
Pleurotus spp. Mycelia Enriched in Magnesium and Zinc Salts as a Potential Functional Food

 , , , ,
, , , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Mushroom Materials
2.1.1. Initial Mycelial Cultures
2.1.2. Experimental Mycelial Cultures
2.1.3. Fruiting Bodies
2.2. Reagents
2.3. Preparation of Artificial Digestive Juices
2.4. Extraction of Metals and Organic Compounds into Artificial Digestive Juices
2.5. Analysis of Organic Compounds—Preparation of the Extract
2.6. Analytical Tools Applied
2.6.1. Analysis of Mg and Zn
2.6.2. Determination of Chloride and Sulfate Ions
2.6.3. Determination of Phenylalanine and Phenolic Acids
2.6.4. Analysis of Indole Compounds
2.6.5. Determination of the Total Phenol Content
2.7. Determination of Antioxidant Activity Using the DPPH Method
2.8. Statistical Analysis
3. Results and Discussion
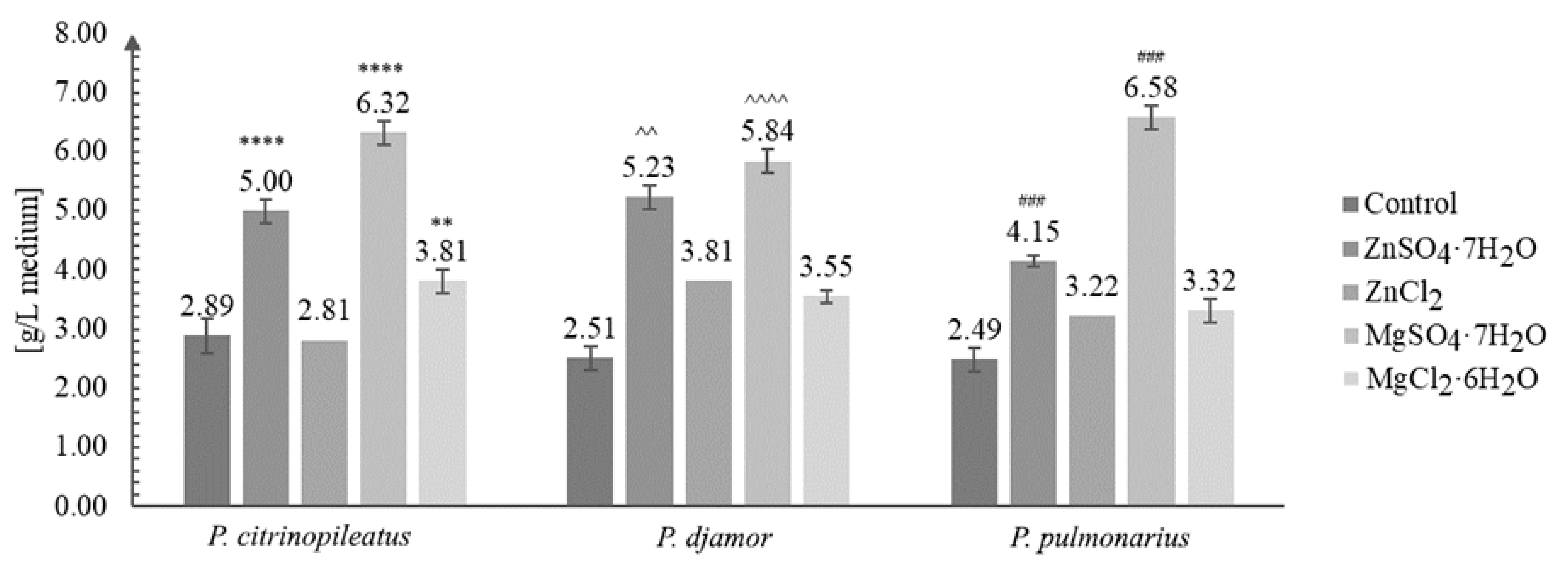
3.1. Biomass Growth Analysis
3.2. Analysis of Metals and Anions Content
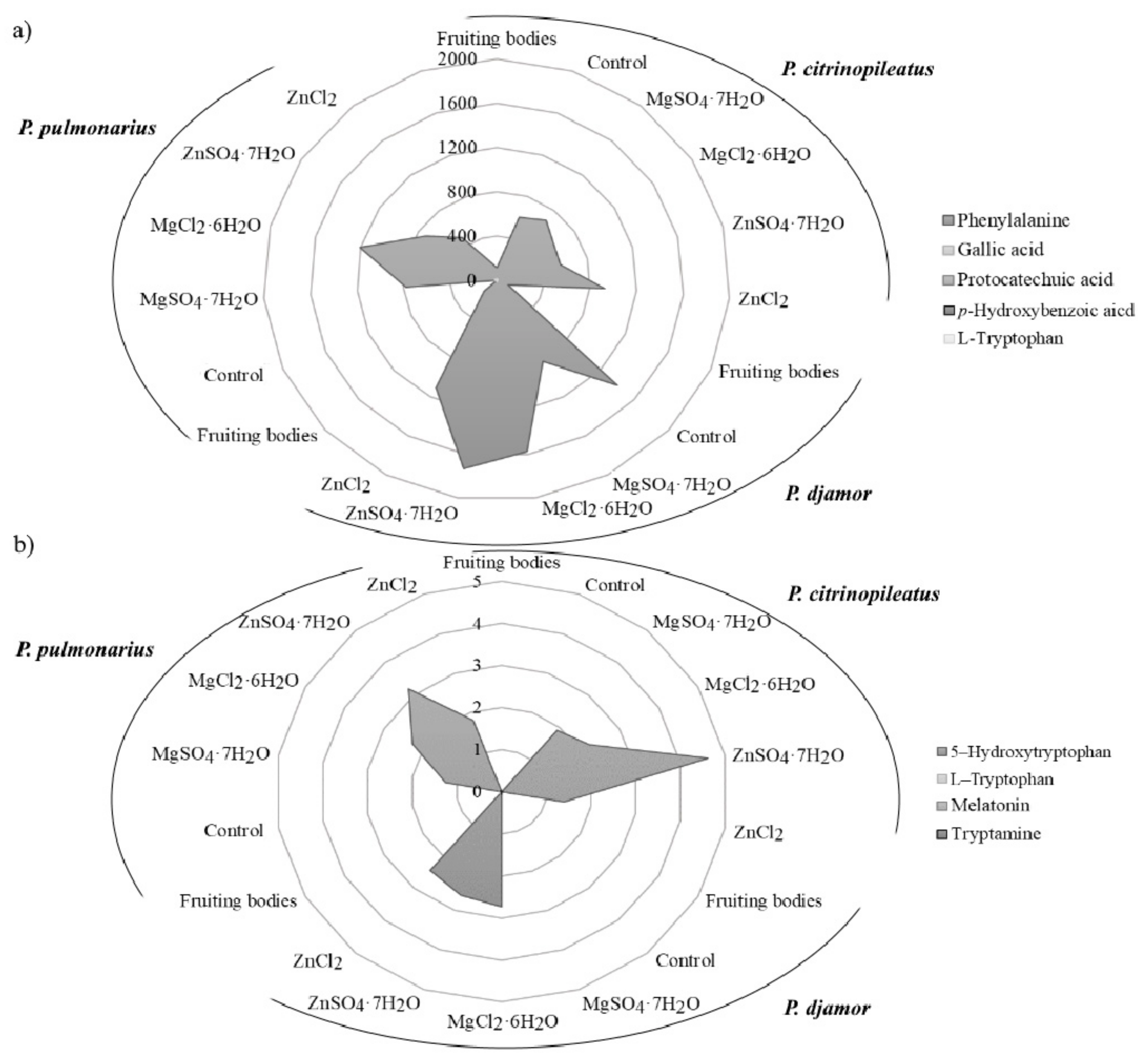
3.3. Determination of Organic Compounds
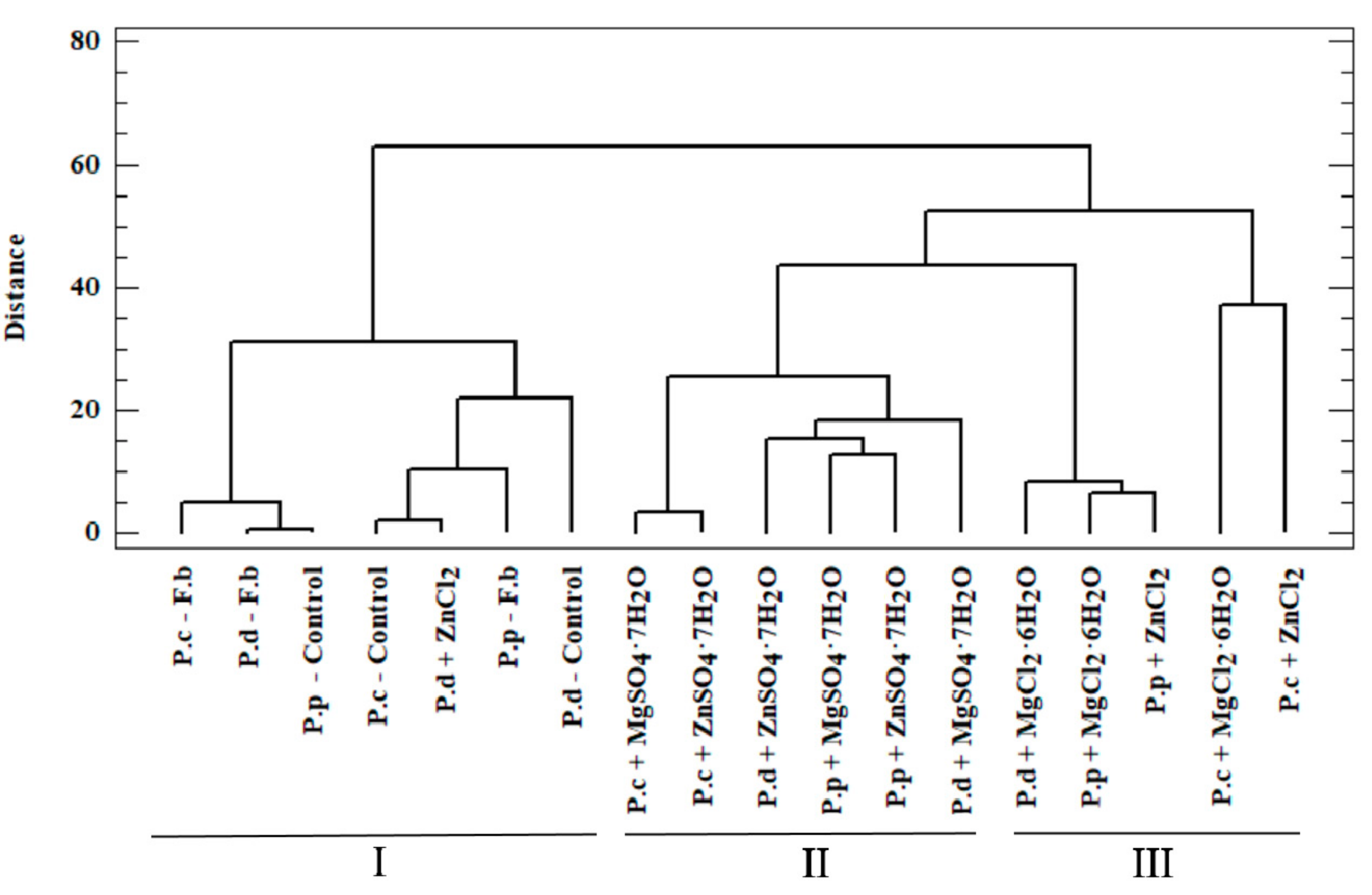
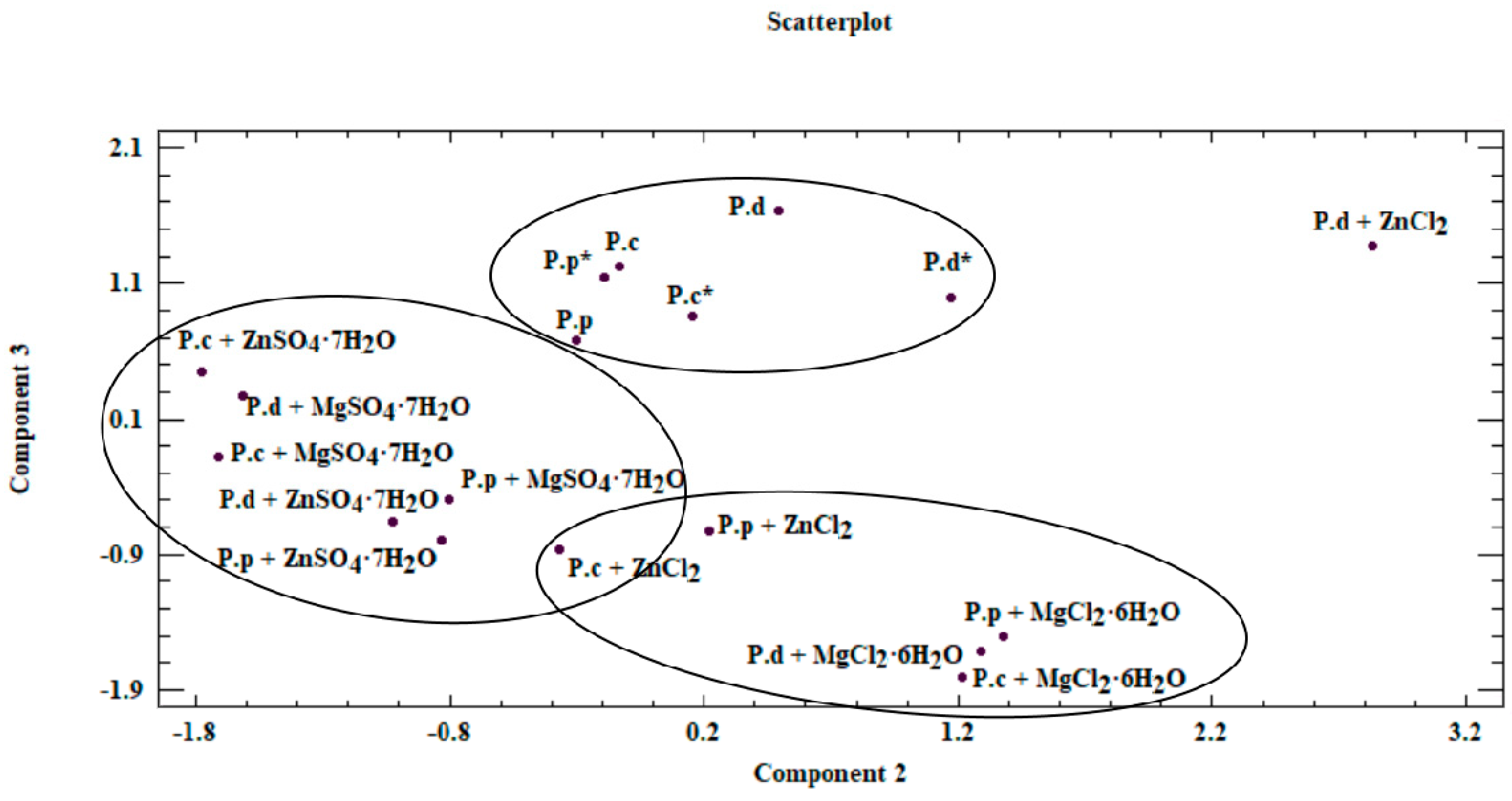
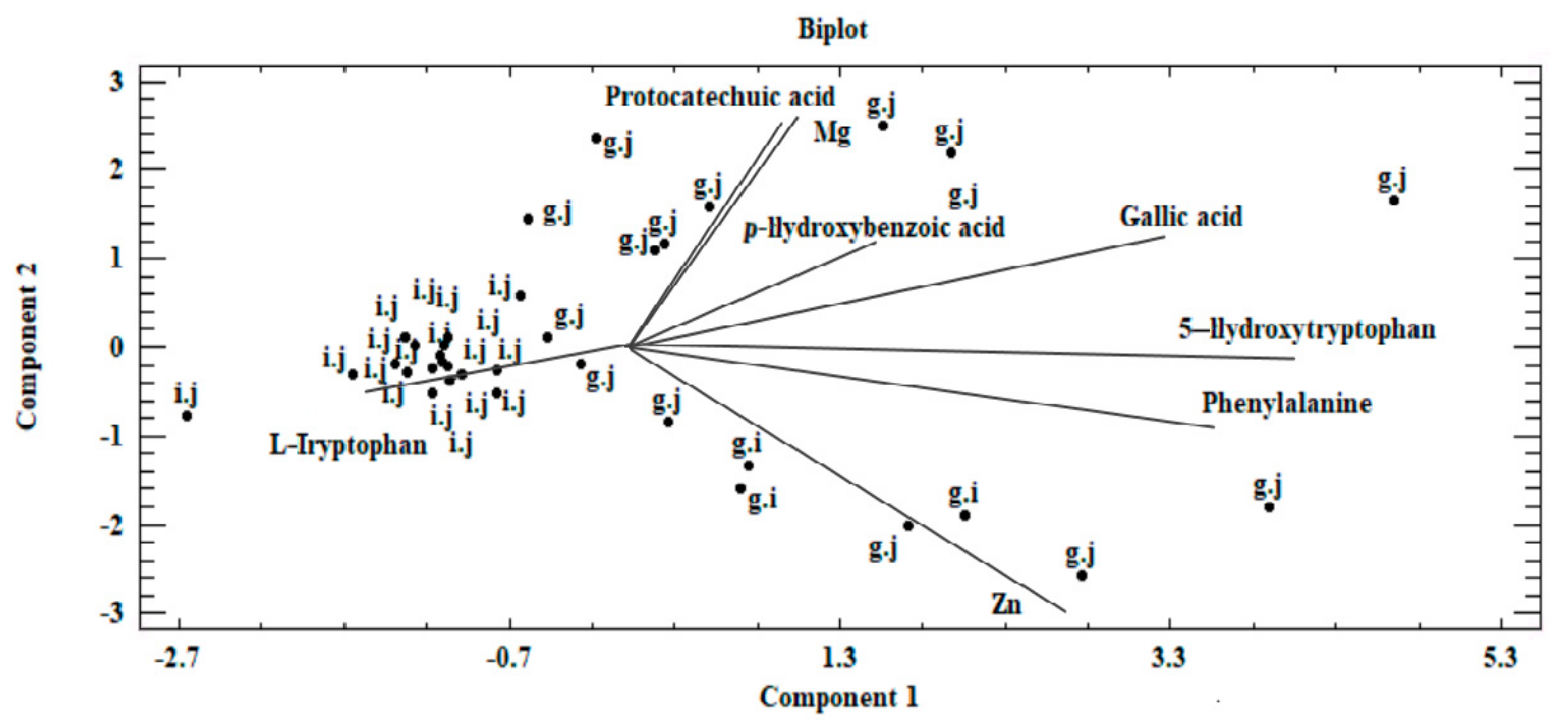
3.4. Chemometric Analysis of Organic Compounds
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Samples Availability
References
- Chang, S.T.; Wasser, S.P. Current and Future Research Trends in Agricultural and Biomedical Applications of Medicinal Mushrooms and Mushroom Products (Review). Int. J. Med. Mushrooms 2018, 20, 1121–1133. [Google Scholar] [CrossRef]
- Muszyńska, B.; Sułkowska-Ziaja, K.; Ekiert, H. Phenolic acids in selected edible Basidiomycota species: Armillaria mellea, Boletus badius, Boletus edulis, Cantharellus cibarius, Lactarius deliciosus and Pleurotus ostreatus. Acta Sci. Pol. Hortorum Cultus 2012, 12, 107–116. [Google Scholar]
- Wasser, S. Medicinal mushroom science: Current perspectives, advances, evidences, and challenges. Biomed. J. 2014, 37, 345–356. [Google Scholar] [CrossRef]
- Rajewska, J.; Bałasińska, B. Biologically active compounds of edible mushrooms and their benefi cial impact on health. Postepy Hig. Med. Dosw. 2004, 58, 352–357. [Google Scholar]
- Chaturvedi, V.K.; Agarwal, S.; Gupta, K.K.; Ramteke, P.W.; Singh, M.P. Medicinal mushroom: Boon for therapeutic applications. 3 Biotech 2018, 8, 334. [Google Scholar] [CrossRef]
- Muszyńska, B.; Grzywacz-Kisielewska, A.; Kała, K.; Gdula-Argasińska, J. Anti-inflammatory properties of edible mushrooms: A review. Food Chem. 2018, 243, 373–381. [Google Scholar] [CrossRef]
- Golak-Siwulska, I.; Kałużewicz, A.; Spiżewski, T.; Siwulski, M.; Sobieralski, K. Bioactive compounds and medicinal properties of Oyster mushrooms (Pleurotus sp.). Folia Hortic. 2018, 30, 191–201. [Google Scholar] [CrossRef] [Green Version]
