
Antitumor Effects of N-Butylidenephthalide Encapsulated in Lipopolyplexs in Colorectal Cancer Cells

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of BP/LPPC on Cell Viability in Colorectal Cancer Cells
2.2. LPPC Encapsulation Stabilized BP Activity for Cytotoxicity of Colorectal Cancer Cells
2.3. LPPC Encapsulation Increased Cell Uptake of BP through Induction of Clathrin-Mediated Endocytosis
2.4. BP/LPPC Induce Cell Cycle Arrest and Cell Apoptosis in Colorectal Cancer Cells
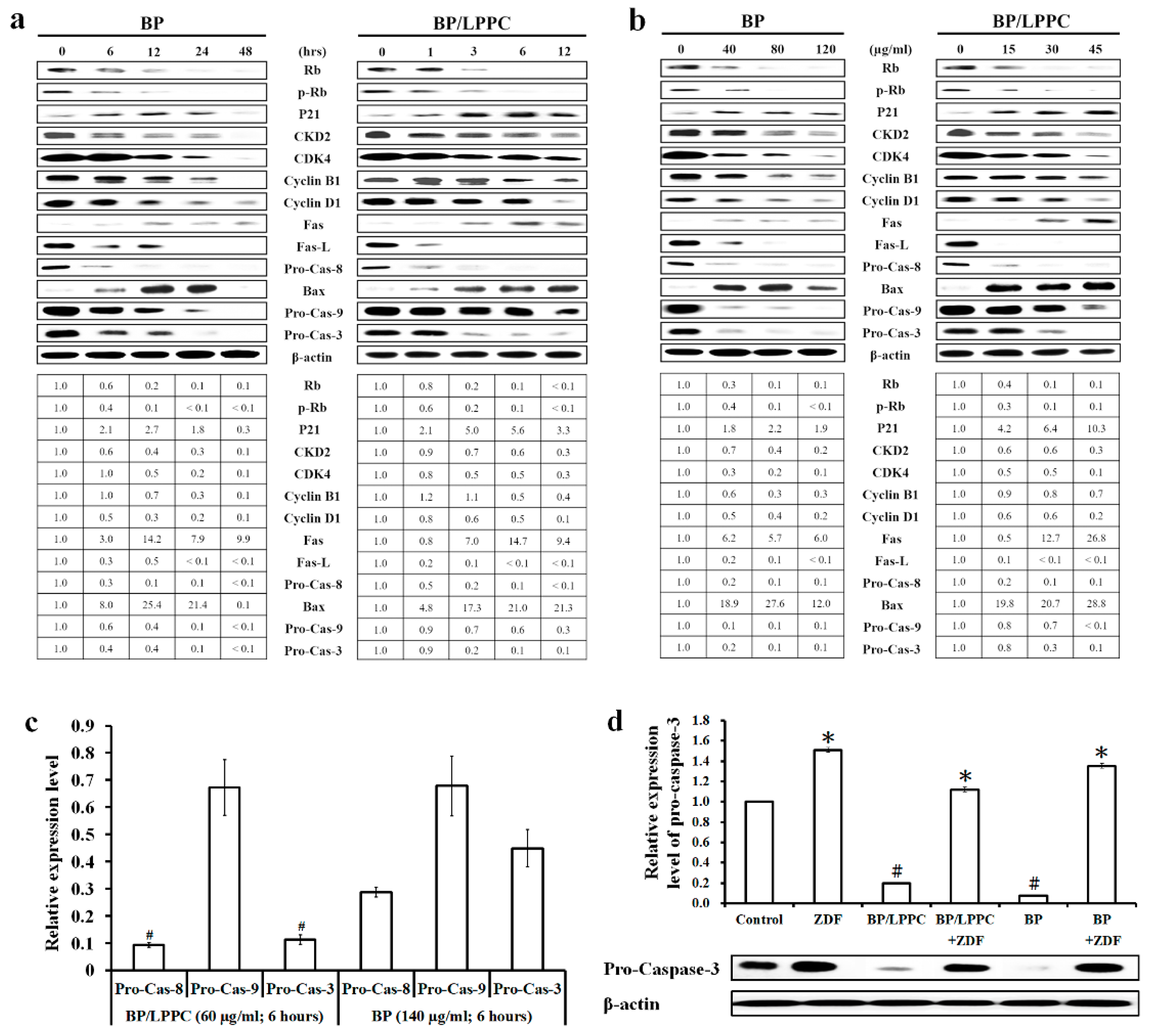
2.5. BP/LPPC Regulated Cell Cycle and Apoptosis Associated Protein Expression
2.6. Combination of BP/LPPC and 5-FU Synergistically Inhibits the Growth of Colorectal Cancer Cells
3. Materials and Methods
3.1. Cell Culture and Reagents
3.2. Analysis of the Cytotoxicity
3.3. Detection of BP Activity
3.4. The Cellular Uptake of BP/LPPC
3.5. Analysis of Endocytosis
3.6. Cell Cycle Analysis
3.7. TUNEL Assay
3.8. Western Blotting
3.9. Analysis of Caspase-3 Activity
3.10. Synergistic Effects of BP Combined with a Clinical Drug
3.11. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hook, I.L. Danggui to Angelica sinensis root: Are potential benefits to European women lost in translation? A review. J. Ethnopharmacol. 2014, 152, 1–13. [Google Scholar] [CrossRef]
- Yeh, J.C.; Cindrova-Davies, T.; Belleri, M.; Morbidelli, L.; Miller, N.; Cho, C.W.; Chan, K.; Wang, Y.T.; Luo, G.A.; Ziche, M.; et al. The natural compound n-butylidenephthalide derived from the volatile oil of Radix Angelica sinensis inhibits angiogenesis in vitro and in vivo. Angiogenesis 2011, 14, 187–197. [Google Scholar] [CrossRef]
- Liu, W.S.; Lin, P.C.; Chang, L.F.; Harn, H.J.; Shiuan, D.; Chiou, T.W.; Jeng, J.R. Inhibitory effect of n-butylidenephthalide on neointimal hyperplasia in balloon injured rat carotid artery. Phytother. Res. PTR 2011, 25, 1494–1502. [Google Scholar] [CrossRef] [PubMed]
- Fu, R.H.; Hran, H.J.; Chu, C.L.; Huang, C.M.; Liu, S.P.; Wang, Y.C.; Lin, Y.H.; Shyu, W.C.; Lin, S.Z. Lipopolysaccharide-stimulated activation of murine DC2.4 cells is attenuated by n-butylidenephthalide through suppression of the NF-kappaB pathway. Biotechnol. Lett. 2011, 33, 903–910. [Google Scholar] [CrossRef] [PubMed]
