
Genomics-Driven Activation of Silent Biosynthetic Gene Clusters in Burkholderia gladioli by Screening Recombineering System

 , and
, and 
Abstract
:
1. Introduction
2. Results and Discussion
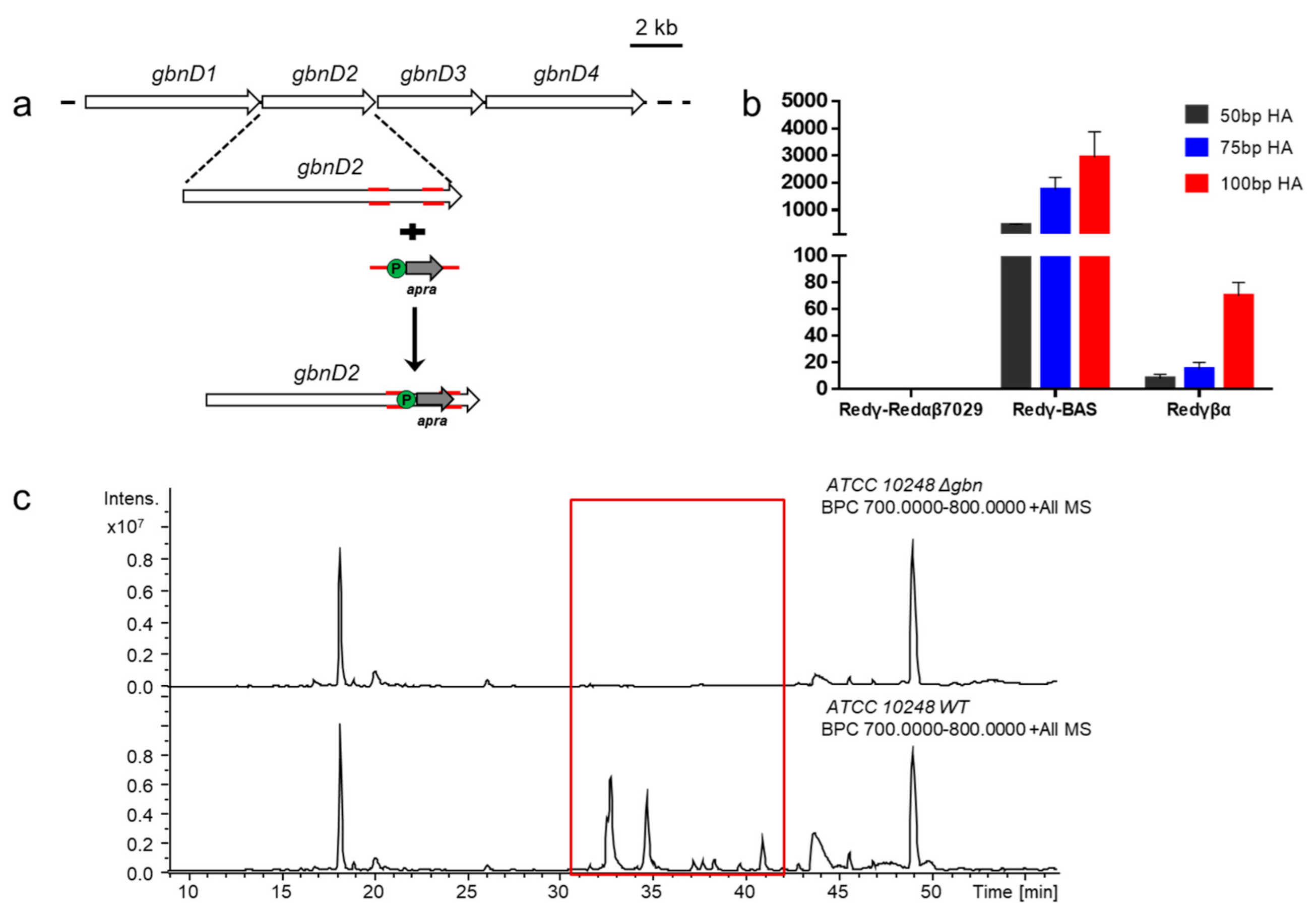
2.1. Bioinformatic Analysis and Manipulation of Silent BGCs in B. gladioli ATCC 10248
2.2. Screening an Available Recombineering Genome Editing System for B. gladioli ATCC 10248
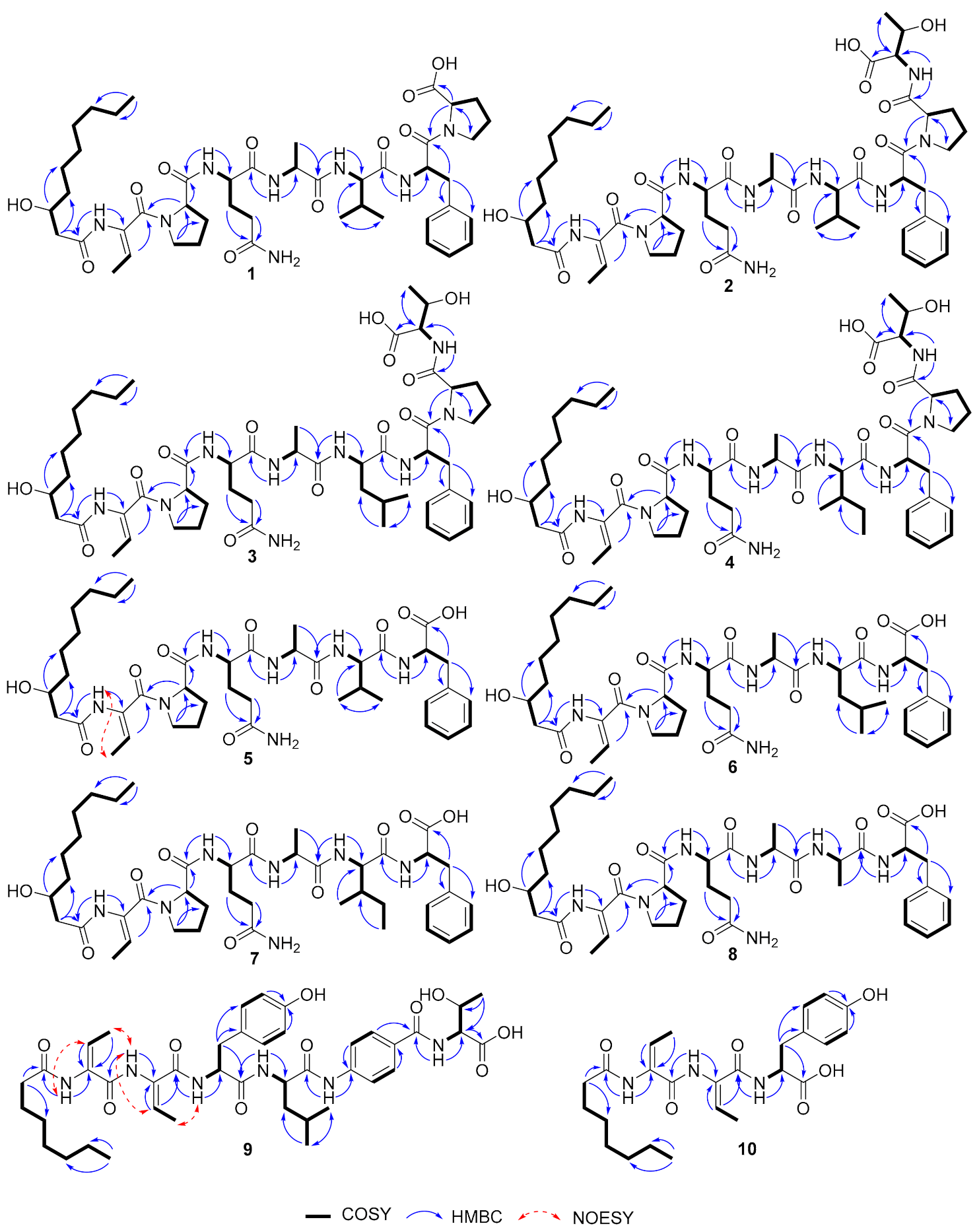
2.3. Activating Two Silent Biosynthetic Gene Clusters and Structure Elucidation of Lipopeptides
2.4. Biosynthesis of Burriogladiodins and Haereogladiodins
2.5. The Bioactivity Assays of Burriogladiodins and Haereogladiodins
3. Materials and Methods
3.1. General Experimental Procedures
3.2. Bacterial Strains, Plasmids and Reagents
3.3. The Construction of Optimized Recombination System in B. gladioli ATCC 10248
3.4. Knockout and Promoter Insertion of the Silent Gene Clusters on the Chromosome of B. gladioli ATCC 10248
3.5. Extraction and Isolation
3.6. Feeding of Labeled Amino Acids
3.7. Antibacterial and Cytotoxic Activities Assay
3.8. Swarming and Swimming Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Sample Availability
References
- Morya, R.; Salvachúa, D.; Thakur, I.S. Burkholderia: An untapped but promising bacterial genus for the conversion of aromatic compounds. Trends Biotechnol. 2020, 38, 963–975. [Google Scholar] [CrossRef]
- Kunakom, S.; Eustaquio, A.S. Burkholderia as a source of natural products. J. Nat. Prod. 2019, 82, 2018–2037. [Google Scholar] [CrossRef]
- Esmaeel, Q.; Pupin, M.; Jacques, P.; Leclere, V. Nonribosomal peptides and polyketides of Burkholderia: New compounds potentially implicated in biocontrol and pharmaceuticals. Environ. Sci. Pollut. Res. Int. 2018, 25, 29794–29807. [Google Scholar] [CrossRef]
- Chen, H.; Bian, Z.; Ravichandran, V.; Li, R.; Sun, Y.; Huo, L.; Fu, J.; Bian, X.; Xia, L.; Tu, Q.; et al. Biosynthesis of polyketides by trans-AT polyketide synthases in Burkholderiales. Crit. Rev. Microbiol. 2019, 45, 162–181. [Google Scholar] [CrossRef] [PubMed]
- Depoorter, E.; Bull, M.J.; Peeters, C.; Coenye, T.; Vandamme, P.; Mahenthiralingam, E. Burkholderia: An update on taxonomy and biotechnological potential as antibiotic producers. Appl. Microbiol. Biotechnol. 2016, 100, 5215–5229. [Google Scholar] [CrossRef] [PubMed]
- Mullins, A.J.; Murray, J.A.H.; Bull, M.J.; Jenner, M.; Jones, C.; Webster, G.; Green, A.E.; Neill, D.R.; Connor, T.R.; Parkhill, J.; et al. Genome mining identifies cepacin as a plant-protective metabolite of the biopesticidal bacterium Burkholderia ambifaria. Nat. Microbiol. 2019, 4, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Esmaeel, Q.; Pupin, M.; Kieu, N.P.; Chataigné, G.; Béchet, M.; Deravel, J.; Krier, F.; Höfte, M.; Jacques, P.; Leclère, V. Burkholderia genome mining for nonribosomal peptide synthetases reveals a great potential for novel siderophores and lipopeptides synthesis. Microbiol. Open 2016, 5, 512–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dose, B.; Ross, C.; Niehs, S.P.; Scherlach, K.; Bauer, J.P.; Hertweck, C. Food-poisoning bacteria employ a citrate synthase and a type II NRPS to synthesize bolaamphiphilic lipopeptide antibiotics. Angew. Chem. Int. Ed. 2020, 59, 21535–21540. [Google Scholar] [CrossRef] [PubMed]
