Viability, Enzymatic and Protein Profiles of Pseudomonas aeruginosa Biofilm and Planktonic Cells after Monomeric/Gemini Surfactant Treatment


Abstract
:1. Introduction
2. Results and Discussion
2.1. Minimal Inhibitory Concentration (MIC)
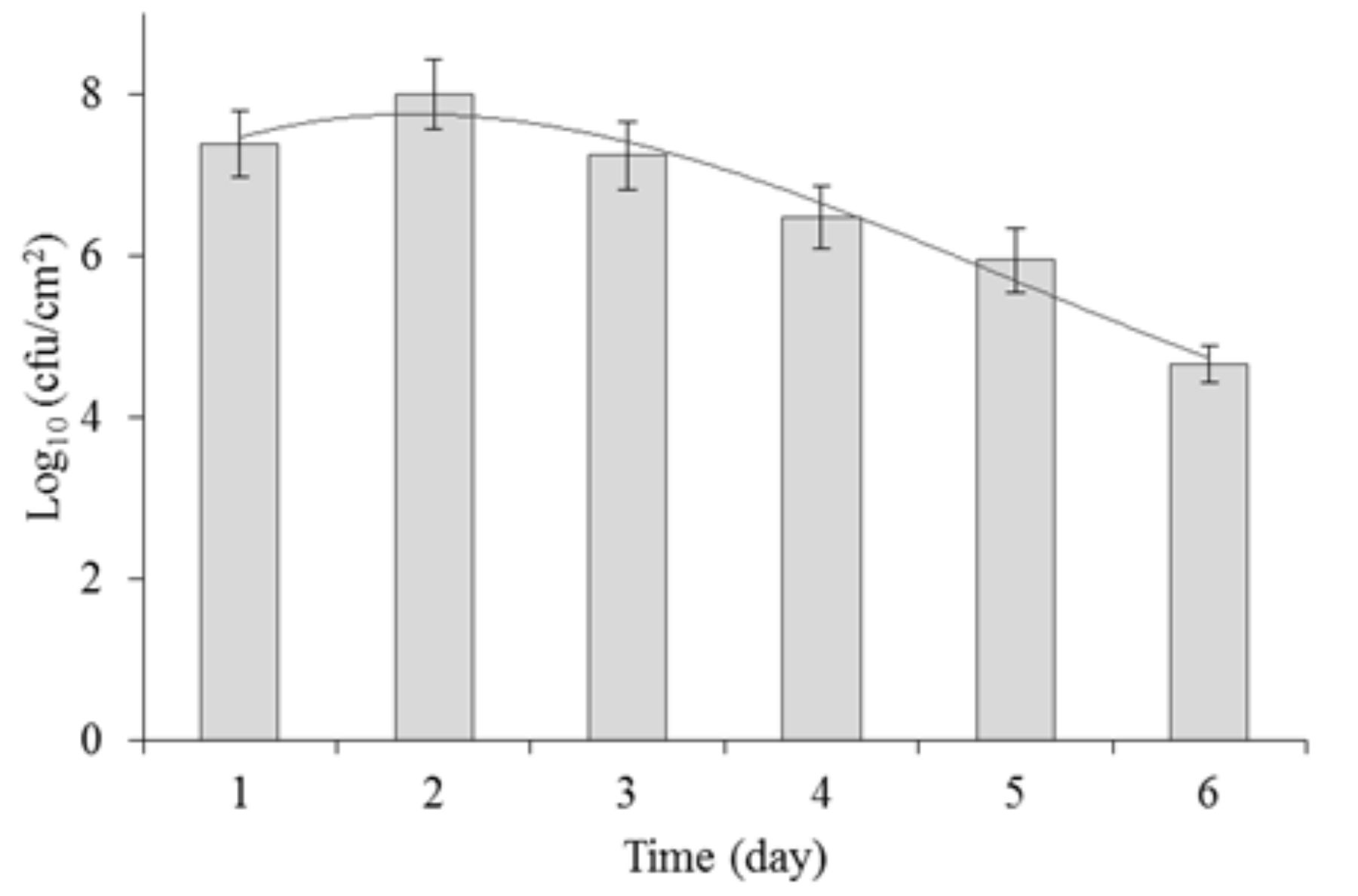
2.2. Biofilm Formation by P. aeruginosa
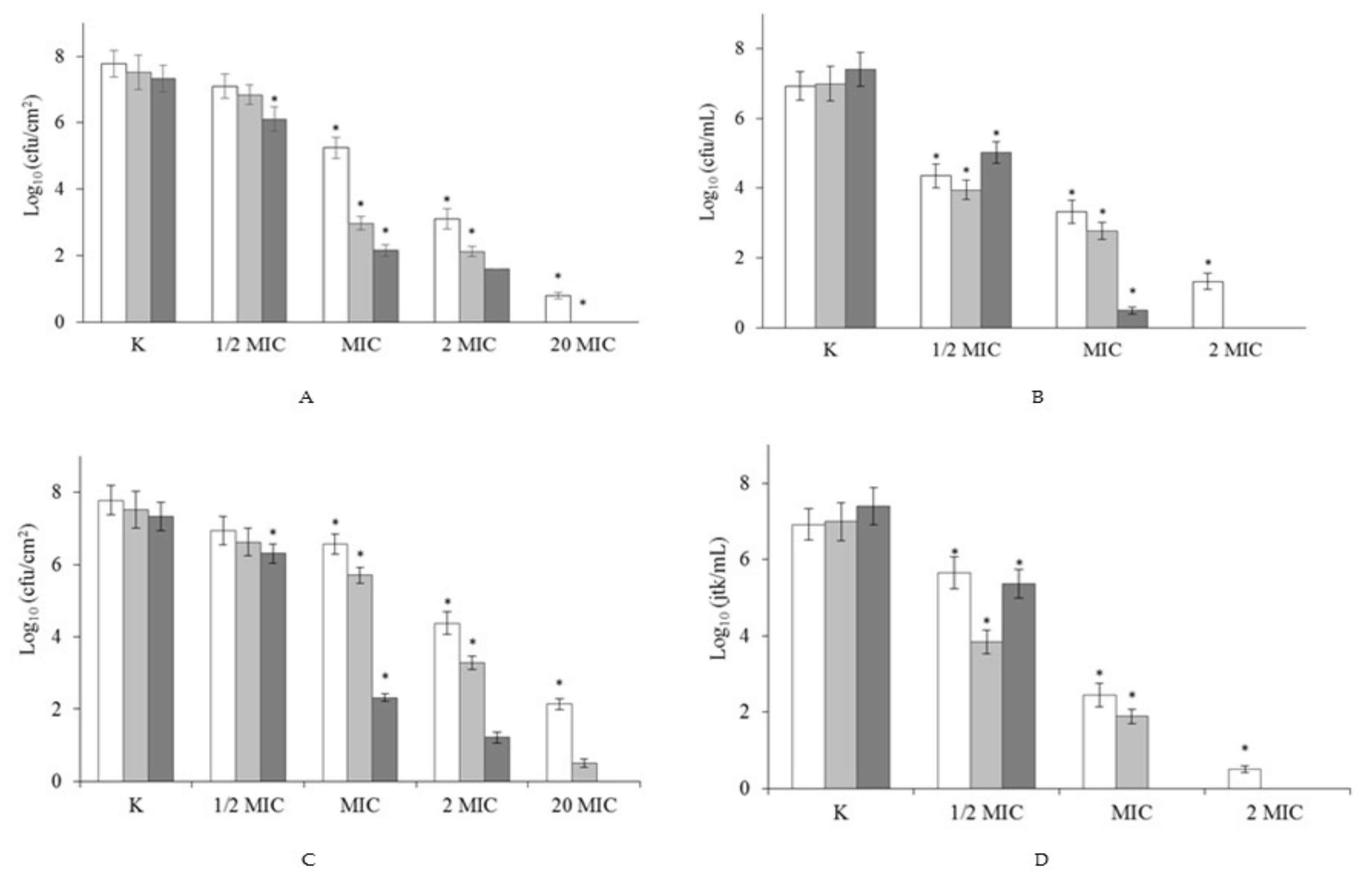
2.3. Effect of Surfactants on Pre-Formed Biofilm
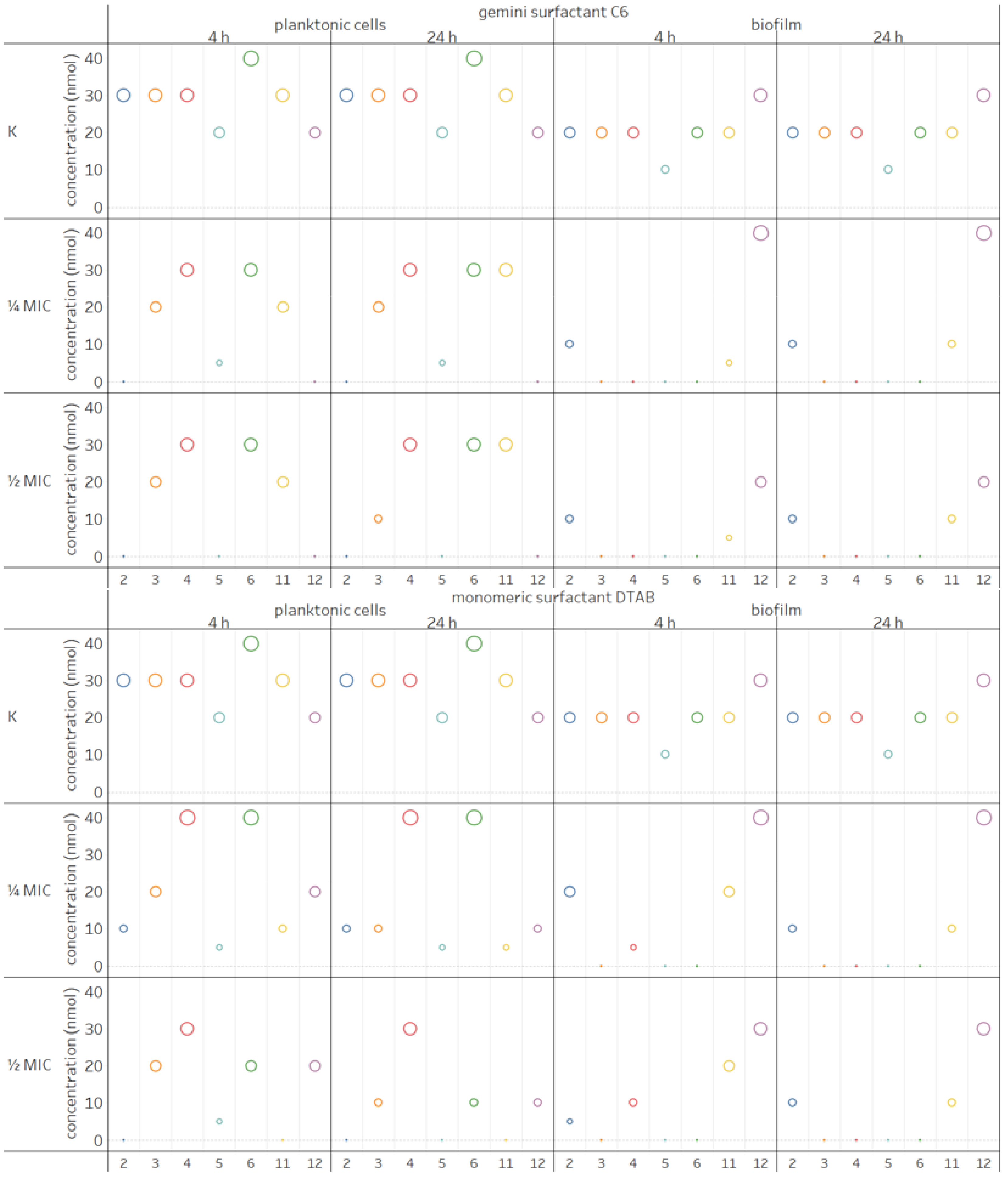
2.4. Influence of Surfactants on Enzymatic Profiles of Biofilms and Planktonic Cells
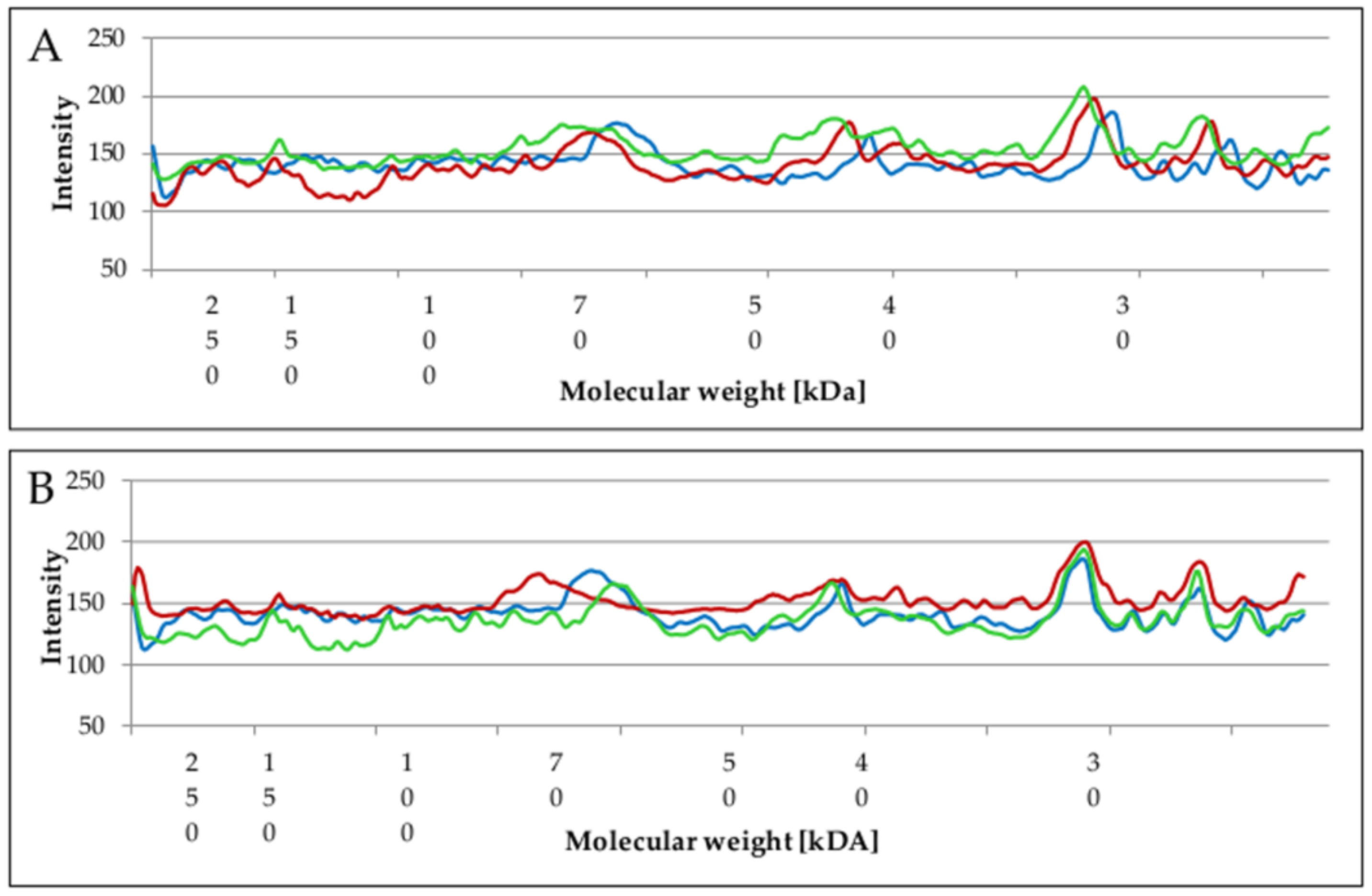
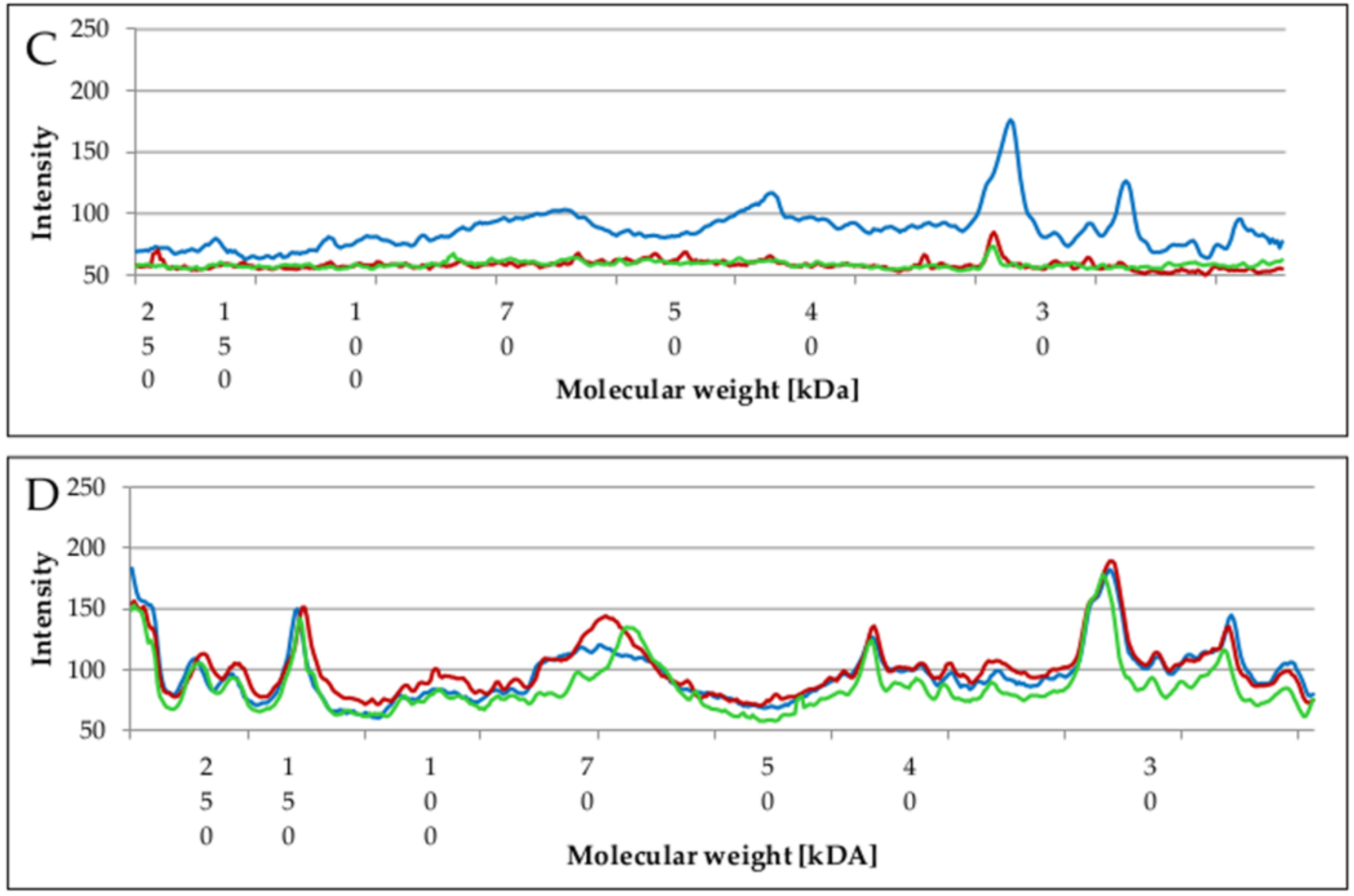
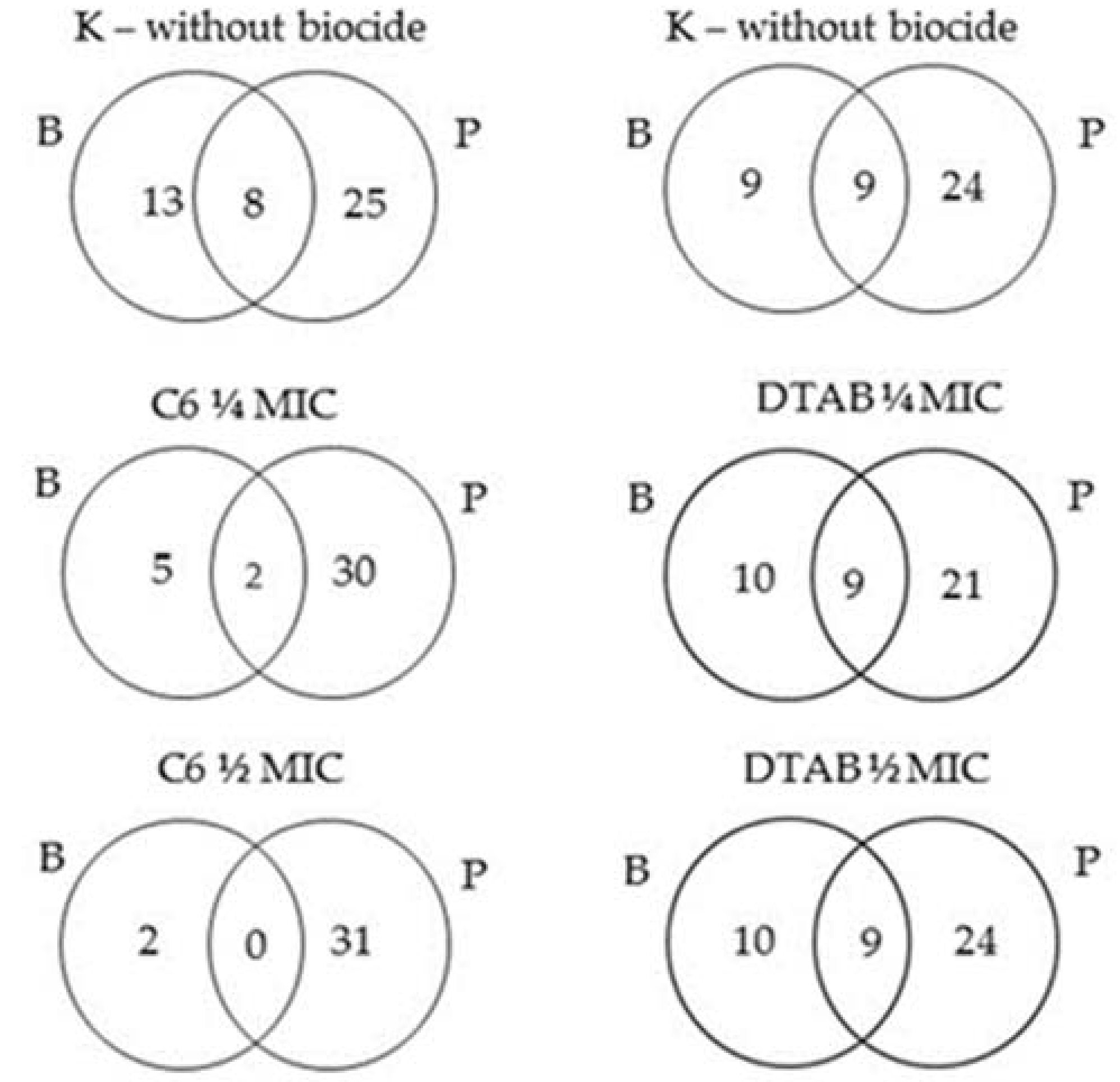
2.5. Changes of Proteins Profile in Biofilm and Planktonic Cells after Surfactant Treatment
3. Materials and Methods
3.1. Microorganisms and Technical Materials
3.2. Antimicrobial Agents
3.3. Minimal Inhibitory Concentration
3.4. Biofilm Formation
3.5. Effect of Surfactants on Pre-Formed Biofilm and Planktonic Cells
3.6. Enzymatic Profile in Biofilm and Planktonic Cells after Surfactant Treatments
3.7. Preparation of Protein Extracts
3.8. Protein Profile Analysis by SDS-PAGE
3.9. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Qu, L.; She, P.; Wang, Y.; Liu, F.; Zhang, D.; Chen, L.; Luo, Z.; Xu, H.; Qi, Y.; Wu, Y. Effect of norspermidine on Pseudomonas aeruginosa biofilm formation and eradication. Microbiol. Open 2016, 5, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Minbiole, K.P.C.; Jennings, M.C.; Ator, L.E.; Black, J.W.; Grenier, M.C.; LaDow, J.E.; Caran, K.L.; Seifert, K.; Wues, W.M. From antimicrobial activity to mechanism of resistance: The multifaceted role of simple quaternary ammonium compounds in bacterial eradication. Tetrahedron 2016, 72, 3559–3566. [Google Scholar] [CrossRef]
- Tielen, P.; Rosenau, F.; Wilhelm, S.; Jaeger, K.-E.; Flemming, H.-C.; Wingender, J. Extracellular enzymes affect biofilm formation of mucoid Pseudomonas aeruginosa. Microbiology 2010, 156, 2239–2252. [Google Scholar] [CrossRef] [PubMed]
- Van Delden, C. Virulence factors in Pseudomonas aeruginosa. In Pseudomonas. Vol. 2 Virulence and Gene Regulations; Ramos, J.-L., Ed.; Kluwer Academic: New York, NY, USA, 2004; pp. 3–45. [Google Scholar]
- Sarabhai, S.; Sharma, P.; Capalash, N. Ellagic acid derivatives from Terminalia chebula Retz. downregulate the expression of quorum sensing genes to attenuate Pseudomonas aeruginosa PAO1 virulence. PLoS ONE 2013, 8, e5344. [Google Scholar] [CrossRef] [PubMed]
- Sauer, K.; Camper, A.K.; Ehrlich, G.D.; Costerton, J.W.; Davies, D.G. Pseudomonas aeruginosa displays multiple phenotypes during development as a biofilm. J. Bacteriol. 2002, 184, 1140–1154. [Google Scholar] [CrossRef] [PubMed]
- Mikkelsen, H.; Duck, Z.; Lilley, K.S.; Welch, M. Interrelationships between colonies, biofilms and planktonic cells of Pseudomonas aeruginosa. J. Bacteriol. 2007, 189, 2411–2416. [Google Scholar] [CrossRef] [PubMed]