- Kan, W.L.; Cho, C.H.; Rudd, J.A.; Lin, G. Study of the anti-proliferative effects and synergy of phthalides from Angelica sinensis on colon cancer cells. J. Ethnopharmacol. 2008, 120, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Tsai, N.M.; Chen, Y.L.; Lee, C.C.; Lin, P.C.; Cheng, Y.L.; Chang, W.L.; Lin, S.Z.; Harn, H.J. The natural compound n-butylidenephthalide derived from Angelica sinensis inhibits malignant brain tumor growth in vitro and in vivo. J. Neurochem. 2006, 99, 1251–1262. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.L.; Jian, M.H.; Lin, C.C.; Kang, J.C.; Chen, S.P.; Lin, P.C.; Hung, P.J.; Chen, J.R.; Chang, W.L.; Lin, S.Z.; et al. The induction of orphan nuclear receptor Nur77 expression by n-butylenephthalide as pharmaceuticals on hepatocellular carcinoma cell therapy. Mol. Pharmacol. 2008, 74, 1046–1058. [Google Scholar] [CrossRef] [Green Version]
- Wei, C.W.; Lin, C.C.; Yu, Y.L.; Lin, C.Y.; Lin, P.C.; Wu, M.T.; Chen, C.J.; Chang, W.; Lin, S.Z.; Chen, Y.L.; et al. n-Butylidenephthalide induced apoptosis in the A549 human lung adenocarcinoma cell line by coupled down-regulation of AP-2alpha and telomerase activity. Acta Pharmacol. Sin. 2009, 30, 1297–1306. [Google Scholar] [CrossRef]
- Schinkovitz, A.; Pro, S.M.; Main, M.; Chen, S.N.; Jaki, B.U.; Lankin, D.C.; Pauli, G.F. Dynamic nature of the ligustilide complex. J. Nat. Prod. 2008, 71, 1604–1611. [Google Scholar] [CrossRef] [Green Version]
- Yan, R.; Ko, N.L.; Li, S.L.; Tam, Y.K.; Lin, G. Pharmacokinetics and metabolism of ligustilide, a major bioactive component in Rhizoma Chuanxiong, in the rat. Drug Metab. Dispos. Biol. Fate Chem. 2008, 36, 400–408. [Google Scholar] [CrossRef] [Green Version]
- Deng, S.; Chen, S.N.; Yao, P.; Nikolic, D.; van Breemen, R.B.; Bolton, J.L.; Fong, H.H.; Farnsworth, N.R.; Pauli, G.F. Serotonergic activity-guided phytochemical investigation of the roots of Angelica sinensis. J. Nat. Prod. 2006, 69, 536–641. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, S.; Mimura, Y.; Naitoh, T.; Kimura, I.; Kimura, M. Chemical structure-activity of cnidium rhizome-derived phthalides for the competence inhibition of proliferation in primary cultures of mouse aorta smooth muscle cells. Jpn. J. Pharmacol. 1993, 63, 353–359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatesan, P.; Puvvada, N.; Dash, R.; Prashanth Kumar, B.N.; Sarkar, D.; Azab, B.; Pathak, A.; Kundu, S.C.; Fisher, P.B.; Mandal, M. The potential of celecoxib-loaded hydroxyapatite-chitosan nanocomposite for the treatment of colon cancer. Biomaterials 2011, 32, 3794–3806. [Google Scholar] [CrossRef] [PubMed]
- Fortina, P.; Kricka, L.J.; Graves, D.J.; Park, J.; Hyslop, T.; Tam, F.; Halas, N.; Surrey, S.; Waldman, S.A. Applications of nanoparticles to diagnostics and therapeutics in colorectal cancer. Trends Biotechnol. 2007, 25, 145–152. [Google Scholar] [CrossRef] [PubMed]
