Evaluation of the Molecular Structural Parameters of Normal Rice Starch and Their Relationships with Its Thermal and Digestion Properties


Abstract
:1. Introduction
2. Results and Discussion
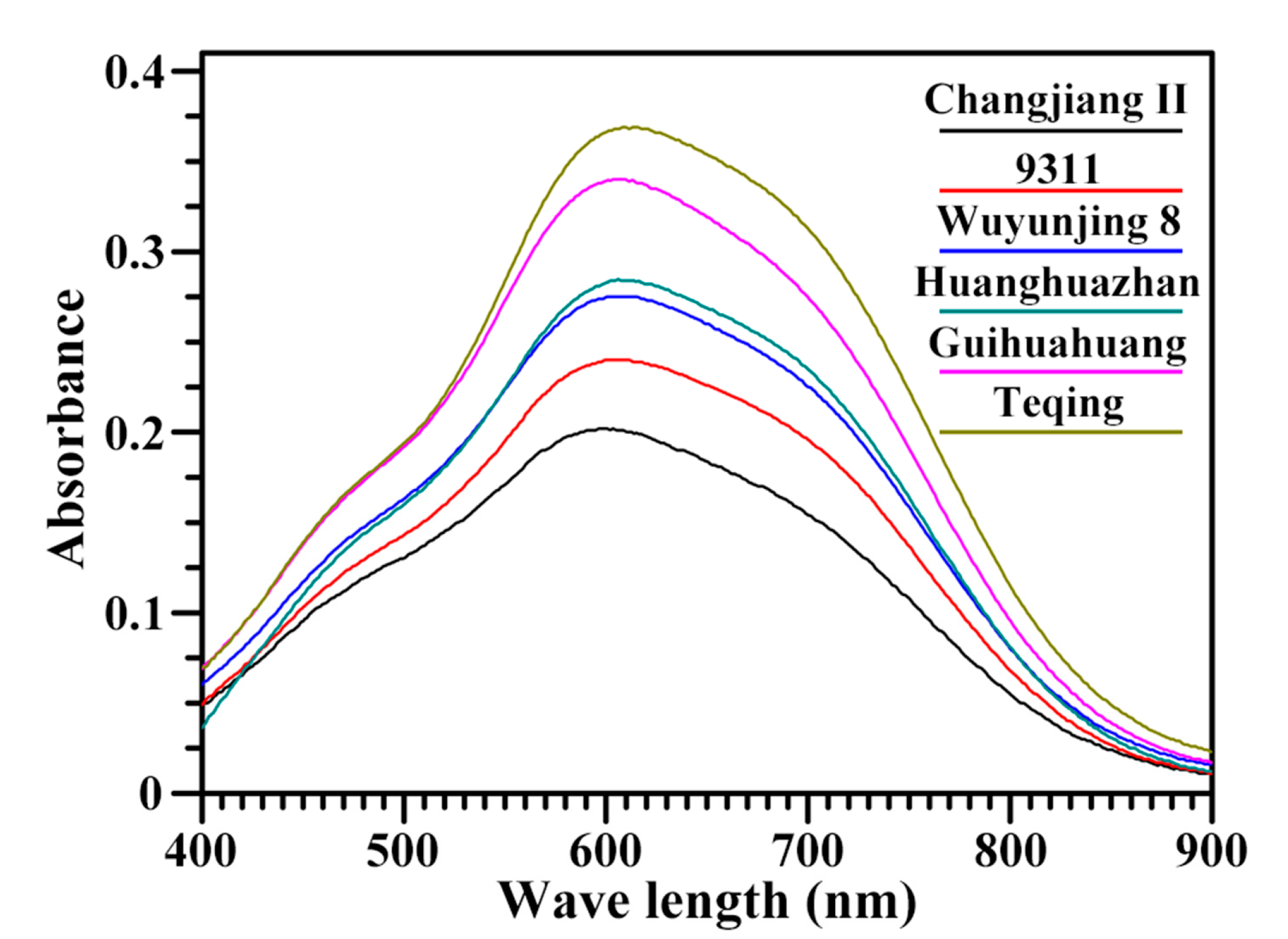
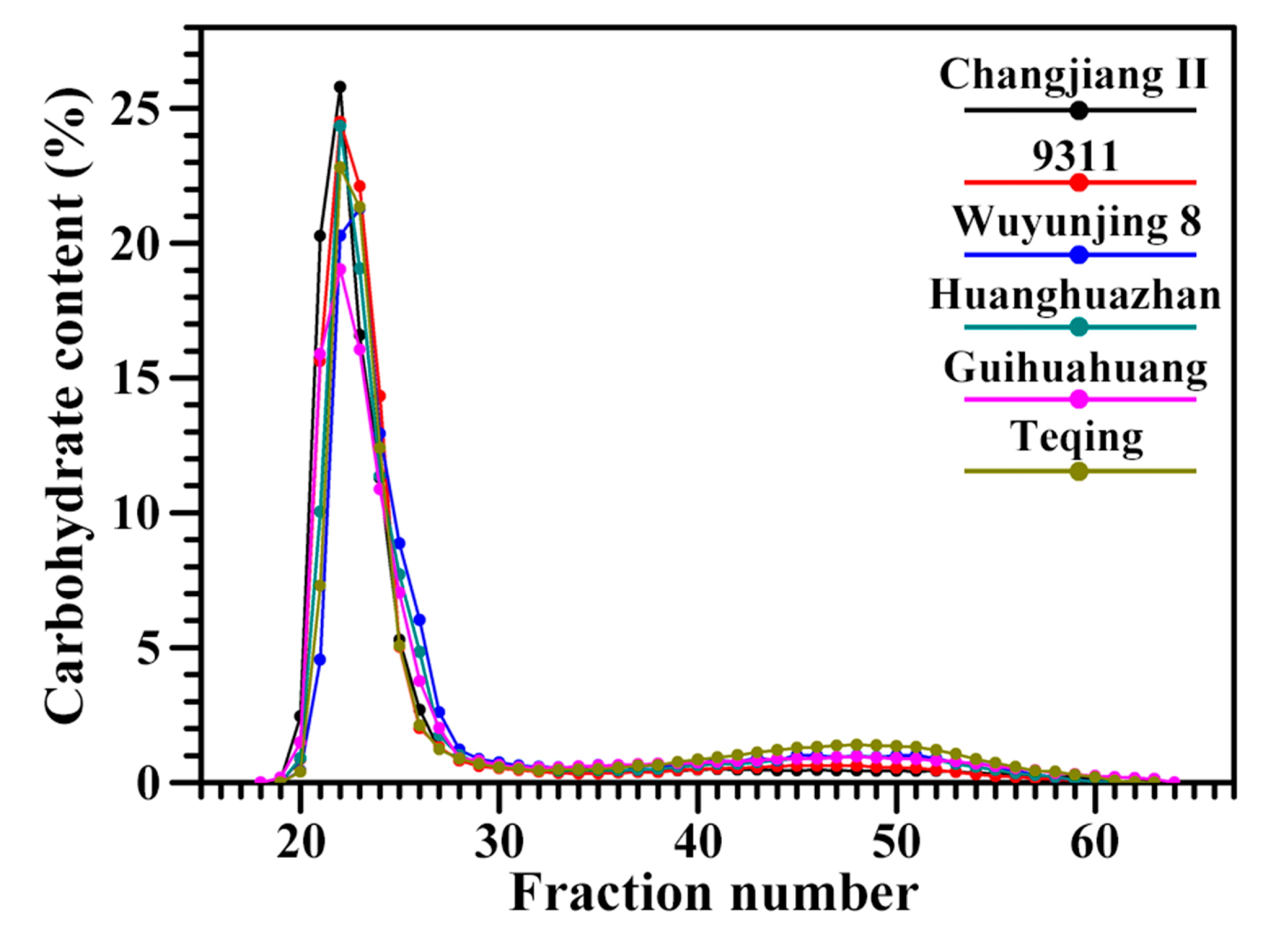
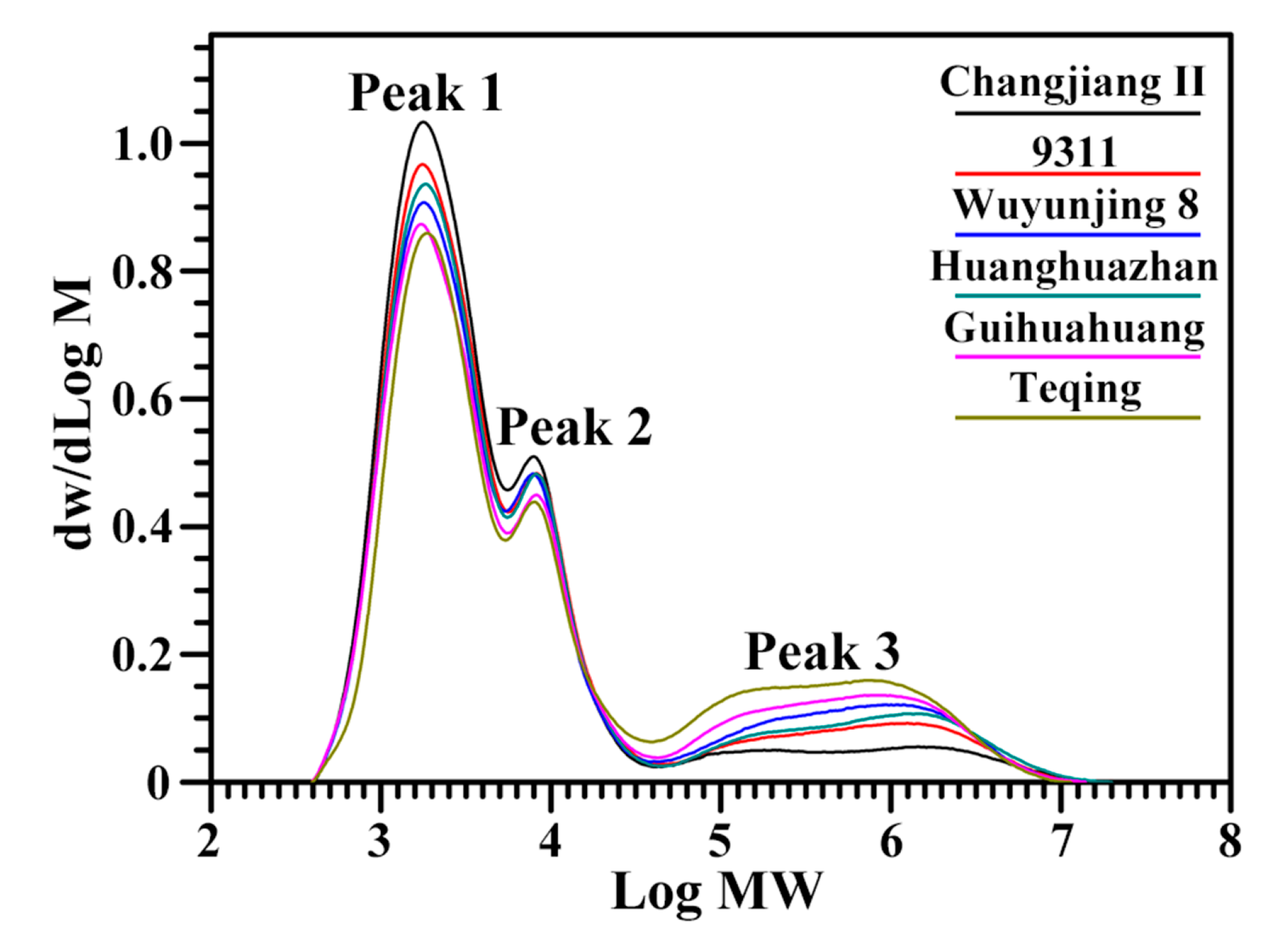
2.1. Molecular Structural Parameters of Starch
2.2. Correlation among Different Molecular Structural Parameters of Starch
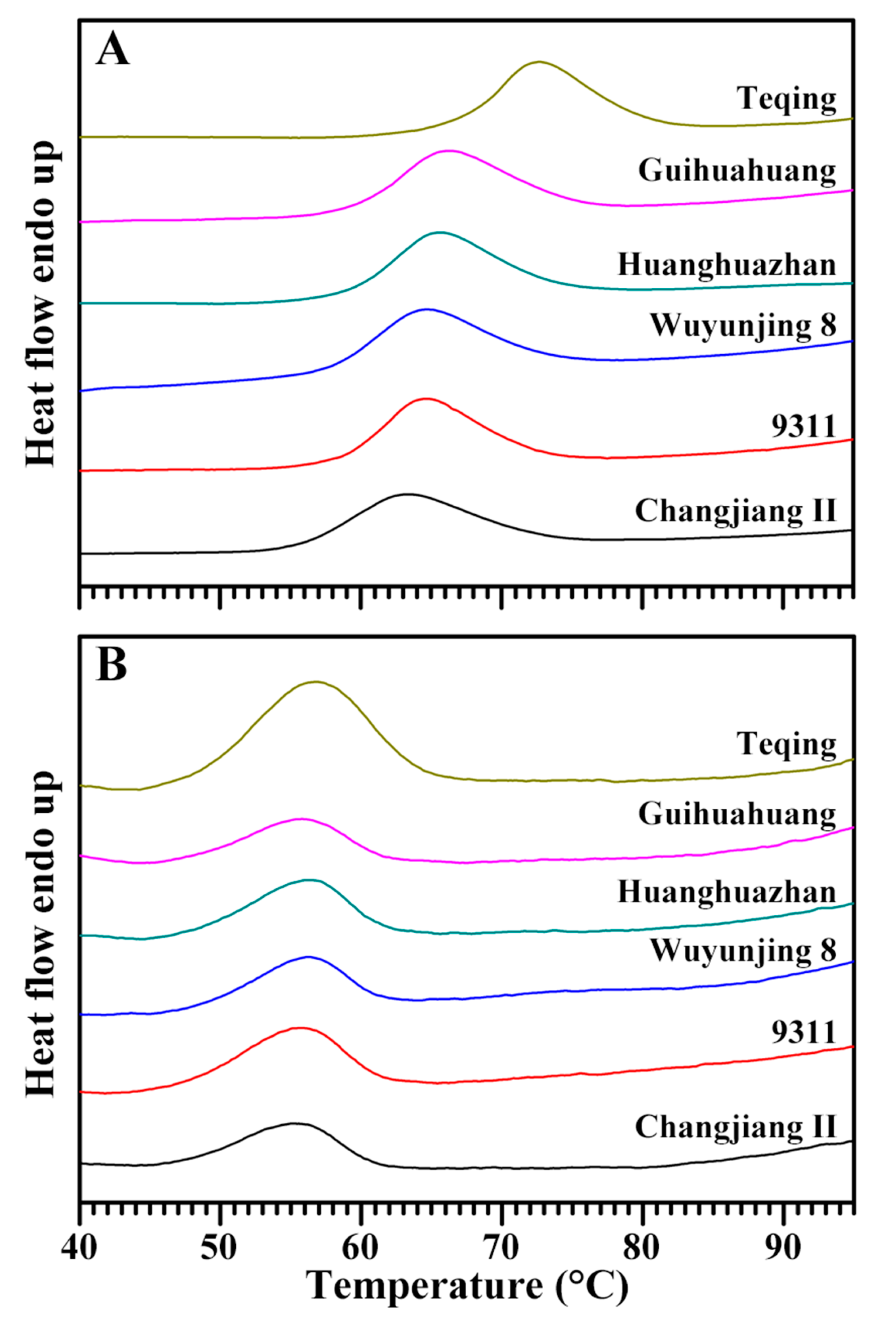
2.3. Thermal Properties of Native and Retrograded Starches
2.4. Correlation between Molecular Structural Parameters and Thermal Properties of Starch
2.5. In Vitro Digestion Properties of Gelatinized and Retrograded Starches
2.6. Correlation between Molecular Structural Parameters and Digestion Properties of Starch
3. Materials and Methods
3.1. Plant Materials
3.2. Isolation of Starch
3.3. Measurements of Iodine Absorption Spectrum and AAC of Starch
3.4. Molecular Weight Distribution of Fully Branched Starch
3.5. Molecular Weight Distribution of Debranched Starch
3.6. Thermal Property Analysis of Starch
3.7. In Vitro Digestion Analysis of Gelatinized and Retrograded Starch
3.8. Statistical Analysis
4. Conclusions
Supplementary Materials
Supplementary File 1Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lin, L.; Guo, D.; Zhao, L.; Zhang, X.; Wang, J.; Zhang, F.; Wei, C. Comparative structure of starches from high-amylose maize inbred lines and their hybrids. Food Hydrocoll. 2016, 52, 19–28. [Google Scholar] [CrossRef]
- Lin, L.; Guo, D.; Huang, J.; Zhang, X.; Zhang, L.; Wei, C. Molecular structure and enzymatic hydrolysis properties of starches from high-amylose maize inbred lines and their hybrids. Food Hydrocoll. 2016, 58, 246–254. [Google Scholar] [CrossRef]
- Lin, L.; Cai, C.; Gilbert, R.G.; Li, E.; Wang, J.; Wei, C. Relationships between amylopectin molecular structures and functional properties of different-sized fractions of normal and high-amylose maize starches. Food Hydrocoll. 2016, 52, 359–368. [Google Scholar] [CrossRef]
