
In Vitro Antioxidant Activity and In Vivo Anti-Fatigue Effect of Sea Horse (Hippocampus) Peptides


Abstract
:1. Introduction
2. Results and Discussion
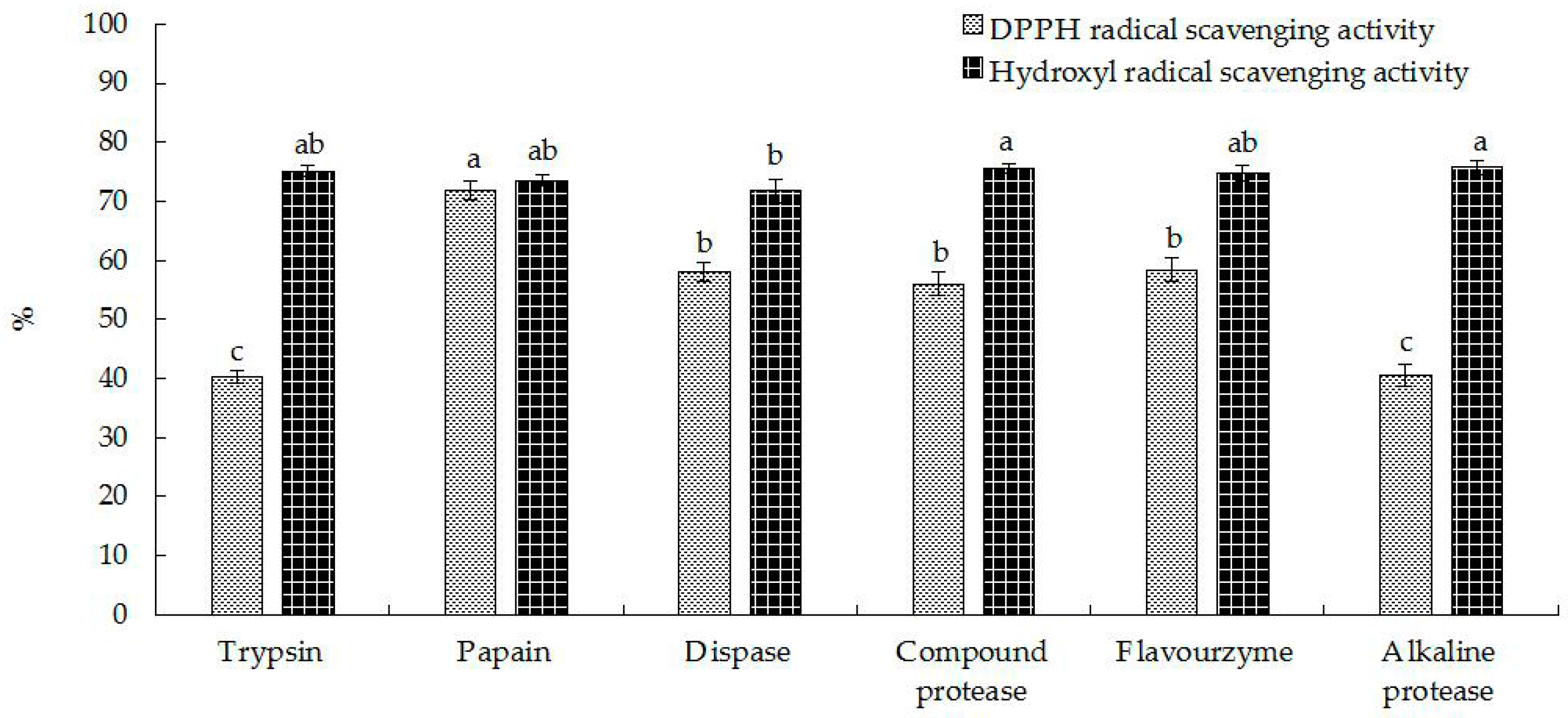
2.1. Preparation of Hippocampal Polypeptides
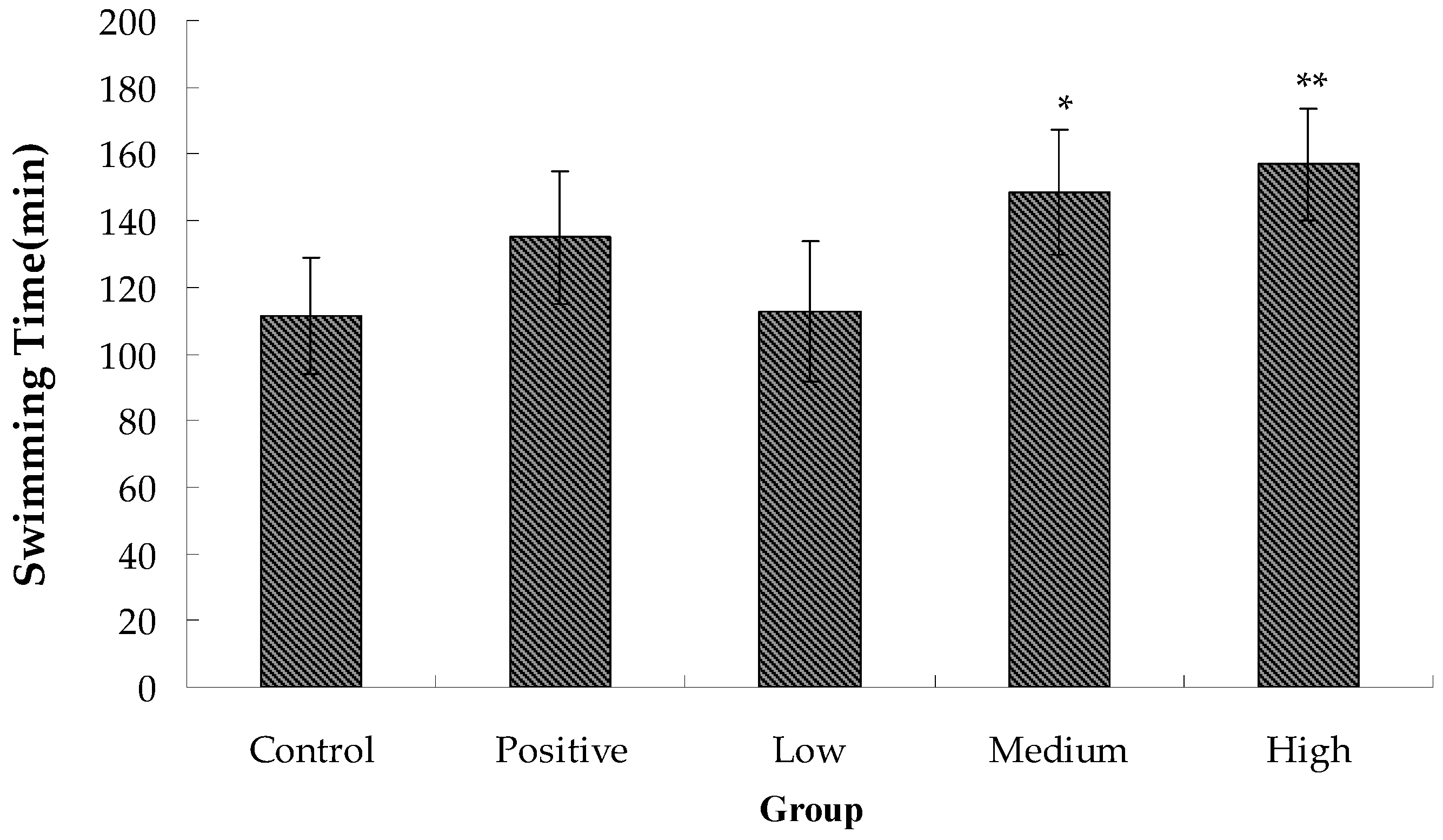
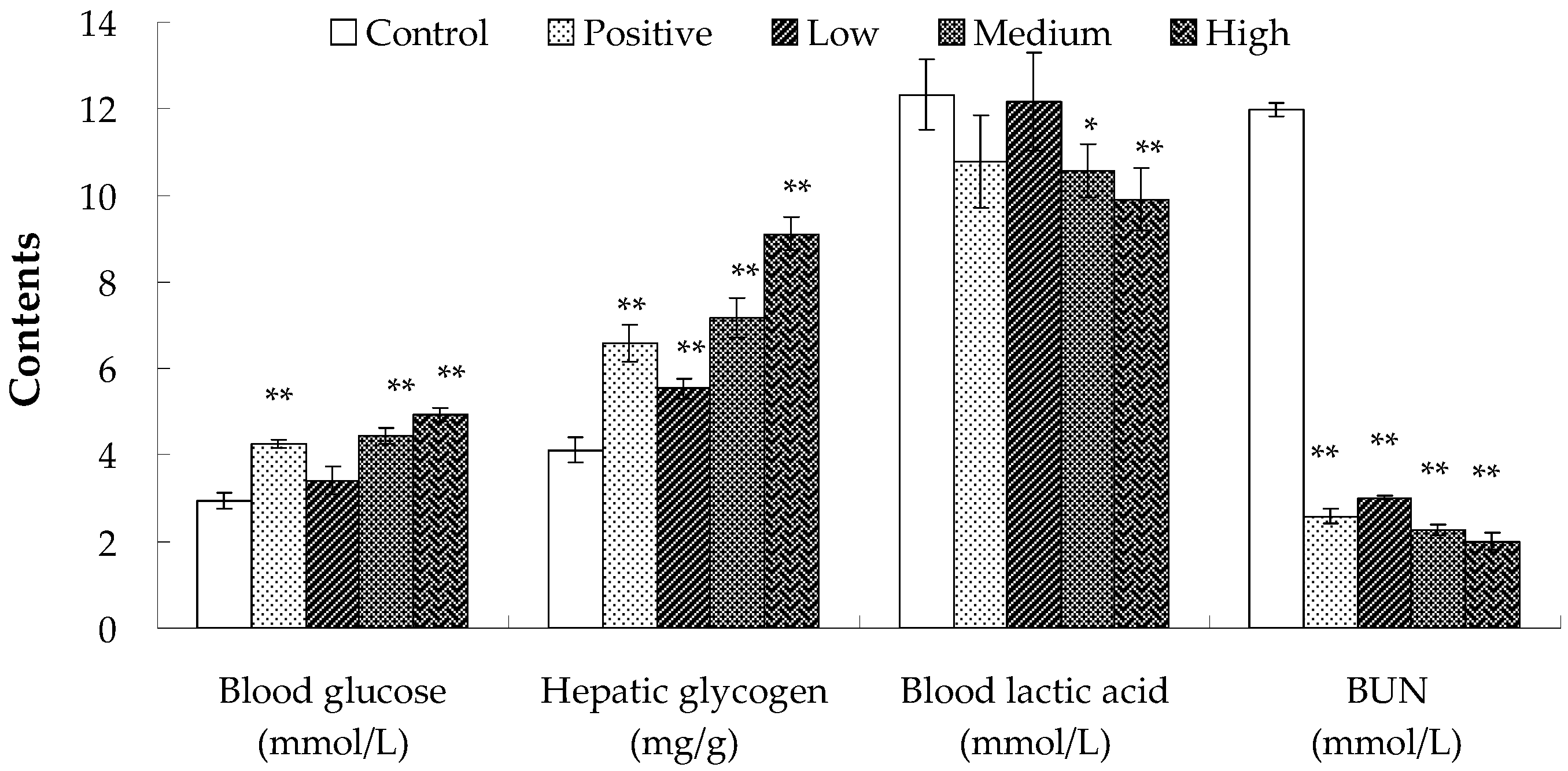
2.2. Anti-Fatigue Activity of Hippocampus Peptide
3. Materials and Methods
3.1. Materials
3.2. Methods
3.2.1. Preparation of Hippocampus Polypeptide
3.2.2. Determination of the Degree of Hydrolysis
3.2.3. Determination of Peptide Yield
3.2.4. Measurement of the Hydroxyl Free Radical Scavenging Rate
3.2.5. Measurement of the DPPH Scavenging Rate
3.2.6. Animal-Experimental Design
3.2.7. Swimming Exhaustion Experiment
3.2.8. Determination of Physiological and Biochemical Indices
3.3. Statistical Analysis
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Zhao, Y.Q.; Zeng, L.; Yang, Z.S.; Huang, F.F.; Ding, G.F.; Wang, B. Anti-fatigue effect by peptide fraction from protein hydrolysate of croceine croaker (Pseudosciaena crocea) swim bladder through inhibiting the oxidative reactions including DNA damage. Mar. Drugs 2016, 14. [Google Scholar] [CrossRef] [PubMed]
- Tan, W.; Yu, K.Q.; Liu, Y.Y.; Ouyang, M.Z.; Yan, M.H.; Luo, R.; Zhao, X.S. Anti-fatigue activity of polysaccharides extract from Radix Rehmanniae Preparata. Int. J. Biol. Macromol. 2012, 50, 59–62. [Google Scholar] [CrossRef] [PubMed]
- Elliott, J.L.; Lal, S. Blood pressure, sleep quality and fatigue in shift working police officers: Effects of a twelve hour roster system on cardiovascular and sleep health. Int. J. Environ. Res. Public Health 2016, 13. [Google Scholar] [CrossRef] [PubMed]