- Costerton, J.W.; Irvin, R.T.; Cheng, K.-J. The bacterial glycocalyx in nature and disease. Annu. Rev. Microbiol. 1981, 35, 299–324. [Google Scholar] [CrossRef] [PubMed]
- Toyofuku, M.; Roschitzki, B.; Riedel, K.; Eberl, L. Identyfication of proteins associated with the Pseudomonas aeruginosa biofilm extracellular matrix. J. Proteome Res. 2012, 11, 4906–4915. [Google Scholar] [CrossRef] [PubMed]
- HØiby, N.; Bjarnsholt, T.; Givskov, M.; Molin, S.; Ciofu, O. Antibiotic resistance of bacterial biofilms. Int. J. Antimicrob. Agents 2010, 35, 322–332. [Google Scholar] [CrossRef] [PubMed]
- Kouidhi, B.; Qurashi, Y.M.A.I.; Chaieb, K. Drug resistance of bacterial dental biofilm and the potential use of natural compounds as alternative for prevention and treatment. Microb. Pathog. 2015, 80, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Dubois-Brissonnet, F.; Greub, G.; Thomas, V.; Briandet, R. Dynamics of the action of biocides in Pseudomonas aeruginosa biofilms. Antimicrob. Agents Chemother. 2011, 55, 2648–2654. [Google Scholar] [CrossRef] [PubMed]
- Taylor, P.K.; Yeung, A.T.Y.; Hancock, R.E.W. Antibiotic resistance in Pseudomonas aeruginosa biofilms: Towards the development of novel anti-biofilm therapies. J. Biotech. 2014, 191, 121–130. [Google Scholar] [CrossRef] [PubMed]