- Man, J.; Lin, L.; Wang, Z.; Wang, Y.; Liu, Q.; Wei, C. Different structures of heterogeneous starch granules from high-amylose rice. J. Agric. Food Chem. 2014, 62, 11254–11263. [Google Scholar] [CrossRef] [PubMed]
- Shaik, S.S.; Carciofi, M.; Martens, H.J.; Hebelstrup, K.H.; Blennow, A. Starch bioengineering affects cereal grain germination and seedling establishment. J. Exp. Bot. 2014, 65, 2257–2270. [Google Scholar] [CrossRef] [PubMed]
- Hoover, R. Composition, molecular structure, and physicochemical properties of tuber and root starches: A review. Carbohydr. Polym. 2001, 45, 253–267. [Google Scholar] [CrossRef]
- Alves, F.V.; Polesi, L.F.; Aguiar, C.L.; Sarmento, S.B.S. Structural and physicochemical characteristics of starch from sugar cane and sweet sorghum stalks. Carbohydr. Polym. 2014, 111, 592–597. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Jane, J. Characterization of barley starches of waxy, normal, and high amylose varieties. Carbohydr. Polym. 2000, 41, 365–377. [Google Scholar] [CrossRef]
- Cooke, D.; Gidley, M.J. Loss of crystalline and molecular order during starch gelatinisation: Origin of the enthalpic transition. Carbohydr. Res. 1992, 227, 103–112. [Google Scholar] [CrossRef]
- Park, I.M.; Ibáñez, A.M.; Zhong, F.; Shoemaker, C.F. Gelatinization and pasting properties of waxy and non-waxy rice starches. Starch 2007, 59, 388–396. [Google Scholar] [CrossRef]
- Chung, H.J.; Liu, Q.; Lee, L.; Wei, D. Relationship between the structure, physicochemical properties and in vitro digestibility of rice starches with different amylose contents. Food Hydrocoll. 2011, 25, 968–975. [Google Scholar] [CrossRef]
- Benmoussa, M.; Moldenhauer, K.A.K.; Hamaker, B.R. Rice amylopectin fine structure variability affects starch digestion properties. J. Agric. Food Chem. 2007, 55, 1475–1479. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Bi, J.; Yan, X.; Wang, H.; Zhu, C.; Wang, J.; Wan, J. In vitro measurement of resistant starch of cooked milled rice and physico-chemical characteristics affecting its formation. Food Chem. 2007, 105, 462–468. [Google Scholar] [CrossRef]