- Trudel, X.; Brisson, C.; Milot, A.; Masse, B.; Vezina, M. Adverse psychosocial work factors, blood pressure and hypertension incidence: Repeated exposure in a 5-year prospective cohort study. J. Epidemiol. Community Health 2016, 70, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Natti, J.; Oinas, T.; Anttila, T. Time pressure, working time control and long-term sickness absence. Occup. Environ. Med. 2015, 72, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Mollica, A.; Pinnen, F.; Azzurra, S.; Costante, R. The evolution of peptide synthesis: From early days to small molecular machines. Curr. Bioact. Compd. 2014, 9, 184–202. [Google Scholar] [CrossRef]
- Ahn, C.B.; Cho, Y.S.; Je, J.Y. Purification and anti-inflammatory action of tripeptide from salmon pectoral fin byproduct protein hydrolysate. Food Chem. 2015, 168, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Chi, C.-F.; Hu, F.-Y.; Wang, B.; Li, T.; Ding, G.-F. Antioxidant and anticancer peptides from the protein hydrolysate of blood clam (Tegillarca granosa) muscle. J. Funct. Foods 2015, 15, 301–313. [Google Scholar] [CrossRef]
- Ennaas, N.; Hammami, R.; Beaulieu, L.; Fliss, I. Purification and characterization of four antibacterial peptides from protamex hydrolysate of Atlantic mackerel (Scomber scombrus) by-products. Biochem. Biophys. Res. Commun. 2015, 462, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Tavares, T.G.; Contreras, M.M.; Amorim, M.; Martín-Álvarez, P.J.; Pintado, M.E.; Recio, I.; Malcata, F.X. Optimisation, by response surface methodology, of degree of hydrolysis and antioxidant and ACE-inhibitory activities of whey protein hydrolysates obtained with cardoon extract. Int. Dairy J. 2011, 21, 926–933. [Google Scholar] [CrossRef]