- Jemal, A.; Bray, F.; Center, M.M.; Ferlay, J.; Ward, E.; Forman, D. Global cancer statistics. CA A Cancer J. Clin. 2011, 61, 69–90. [Google Scholar] [CrossRef] [Green Version]
- O’Connell, J.B.; Maggard, M.A.; Ko, C.Y. Colon cancer survival rates with the new American Joint Committee on Cancer sixth edition staging. J. Nat. Cancer Inst. 2004, 96, 1420–1425. [Google Scholar] [CrossRef]
- Smitha, K.T.; Anitha, A.; Furuike, T.; Tamura, H.; Nair, S.V.; Jayakumar, R. In vitro evaluation of paclitaxel loaded amorphous chitin nanoparticles for colon cancer drug delivery. Coll. Surf. B Biointerfaces 2013, 104, 245–253. [Google Scholar] [CrossRef] [Green Version]
- Ashwanikumar, N.; Kumar, N.A.; Nair, S.A.; Kumar, G.S.V. Dual drug delivery of 5-fluorouracil (5-FU) and methotrexate (MTX) through random copolymeric nanomicelles of PLGA and polyethylenimine demonstrating enhanced cell uptake and cytotoxicity. Coll. Surf. B Biointerfaces 2014, 122, 520–528. [Google Scholar] [CrossRef]
- Tan, M.L.; Choong, P.F.; Dass, C.R. Recent developments in liposomes, microparticles and nanoparticles for protein and peptide drug delivery. Peptides 2010, 31, 184–193. [Google Scholar] [CrossRef]
- Malam, Y.; Loizidou, M.; Seifalian, A.M. Liposomes and nanoparticles: Nanosized vehicles for drug delivery in cancer. Trends Pharmacol. Sci. 2009, 30, 592–599. [Google Scholar] [CrossRef]
- Schroeder, A.; Kost, J.; Barenholz, Y. Ultrasound, liposomes, and drug delivery: Principles for using ultrasound to control the release of drugs from liposomes. Chem. Phys. Lipids 2009, 162, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Allen, T.M.; Cullis, P.R. Drug delivery systems: Entering the mainstream. Science 2004, 303, 1818–1822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Celia, C.; Cosco, D.; Paolino, D.; Fresta, M. Gemcitabine-loaded innovative nanocarriers vs GEMZAR: Biodistribution, pharmacokinetic features and in vivo antitumor activity. Expert Opin. Drug Deliv. 2011, 8, 1609–1629. [Google Scholar] [CrossRef] [PubMed]
- Koshkaryev, A.; Piroyan, A.; Torchilin, V.P. Increased apoptosis in cancer cells in vitro and in vivo by ceramides in transferrin-modified liposomes. Cancer Biol. Ther. 2012, 13, 50–60. [Google Scholar] [CrossRef] [Green Version]