- Muszyńska, B.; Kała, K.; Włodarczyk, A.; Krakowska, A.; Ostachowicz, B.; Gdula-Argasińska, J.; Suchocki, P. Lentinula edodes as a Source of Bioelements Released into Artificial Digestive Juices and Potential Anti-inflammatory Material. Biol. Trace Elem. Res. 2020, 194, 603–613. [Google Scholar] [CrossRef] [Green Version]
- Patel, Y.; Naraian, R.; Singh, V.K. Medicinal Properties of Pleurotus Species (Oyster Mushroom). WJFPB. 2012, 3, 1–12. [Google Scholar] [CrossRef]
- Royse, D.J.; Baars, J.; Tan, Q. Current Overview of Mushroom Production in the World. In Edible and Medicinal Mushrooms: Technology and Applicaions; Diego, C.Z., Pardo-Giménez, A., Eds.; John Wiley and Sons Ltd.: Hoboken, NJ, USA, 2017; pp. 5–13. [Google Scholar] [CrossRef]
- Yamanaka, K. Mushroom cultivation in Japan. Mushroom Production Bulletin Number 2011; World Society for Mushroom Biology and Mushroom Products: Kyoto, Japan, 4 January 2011; pp. 455–459. [Google Scholar]
- Gapiński, M.; Woźniak, W.; Ziobra, M. Oyster Mushroom. Technology of Cultivation and Processing; State Agricultural and Forest Publishing House: Warsaw, Poland, 2010; pp. 12–15. [Google Scholar]
- Bellettini, M.B.; Fiorda, F.A.; Maieves, H.A.; Teixeira, G.L.; Ávila, S.; Hornung, P.S.; Júnior, A.M.; Ribani, R.H. Factors affecting mushroom Pleurotus spp. Saudi J. Biol. Sci. 2019, 26, 633–646. [Google Scholar] [CrossRef]
- Krakowska, A.; Zięba, P.; Włodarczyk, A.; Kała, K.; Sułkowska-Ziaja, K.; Bernaś, E.; Sękara, A.; Ostachowicz, B.; Muszyńska, B. Selected edible medicinal mushrooms from Pleurotus genus as an answer for human civilization diseases. Food Chem. 2020, 327, 127084. [Google Scholar] [CrossRef]
- Zięba, P.; Kała, K.; Włodarczyk, A.; Szewczyk, A.; Kunicki, E.; Sękara, A.; Muszyńska, B. Selenium and Zinc Biofortification of Pleurotus eryngii Mycelium and Fruiting Bodies as a Tool for Controlling Their Biological Activity. Molecules 2020, 25, 889. [Google Scholar] [CrossRef] [Green Version]
- Friese, W. Mineral composition of macrofungi. Z Unters Lebensm Forsch 1929, 57, 604–613. [Google Scholar]
- Kała, K.; Krakowska, A.; Gdula-Argasińska, J.; Opoka, W.; Muszyńska, B. Assessing the Bioavailability of Zinc and Indole Compounds from Mycelial Cultures of the Bay Mushroom Imleria badia (Agaricomycetes) Using In Vitro Models. Int. J. Med. Mushrooms 2019, 21, 343–352. [Google Scholar] [CrossRef]
- Campos, J.A.; Tejera, N.A.; Sánchez, C.J. Substrate role in the accumulation of heavy metals in sporocarps of wild fungi. BioMetals 2009, 22, 835–841. [Google Scholar] [CrossRef]