- Zhang, L.; Li, J.; Zhou, K. Chelating and radical scavenging activities of soy protein hydrolysates prepared from microbial proteases and their effect on meat lipid peroxidation. Bioresour. Technol. 2010, 101, 2084–2089. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.-F.; Li, Y.-Y.; Xu, J.-J.; Su, X.-R.; Gao, X.; Yue, F.-P. Study on effect of jellyfish collagen hydrolysate on anti-fatigue and anti-oxidation. Food Hydrocoll. 2011, 25, 1350–1353. [Google Scholar] [CrossRef]
- You, L.; Zhao, M.; Regenstein, J.M.; Ren, J. In vitro antioxidant activity and in vivo anti-fatigue effect of loach (Misgurnus anguillicaudatus) peptides prepared by papain digestion. Food Chem. 2011, 124, 188–194. [Google Scholar] [CrossRef]
- Kim, G.-N.; Jang, H.-D.; Kim, C.-I. Antioxidant capacity of caseinophosphopeptides prepared from sodium caseinate using Alcalase. Food Chem. 2007, 104, 1359–1365. [Google Scholar]
- Zhang, Z.H.; Xu, G.J.; Xu, L.S.; Wang, Q. Physicaland chemieal analysis of medieinal animals of syngnathidae. J. Chin. Med. Mater. 1997, 30, 140–144. [Google Scholar]
- Yan, J.B.; Ma, R.D.; Yu, L. Medicinal values of Hippocampus. J. China Pharm. Univ. 2002, 90, 48–52. [Google Scholar]
- Sarmadi, B.H.; Ismail, A. Antioxidative peptides from food proteins: A review. Peptides 2010, 31, 1949–1956. [Google Scholar] [CrossRef] [PubMed]
- Su, G.; Ren, J.; Yang, B.; Cui, C.; Zhao, M. Comparison of hydrolysis characteristics on defatted peanut meal proteins between a protease extract from Aspergillus oryzae and commercial proteases. Food Chem. 2011, 126, 1306–1311. [Google Scholar] [CrossRef]
- Zhou, K.; Canning, C.; Sun, S. Effects of rice protein hydrolysates prepared by microbial proteases and ultrafiltration on free radicals and meat lipid oxidation. LWT-Food Sci. Technol. 2013, 50, 331–335. [Google Scholar] [CrossRef]