- Dashti, Y.; Nakou, I.; Mullins, A.; Webster, G.; Jian, X.; Mahenthiralingam, E.; Challis, G.L. Discovery and biosynthesis of bolagladins: Unusual lipodepsipeptides from Burkholderia gladioli clinical isolates. Angew. Chem. Int. Ed. 2020, 59, 21553–21561. [Google Scholar] [CrossRef] [PubMed]
- Niehs, S.P.; Kumpfmuller, J.; Dose, B.; Little, R.F.; Ishida, K.; Florez, L.V.; Kaltenpoth, M.; Hertweck, C. Insect-associated bacteria assemble the antifungal butenolide gladiofungin by non-canonical polyketide chain termination. Angew. Chem. Int. Ed. 2020, 59, 23122–23126. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Henkes, L.M.; Doughty, L.B.; He, M.; Wang, D.; Meyer-Almes, F.J.; Cheng, Y.Q. Thailandepsins: Bacterial products with potent histone deacetylase inhibitory activities and broad-spectrum antiproliferative activities. J. Nat. Prod. 2011, 74, 2031–2038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biggins, J.B.; Gleber, C.D.; Brady, S.F. Acyldepsipeptide HDAC inhibitor production induced in Burkholderia thailandensis. Org. Lett. 2011, 13, 1536–1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Zhou, H.; Chen, H.; Jing, X.; Zheng, W.; Li, R.; Sun, T.; Liu, J.; Fu, J.; Huo, L.; et al. Discovery of recombinases enables genome mining of cryptic biosynthetic gene clusters in Burkholderiales species. Proc. Natl. Acad. Sci. USA 2018, 115, E4255–E4263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, H.; Zhou, H.; Sun, T.; Xu, J.; Tu, Q.; Yang, J.; Zhang, Y.; Bian, X. Identification of Holrhizins E-Q Reveals the Diversity of Nonribosomal Lipopeptides in Paraburkholderia rhizoxinica. J. Nat. Prod. 2020, 83, 537–541. [Google Scholar] [CrossRef] [PubMed]
- Zheng, W.; Wang, X.; Zhou, H.; Zhang, Y.; Li, A.; Bian, X. Establishment of recombineering genome editing system in Paraburkholderia megapolitana empowers activation of silent biosynthetic gene clusters. Microb. Biotechnol. 2020, 13, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Hildebrand, D.C.; Palleroni, N.J.; Doudoroff, M. Synonymy of Pseudomonas gladioli Severini 1913 and Pseudomonas marginata (McCulloch 1921) Stapp 1928. Int. J. Syst. Bacteriol. 1973, 23, 433–437. [Google Scholar] [CrossRef] [Green Version]
- Johnson, S.L.; Bishop-Lilly, K.A.; Ladner, J.T.; Daligault, H.E.; Davenport, K.W.; Jaissle, J.; Frey, K.G.; Koroleva, G.I.; Bruce, D.C.; Coyne, S.R.; et al. Complete genome sequences for 59 burkholderia isolates, both pathogenic and near neighbor. Genome Announc. 2015, 3, e00159-15. [Google Scholar] [CrossRef] [Green Version]
- Blin, K.; Shaw, S.; Steinke, K.; Villebro, R.; Ziemert, N.; Lee, S.Y.; Medema, M.H.; Weber, T. AntiSMASH 5.0: Updates to the secondary metabolite genome mining pipeline. Nucleic Acids Res. 2019, 47, W81–W87. [Google Scholar] [CrossRef] [Green Version]