- Ueno, Y.; Sonoda, S.; Suzuki, R.; Yokouchi, M.; Kawasoe, Y.; Tachibana, K.; Maruyama, K.; Sakamoto, T.; Komiya, S. Combination of ultrasound and bubble liposome enhance the effect of doxorubicin and inhibit murine osteosarcoma growth. Cancer Biol. Ther. 2011, 12, 270–277. [Google Scholar] [CrossRef] [Green Version]
- Jiang, Y.; DiVittore, N.A.; Kaiser, J.M.; Shanmugavelandy, S.S.; Fritz, J.L.; Heakal, Y.; Tagaram, H.R.; Cheng, H.; Cabot, M.C.; Staveley-O’Carroll, K.F.; et al. Combinatorial therapies improve the therapeutic efficacy of nanoliposomal ceramide for pancreatic cancer. Cancer Biol. Ther. 2011, 12, 574–585. [Google Scholar] [CrossRef] [Green Version]
- Deng, H.; Jiang, Q.; Yang, Y.; Zhang, S.; Ma, Y.; Xie, G.; Chen, X.; Qian, Z.; Wen, Y.; Li, J.; et al. Intravenous liposomal delivery of the short hairpin RNAs against Plk1 controls the growth of established human hepatocellular carcinoma. Cancer Biol. Ther. 2014, 11, 401–409. [Google Scholar] [CrossRef] [Green Version]
- Lonez, C.; Vandenbranden, M.; Ruysschaert, J.M. Cationic liposomal lipids: From gene carriers to cell signaling. Progress Lipid Res. 2008, 47, 340–347. [Google Scholar] [CrossRef]
- Liu, Y.K.; Lin, Y.L.; Chen, C.H.; Lin, C.M.; Ma, K.L.; Chou, F.H.; Tsai, J.S.; Lin, H.Y.; Chen, F.R.; Cheng, T.L.; et al. A unique and potent protein binding nature of liposome containing polyethylenimine and polyethylene glycol: A nondisplaceable property. Biotechnol. Bioeng. 2011, 108, 1318–1327. [Google Scholar] [CrossRef]
- Zuhorn, I.S.; Hoekstra, D. On the mechanism of cationic amphiphile-mediated transfection. To fuse or not to fuse: Is that the question? J. Membrane Biol. 2002, 189, 167–179. [Google Scholar] [CrossRef]
- Rawat, A.; Vaidya, B.; Khatri, K.; Goyal, A.K.; Gupta, P.N.; Mahor, S.; Paliwal, R.; Rai, S.; Vyas, S.P. Targeted intracellular delivery of therapeutics: An overview. Die Pharmazie 2007, 62, 643–658. [Google Scholar] [PubMed]
- Chen, C.H.; Lin, Y.L.; Liu, Y.K.; He, P.J.; Lin, C.M.; Chiu, Y.H.; Wu, C.J.; Cheng, T.L.; Liu, S.J.; Liao, K.W. Liposome-based polymer complex as a novel adjuvant: Enhancement of specific antibody production and isotype switch. Int. J. Nanomed. 2012, 7, 607–621. [Google Scholar]
- Immordino, M.L.; Dosio, F.; Cattel, L. Stealth liposomes: Review of the basic science, rationale, and clinical applications, existing and potential. Int. J. Nanomed. 2006, 1, 297–315. [Google Scholar]
- Lin, Y.L.; Liu, Y.K.; Tsai, N.M.; Hsieh, J.H.; Chen, C.H.; Lin, C.M.; Liao, K.W. A Lipo-PEG-PEI complex for encapsulating curcumin that enhances its antitumor effects on curcumin-sensitive and curcumin-resistance cells. Nanomed. Nanotechnol. Biol. Med. 2012, 8, 318–327. [Google Scholar] [CrossRef]