- Wagenmakers, A.J.; Beckers, E.J.; Brouns, F.; Kuipers, H.; Soeters, P.B.; van der Vusse, G.J. Carbohydrate supplementation, glycogen depletion, and amino acid metabolism during exercise. Am. J. Physiol. Endocrinol. Metab. 1991, 260, 883–890. [Google Scholar]
- Jia, J.M.; Wu, C.F. Antifatigue activity of tissue culture extracts of Saussurea involucrata. Pharm. Biol. 2008, 46, 433–436. [Google Scholar] [CrossRef]
- Jung, K.A.; Han, D.; Kwon, E.K.; Lee, C.H.; Kim, Y.E. Antifatigue effect of Rubus coreanus Miquel extract in mice. J. Med. Food 2007, 10, 689–693. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.J.; Shieh, M.J.; Kuo, S.L.; Lee, C.L.; Pan, T.M. Effect of red mold rice on antifatigue and exercise-related changes in lipid peroxidation in endurance exercise. Appl. Microbiol. Biotechnol. 2006, 70, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Cairns, S.P. Lactic acid and exercise performance-Culprit or friend? Sports Med. 2006, 36, 279–291. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Moon, P.D.; Pak, S.C.; Kim, H.M.; Jeong, H.J. Anti-fatigue effect of Zizania Caudiflora (Turczaninow) Nakai. Am. J. Chin. Med. 2012, 40, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, H.L.; Lu, R.; Zhou, Y.J.; Ma, R.; Lv, J.Q.; Li, X.L.; Chen, L.J.; Yao, Z. The decapeptide CMS001 enhances swimming endurance in mice. Peptides 2008, 29, 1176–1182. [Google Scholar] [CrossRef] [PubMed]
- Li, X.L.; Zhang, H.B.; Xu, H.B. Analysis of chemical components of shiitake polysaccharides and its anti-fatigue effect under vibration. Int. J. Biol. Macromol. 2009, 45, 377–380. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yao, X.B.; Bao, B.L.; Zhang, Y. Anti-fatigue activity of a triterpenoid-rich extract from Chinese bamboo shavings (Caulis bamfusae in taeniam). Phytother. Res. 2006, 20, 872–876. [Google Scholar] [CrossRef] [PubMed]