- Zhu, L.; Liu, Q.; Wilson, J.D.; Gu, M.; Shi, Y.C. Digestibility and physicochemical properties of rice (Oryza sativa L.) flours and starches differing in amylose content. Carbohydr. Polym. 2011, 86, 1751–1759. [Google Scholar] [CrossRef]
- Nishi, A.; Nakamura, Y.; Tanaka, N.; Satoh, H. Biochemical and genetic analysis of the effects of amylose-extender mutation in rice endosperm. Plant Physiol. 2001, 127, 459–472. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhao, S.; Xiong, S. Morphology and physicochemical properties of mechanically activated rice starch. Carbohydr. Polym. 2010, 79, 341–348. [Google Scholar] [CrossRef]
- Pinto, V.Z.; Vanier, N.L.; Deon, V.G.; Moomand, K.; El Halal, S.L.M.; da Rosa Zavareze, E.; Lin, L.T.; Dias, A.R.G. Effects of single and dual physical modifications on pinhão starch. Food Chem. 2015, 187, 98–105. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.J.; White, P.J.; Pollak, L.; Jane, J. Characterization of starch structures of 17 maize endosperm mutant genotypes with Oh43 inbred line background. Cereal Chem. 1993, 70, 171–179. [Google Scholar]
- Li, L.; Jiang, H.; Campbell, M.; Blanco, M.; Jane, J.L. Characterization of maize amylose-extender (ae) mutant starches. Part I: Relationship between resistant starch contents and molecular structures. Carbohydr. Polym. 2008, 74, 396–404. [Google Scholar] [CrossRef]
- Jiang, H.; Campbell, M.; Wu, Y.; Du, S.; Srichuwong, S.; Jane, J.L. Dosage effect of high-amylose modifier gene(s) on the starch structure of maize amylose-extender mutant. J. Agric. Food Chem. 2015, 63, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Cai, J.; Man, J.; Huang, J.; Liu, Q.; Wei, W.; Wei, C. Relationship between structure and functional properties of normal rice starches with different amylose contents. Carbohydr. Polym. 2015, 125, 35–44. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chen, S.; Ren, X.; Lu, Y.; Liu, D.; Cai, X.; Li, Q.; Gao, J.; Liu, Q. Molecular structure and physicochemical properties of starches from rice with different amylose contents resulting from modification of OsGBSSI activity. J. Agric. Food Chem. 2017, 65, 2222–2232. [Google Scholar] [CrossRef] [PubMed]