- Song, L.; Jenner, M.; Masschelein, J.; Cerith, J.; Bull, M.J.; Harris, S.R.; Hartkoorn, R.C.; Vocat, A.; Romero-Canelon, I.; Coupland, P.; et al. Discovery and biosynthesis of gladiolin: A Burkholderia gladioli antibiotic with promising activity against mycobacterium tuberculosis. J. Am. Chem. Soc. 2017, 139, 7974–7981. [Google Scholar] [CrossRef] [Green Version]
- Dose, B.; Niehs, S.P.; Scherlach, K.; Florez, L.V.; Kaltenpoth, M.; Hertweck, C. Unexpected bacterial origin of the antibiotic icosalide: Two-tailed depsipeptide assembly in multifarious Burkholderia symbionts. ACS Chem. Biol. 2018, 13, 2414–2420. [Google Scholar] [CrossRef]
- Horsman, M.E.; Marous, D.R.; Li, R.; Oliver, R.A.; Byun, B.; Emrich, S.J.; Boggess, B.; Townsend, C.A.; Mobashery, S. Whole-genome shotgun sequencing of two β-proteobacterial species in search of the bulgecin biosynthetic cluster. ACS Chem. Biol. 2017, 12, 2552–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baral, B.; Akhgari, A.; Metsä-Ketelä, M. Activation of microbial secondary metabolic pathways: Avenues and challenges. Synth. Syst. Biotechnol. 2018, 3, 163–178. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Bian, X.; Hu, S.; Wang, H.; Huang, F.; Seibert, P.M.; Plaza, A.; Xia, L.; Müller, R.; Stewart, A.F.; et al. Full-length RecE enhances linear-linear homologous recombination and facilitates direct cloning for bioprospecting. Nat. Biotechnol. 2012, 30, 440–446. [Google Scholar] [CrossRef] [PubMed]
- Tang, B.; Yu, Y.; Liang, J.; Zhang, Y.; Bian, X.; Zhi, X.; Ding, X. Reclassification of Polyangium brachysporum DSM 7029 as Schlegelella brevitalea sp. nov. Int. J. Syst. Evol. Microbiol. 2019, 69, 2877–2883. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Zheng, W.; Gao, Y.; Jiang, C.; Shi, H.; Diao, X.; Li, S.; Chen, H.; Wang, H.; Li, R.; et al. Single-stranded DNA-binding protein and exogenous RecBCD inhibitors enhance phage-derived homologous recombination in Pseudomonas. iScience 2019, 14, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Götze, S.; Stallforth, P. Structure, properties, and biological functions of nonribosomal lipopeptides from pseudomonads. Nat. Prod. Rep. 2020, 37, 29–54. [Google Scholar] [CrossRef] [PubMed]
- Hamley, I.W. Lipopeptides: From self-assembly to bioactivity. Chem. Commun. 2015, 51, 8574–8583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thongkongkaew, T.; Ding, W.; Bratovanov, E.; Oueis, E.; Garcia-Altares, M.; Zaburannyi, N.; Harmrolfs, K.; Zhang, Y.; Scherlach, K.; Müller, R.; et al. Two types of threonine-tagged lipopeptides synergize in host colonization by pathogenic Burkholderia species. ACS Chem. Biol. 2018, 13, 1370–1379. [Google Scholar] [CrossRef]
- Yoshimura, A.; Covington, B.C.; Gallant, E.; Zhang, C.; Li, A.; Seyedsayamdost, M.R. Unlocking cryptic metabolites with mass spectrometry-guided transposon mutant selection. ACS Chem. Biol. 2020, 15, 2766–2774. [Google Scholar] [CrossRef]
- Kang, Y.; Norris, M.H.; Wilcox, B.A.; Tuanyok, A.; Keim, P.S.; Hoang, T.T. Knockout and pullout recombineering for naturally transformable Burkholderia thailandensis and Burkholderia pseudomallei. Nat. Protoc. 2011, 6, 1085–1104. [Google Scholar] [CrossRef] [Green Version]