- Falandysz, J.; Kojta, A.K.; Jarzyńska, G.; Drewnowska, M.; Dryżałowska, A.; Wydmańska, D.; Kowalewska, I.; Wacko, A.; Szlosowska, M.; Kannan, K.; et al. Mercury in bay bolete (Xerocomus badius): Bioconcentration by fungus and assessment of element intake by humans eating fruiting bodies. Food Addit. Contam. Part A 2012, 29, 951–961. [Google Scholar] [CrossRef] [Green Version]
- Mleczek, M.; Siwulski, M.; Stuper-Szablewska, K.; Rissmann, I.; Sobieralski, K.; Goliński, P. Accumulation of elements by edible mushroom species: Part I. Problem of trace element toxicity in mushrooms. J. Environ. Sci. Heal. Part B 2012, 48, 69–81. [Google Scholar] [CrossRef]
- Krakowska, A.; Reczyński, W.; Muszyńska, B. Optimization of the Liquid Culture Medium Composition to Obtain the Mycelium of Agaricus bisporus Rich in Essential Minerals. Biol. Trace Elem. Res. 2016, 173, 231–240. [Google Scholar] [CrossRef]
- Reczyński, W.; Muszyńska, B.; Opoka, W.; Smalec, A.; Sułkowska-Ziaja, K.; Malec, M. Comparative Study of Metals Accumulation in Cultured In Vitro Mycelium and Naturally Grown Fruiting Bodies of Boletus badius and Cantharellus cibarius. Biol. Trace Elem. Res. 2013, 153, 355–362. [Google Scholar] [CrossRef] [Green Version]
- Bagherani, N.; Smoller, B.R. An overview of zinc and its importance in dermatology- Part I: Importance and function of zinc in human beings. Glob. Dermatol. 2016, 3, 330–336. [Google Scholar] [CrossRef]
- Corbo, M.D.; Lam, J. Zinc deficiency and its management in the pediatric population: A literature review and proposed etiologic classification. J. Am. Acad. Dermatol. 2013, 69, 616–624.e1. [Google Scholar] [CrossRef]
- Jahnen-Dechent, W.; Ketteler, M. Magnesium basics. Clin. Kidney J. 2012, 5, i3–i14. [Google Scholar] [CrossRef] [Green Version]
- Schwalfenberg, G.K.; Genuis, S.J. The Importance of Magnesium in Clinical Healthcare. Scientifica 2017, 2017, 1–14. [Google Scholar] [CrossRef]
- Sikorski, Z.E. Food Chemistry; WNT: Warsaw, Poland, 2012. [Google Scholar]
- Oddoux, L. Research on Secondary Mycelia of Homobasidia in Pure Culture: Morphology, Cytology, Dietary Requirements; Trevoux: Lyon, France, 1957. [Google Scholar]
- Arvidson, K.; Johansson, E.G. Galvanic currents between dental alloys in vitro. Scand. J. Dent. Res. 1985, 93, 467–473. [Google Scholar]
- Neumann, M.; Goderska, K.; Grajek, K.; Grajek, W. In vitro gastrointestinal models for nutrient bioavailability studies. Food Sci. Technol. Qual. 2006, 1, 30–45. [Google Scholar]
- Polish Pharmakopeia PTFarm. Wydanie X, PZWL; Polish Pharmakopeia PTFarm: Warszawa, Poland, 2014. [Google Scholar]
- Opoka, W.; Muszyńska, B.; Rojowski, J.; Rumian, J. Gastroel–2014. Poland Patent Application P 417238, 2016. [Google Scholar]