- Yu, B.; Lu, Z.-X.; Bie, X.-M.; Lu, F.-X.; Huang, X.-Q. Scavenging and anti-fatigue activity of fermented defatted soybean peptides. Eur. Food Res. Technol. 2008, 226, 415–421. [Google Scholar] [CrossRef]
- Correa, A.P.; Daroit, D.J.; Fontoura, R.; Meira, S.M.; Segalin, J.; Brandelli, A. Hydrolysates of sheep cheese whey as a source of bioactive peptides with antioxidant and angiotensin-converting enzyme inhibitory activities. Peptides 2014, 61, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Zhou, D.Y.; Zhu, B.W.; Qiao, L.; Wu, H.T.; Li, D.M.; Yang, J.F.; Murata, Y. In vitro antioxidant activity of enzymatic hydrolysates prepared from abalone (Haliotis discus hannai Ino) viscera. Food Bioprod. Process. 2012, 90, 148–154. [Google Scholar] [CrossRef]
- Miller, J.; Young, C.T. Protein nutritional quality of florunner peanut meal as measured by rat bioassay. J. Agric. Food Chem. 1997, 25, 653–657. [Google Scholar] [CrossRef]
- Zhuang, Y.L.; Sun, L.P.; Zhao, X.; Hou, H.; Li, B.F. Investigation of gelatin polypeptides of jellyfish (Rhopilema esculentum) for their antioxidant activity in vitro. Food Technol. Biotechnol. 2010, 48, 222–228. [Google Scholar]
- Yang, J.I.; Liang, W.S.; Chow, C.J. Siebert, K.J. Process for the production of tilapia retorted skin gelatin hydrolysates with optimized antioxidative properties. Process Biochem. 2009, 44, 1152–1157. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the compounds are not available from the authors.




{kind=link}
{kind=link}
{kind=link}
| Factors/Levels | DPPH Free Radical Scavenging Rate (%) | Hydroxyl Radical Scavenging Rate (%) | Degree of Hydrolysis (%) | Yield of Peptide (%) |
|---|---|---|---|---|
| pH | ||||
