
Anti-Schistosomal Activity of Cinnamic Acid Esters: Eugenyl and Thymyl Cinnamate Induce Cytoplasmic Vacuoles and Death in Schistosomula of Schistosoma mansoni

Abstract
:1. Introduction
2. Results and Discussion
2.1. Screen Results

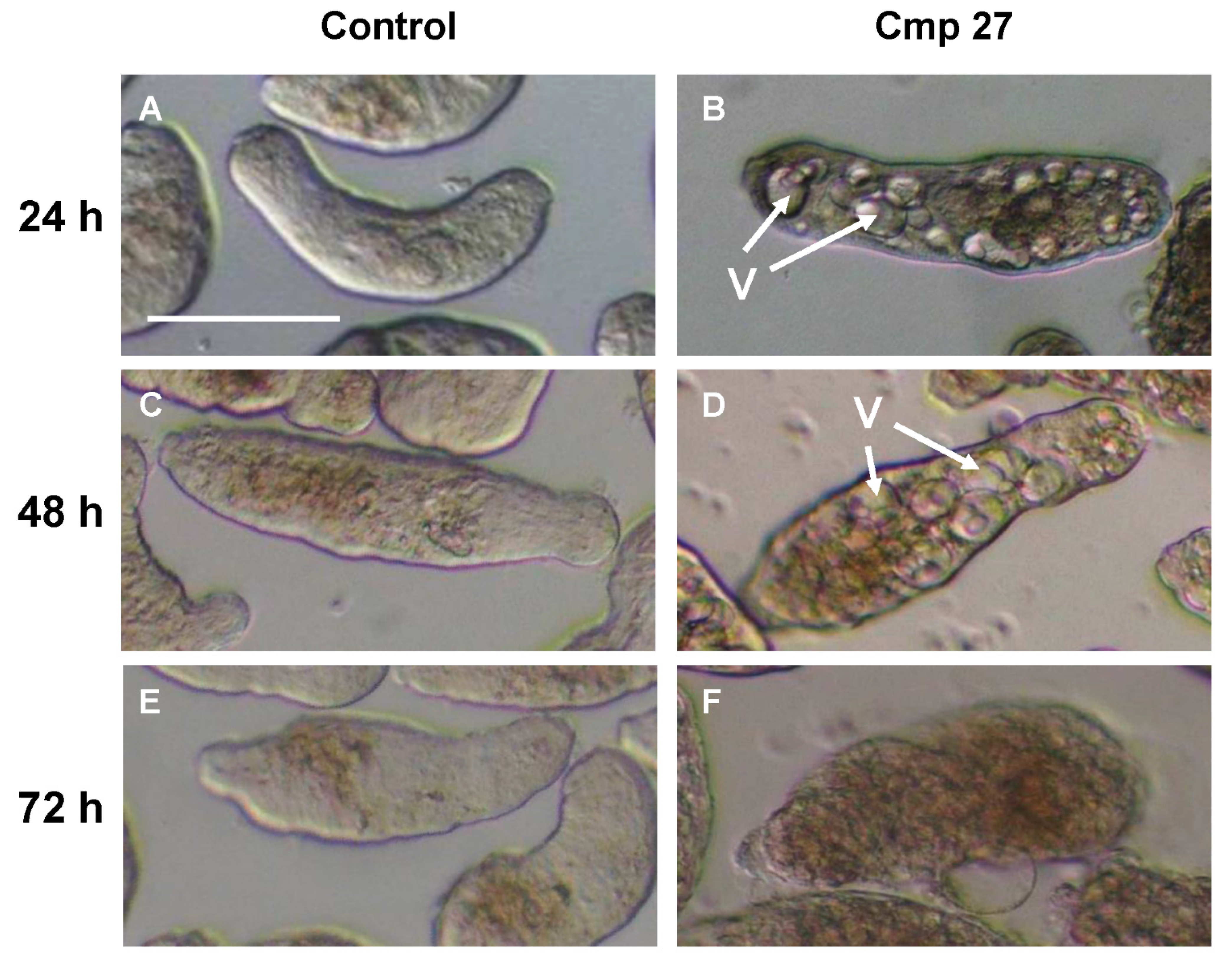{kind=link}
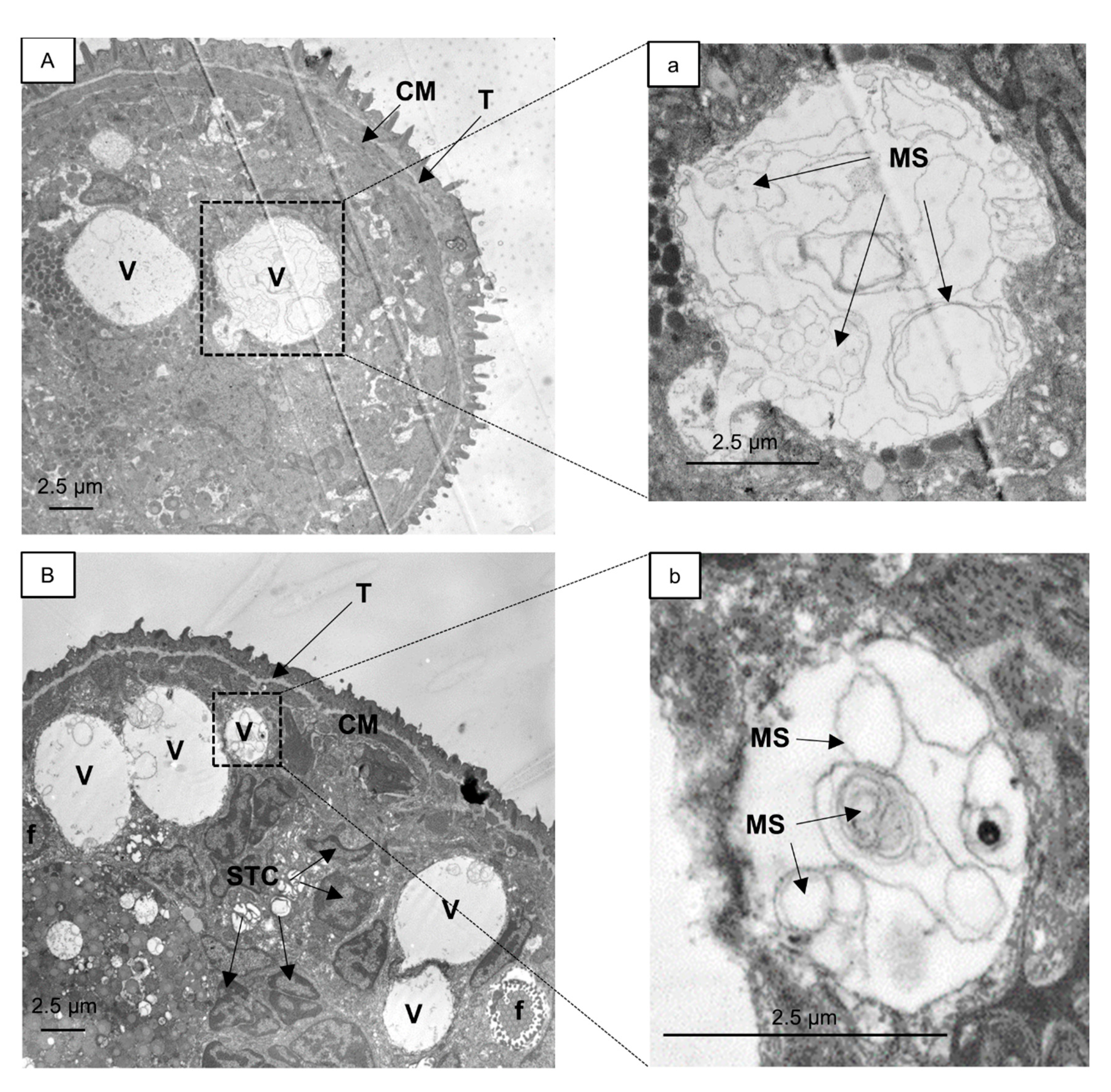{kind=link}
 | Observed Phenotype After (c = 5 μM) | |||||||
|---|---|---|---|---|---|---|---|---|
| Compound | R1 | R2 | R3 | R4 | 24 h | 48 h | 72 h | J774.1 [μM] [15] |
| 1 | -OH | -OH | -H | -H | - | - | - | 8.3 |
| 2 | -OH | -OCH3 | -H | -H | R | R, Dark | R, Dark | 48.7 |
| 3 | -H | -H | -H | -H | - | R, Dark | R, Deg | 45.2 |
| 4 | -H | -H | -Cl | -Cl | R | - | - | 58.9 |
| 5 | -H | -Cl | -H | -Cl | R | - | - | 60.6 |
| 6 | -H | -OCH3 | -H | -H | R | - | - | 42.6 |
| 7 | -H | -Br | -H | -H | R | - | Dark | 54.6 |
| 8 | -H | -N(CH3)2 | -H | -H | R | - | - | >100 |
| 9 | -H | -OBn | -H | -H | R | - | - | >100 |
| 10 | -H | -Cl | -H | -H | - | Dark | - | 49.5 |
| PZQ | - | - | - | - | O, Dark | O, Deg | Deg | - |
 | Observed Phenotype After (c = 5 μM) | ||||||
|---|---|---|---|---|---|---|---|
| Compound | R1 | R2 | R3 | 24 h | 48 h | 72 h | J774.1 [μM] [15] |
| 11 | isoborneol | -OH | -OH | - | - | - | 8.8 |
| 12 | isoborneol | -OH | -OCH3 | - | R | - | 44.3 |
| 13 | isoborneol | -H | -H | - | - | - | 46.0 |
| 14 | isoborneol | -H | -NO2 | - | - | - | >100 |
| 15 | cyclohexanol | -H | -H | - | - | - | >100 |
| 16 | geraniol | -H | -H | - | - | - | >100 |