- Rausch, C.; Hoof, I.; Weber, T.; Wohlleben, W.; Huson, D.H. Phylogenetic analysis of condensation domains in NRPS sheds light on their functional evolution. BMC Evol. Biol. 2007, 7, 1–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, C.; Mannaa, M.; Kim, N.; Kim, J.; Choi, Y.; Kim, S.H.; Jung, B.; Lee, H.H.; Lee, J.; Seo, Y.S. Stress tolerance and virulence-related roles of lipopolysaccharide in Burkholderia glumae. Plant. Pathol. J. 2019, 35, 445–458. [Google Scholar] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No | 1 | 2 | |||
|---|---|---|---|---|---|
| δC | δH (J in Hz) | δC | δH (J in Hz) | ||
| β-OH-Dec | 1 | 172.1, C | 172.1, C | ||
| 2 | 42.6, CH2 | 2.37, d (6.4) | 42.6, CH2 | 2.36, d (6.5) | |
| 3 | 67.0, CH | 3.88, m | 67.0, CH | 3.87, m | |
| 4 | 37.3, CH2 | 1.30, m | 37.3, CH2 | 1.30, m | |
| 5 | 25.0, CH2 | 1.23, m a | 25.0, CH2 | 1.23, m a | |
| 6 | 29.1, CH2 | 1.23, m a | 29.1, CH2 | 1.23, m a | |
| 7 | 28.7, CH2 | 1.23, m a | 28.8, CH2 | 1.23, m a | |
| 8 | 31.3, CH2 | 1.23, m a | 31.3, CH2 | 1.23, m a | |
| 9 | 22.1, CH2 | 1.23, m a | 22.1, CH2 | 1.23, m a | |
| 10 | 14.0, CH3 | 0.86, t (6.7) | 14.0, CH3 | 0.86, t (7.1) | |
| Dhb | 1 | 166.9, C | |||
| 2 | 132.1, C | 166.9, C | |||
| 3 | 120.7, CH | 5.64, q (6.8) | 132.1, C | 5.64, q (6.8) | |
| 4 | 11.9, CH3 | 1.67, d (6.8) | 120.8, CH | 1.66, d (6.8) | |
| 2-NH | 166.9, C | 9.71, s | 12.0, CH3 | 9.71, s | |
| Pro | 1 | 171.2, C | 171.3, C | ||
| 2 | 61.0, CH | 4.20, t (7.6) | 61.0, CH | 4.20, t (7.2) | |
| 3a | 29.3, CH2 | 2.16, m | 29.3, CH2 | 2.16, m | |
| 3b | 1.75, m a | 1.74, m a | |||
| 4a | 24.9, CH2 | 1.84, m a | 24.9, CH2 | 1.84, m a | |
| 4b | 1.75, m a | 1.74, m a | |||
| 5a | 48.7, CH2 | 3.57, m | 48.7, CH2 | 3.57, m | |
| 5b | 171.2, C | 3.40, m | 171.3, C | 3.40, m | |
| Gln | 1 | 171.0, C | 171.0, C | ||
| 2 | 52.6, CH | 4.07, m a | 52.6, CH | 4.07, m a | |
| 3a | 27.1, CH2 | 2.04, m a | 27.1, CH2 | 2.04, m a | |
| 3b | 1.84, m a | 1.84, m a | |||
| 4 | 32.2, CH2 | 2.04, m a | 32.2, CH2 | 2.04, m a | |
| 5 | 174.2, C | 174.2, C | |||
| 2-NH | 171.0, C | 7.77, d (8.7) | 171.0, C | 7.77, d (8.5) | |
| Ala | 1 | 172.0, C | 172.0, C | ||
| 2 | 49.4, CH | 4.07, m a | 49.5, CH | 4.07, m a | |
| 3 | 17.7, CH3 | 1.19, d (7.3) | 17.8, CH3 | 1.19, d (7.3) | |
| 2-NH | 7.52, d (6.4) | 172.0, C | 7.52, d (6.3) | ||
| Val | 1 | 170.1, C | 170.1, C | ||
| 2 | 56.8, CH | 4.14, dd (6.9, 9.4) | 56.7, CH | 4.14, m a | |
| 3 | 30.8, CH | 1.66, m | 30.8, CH | 1.65, m | |
| 5 | 18.9, CH3 | 0.46, d (6.9) | 18.9, CH3 | 0.44, d (6.2) | |
| 6 | 17.4, CH3 | 0.44, d (6.9) | 17.4, CH3 | 0.43, d (6.2) | |
| 2-NH | 170.1, C | 7.21, d (7.9) | 170.1, C | 7.18, d (7.8) | |
| Phe | 1 | 169.6, C | 169.8, C | ||
| 2 | 51.8, CH | 4.72, m | 52.0, CH | 4.71, m | |
| 3a | 36.8, CH2 | 2.97, dd (4.2, 14.0) | 36.9, CH2 | 2.97, dd (3.4, 14.0) | |
| 3b | 2.72, dd (10.3, 14.0) | 2.72, dd (10.7, 14.0) | |||
| 4 | 137.6, C | 137.8, C | |||
| 5/9 | 129.3, CH | 7.27, d (7.2) | 129.3, CH | 7.27, d (7.3) | |