- Bridier, A.; Briandet, R.; Thomas, V.; Dubois-Brissonnet, F. Resistance of bacterial biofilms to disinfectants: A review. Biofouling 2011, 27, 1017–1032. [Google Scholar] [CrossRef] [PubMed]
- Chang, C.-Y. Surface sensing for biofilm formation in Pseudomonas aeruginosa. Front. Microbiol. 2018, 8, 2671–2678. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.C.; Ator, L.E.; Paniak, T.J.; Minbiole, K.P.C.; Wuest, W.M. Biofilm-eradicating properties of Quaternary Ammonium Amphiphiles: Simple mimics of antimicrobial peptides. ChemBioChem 2014, 15, 2211–2215. [Google Scholar] [CrossRef] [PubMed]
- Koziróg, A.; Kręgiel, D.; Brycki, B. Action of monomeric/gemini surfactants on free cells and biofilm of Asaia lannensis. Molecules 2017, 22, 2036–2049. [Google Scholar] [CrossRef] [PubMed]
- Paniak, T.J.; Jennings, M.C.; Shanahan, P.C.; Joyce, M.D.; Santiago, C.N.; Wuest, W.M.; Minbiole, K.P.C. The antimicrobial activity of mono-, bis-, tris-, and tetracationic amphiphiles derived from simple polyamine platforms. Bioorg. Med. Chem. Lett. 2014, 24, 5824–5828. [Google Scholar] [CrossRef] [PubMed]
- Murguia, M.C.; Vaillard, V.A.; Sánchez, V.G.; Di Conza, J.; Grau, R.J. Synthesis, surface-active properties and antimicrobial activities of new double-chain gemini surfactants. J. Oleo Sci. 2008, 57, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Koziróg, A.; Brycki, B. Monomeric and gemini surfactants as antimicrobial agents—Influence on environmental and reference strains. Acta Biochim. Pol. 2015, 62, 879–883. [Google Scholar] [CrossRef] [PubMed]