- Chung, H.J.; Lim, H.S.; Lim, S.T. Effect of partial gelatinization and retrogradation on the enzymatic digestion of waxy rice starch. J. Cereal Sci. 2006, 43, 353–359. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Z.; Li, X.; Wang, Z. Retrogradation properties of high amylose rice flour and rice starch by physical modification. LWT Food Sci. Technol. 2010, 43, 492–497. [Google Scholar] [CrossRef]
- Vandeputte, G.E.; Vermeylen, R.; Geeroms, J.; Delcour, J.A. Rice starches. I. Structural aspects provide insight into crystallinity characteristics and gelatinisation behaviour of granular starch. J. Cereal Sci. 2003, 38, 43–52. [Google Scholar] [CrossRef]
- Wu, Y.; Chen, Z.; Li, X.; Li, M. Effect of tea polyphenols on the retrogradation of rice starch. Food Res. Int. 2009, 42, 221–225. [Google Scholar] [CrossRef]
- Shi, Y.C.; Seib, P.A. The structure of four waxy starches related to gelatinization and retrogradation. Carbohydr. Res. 1992, 227, 131–145. [Google Scholar] [CrossRef]
- Cai, J.; Cai, C.; Man, J.; Zhou, W.; Wei, C. Structural and functional properties of C-type starches. Carbohydr. Polym. 2014, 101, 289–300. [Google Scholar] [CrossRef] [PubMed]
- Frei, M.; Siddhuraju, P.; Becker, K. Studies on the in vitro starch digestibility and the glycemic index of six different indigenous rice cultivars from the Philippines. Food Chem. 2003, 83, 395–402. [Google Scholar] [CrossRef]
- Syahariza, Z.A.; Sar, S.; Hasjim, J.; Tizzotti, M.J.; Gilbert, R.G. The importance of amylose and amylopectin fine structures for starch digestibility in cooked rice grains. Food Chem. 2013, 136, 742–749. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.; Xu, B.; Qin, F.; Yu, H.; Chen, C.; Meng, X.; Zhu, L.; Wang, Y.; Gu, M.; Liu, Q. C-type starch from high-amylose rice resistant starch granules modified by antisense RNA inhibition of starch branching enzyme. J. Agric. Food Chem. 2010, 58, 7383–7388. [Google Scholar] [CrossRef] [PubMed]