| 6/8 | 128.0, CH | 7.21, t (7.2) | 128.0, CH | 7.21, t (7.3) | |
| 7 | 126.2, CH | 7.14, t (7.2) | 126.2, CH | 7.13, t (7.3) | |
| 2-NH | 169.6, C | 8.45, d (8.5) | 8.44, d (8.5) | ||
| Pro | 1 | 173.2, C | 171.8, C | ||
| 2 | 58.6, CH | 4.27, dd (4.1, 8.8) | 59.3, CH | 4.54, dd (3.0, 8.3) | |
| 3a | 28.6, CH2 | 2.12, m | 28.8, CH2 | 2.04, m a | |
| 3b | 1.88, m a | 1.90, m a | |||
| 4 | 24.5, CH2 | 1.88, m a | 24.5, CH2 | 1.90, m a | |
| 5 | 46.4, CH2 | 3.64, m | 46.8, CH2 | 3.61, m | |
| Thr | 1 | 172.1, C | |||
| 2 | 57.5, CH | 4.21, m a | |||
| 3 | 66.4, CH | 4.14, m a | |||
| 4 | 20.4, CH3 | 1.08, d (6.4) | |||
| 2-NH | 172.1, C | 7.76, d (8.6) | |||
| No | 3 | 4 | |||
|---|---|---|---|---|---|
| δC | δH (J in Hz) | δC | δH (J in Hz) | ||
| β-OH-Dec | 1 | 172.3, C | 172.3, C | ||
| 2a | 42.6, CH2 | 2.37, dd (9.6, 14.4) | 42.7, CH2 | 2.37, dd (9.6, 14.4) | |
| 2b | 2.24, dd (2.3, 14.4) | 2.24, dd (2.3, 14.4) | |||
| 3 | 67.0, CH | 3.90, m | 67.0, CH | 3.90, m | |
| 4 | 37.4, CH2 | 1.32, m | 37.3, CH2 | 1.32, m | |
| 5 | 25.0, CH2 | 1.24, m a | 25.0, CH2 | 1.24, m a | |
| 6 | 29.2, CH2 | 1.24, m a | 29.1, CH2 | 1.24, m a | |
| 7 | 28.8, CH2 | 1.24, m a | 28.9, CH2 | 1.24, m a | |
| 8 | 31.3, CH2 | 1.24, m a | 31.4, CH2 | 1.24, m a | |
| 9 | 22.2, CH2 | 1.24, m a | 22.2, CH2 | 1.24, m a | |
| 10 | 14.0, CH3 | 0.86, t (6.3) | 14.0, CH3 | 0.85, t (6.3) | |
| Dhb | 1 | 167.1, C | 166.9, C | ||
| 2 | 132.1, C | 132.1, C | |||
| 3 | 121.3, CH | 5.67, q (6.8) | 120.6, CH | 5.62, q (6.8) | |
| 4 | 12.0, CH3 | 1.68, d (6.8) | 12.0, CH3 | 1.67, d (6.8) | |
| 2-NH | 9.74, s | 9.69, s | |||
| Pro | 1 | 171.2, C | 171.3, C | ||
| 2 | 61.2, CH | 4.20, m a | 61.0, CH | 4.20, m a | |
| 3a | 29.3, CH2 | 2.17, m | 29.3, CH2 | 2.17, m | |
| 3b | 1.76, m a | 1.76, m a | |||
| 4a | 25.0, CH2 | 1.83, m a | 24.9, CH2 | 1.83, m a | |
| 4b | 1.76, m a | 1.76, m a | |||
| 5a | 48.6, CH2 | 3.68, m | 48.7, CH2 | 3.68, m | |
| 5b | 3.38, m | 3.38, m | |||
| Gln | 1 | 170.9, C | 171.1, C | ||
| 2 | 52.6, CH | 4.06, m a | 52.7, CH | 4.06, m a | |
| 3a | 27.2, CH2 | 2.03, m a | 27.1, CH2 | 2.03, m a | |
| 3b | 1.83, m a | 1.83, m a | |||
| 4 | 32.3, CH2 | 2.03, m a | 32.2, CH2 | 2.03, m a | |
| 5 | 174.3, C | 174.2, C | |||
| 2-NH | 7.71, d (8.9) | 7.76, d (8.9) | |||
| Ala | 1 | 172.1, C | 172.0, C | ||
| 2 | 49.6, CH | 4.00, m a | 49.4, CH | 4.00, m a | |
| 3 | 17.6, CH3 | 1.19, d (7.2) | 17.8, CH3 | 1.19, d (7.2) | |
| 2-NH | 7.52, d (6.5) | 7.53, d (6.5) | |||
| Leu/Ile | 1 | 171.9, C | 170.4, C | ||
| 2 | 50.1, CH | 4.30, dd (4.5, 9.5) | 55.0, CH | 4.27, dd (6.1, 9.5) | |
| 3a | 41.4, CH2 | 1.01, m | 37.3, CH | 1.53, m | |
| 3b | 0.92, m | ||||
| 4a | 23.7, CH | 1.24, m a | 25.4, CH2 | 0.92, m a | |
| 4b | 0.73, m a | ||||
| 5 | 23.0, CH3 | 0.69, d (6.8) | 11.5, CH3 | 0.66, t (7.5) | |
| 6 | 21.5, CH3 | 0.68, d (6.8) | 13.9, CH3 | 0.39, d (6.7) | |
| 2-NH | 7.20, d a | 7.26, d a | |||
| Phe | 1 | 169.8, C | 169.9, C | ||
| 2 | 51.8, CH | 4.67, m | 52.1, CH | 4.67, m | |
| 3a | 37.0, CH2 | 2.96, m | 36.9, CH2 | 2.96, m | |