- Confortin, F.G.; Marchetto, R.; Bettin, F.; Camassola, M.; Salvador, M.; Dillon, A.J.P. Production of Pleurotus sajor-caju strain PS-2001 biomass in submerged culture. J. Ind. Microbiol. Biotechnol. 2008, 35, 1149–1155. [Google Scholar] [CrossRef]
- Burns, P.; Yeo, P.; Keshavarz, T.; Roller, S.; Evans, C. Physiological studies of exopolysaccharide production from the basidiomycete Pleurotus sp. florida. Enzym. Microb. Technol. 1994, 16, 566–572. [Google Scholar] [CrossRef]
- Rosado, F.R.; Germano, S.; Carbonero, E.R.; Da Costa, S.M.G.; Iacomini, M.; Kemmelmeier, C. Biomass and exopolysaccharide production in submerged cultures of Pleurotus ostreatoroseus Sing. and Pleurotus ostreatus “florida” (Jack.: Fr.) Kummer. J. Basic Microbiol. 2003, 43, 230–237. [Google Scholar] [CrossRef]
- Gąsecka, M.; Mleczek, M.; Siwulski, M.; Niedzielski, P. Phenolic composition and antioxidant properties of Pleurotus ostreatus and Pleurotus eryngii enriched with selenium and zinc. Eur. Food Res. Technol. 2016, 242, 723–732. [Google Scholar] [CrossRef] [Green Version]
- Poursaeid, N.; Azadbakht, A.; Balali, G.R. Improvement of Zinc Bioaccumulation and Biomass Yield in the Mycelia and Fruiting Bodies of Pleurotus florida Cultured on Liquid Media. Appl. Biochem. Biotechnol. 2015, 175, 3387–3396. [Google Scholar] [CrossRef]
- Muszyńska, B.; Szacawa, E.; Bederska-Łojewska, D.; Dudek, K.; Pomierny, B.; Włodarczyk, A.; Kała, K.; Lazur, J.; Suchocki, P.; Budziszewska, B.; et al. Preliminary study on Se-enriched Lentinula edodes mycelium as a proposal of new feed additive in selenium deficiency. PLoS ONE 2020, 15, e0233456. [Google Scholar] [CrossRef]
- Araújo, A.C.M.F.; Araújo, W.M.C.; Marquez, U.M.L.; Akutsu, R.D.C.C.A.; Nakano, E.Y.; Baumgartner, M.R.; Patterson, M.; Rahman, S.; Peters, V.; Morava, E.; et al. Table of Phenylalanine Content of Foods: Comparative Analysis of Data Compiled in Food Composition Tables. JIMD Reports 2016, 34, 87–96. [Google Scholar] [CrossRef] [Green Version]
- Muszyńska, B.; Krakowska, A.; Sułkowska-Ziaja, K.; Szewczyk, A.; Reczyński, W.; Opoka, W.; Muszyńska, B. Kinetics of extracted bioactive components from mushrooms in artificial digestive juices. Int. J. Food Prop. 2016, 20, 1796–1817. [Google Scholar] [CrossRef] [Green Version]
- Miller, J.N.; Miller, J.C. Statistics and Chemometrics for Analytical Chemistry; Prentice Hall: London, UK, 1999. [Google Scholar]
- Mazerski, J. Fundamentals of Chemometry; Publisher Politechniki Gdańskiej: Gdańsk, Poland, 2000. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Sample Weight (g) |
|---|---|
| Saliva (pH = 6.7) | |
| KH2PO4 (25 mmol/L) | 0.34 |
| Na2HPO4 (24 mmol/L) | 0.34 |
| KHCO3 (150 mmol/L) | 1.50 |