- Garrett, R.; Bhakoo, M.; Zhang, Z. Bacterial adhesion and biofilms on surfaces. Prog. Nat. Sci. 2008, 18, 1049–1056. [Google Scholar] [CrossRef]
- Meira, Q.G.S.; Athayde, A.J.A.A.; Siqueira-Júnior, J.P.; Souza, E.L. Influence of temperature and surface kind on biofilm formation by Staphylococcus aureus form food-contact surfaces and sensitivity to sanitizers. Food Control 2012, 25, 469–475. [Google Scholar] [CrossRef]
- Rühs, P.A.; Böni, L.; Fuller, G.G.; Inglis, R.F.; Fischer, P. In-situ quantification of the interfacial rheological response of bacterial biofilms to environmental stimuli. PLoS ONE 2013, 8, e78524. [Google Scholar] [CrossRef] [PubMed]
- Tré-Hardy, M.; Vanderbist, F.; Traore, H.; Devleeschouwer, M.J. In vitro activity of antibiotic combinations against Pseudomonas aeruginosa biofilm and planktonic culture. Int. J. Antimicrob. Agents 2008, 321, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Wilhelm, S.; Gdynia, A.; Tielen, P.; Rosenau, F.; Jaeger, K.-E. The autotransporter esterase EstA of Pseudomonas aeruginosa is required for rhamnolipid production, cell motility and biofilm formation. J. Bacteriol. 2007, 189, 6695–6703. [Google Scholar] [CrossRef] [PubMed]
- Wingender, J.; Neu, T.R.; Flemming, H.-C. What are bacterial extracellular polymeric substances. In Microbial Eextracellular Polymeric Substances; Wingender, J., Neu, T.R., Flemming, H.-C., Eds.; Springer-Verlag: Berlin/Heidelberg, Germany, 1999; pp. 1–19. [Google Scholar]