| 5.0 | 88.45 ± 1.43 a | 72.46 ± 0.44 cd | 16.35 ± 0.43 ab | 33.12 ± 0.85 a |
| 5.5 | 87.29 ± 0.50 a | 71.52 ± 0.71 d | 17.48 ± 0.75 a | 33.03 ± 0.63 a |
| 6.0 | 83.46 ± 0.93 b | 74.17 ± 0.70 c | 17.70 ± 0.50 a | 32.55 ± 0.50 a |
| 6.5 | 70.98 ± 0.58 c | 74.34 ± 0.64 c | 16.72 ± 0.82 ab | 32.96 ± 0.88 a |
| 7.0 | 56.07 ± 0.70 d | 80.68 ± 0.31 b | 16.54 ± 0.27 ab | 33.42 ± 0.36 a |
| 7.5 | 42.08 ± 0.90 e | 83.30 ± 1.10 a | 15.81 ± 0.36 b | 33.97 ± 0.64 a |
| Temperature (°C) | ||||
| 50 | 73.76 ± 0.36 a | 75.79 ± 1.19 ab | 16.21 ± 0.42 c | 34.06 ± 0.23 a |
| 55 | 74.49 ± 0.24 a | 76.22 ± 0.52 a | 16.65 ± 0.27 bc | 33.81 ± 0.41 ab |
| 60 | 74.49 ± 0.48 a | 76.96 ± 0.26 a | 17.27 ± 0.61 abc | 32.96 ± 0.28 bc |
| 65 | 74.61 ± 0.85 a | 74.24 ± 0.42 bc | 18.14 ± 0.42 a | 32.87 ± 0.37 c |
| 70 | 74.72 ± 0.26 a | 73.67 ± 0.49 c | 17.70 ± 0.21 ab | 33.01 ± 0.31 bc |
| 75 | 74.49 ± 1.13 a | 74.40 ± 0.46 bc | 16.97 ± 0.51 abc | 32.60 ± 0.39 c |
| Time of hydrolysis (min) | ||||
| 20 | 69.69 ± 1.04 a | 73.87 ± 0.29 b | 13.69 ± 0.28 e | 34.88 ± 0.66 a |
| 40 | 71.48 ± 0.73 a | 76.53 ± 0.88 a | 15.08 ± 0.41 d | 34.61 ± 0.28 a |
| 60 | 70.28 ± 0.84 a | 77.18 ± 0.84 a | 16.21 ± 0.65 cd | 33.69 ± 0.58 a |
| 80 | 69.36 ± 0.55 a | 73.20 ± 1.01 b | 17.38 ± 0.25 bc | 31.98 ± 0.74 b |
| 100 | 65.30 ± 0.89 b | 72.18 ± 0.90 bc | 18.14 ± 0.14 ab | 31.93 ± 0.16 b |
| 120 | 60.30 ± 0.96 c | 70.34 ± 0.31 c | 19.13 ± 0.85 a | 31.73 ± 0.46 b |
| Enzyme concentration (U/g) | ||||
| 1000 | 73.57 ± 0.34 a | 78.43 ± 0.54 a | 12.30 ± 0.49 d | 33.30 ± 0.47 a |
| 2000 | 73.38 ± 1.09 a | 77.94 ± 0.46 a | 14.02 ± 0.35 c | 33.10 ± 0.22 ab |
| 3000 | 72.93 ± 0.28 ab | 77.56 ± 0.52 a | 14.89 ± 0.57 c | 32.07 ± 0.83 abc |
| 4000 | 71.40 ± 0.64 bc | 75.12 ± 0.23 b | 16.35 ± 0.12 b | 31.84 ± 0.60 bcd |