| MgCl2 (1.5 mmol/L) | 0.01 |
| C6H8O7 (25 mmol/L) | 0.03 |
| CaCl2 (15 mmol/L) | 0.17 |
| Gastric juice (pH = 2) | |
| NaCl | 2.00 |
| Pepsin | 3.20 |
| HCl | 0.12 |
| Intestinal juice (pH = 8) | |
| Pancreatic extract | 0.02 |
| Bile salt | 0.12 |
| NaHCO3 | 8.40 |
| Species | Sample | Addition | Mg (mg/100 g d.w.) | Zn (mg/100 g d.w.) | Cl− (mg/100 g d.w.) | SO42− (mg/100 g d.w.) | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | |||
| P.c | Fruiting bodies | 103.4 ± 9.2 b | 28.1 ± 1.6 b | 12.2 ± 1.2 a | 2.1 ± 0.4 a | 667 ± 5 b | 464 ± 2 c | 404 ± 0 a | 196 ± 1 b | |
| Mycelium | 51.2 ± 4.7 a | 20.2 ± 0.5 a | 10.5 ± 0.1 a | 1.1 ± 0.1 a | 160 ± 4 a | 78 ± 2 a | 416 ± 1 b | 93 ± 1 a | ||
| Mycelium | MgSO4·7H2O | 906.2 ± 25.1 d | 88.4 ± 4.1 d | - | - | - | - | 1302 ± 3 c | 90 ± 6 a | |
| Mycelium | MgCl2·6H2O | 824.9 ± 17.7 c | 76.7 ± 2.0 c | - | - | 867 ± 9 c | 285 ± 3 b | - | - | |
| Mycelium | ZnSO4·7H2O | - | - | 166.3 ± 3.4 c | 11.7 ± 1.5 c | - | - | 408 ± 2 a | 199 ± 1 b | |
| Mycelium | ZnCl2 | - | - | 111.3 ± 0.8 b | 6.4 ± 0.6 b | 1067 ± 8 d | 464 ± 6 c | - | - | |
| P.d | Fruiting bodies | 71.1 ± 5.7 a | 8.2 ± 0.8 a | 10.3 ± 0.1 a | 1.8 ± 0.1 a | 200 ± 2 b | 70 ± 1 a | 226 ± 3 a | 121 ± 2 b | |
| Mycelium | 59.2 ± 4.7 a | 10.4 ± 1.3 a | 13.9 ± 0.3 a | 0.9 ± 0.1 a | 400 ± 2 c | 133 ± 1 b | 626 ± 1 b | 262 ± 1 d | ||
| Mycelium | MgSO4·7H2O | 842.8 ± 8.5 c | 155.9 ± 5.8 c | - | - | - | - | 2462 ± 5 d | 91 ± 3 a | |
| Mycelium | MgCl2·6H2O | 697.8 ± 12.5 b | 117.0 ± 3.4 b | - | - | 667 ± 3 d | 305 ± 18 c | - | - | |
| Mycelium | ZnSO4·7H2O | - | - | 145.8 ± 6.7 c | 9.6 ± 1.7 b | - | - | 678 ± 1 c | 253 ± 1 c | |
| Mycelium | ZnCl2 | - | - | 128.1 ± 4.2 b | 11.6 ± 2.3 b | 133 ± 8 a | 68 ± 13 a | - | - | |
| P.p | Fruiting bodies | 59.1 ± 4.8 a | 10.4 ± 1.3 a | 6.9 ± 0.6 a | 1.4 ± 0.2 a | 600 ± 1 c | 410 ± 2 c | 287 ± 0 a | 21 ± 1 a | |
| Mycelium | 41.1 ± 4.2 a | 8.3 ± 0.7 a | 15.3 ± 0.7 a | 2.0 ± 0.3 a | 1000 ± 2 d | 413 ± 2 c | 449 ± 0 b | 68 ± 1 b | ||
| Mycelium | MgSO4·7H2O | 1280.4 ± 16.5 c | 82.3 ± 3.9 b | - | - | - | - | 3636 ± 3 d | 399 ± 3 d | |
| Mycelium | MgCl2·6H2O | 526.9 ± 19.7 b | 131.6 ± 5.1 c | - | - | 400 ± 4 a | 133 ± 3 a | - | - | |
| Mycelium | ZnSO4·7H2O | - | - | 121.8 ± 3.6 c | 22.3 ± 0.2 c | - | - | 818 ± 2 c | 385 ± 1 c | |
| Mycelium | ZnCl2 | - | - | 98.8 ± 5.5 b | 16.4 ± 1.1 b | 533 ± 5 b | 385 ± 3 b | - | - | |
| Species | Sample | Addition | Phenylalanine and Phenolic Acids [mg/100 g d.w.] | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Phenylalanine | Gallic Acid | Protocatechuic Acid | p-Hydroxybenzoic Acid | |||||||||
| Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | |||||