- Zhang, W.; Sun, J.; Ding, W.; Lin, J.; Tian, R.; Lu, L.; Liu, X.; Shen, X.; Qian, P.-Y. Extracellular matrix- associated proteins from an integral and dynamic system during Pseudomonas aeruginosa biofilm development. Front. Cell. Infect. Microbiol. 2015, 5, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Obłąk, E.; Piecuch, A.; Guz-Regner, K.; Dworniczek, E. Antibacterial activity of gemini quaternary ammonium salts. FEMS Microbiol. Lett. 2014, 350, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Kadurugamuwa, J.I.; Beveridge, T.J. Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: A novel mechanism of enzyme secretion. J. Bacteriol. 1995, 177, 3998–4008. [Google Scholar] [CrossRef] [PubMed]
- Shafiei, M.; Abdi-Ali, A.; Shahcheraghi, F.; Vali, H.; Zahiri, H.S.; Noghabi, K.A. Analysis of Pseudomonas aeruginosa PAO1 biofilm protein profile after exposure to n-butanolic Cyclamen coum extract alone and in combination with ciprofloxacin. Appl. Biochem. Biotechnol. 2017, 182, 1444–1457. [Google Scholar] [CrossRef] [PubMed]
- Couto, N.; Schooling, S.R.; Dutcher, J.R.; Barber, J. Proteom profiles of outer membrane vesicles and extracellular matrix of Pseudomonas aeruginosa biofilm. J. Proteome Res. 2015, 14, 4207–4222. [Google Scholar] [CrossRef] [PubMed]