- Konik-Rose, C.; Thistleton, J.; Chanvrier, H.; Tan, I.; Halley, P.; Gidley, M.; Kosar-Hashemi, B.; Wang, H.; Larroque, O.; Ikea, J.; et al. Effects of starch synthase IIa gene dosage on grain, protein and starch in endosperm of wheat. Theor. Appl. Genet. 2007, 115, 1053–1065. [Google Scholar] [CrossRef] [PubMed]
- Tran, T.T.B.; Shelat, K.J.; Tang, D.; Li, E.; Gilbert, R.G.; Hasjim, J. Milling of rice grains. The degradation on three structural levels of starch in rice flour can be independently controlled during grinding. J. Agric. Food Chem. 2011, 59, 3964–3973. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Lin, L.; Wang, J.; Wang, Z.; Liu, Q.; Wei, C. In vitro digestion properties of heterogeneous starch granules from high-amylose rice. Food Hydrocoll. 2016, 54, 1022. [Google Scholar] [CrossRef]
Sample Availability: Samples of the six rice starches are available from the authors. |





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Rice Variety | λmax (nm) | BV b | OD 620/550 | AAC (%) b |
|---|---|---|---|---|
| Changjiang II | 599.2 ± 0.8 a | 0.166 ± 0.004 a | 1.150 ± 0.007 a | 10.4 ± 0.4 a |
| 9311 | 605.8 ± 0.6 b | 0.211 ± 0.001 b | 1.219 ± 0.001 b | 15.5 ± 0.4 b |
| Wuyunjing 8 | 607.8 ± 0.6 bc | 0.243 ± 0.001 c | 1.225 ± 0.013 bc | 18.4 ± 0.2 c |
| Huanghuazhan | 609.8 ± 1.0 cd | 0.255 ± 0.004 d | 1.264 ± 0.019 cd | 19.6 ± 0.4 d |
| Guihuahuang | 610.2 ± 2.8 cd | 0.300 ± 0.004 e | 1.273 ± 0.033 d | 24.7 ± 0.3 e |
| Teqing | 612.3 ± 0.6 d | 0.334 ± 0.002 f | 1.305 ± 0.015 d | 27.9 ± 0.4 f |
| Rice Variety | Fully Branched Starch | Debranched Starch | |||||
|---|---|---|---|---|---|---|---|
| AP (%) b | IC (%) b | ACB (%) b | AP-S (%) c | AP-L (%) c | ACD (%) c | AP-S/L c | |
| Changjiang II | 87.1 ± 0.7 d | 6.6 ± 0.6 a | 6.3 ± 0.3 a | 68.7 ± 0.2 d | 21.1 ± 0.2 a | 10.2 ± 0.1 a | 3.25 ± 0.04 c |
| 9311 | 85.2 ± 1.2 d | 6.7 ± 0.6 a | 8.0 ± 1.0 a | 63.6 ± 0.9 c | 20.8 ± 0.4 a | 15.6 ± 1.3 b | 3.05 ± 0.01 bc |
| Wuyunjing 8 | 79.0 ± 0.7 b | 8.6 ± 0.6 bc | 12.4 ± 0.4 bc | 61.1 ± 1.2 c | 20.5 ± 0.6 a | 18.4 ± 0.6 c | 2.98 ± 0.15 bc |
| Huanghuazhan | 81.8 ± 0.7 c | 7.5 ± 0.6 ab | 10.7 ± 0.8 b | 61.9 ± 0.7 c | 20.7 ± 0.4 a | 17.5 ± 1.1 bc | 3.00 ± 0.02 bc |