| P.c | Fruiting bodies | 108.3 ± 13.9 a | n.d. | n.d | n.d | 0.14 ± 0.01 a | n.d. | 0.14 ± 0.01 cd | n.d. | |||
| Mycelium | 395.2 ± 57.9 d | n.d. | 4.49 ± 0.75 d | n.d | 0.13 ± 0.02 a | n.d. | 0.13 ± 0.01 bc | n.d. | ||||
| Mycelium | MgSO4·7H2O | 271.0 ± 1.7 c | 37.2 ± 1.9 d | 0.78 ± 0.01 b | 0.35 ± 0.01 b | 0.15 ± 0.02 a | 0.35 ± 0.01 b | 0.15 ± 0.01 d | 0.22 ± 0.01 c | |||
| Mycelium | MgCl2·6H2O | 168.7 ± 2.9 b | 24.2 ± 0.4 b | 0.25 ± 0.11 a | n.d | n.d. | n.d. | 0.07 ± 0.01 a | 0.02 ± 0.00 b | |||
| Mycelium | ZnSO4·7H2O | 448.0 ± 22.1 d | 26.7 ± 0.5 c | 2.43 ± 0.08 c | 0.15 ± 0.05 a | n.d. | n.d. | 0.12 ± 0.01 b | 0.01 ± 0.00 a | |||
| Mycelium | ZnCl2 | 127.8 ± 6.1 ab | 12.8 ± 1.2 a | n.d | 0.12 ± 0.00 a | n.d. | 0.16 ± 0.01 b | 0.22 ± 0.01 e | n.d. | |||
| P.d | Fruiting bodies | 24.9 ± 1.7 a | n.d. | n.d. | n.d. | n.d. | 0.03 ± 0.01 a | 0.03 ± 0.01 a | n.d. | |||
| Mycelium | 657.5 ± 13.5 e | n.d. | 2.10 ± 0.21 b | n.d. | n.d. | n.d. | 0.23 ± 0.01 d | n.d. | ||||
| Mycelium | MgSO4·7H2O | 563.9 ± 1.6 d | 32.7 ± 0.9 a | n.d. | n.d. | n.d. | n.d. | 0.12 ± 0.03 c | n.d. | |||
| Mycelium | MgCl2·6H2O | 262.0 ± 8.7 b | 33.2 ± 10.4 a | 1.13 ± 0.31 a | n.d. | n.d. | 0.14 ± 0.02 b | 0.79 ± 0.01 e | 0.14 ± 0.00 b | |||
| Mycelium | ZnSO4·7H2O | 721.6 ± 1.1f | 217.5 ± 32.7 b | n.d. | n.d. | n.d. | 0.03 ± 0.02 a | 0.06 ± 0.01 b | 0.03 ± 0.00 a | |||
| Mycelium | ZnCl2 | 547.5 ± 1.1 c | 42.8 ± 0.5 a | n.d. | n.d. | n.d. | n.d | 0.23 ± 0.01 d | n.d. | |||
| P.p | Fruiting bodies | 137.7 ± 3.3 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |||
| Mycelium | 49 ± 5.1 a | n.d | 0.26 ± 0.01 b | n.d. | n.d. | n.d. | n.d. | n.d. | ||||
| Mycelium | MgSO4·7H2O | 274.4 ± 9.6 d | 23.8 ± 2.5 b | 0.15 ± 0.02 a | n.d. | n.d. | n.d. | n.d. | n.d. | |||
| Mycelium | MgCl2·6H2O | 35.7 ± 0.1 a | 3.7 ± 0.1 a | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |||
| Mycelium | ZnSO4·7H2O | 320.1 ± 0.8 e | 55.3 ± 3.8 d | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |||
| Mycelium | ZnCl2 | 183.7 ± 34.3 c | 36.8 ± 1.7 c | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |||
| Species | Sample | Addition | Indole Compounds [mg/100 g d.w.] | |||||||||
| 5-Hydroxytryptophan | L-Tryptophan | Melatonin | Tryptamine | Serotonin | ||||||||
| Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | Gastric Juice | Intestinal Juice | |||
| P.c | Fruiting bodies | 0.47 ± 0.01 a | 0.17 ± 0.01 bc | 3.15 ± 0.01 a | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | 1.63 ± 0.07 d | 0.19 ± 0.01 c | 10.61 ± 0.50 c | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