- Song, T.; Duperthuy, M.; Wai, S.N. Sub-optimal treatment of bacterial biofilms. Antibiotics 2016, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Aka, S.T.; Haji, S.H. Sub-MIC of antibiotics induced biofilm formation of Pseudomonas aeruginosa in the presence of chlorhexidine. Braz. J. Microbiol. 2015, 46, 149–154. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.C.; Minbiole, K.P.C.; Wuest, W.M. Quaternary Ammonium Compounds: An antimicrobial mainstay and platform for innovation to address bacterial resistance. Infect. Dis. 2016, 1, 288–303. [Google Scholar] [CrossRef] [PubMed]
- Kręgiel, D. Attachment of Asaia lannensis to materials commonly used in beverage industry. Food Control 2013, 32, 537–542. [Google Scholar] [CrossRef]
- European Normative EN 1276. Chemical Disinfectants and Antiseptics—Quantitative Suspension Test for the Evaluation of Bactericidal Activity of Chemical Disinfectants and Antiseptics Used in Food, Industrial, Domestic and Institutional Areas—Test Method and Requirements (Phase 2, Step 1); BSI: London, UK, 2010. [Google Scholar]
- Campanac, C.; Pineau, L.; Payard, A.; Baziard-Mouysset, G.; Roques, C. Interaction between biocide cationic agents and bacterial biofilms. Antimicrob. Agents Chemother. 2002, 46, 1469–1474. [Google Scholar] [CrossRef] [PubMed]
- Laemmli, U.K. Cleavage of structural proteins during assembly of head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1989. [Google Scholar]