| Guihuahuang | 76.9 ± 0.9 b | 8.8 ± 1.0 bc | 14.3 ± 1.9 cd | 58.7 ± 0.7 b | 20.1 ± 0.4 a | 21.2 ± 0.3 d | 2.92 ± 0.09 b |
| Teqing | 74.3 ± 1.2 a | 9.2 ± 0.4 c | 16.4 ± 1.2 d | 53.8 ± 0.4 a | 20.2 ± 0.2 a | 26.0 ± 0.2 e | 2.66 ± 0.05 a |
| λmax | BV | OD 620/550 | AAC | AP | IC | ACB | AP-S | AP-L | ACD | |
|---|---|---|---|---|---|---|---|---|---|---|
| BV | 0.936 ** | |||||||||
| OD 620/550 | 0.983 ** | 0.962 ** | ||||||||
| AAC | 0.936 ** | 0.999 ** | 0.963 ** | |||||||
| AP | ‒0.873 * | ‒0.960 ** | ‒0.870 * | ‒0.956 ** | ||||||
| IC | 0.798 | 0.899 * | 0.778 | 0.893 * | ‒0.985 ** | |||||
| ACB | 0.890 * | 0.973 ** | 0.892 * | 0.969 ** | ‒0.999 ** | 0.976 ** | ||||
| AP-S | −0.878 * | −0.945 ** | −0.876 * | −0.949 ** | 0.947 ** | −0.919 ** | −0.965 ** | |||
| AP-L | −0.929 ** | −0.979 ** | −0.945 ** | −0.979 ** | 0.954 ** | −0.897 * | −0.949 ** | 0.904 * | ||
| ACD | 0.931 ** | 0.983 ** | 0.946 ** | 0.983 ** | −0.959 ** | 0.904 * | 0.970 ** | −1.000 ** | −0.917 * | |
| AP-S/L | −0.906 * | −0.955 ** | −0.929 ** | −0.953 ** | 0.921 ** | −0.858 * | −0.934 ** | 0.992 ** | 0.842 * | −0.987 ** |
| Rice Variety | Native Starch | Retrograded Starch | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| To (°C) b | Tp (°C) b | Tc (°C) b | ΔT (°C) b | ΔH (J/g) b | To (°C) | Tp (°C) | Tc (°C) | ΔT (°C) | ΔH (J/g) | Retro (%) b | |
| Changjiang II | 56.1 ± 0.2 a | 63.1 ± 0.4 a | 72.5 ± 0.6 a | 16.5 ± 0.8 d | 9.88 ± 0.05 a | 47.2 ± 1.6 a | 54.8 ± 1.2 a | 60.2 ± 1.3 a | 13.0 ± 0.3 a | 1.86 ± 0.02 a | 18.8 ± 0.1 a |
| 9311 | 58.4 ± 0.6 bc | 64.7 ± 0.1 b | 72.3 ± 0.4 a | 13.9 ± 0.2 ab | 10.01 ± 0.08 a | 47.3 ± 3.1 a | 55.2 ± 1.1 a | 61.0 ± 0.5 a | 13.7 ± 2.6 a | 2.74 ± 0.30 ab | 27.4 ± 3.2 b |
| Wuyunjing 8 | 58.1 ± 0.1 b | 64.8 ± 0.1 b | 73.2 ± 0.4 ab | 15.1 ± 0.4 bcd | 10.19 ± 0.47 a | 47.2 ± 1.5 a | 55.4 ± 1.2 a | 60.7 ± 0.4 a | 13.5 ± 1.1 a | 2.03 ± 0.32 a | 19.9 ± 2.2 a |
| Huanghuazhan | 59.2 ± 0.1 cd | 65.7 ± 0.1 c | 74.1 ± 0.2 b | 14.9 ± 0.3 bc | 10.29 ± 0.43 a | 47.3 ± 1.7 a | 55.8 ± 0.8 a | 61.5 ± 0.2 a | 14.2 ± 1.5 a | 3.06 ± 0.03 b | 29.8 ± 1.6 b |