| 3b | 2.73, m | 2.73, m | |||
| 4 | 137.8, C | 137.8, C | |||
| 5/9 | 129.5, CH | 7.26, d (7.3) | 129.3, CH | 7.26, d (7.2) | |
| 6/8 | 127.9, CH | 7.20, t (7.3) | 128.0, CH | 7.20, t (7.2) | |
| 7 | 126.1, CH | 7.13, t (7.3) | 126.2, CH | 7.14, t (7.2) | |
| 2-NH | 8.42, d (8.5) | 8.42, d (8.5) | |||
| Pro | 1 | 171.8, C | 171.9, C | ||
| 2 | 59.3, CH | 4.54, dd (2.1, 7.8) | 59.3, CH | 4.54, dd (2.1, 7.8) | |
| 3a | 28.8, CH2 | 2.03, m a | 28.9, CH2 | 2.03, m a | |
| 3b | 1.92, m a | 1.92, m a | |||
| 4a | 2.03, m a | 24.5, CH2 | 2.03, m a | ||
| 4b | 24.5, CH2 | 1.92, m a | 1.92, m a | ||
| 5 | 46.8, CH2 | 3.60, m | 46.8, CH2 | 3.60, m | |
| Thr | 1 | 172.1, C | 172.1, C | ||
| 2 | 57.5, CH | 4.20, m a | 57.5, CH | 4.20, m a | |
| 3 | 66.5, CH | 4.14, m | 66.5, CH | 4.14, m | |
| 4 | 20.4, CH3 | 1.08, d (6.3) | 20.4, CH3 | 1.08, d (6.3) | |
| 2-NH | 7.76, d (8.6) | 7.77, d (8.6) | |||
| No | 5 | 8 | |||
|---|---|---|---|---|---|
| δC | δH (J in Hz) | δC | δH (J in Hz) | ||
| β-OH-Dec | 1 | 172.1, C | 171.7, C | ||
| 2a | 42.7, CH2 | 2.42, dd (4.1, 14.6) | 42.7, CH2 | 2.36, dd (8.7, 13.1) | |
| 2b | 2.37, dd (8.8, 14.6) | ||||
| 3 | 67.0, CH | 3.90, m | 67.1, CH | 3.89, m | |
| 4 | 37.3, CH2 | 1.32, m | 37.2, CH2 | 1.34, m | |
| 5 | 24.9, CH2 | 1.22, m a | 24.9, CH2 | 1.23, m a | |
| 6 | 29.0, CH2 | 1.22, m a | 29.0, CH2 | 1.23, m a | |
| 7 | 28.7, CH2 | 1.22, m a | 28.7, CH2 | 1.23, m a | |
| 8 | 31.3, CH2 | 1.22, m a | 31.3, CH2 | 1.23, m a | |
| 9 | 22.1, CH2 | 1.22, m a | 22.1, CH2 | 1.23, m a | |
| 10 | 13.9, CH3 | 0.85, t (6.7) | 14.0, CH3 | 0.85, t (7.1) | |
| Dhb | 1 | 166.9, C | 166.9, C | ||
| 2 | 132.0, C | 131.9, C | |||
| 3 | 120.9, CH | 5.64, q (6.9) | 120.5, CH | 5.61, q (7.1) | |
| 4 | 12.0, CH3 | 1.67, d (6.9) | 12.0, CH3 | 1.66, d (7.1) | |
| 2-NH | 9.69, s | 9.71, s | |||
| Pro | 1 | 171.3, C | 171.3, C | ||
| 2 | 60.9, CH | 4.21, t (7.5) | 60.8, CH | 4.20, m a | |
| 3a | 29.2, CH2 | 2.16, m | 29.3, CH2 | 2.16, m | |
| 3b | 1.74, m a | 1.75, m a | |||
| 4a | 24.9, CH2 | 1.83, m a | 24.8, CH2 | 1.84, m a | |
| 4b | 1.74, m a | 1.75, m a | |||
| 5a | 48.7, CH2 | 3.67, m | 48.8, CH2 | 3.63, m | |
| 5b | 3.41, m | 3.42, m | |||
| Gln | 1 | 171.0, C | 171.2, C | ||
| 2 | 52.6, CH | 4.09, m a | 52.7, CH | 4.08, m | |
| 3a | 27.1, CH2 | 2.06, m a | 26.9, CH2 | 2.07, m a | |
| 3b | 1.84, m a | 1.84, m a | |||
| 4 | 32.2, CH2 | 2.06, m a | 32.0, CH2 | 2.07, m a | |
| 5 | 174.1, C | 174.0, C | |||
| 2-NH | 7.77, d (8.8) | 7.76, d (8.5) | |||
| Ala | 1 | 172.1, C | 171.7, C | ||
| 2 | 49.5, CH | 4.09, m a | 49.3, CH | 4.03, m | |
| 3 | 17.7, CH3 | 1.21, d (7.4) | 17.4, CH3 | 1.18, d (7.2) | |
| 2-NH | 7.55, d (6.4) | 7.57, d (6.3) | |||
| Val/Ala | 1 | 170.3, C | 171.6, C | ||
| 2 | 56.9, CH | 4.14, dd (6.9, 9.3) | 47.5, CH | 4.24, m | |
| 3 | 30.6, CH | 1.72, m | 18.3, CH3 | 0.93, d (7.1) | |
| 4 | 18.9, CH3 | 0.56, d (6.7) | |||
| 5 | 17.3, CH3 | 0.51, d (6.7) | |||
| 2-NH | 170.3, C | 7.25, d (9.2) | 7.52, d (8.0) | ||
| Phe | 1 | 172.8, C | 172.8, C | ||
| 2 | 53.4, CH | 4.40, m | 53.2, CH | 4.42, m | |