- Lin, Y.L.; Chen, C.H.; Wu, H.Y.; Tsai, N.M.; Jian, T.Y.; Chang, Y.C.; Lin, C.H.; Wu, C.H.; Hsu, F.T.; Leung, T.K.; et al. Inhibition of breast cancer with transdermal tamoxifen-encapsulated lipoplex. J. Nanobiotechnol. 2016, 14, 11. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.-L.; Tsai, N.-M.; Chen, C.-H.; Liu, Y.-K.; Lee, C.-J.; Chan, Y.-L.; Wang, Y.-S.; Chang, Y.-C.; Lin, C.-H.; Huang, T.-H.; et al. Specific drug delivery efficiently induced human breast tumor regression using a lipoplex by non-covalent association with anti-tumor antibodies. J. Nanobiotechnol. 2019, 17, 1. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.L.; Chang, K.F.; Huang, X.F.; Hung, C.L.; Chen, S.C.; Chao, W.R.; Liao, K.W.; Tsai, N.M. Liposomal n-butylidenephthalide protects the drug from oxidation and enhances its antitumor effects in glioblastoma multiforme. Int. J. Nanomed. 2015, 10, 6009–6020. [Google Scholar]
- Gao, H.-W.; Chang, K.-F.; Huang, X.-F.; Lin, Y.-L.; Weng, J.-C.; Liao, K.-W.; Tsai, N.-M. Antitumor Effect of n-Butylidenephthalide Encapsulated on B16/F10 Melanoma Cells In Vitro with a Polycationic Liposome Containing PEI and Polyethylene Glycol Complex. Molecules 2018, 23, 3224. [Google Scholar] [CrossRef] [Green Version]
- Lin, Y.L.; Huang, X.F.; Chang, K.F.; Liao, K.W.; Tsai, N.M. Encapsulated n-Butylidenephthalide Efficiently Crosses the Blood-Brain Barrier and Suppresses Growth of Glioblastoma. Int. J. Nanomed. 2020, 15, 749–760. [Google Scholar] [CrossRef] [Green Version]
- Blau, S.; Jubeh, T.T.; Haupt, S.M.; Rubinstein, A. Drug targeting by surface cationization. Crit. Rev. Ther. Drug Carrier Syst. 2000, 17, 425–465. [Google Scholar] [CrossRef]
- Emam, S.E.; Ando, H.; Lila, A.S.A.; Shimizu, T.; Okuhira, K.; Ishima, Y.; Mahdy, M.A.; Ghazy, F.S.; Sagawa, I.; Ishida, T. Liposome co-incubation with cancer cells secreted exosomes (extracellular vesicles) with different proteins expressions and different uptake pathways. Sci. Rep. 2018, 8, 14493. [Google Scholar] [CrossRef] [PubMed]
- Longley, D.B.; Harkin, D.P.; Johnston, P.G. 5-fluorouracil: Mechanisms of action and clinical strategies. Nat. Rev. Cancer 2003, 3, 330–338. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Huang, S.; Wei, Y.; Cao, S.; Pi, C.; Feng, T.; Liang, J.; Zhao, L.; Ren, G. Curcumin Enhances the Anticancer Effect Of 5-fluorouracil against Gastric Cancer through Down-Regulation of COX-2 and NF- kappaB Signaling Pathways. J. Cancer 2017, 8, 3697–3706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bray, F.; Ferlay, J.; Soerjomataram, I.; Siegel, R.L.; Torre, L.A.; Jemal, A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA A Cancer J. Clin. 2018, 68, 394–424. [Google Scholar] [CrossRef] [Green Version]