- Dice, L.R. Measures of the amount of ecologic association between species. Ecology 1945, 26, 297–302. [Google Scholar] [CrossRef]
Sample Availability: Samples of the compounds C6 and DTAB are available from the authors. |







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Biocide | Concentration | Planktonic Cells | Biofilm |
|---|---|---|---|
| K (control without biocide) | 100 | 100 | |
| C6 | ¼ MIC 4 h | 96.8 | 100 |
| ¼ MIC 24 h | 98.5 | 50 | |
| ½ MIC 4 h | 98.5 | 100 | |
| ½ MIC 24 h | 96.8 | 17.4 | |
| DTAB | ¼ MIC 4 h | 95.2 | 84.8 |
| ¼ MIC 24 h | 95.2 | 97.3 | |
| ½ MIC 4 h | 96.9 | 97.3 | |
| ½ MIC 24 h | 100 | 97.3 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Koziróg, A.; Otlewska, A.; Brycki, B. Viability, Enzymatic and Protein Profiles of Pseudomonas aeruginosa Biofilm and Planktonic Cells after Monomeric/Gemini Surfactant Treatment. Molecules 2018, 23, 1294. https://doi.org/10.3390/molecules23061294
Koziróg A, Otlewska A, Brycki B. Viability, Enzymatic and Protein Profiles of Pseudomonas aeruginosa Biofilm and Planktonic Cells after Monomeric/Gemini Surfactant Treatment. Molecules. 2018; 23(6):1294. https://doi.org/10.3390/molecules23061294
Chicago/Turabian StyleKoziróg, Anna, Anna Otlewska, and Bogumił Brycki. 2018. "Viability, Enzymatic and Protein Profiles of Pseudomonas aeruginosa Biofilm and Planktonic Cells after Monomeric/Gemini Surfactant Treatment" Molecules 23, no. 6: 1294. https://doi.org/10.3390/molecules23061294