| 17 | adamantol | -H | -H | - | - | - | 46.7 |
| 18 | 4-thujanol | -H | -H | - | - | - | >100 |
| 19 | naphthol | -H | -H | - | - | - | 44.5 |
| 20 | α-bisabolol | -H | -H | R | R | - | >100 |
| 21 | thymol | -OH | -OH | O | - | - | 8.8 |
| 22 | thymol | -OH | -OCH3 | - | R, Dark | Dark, R | 45.6 |
| 23 | thymol | -H | -H | Deg, V | Deg, V | Deg | 44.8 |
| 24 | menthol | -OH | -OH | - | R, Dark | Dark, R | 2.0 |
| 25 | menthol | -OH | -OCH3 | R | R, Dark | R, Deg | 45.6 |
| 26 | menthol | -H | -H | R | R, Dark | R, Deg | 44.8 |
| 27 | eugenol | -H | -H | Deg, V | Deg | D | 32.2 |
| 28 | thymol | - | - | - | - | - | >100 |
| 29 | eugenol | - | - | - | - | - | >100 |
| 30 | borneol | - | - | - | - | - | >100 |
| PZQ | - | - | - | O, R | O, Deg | Deg | - |
2.2. Analysis of Vacuoles


3. Experimental Section
3.1. General Information
3.2. Synthesis and Cytotoxicity Assays
3.3. Schistosoma Screens and TEM
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Bilharz, T.; Siebold, C.T.E. Ein Beitrag zur Helminthographia Humana; W. Engelmann: Leipzig, Germany, 1852; pp. 1–77. [Google Scholar]
- Chitsulo, L.; Engels, D.; Montresor, A.; Savioli, L. The global status of schistosomiasis and its control. Acta Trop. 2000, 77, 41–51. [Google Scholar] [CrossRef]
- Engels, D.; Chitsulo, L.; Montresor, A.; Savioli, L. The global epidemiological situation of schistosomiasis and new approaches to control and research. Acta Trop. 2002, 82, 139–146. [Google Scholar] [CrossRef]