- Kastan, M.B.; Bartek, J. Cell-cycle checkpoints and cancer. Nature 2004, 432, 316–323. [Google Scholar] [CrossRef]
- Li, Y.; Pan, J.; Gou, M. The Anti-Proliferation, Cycle Arrest and Apoptotic Inducing Activity of Peperomin E on Prostate Cancer PC-3 Cell Line. Molecules 2019, 24, 1472. [Google Scholar] [CrossRef] [Green Version]
- Zhao, L.; Wientjes, M.G.; Au, J.L. Evaluation of combination chemotherapy: Integration of nonlinear regression, curve shift, isobologram, and combination index analyses. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2004, 10, 7994–8004. [Google Scholar] [CrossRef] [Green Version]
Sample Availability: Samples of the compounds are not available from the authors. |






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cell Line | Tumor Type | BP/LPPC | BP | BP/Liposome | 5-FU |
|---|---|---|---|---|---|
| CRC cells | |||||
| HT-29 | Human colorectal adenocarcinoma | 9.61 ± 2.97 a,b | 73.91 ± 2.98 | 139.33 ± 2.32 | >10 |
| CT26 | Mouse colorectal adenocarcinoma | 11.01 ± 3.96 a,b,c | 47.87 ± 2.30 | 69.61 ± 1.74 | 1.25 ± 1.86 |
| Normal cells | |||||
| SVEC | Mouse vascular endothelial cell | 24.15 ± 0.40 a,c | 97.48 ± 4.80 | ND | 7.33 ± 2.39 |
| MDCK | Canine kidney epithelial cell | 26.74 ± 3.82 a,b | 116.62 ± 0.73 | 148.69 ± 12.21 | >10 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, K.-F.; Chang, J.T.; Huang, X.-F.; Lin, Y.-L.; Liao, K.-W.; Huang, C.-W.; Tsai, N.-M. Antitumor Effects of N-Butylidenephthalide Encapsulated in Lipopolyplexs in Colorectal Cancer Cells. Molecules 2020, 25, 2394. https://doi.org/10.3390/molecules25102394
Chang K-F, Chang JT, Huang X-F, Lin Y-L, Liao K-W, Huang C-W, Tsai N-M. Antitumor Effects of N-Butylidenephthalide Encapsulated in Lipopolyplexs in Colorectal Cancer Cells. Molecules. 2020; 25(10):2394. https://doi.org/10.3390/molecules25102394
Chicago/Turabian StyleChang, Kai-Fu, Jinghua Tsai Chang, Xiao-Fan Huang, Yu-Ling Lin, Kuang-Wen Liao, Chien-Wei Huang, and Nu-Man Tsai. 2020. "Antitumor Effects of N-Butylidenephthalide Encapsulated in Lipopolyplexs in Colorectal Cancer Cells" Molecules 25, no. 10: 2394. https://doi.org/10.3390/molecules25102394