| Guihuahuang | 59.7 ± 0.1 d | 66.3 ± 0.1 d | 75.2 ± 0.1 c | 15.5 ± 0.1 cd | 10.48 ± 0.09 a | 47.3 ± 1.1 a | 55.5 ± 1.0 a | 61.3 ± 0.1 a | 14.0 ± 1.1 a | 3.13 ± 0.14 b | 29.8 ± 1.1 b |
| Teqing | 66.8 ± 0.2 e | 72.2 ± 0.1 e | 79.4 ± 0.1 d | 12.7 ± 0.2 a | 10.36 ± 0.13 a | 48.3 ± 0.9 a | 56.6 ± 0.4 a | 64.2 ± 0.1 b | 15.9 ± 1.0 a | 6.41 ± 0.46 c | 61.9 ± 3.6 c |
| λmax | BV | OD 620/550 | AAC | AP | IC | ACB | AP-S | AP-L | ACD | AP-S/L | |
|---|---|---|---|---|---|---|---|---|---|---|---|
| N-To | 0.757 | 0.860 * | 0.828 * | 0.857 * | ‒0.796 | 0.716 | 0.815 * | −0.906 * | −0.674 | 0.896 * | ‒0.946 ** |
| N-Tp | 0.757 | 0.873 * | 0.831 * | 0.869 * | −0.814 * | 0.738 | 0.832 * | −0.911 * | −0.693 | 0.901 * | ‒0.947 ** |
| N-Tc | 0.717 | 0.880 * | 0.800 | 0.873 * | −0.842 * | 0.780 | 0.855 * | −0.885 * | −0.720 | 0.879 * | −0.908 * |
| N-ΔT | −0.659 | −0.619 | −0.691 | −0.623 | 0.518 | −0.421 | −0.543 | 0.737 | 0.424 | −0.720 | 0.801 |
| N-ΔH | 0.904 * | 0.921 ** | 0.902 * | 0.921 ** | −0.883 * | −0.834 * | 0.893 * | −0.840 * | −0.950 ** | 0.853 * | −0.775 |
| R-To | 0.557 | 0.706 | 0.658 | 0.701 | −0.637 | 0.558 | 0.656 | −0.764 | −0.481 | 0.749 | −0.827 * |
| R-Tp | 0.876 * | 0.901 * | 0.917 * | 0.893 * | −0.827 * | 0.740 | 0.849 * | −0.926 ** | −0.713 | 0.916 * | −0.955 ** |
| R-Tc | 0.722 | 0.820 * | 0.804 | 0.815 * | −0.738 | 0.648 | 0.760 | −0.863 * | −0.606 | 0.850 * | −0.912 * |
| R-ΔT | 0.780 | 0.856 * | 0.852 * | 0.851 * | −0.769 | 0.675 | 0.792 | −0.891 * | −0.649 | 0.880 * | −0.934 ** |
| R-ΔH | 0.689 | 0.807 | 0.784 | 0.804 | −0.713 | 0.619 | 0.737 | −0.845 * | −0.598 | 0.833 * | −0.893 * |
| Retro | 0.675 | 0.792 | 0.770 | 0.789 | −0.696 | 0.601 | 0.720 | −0.833 * | −0.578 | 0.820 * | −0.884 * |
| Rice Variety | Gelatinized Starch | Retrograded Starch | ||||
|---|---|---|---|---|---|---|
| RDS (%) b | SDS (%) b | RS (%) b | RDS (%) | SDS (%) | RS (%) | |
| Changjiang II | 83.7 ± 1.0 d | 8.9 ± 0.6 a | 7.3 ± 0.9 a | 76.7 ± 1.3 d | 7.6 ± 1.0 a | 15.7 ± 0.3 a |
| 9311 | 81.1 ± 0.6 c | 9.4 ± 0.1 a | 9.5 ± 0.6 b | 76.6 ± 0.6 d | 7.5 ± 0.3 a | 15.9 ± 0.8 a |