- Hall, S.C.; Kehoe, E. Case reports. Prolonged survival of Schistosoma japonicum. Calif. Med. 1970, 113, 75–77. [Google Scholar] [PubMed]
- Gryseels, B.; Polman, K.; Clerinx, J.; Kestens, L. Human schistosomiasis. Lancet 2006, 368, 1106–1118. [Google Scholar] [CrossRef]
- Morgan, J.; Dejong, R.; Snyder, S.; Mkoji, G.; Loker, E. Schistosoma mansoni and Biomphalaria: Past history and future trends. Parasitology 2001, 123, 211–228. [Google Scholar] [CrossRef]
- Van der Werf, M.J.; de Vlas, S.J.; Brooker, S.; Looman, C.W.; Nagelkerke, N.J.; Habbema, J.D.F.; Engels, D. Quantification of clinical morbidity associated with schistosome infection in sub-Saharan Africa. Acta Trop. 2003, 86, 125–139. [Google Scholar] [CrossRef]
- Warren, K.S. The pathology, pathobiology and pathogenesis of schistosomiasis. Nature 1978, 273, 609–612. [Google Scholar] [CrossRef] [PubMed]
- Boros, D. Immunopathology of Schistosoma mansoni infection. Clin. Microbiol. Rev. 1989, 2, 250–269. [Google Scholar] [PubMed]
- Smith, J.H.; Christie, J.D. The pathobiology of Schistosoma haematobium infection in humans. Hum. Pathol. 1986, 17, 333–345. [Google Scholar] [CrossRef] [PubMed]
- Gönnert, R.; Andrews, P. Praziquantel, a new broad-spectrum antischistosomal agent. Z. Parasitenkd. 1977, 52, 129–150. [Google Scholar] [CrossRef] [PubMed]