| 3a | 36.8, CH2 | 3.06, dd (4.5, 13.8) | 37.0, CH2 | 3.06, dd (4.9, 13.8) | |
| 3b | 2.83, dd (10.4, 13.8) | 2.83, dd (9.6, 13.8) | |||
| 4 | 137.7, C | 7.22, m a | 137.4, C | ||
| 5/9 | 129.1, CH | 7.22, m a | 129.2, CH | 7.24, m a | |
| 6/8 | 128.0, CH | 7.15, t (6.7) | 128.1, CH | 7.18, m a | |
| 7 | 126.3, CH | 8.22, d (8.3) | 126.3, CH | 7.18, m a | |
| 2-NH | 8.09, d (8.3) | ||||
| No | 6 | 7 | |||
|---|---|---|---|---|---|
| δC | δH (J in Hz) | δC | δH (J in Hz) | ||
| β-OH-Dec | 1 | 172.0, C | 171.9, C | ||
| 2a | 42.6, CH2 | 2.38, dd (9.7, 14.5) | 42.7, CH2 | 2.39, dd (9.7, 14.5) | |
| 2b | 2.33, dd (3.4, 14.5) | 2.38, dd (4.8, 14.5) | |||
| 3 | 67.1, CH | 3.90, m | 67.0, CH | 3.90, m | |
| 4 | 37.3, CH2 | 1.33, m | 37.3, CH2 | 1.33, m | |
| 5 | 24.9, CH2 | 1.23, m a | 24.9, CH2 | 1.23, m a | |
| 6 | 29.1, CH2 | 1.23, m a | 29.1, CH2 | 1.23, m a | |
| 7 | 28.7, CH2 | 1.23, m a | 28.7, CH2 | 1.23, m a | |
| 8 | 31.3, CH2 | 1.23, m a | 31.3, CH2 | 1.23, m a | |
| 9 | 22.1, CH2 | 1.23, m a | 22.1, CH2 | 1.23, m a | |
| 10 | 14.0, CH3 | 0.85, t (6.7) | 14.0, CH3 | 0.85, t (6.7) | |
| Dhb | 1 | 167.1, C | 166.9, C | ||
| 2 | 131.9, C | 131.9, C | |||
| 3 | 121.2, CH | 5.65, q (6.9) | 120.7, CH | 5.62, q (7.0) | |
| 4 | 12.0, CH3 | 1.68, d (6.9) | 12.0, CH3 | 1.66, d (7.0) | |
| 2-NH | 9.74, s | 9.68, s | |||
| Pro | 1 | 171.3, C | 171.3, C | ||
| 2 | 61.1, CH | 4.20, t (7.6) | 60.9, CH | 4.20, t (7.6) | |
| 3a | 29.3, CH2 | 2.17, m | 29.3, CH2 | 2.17, m | |
| 3b | 1.75, m a | 1.75, m a | |||
| 4a | 24.9, CH2 | 1.83, m a | 24.8, CH2 | 1.83, m a | |
| 4b | 1.75, m a | 1.75, m a | |||
| 5a | 48.6, CH2 | 3.68, m | 48.7, CH2 | 3.68, m | |
| 5b | 3.42, m a | 171.3, C | 3.42, m a | ||
| Gln | 1 | 171.0, C | 171.1, C | ||
| 2 | 52.6, CH | 4.09, m a | 52.7, CH | 4.09, m a | |
| 3a | 27.1, CH2 | 2.06, m a | 27.1, CH2 | 2.06, m a | |
| 3b | 1.83, m a | 1.83, m a | |||
| 4 | 32.3, CH2 | 2.06, m a | 32.2, CH2 | 2.06, m a | |
| 5 | 174.2, C | 174.1, C | |||
| 2-NH | 7.71, d (8.8) | 7.77, d (8.7) | |||
| Ala | 1 | 172.2, C | 172.2, C | ||
| 2 | 49.6, CH | 4.02, m | 49.4, CH | 4.09, m a | |
| 3 | 17.5, CH3 | 1.20, d (7.2) | 17.7, CH3 | 1.20, d (7.2) | |
| 2-NH | 7.55, d (5.9) | 7.54, d (5.8) | |||
| Leu/Ile | 1 | 171.5, C | 170.6, C | ||
| 2 | 50.1, CH | 4.30, dd (4.8, 9.5) | 55.1, CH | 4.27, dd (6.1, 9.4) | |
| 3 | 41.2, CH2 | 1.09, m | 37.0, CH | 1.52, m | |
| 4a | 23.7, CH | 1.32, m a | 25.5, CH2 | 1.02, m a | |
| 4b | 0.84, m a | ||||
| 5 | 23.0, CH3 | 0.71, d (6.5) | 11.4, CH3 | 0.70, t (7.7) | |
| 6 | 21.5, CH3 | 0.71, d (6.5) | 13.9, CH3 | 0.46, d (6.8) | |
| 2-NH | 7.33, d (8.8) | 7.26, d (9.4) | |||
| Phe | 1 | 172.9, C | 172.9, C | ||
| 2 | 53.3, CH | 4.38, m | 53.4, CH | 4.42, m | |
| 3a | 37.0, CH2 | 3.06, dd (3.8, 13.5) | 36.8, CH2 | 3.06, dd (4.0, 13.8) | |
| 3b | 2.81, dd (4.4, 13.5) | 2.83, dd (4.8, 13.8) | |||
| 4 | 137.6, C | 137.7, C | |||
| 5/9 | 129.2, CH | 7.21, m a | 129.1, CH | 7.21, m a | |
| 6/8 | 128.0, CH | 7.21, m a | 128.1, CH | 7.21, m a | |
| 7 | 126.3, CH | 7.16, t (6.8) | 126.3, CH | 7.16, t (6.8) | |