| Mycelium | MgSO4·7H2O | 0.77 ± 0.06 b | 0.13 ± 0.08 | n.d. | 2.87 ± 0.01 a | * | * | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | MgCl2·6H2O | 0.39 ± 0.11 a | 0.11 ± 0.01 abc | 4.07 ± 0.10 b | 2.99 ± 0.01 b | * | n.d. | * | n.d. | n.d. | n.d. | |
| Mycelium | ZnSO4·7H2O | 1.06 ± 0.06 c | 0.05 ± 0.03 a | n.d. | 3.19 ± 0.01 c | * | * | n.d. | n.d. | 0.04 ± 0.0 | n.d. | |
| Mycelium | ZnCl2 | 0.45 ± 0.02 a | 0.07 ± 0.04 ab | 3.74 ± 0.01 b | 3.69 ± 0.01 d | * | * | n.d. | n.d. | n.d | n.d. | |
| P.d | Fruiting bodies | 0.29 ± 0.04 a | n.d. | 4.06 ± 0.01 d | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | n.d. | n.d. | 2.74 ± 0.01 a | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
| Mycelium | MgSO4·7H2OO | 0.29 ± 0.01 a | 0.11 ± 0.01 a | 4.24 ± 0.03 e | 3.53 ± 0.01 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | MgCl2·6H2O | 0.51 ± 0.01 b | 0.11 ± 0.00 a | 5.01 ± 0.01 f | 3.63 ± 0.01 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | ZnSO4·7H2O | 0.83 ± 0.04 d | 0.29 ± 0.02 b | 3.33 ± 0.01 b | 3.31 ± 0.02 a | * | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | ZnCl2 | 0.57 ± 0.01 c | 0.12 ± 0.01 a | 3.64 ± 0.16 c | 4.10 ± 0.15 c | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| P.p | Fruiting bodies | 0.02 ± 0.01 a | n.d. | 10.61 ± 0.62 d | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | 0.31 ± 0.15 b | 0.38 ± 0.02 d | 3.68 ± 0.02 a | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | ||
| Mycelium | MgSO4·7H2O | 0.41 ± 0.08 b | 0.13 ± 0.01 b | 6.41 ± 0.02 c | 4.60 ± 0.02 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | MgCl2·6H2O | 0.30 ± 0.01 b | 0.05 ± 0.03 a | 4.72 ± 0.08 b | 3.78 ± 0.01 a | n.d. | n.d. | n.d. | * | n.d. | n.d. | |
| Mycelium | ZnSO4·7H2O | 0.93 ± 0.03 d | 0.22 ± 0.03 c | 3.24 ± 0.01 a | 5.33 ± 0.77 c | n.d. | * | n.d. | n.d. | n.d. | n.d. | |
| Mycelium | ZnCl2 | 0.60 ± 0.05 d | 0.15 ± 0.01 c | 5.02 ± 0.01 a | 4.54 ± 0.05 c | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. | |
| PC1 | PC2 | PC3 | |
|---|---|---|---|
| Phenylalanine | 0.46508 | −0.49099 | −0.07265 |
| Gallic acid | 0.53583 | 0.04218 | 0.32777 |
| Protocatechuic acid | 0.23264 | 0.83861 | 0.16821 |
| p-Hydroxybenzoic acid | 0.22460 | 0.19685 | −0.58975 |
| L-Tryptophan | −0.23189 | −0.08289 | 0.68890 |
| 5-Hydroxytryptophan | 0.58157 | −0.09077 | 0.19126 |
| Species | Sample | Addition | TP | DPPH | TP | DPPH | TP | DPPH |
|---|---|---|---|---|---|---|---|---|
| Dry Material | Dry Material | Gastric Juice | Gastric Juice | Intestinal Juice | Intestinal Juice | |||
| P.c | Fruiting bodies | 297 ± 24 e | 44 ± 0.5 d | 4.33 ± 0.03 b | 8.9 ± 0.41 b | 1.3 ± 0.01 c | 1.18 ± 0.21 c | |
| Mycelium | 243 ± 6 d | 14.4 ± 0.2 c | 8.39 ± 0.12 f | 33.86 ± 0.22 f | 2.28 ± 0.01 e | 0.51 ± 0.14 ab | ||