| 5000 | 70.96 ± 0.22 c | 75.03 ± 0.76 b | 16.79 ± 0.35 b | 31.61 ± 0.23 cd |
| 6000 | 68.28 ± 0.58 d | 74.87 ± 0.60 b | 18.21 ± 0.62 a | 30.61 ± 0.42 d |
| Solid-liquid ratio | ||||
| 1:5 | 78.15 ± 0.59 a | 70.43 ± 0.27 d | 13.54 ± 0.66 f | 13.12 ± 0.49 e |
| 1:10 | 78.21 ± 0.41 a | 74.24 ± 0.65 c | 15.20 ± 0.32 e | 23.86 ± 0.31 d |
| 1:15 | 74.39 ± 0.56 b | 76.51 ± 0.31 b | 16.72 ± 0.43 d | 32.34 ± 0.33 c |
| 1:20 | 69.00 ± 0.29 c | 76.53 ± 0.46 b | 18.22 ± 0.66 c | 34.53 ± 0.62 b |
| 1:25 | 62.12 ± 1.15 d | 76.67 ± 0.60 b | 20.80 ± 0.37 b | 35.84 ± 0.64 ab |
| 1:30 | 54.10 ± 0.29 e | 84.52 ± 0.38 a | 23.15 ± 0.68 a | 36.29 ± 0.41 a |
| Protease | Loading Amount (U/g) | pH | Enzymolysis Time (h) | Enzymolysis Temperature (℃) | Material/Solvent |
|---|---|---|---|---|---|
| Neutral protease | 4000 | 7.0 | 2 | 50 | 1:15 |
| Flavorzyme | 4000 | 7.0 | 2 | 50 | 1:15 |
| Compound protease | 4000 | 7.0 | 2 | 50 | 1:15 |
| Papain | 4000 | 6.5 | 2 | 55 | 1:15 |
| Trypsin | 4000 | 7.5 | 2 | 45 | 1:15 |
| Alkaline protease | 4000 | 8.0 | 2 | 50 | 1:15 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Z.; Lin, D.; Guo, J.; Zhang, Y.; Zheng, B. In Vitro Antioxidant Activity and In Vivo Anti-Fatigue Effect of Sea Horse (Hippocampus) Peptides. Molecules 2017, 22, 482. https://doi.org/10.3390/molecules22030482
Guo Z, Lin D, Guo J, Zhang Y, Zheng B. In Vitro Antioxidant Activity and In Vivo Anti-Fatigue Effect of Sea Horse (Hippocampus) Peptides. Molecules. 2017; 22(3):482. https://doi.org/10.3390/molecules22030482
Chicago/Turabian StyleGuo, Zebin, Duanquan Lin, Juanjuan Guo, Yi Zhang, and Baodong Zheng. 2017. "In Vitro Antioxidant Activity and In Vivo Anti-Fatigue Effect of Sea Horse (Hippocampus) Peptides" Molecules 22, no. 3: 482. https://doi.org/10.3390/molecules22030482