| 2-NH | 8.15, d (8.4) | 8.18, d (8.4) | |||
| No | 9 | 10 | |||
|---|---|---|---|---|---|
| δC | δH (J in Hz) | δC | δH (J in Hz) | ||
| Octanoate acid | 1 | 172.1, C | 172.1, C | ||
| 2 | 34.8, CH2 | 2.23, m | 35.6, CH2 | 2.17, t (5.4) | |
| 3 | 24.7, CH2 | 1.49, m a | 25.3, CH2 | 1.49, m | |
| 4 | 28.6, CH2 | 1.23, m a | 28.9, CH2 | 1.22, m a | |
| 5 | 28.7, CH2 | 1.23, m a | 29.1, CH2 | 1.22, m a | |
| 6 | 31.2, CH2 | 1.19, m a | 31.6, CH2 | 1.22, m a | |
| 7 | 22.1, CH2 | 1.23, m a | 22.5, CH2 | 1.22, m a | |
| 8 | 13.9, CH3 | 0.79, t (6.9) | 14.4, CH3 | 0.83, t (6.0) | |
| Dhb | 1 | 164.4, C | 164.2, C | ||
| 2 | 131.7, C | 132.4, C | |||
| 3 | 118.1, CH | 5.52, q (7.2) | 118.9, CH | 5.62, q (6.0) | |
| 4 | 12.7, CH3 | 1.73, d (7.2) | 13.3, CH3 | 1.74, d (6.0) | |
| 2-NH | 9.83, s | 9.18, s | |||
| Dhb | 1 | 164.2, C | 164.0, C | ||
| 2 | 129.3, C | 130.6, C | |||
| 3 | 127.3, CH | 5.71, q (7.3) | 125.4, CH | 5.89, q (5.8) | |
| 4 | 13.3, CH3 | 1.79, d (7.3) | 13.8, CH3 | 1.82, d (5.8) | |
| 2-NH | 9.85, s | 9.51, s | |||
| Tyr | 1 | 171.4, C | 172.6, C | 4.17, m | |
| 2 | 56.0, CH | 4.26, m | 55.6, CH | 2.93, d (10.5) | |
| 3a | 35.5, CH2 | 3.03, dd (5.2, 14.0), | 36.5, CH2 | ||
| 3b | 2.92, dd (10.3, 14.0) | ||||
| 4 | 128.2, C | 129.1, C | 6.98, d (8.0) | ||
| 5/9 | 129.8, CH | 7.03, d (8.4) | 130.6, CH | 6.58, d (8.0) | |
| 6/8 | 115.0, CH | 6.62, d (8.4) | 115.2, CH | ||
| 7 | 155.9, C | 156.1, C | 7.62, d (6.0) | ||
| 2-NH | 8.19, d (6.8) | ||||
| 7-OH | 9.23, s | ||||
| Leu | 1 | 171.3, C | |||
| 2 | 52.3, CH | 4.31, m | |||
| 3a | 39.9, CH2 | 1.63, m a | |||
| 3b | 1.49, m a | ||||
| 4a | 24.0, CH | 1.63, m a | |||
| 4b | |||||
| 5 | 23.1, CH3 | 0.83, d (6.3) | |||
| 6 | 21.4, CH3 | 0.80, d (6.3) | |||
| 2-NH | 8.00, d (7.6) | ||||
| PABA | 1 | 166.1, C | |||
| 2 | 128.6, C | ||||
| 3/7 | 128.1, CH | 7.86, d (8.7) | |||
| 4/6 | 118.6, CH | 7.76, d (8.7) | |||
| 5 | 141.7, C | ||||
| 5-NH | 9.83, s | ||||
| Thr | 1 | 172.3, C | |||
| 2 | 58.7, CH | 4.40, dd (3.5, 8.3) | |||
| 3 | 66.6, CH | 4.19, m | |||
| 4 | 20.5, CH3 | 1.13, d (6.4) | |||
| 2-NH | 7.94, d (8.3) | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, H.; Sun, T.; Bai, X.; Yang, J.; Yan, F.; Yu, L.; Tu, Q.; Li, A.; Tang, Y.; Zhang, Y.; et al. Genomics-Driven Activation of Silent Biosynthetic Gene Clusters in Burkholderia gladioli by Screening Recombineering System. Molecules 2021, 26, 700. https://doi.org/10.3390/molecules26030700
Chen H, Sun T, Bai X, Yang J, Yan F, Yu L, Tu Q, Li A, Tang Y, Zhang Y, et al. Genomics-Driven Activation of Silent Biosynthetic Gene Clusters in Burkholderia gladioli by Screening Recombineering System. Molecules. 2021; 26(3):700. https://doi.org/10.3390/molecules26030700
Chicago/Turabian StyleChen, Hanna, Tao Sun, Xianping Bai, Jie Yang, Fu Yan, Lei Yu, Qiang Tu, Aiying Li, Yajie Tang, Youming Zhang, and et al. 2021. "Genomics-Driven Activation of Silent Biosynthetic Gene Clusters in Burkholderia gladioli by Screening Recombineering System" Molecules 26, no. 3: 700. https://doi.org/10.3390/molecules26030700