| Wuyunjing 8 | 78.9 ± 0.4 b | 9.6 ± 0.3 a | 11.5 ± 0.1 c | 73.5 ± 1.0 c | 10.1 ± 0.6 b | 16.5 ± 0.7 a |
| Huanghuazhan | 80.7 ± 0.9 bc | 9.4 ± 0.3 a | 9.9 ± 0.6 b | 74.7 ± 0.3 c | 10.1 ± 0.6 b | 15.2 ± 0.8 a |
| Guihuahuang | 78.9 ± 0.6 b | 9.7 ± 0.5 a | 11.4 ± 0.8 c | 71.6 ± 0.6 b | 10.0 ± 0.3 b | 18.5 ± 0.5 b |
| Teqing | 73.6 ± 0.9 a | 11.4 ± 0.4 b | 15.0 ± 0.4 d | 68.7 ± 0.3 a | 8.4 ± 0.2 a | 22.8 ± 0.4 c |
| G-RDS | G-SDS | G-RS | R-RDS | R-SDS | R-RS | |
|---|---|---|---|---|---|---|
| λmax | −0.830 * | 0.723 | 0.853 * | −0.799 | 0.575 | 0.603 |
| BV | −0.914 * | 0.849 * | 0.922 ** | −0.947 ** | 0.457 | 0.813 * |
| OD 620/550 | −0.851 * | 0.784 | 0.862 * | −0.834 * | 0.472 | 0.686 |
| AAC | −0.910 * | 0.845 * | 0.919 ** | −0.942 ** | 0.446 | 0.812 * |
| AP | 0.920 ** | −0.825 * | −0.938 ** | 0.972 ** | −0.526 | −0.810 |
| IC | −0.880 * | 0.769 | 0.904 * | −0.948 ** | 0.552 | 0.772 |
| ACB | −0.927 ** | 0.837 * | 0.943 ** | −0.973 ** | 0.515 | 0.816 * |
| AP-S | 0.970 ** | −0.908 * | −0.976 ** | 0.943 ** | −0.365 | −0.849 * |
| AP-L | 0.806 | −0.686 | −0.834 * | 0.888 * | −0.564 | −0.703 |
| ACD | −0.964 ** | 0.898 * | 0.972 ** | −0.945 ** | 0.381 | 0.844 * |
| AP-S/L | 0.984 ** | −0.945 ** | −0.983 ** | 0.928 ** | −0.284 | −0.869 * |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lin, L.; Zhang, Q.; Zhang, L.; Wei, C. Evaluation of the Molecular Structural Parameters of Normal Rice Starch and Their Relationships with Its Thermal and Digestion Properties. Molecules 2017, 22, 1526. https://doi.org/10.3390/molecules22091526
Lin L, Zhang Q, Zhang L, Wei C. Evaluation of the Molecular Structural Parameters of Normal Rice Starch and Their Relationships with Its Thermal and Digestion Properties. Molecules. 2017; 22(9):1526. https://doi.org/10.3390/molecules22091526
Chicago/Turabian StyleLin, Lingshang, Qing Zhang, Long Zhang, and Cunxu Wei. 2017. "Evaluation of the Molecular Structural Parameters of Normal Rice Starch and Their Relationships with Its Thermal and Digestion Properties" Molecules 22, no. 9: 1526. https://doi.org/10.3390/molecules22091526