| Mycelium | MgSO4·7H2O | 98 ± 3 a | 7 ± 0.2 a | 5.77 ± 0.01 d | 20.81 ± 0.55 d | 1.07 ± 0.02 b | 0.54 ± 0.06 ab | |
| Mycelium | MgCl2·6H2O | 229 ± 2 d | 12.9 ± 0.6 b | 4.49 ± 0.03 c | 10.82 ± 0.68 c | 1.09 ± 0.02 b | 2.38 ± 0.17 d | |
| Mycelium | ZnSO4·7H2O | 135 ± 1 b | 4.7 ± 0.2 a | 6.91 ± 0.02 e | 31.84 ± 0.07 e | 1.4 ± 0.03 d | 0.85 ± 0.01 bc | |
| Mycelium | ZnCl2 | 192 ± 12 c | 5.6 ± 0.8 a | 4.13 ± 0.01 a | 7.25 ± 0.22 a | 0.13 ± 0.01 a | 0.25 ± 0.09 a | |
| P.d | Fruiting bodies | 161 ± 1 c | 10.3 ± 0.4 b | 3.73 ± 0.03 a | 5.51 ± 0.36 a | 1.8 ± 0.01 d | 9.15 ± 0.14 d | |
| Mycelium | 193 ± 8 d | 11.5 ± 0.5 b | 7.67 ± 0.01 f | 25.16 ± 0.43 e | 2.2 ± 0.02 f | 10.11 ± 1.15 d | ||
| Mycelium | MgSO4·7H2O | 105 ± 6 a | 6 ± 0.2 a | 4.41 ± 0.06 b | 9.52 ± 0.21 b | 0.88 ± 0.01 a | 2.04 ± 0.4 c | |
| Mycelium | MgCl2·6H2O | 186 ± 3 d | 11.3 ± 2.2 b | 5.18 ± 0.02 c | 10.66 ± 0.36 c | 1.35 ± 0.01 c | 1.01 ± 0.16 b | |
| Mycelium | ZnSO4·7H2O | 142 ± 3 b | 9 ± 1.1 b | 6.22 ± 0.02 e | 20.34 ± 0.41 d | 1.95 ± 0.01 e | 0.29 ± 0.01 a | |
| Mycelium | ZnCl2 | 230 ± 1 e | 9.5 ± 0.3 b | 5.83 ± 0.01 d | 9.93 ± 0.08 bc | 1.01 ± 0.02 b | 0.29 ± 0.01 a | |
| P.p | Fruiting bodies | 92 ± 13 a | 7.8 ± 0.5 ab | 8.34 ± 0.03 d | 43.43 ± 0.07 f | 2.13 ± 0.01 e | 5.82 ± 0.07 d | |
| Mycelium | 259 ± 5 c | 19 ± 0.7 e | 4.88 ± 0.01 b | 4.38 ± 0.66 b | 2.32 ± 0.01 f | 3.2 ± 0.36 c | ||
| Mycelium | MgSO4·7H2O | 141 ± 3 b | 3.9 ± 0.3 a | 4.73 ± 0.17 b | 7.27 ± 0.43 c | 0.97 ± 0.01 b | 5.16 ± 0.39 d | |
| Mycelium | MgCl2·6H2O | 253 ± 5 c | 14 ± 0.7 c | 3.89 ± 0.03 a | 3.22 ± 0.21 a | 0.91 ± 0.01 a | 1.24 ± 0.3 b | |
| Mycelium | ZnSO4·7H2O | 125 ± 0 b | 10 ± 0.2 bc | 6.33 ± 0.06 c | 18.36 ± 0.22 e | 1.71 ± 0.01 d | 3.35 ± 0.07 c | |
| Mycelium | ZnCl2 | 104 ± 2 a | 3.9 ± 0.2 a | 6.45 ± 0.04 c | 14.01 ± 0.13 d | 1.49 ± 0.02 c | 0.29 ± 0.01 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Włodarczyk, A.; Krakowska, A.; Sułkowska-Ziaja, K.; Suchanek, M.; Zięba, P.; Opoka, W.; Muszyńska, B. Pleurotus spp. Mycelia Enriched in Magnesium and Zinc Salts as a Potential Functional Food. Molecules 2021, 26, 162. https://doi.org/10.3390/molecules26010162
Włodarczyk A, Krakowska A, Sułkowska-Ziaja K, Suchanek M, Zięba P, Opoka W, Muszyńska B. Pleurotus spp. Mycelia Enriched in Magnesium and Zinc Salts as a Potential Functional Food. Molecules. 2021; 26(1):162. https://doi.org/10.3390/molecules26010162
Chicago/Turabian StyleWłodarczyk, Anna, Agata Krakowska, Katarzyna Sułkowska-Ziaja, Małgorzata Suchanek, Piotr Zięba, Włodzimierz Opoka, and Bożena Muszyńska. 2021. "Pleurotus spp. Mycelia Enriched in Magnesium and Zinc Salts as a Potential Functional Food" Molecules 26, no. 1: 162. https://doi.org/10.3390/molecules26010162