilitating the formation of the cell wall, cell expansion, nourishes respiration and acid metabolism [
58,
59].
The third relevant cambuci metabolism was the TCA cycle, with malic acid, citric acid and succinic acid as the most noteworthy acids (
Table 6,
Figure 3). During fruit ripening, sugars may no longer be available for respiration, causing a change from sugars to organic acids (in particular citric acid) as a respiratory substrate [
60]. In this phase, increased respiration may stimulate the conversion of citric acid into malate, in order to maintain the set of intermediate products in the TCA cycle constant [
60]. All metabolic pathways linked to the TCA cycle depend on the transient formation of succinic acid, including the carbon metabolism and lipid, amino acid and GABA syntheses [
61].
The fourth most representative pathway (butanoate metabolism) in cambuci is related to γ-aminobutyric acid (GABA, or 4-aminobutyrate), also derived from glutamic acid (
Table 6,
Figure 3). Glutamic acid is decarboxylated into GABA by the action of glutamic acid decarboxylate (GAD) in cytosol [
54]. GABA plays a dual role as a signaling molecule and as a metabolite. Among other possibilities, stresses such as cold, heat, salt and soft or transient environmental factors such as touch, wind or rain, inherent to cambuci production and postharvest stages, are known to increase cellular Ca
2+ levels [
62]. In addition, the versatility of GABA in dealing with biotic stress factors shapes plant immune responses against pathogens by modulating the balance of reactive oxygen species in plant tissues [
63].
Some amino acids are synthesized as a consequence of the association between GABA production and the C and N metabolism, both under both normal and stress conditions [
64,
65], such as threonine. Threonine is part of the glycine, serine and threonine metabolism, the fifth metabolic pathway recognized as important in cambucis. In tomatoes, a significant increase in threonine levels at the stage classified as mature green has been observed, which remained high until the end of this fruit’s postharvest life [
66]. In kiwis, an increase in threonine in the fruit pulp was observed up to the stage classified as eating-ripe (soft), decaying from the stage classified as overripe [
67]. Another nitrogen compound involved in this pathway is choline, an amino acid-like nutrient already described as one of the main metabolites responsible for the senescence in bananas [
6].



 ,
, 




{kind=link}
{kind=link}
{kind=link}
{kind=link}