- Fenwick, A.; Savioli, L.; Engels, D.; Robert Bergquist, N.; Todd, M.H. Drugs for the control of parasitic diseases: Current status and development in schistosomiasis. Trends Parasitol. 2003, 19, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Doenhoff, M.J.; Pica-Mattoccia, L. Praziquantel for the treatment of schistosomiasis: Its use for control in areas with endemic disease and prospects for drug resistance. Expert Rev. Anti. Infect. Ther. 2006, 4, 199–210. [Google Scholar] [CrossRef] [PubMed]
- Glaser, J.; Schultheis, M.; Moll, H.; Hazra, B.; Holzgrabe, U. Antileishmanial and cytotoxic compounds from Valeriana wallichii and identification of a novel nepetolactone derivative. Molecules 2015, 20, 5740–5753. [Google Scholar] [CrossRef] [PubMed]
- Glaser, J.; Schultheis, M.; Hazra, S.; Hazra, B.; Moll, H.; Schurigt, U.; Holzgrabe, U. Antileishmanial lead structures from nature: Analysis of structure-activity relationships of a compound library derived from caffeic acid bornyl ester. Molecules 2014, 19, 1394–1410. [Google Scholar] [CrossRef] [PubMed]
- Abdulla, M.H.; Ruelas, D.S.; Wolff, B.; Snedecor, J.; Lim, K.C.; Xu, F.; Renslo, A.R.; Williams, J.; McKerrow, J.H.; Caffrey, C.R. Drug discovery for schistosomiasis: Hit and lead compounds identified in a library of known drugs by medium-throughput phenotypic screening. PLoS Negl. Trop. Dis. 2009. [Google Scholar] [CrossRef] [PubMed]
- Baell, J.; Walters, M.A. Chemistry: Chemical con artists foil drug discovery. Nature News 2014, 513, 481–483. [Google Scholar] [CrossRef] [PubMed]
- Schweigert, N.; Zehnder, A.J.; Eggen, R.I. Chemical properties of catechols and their molecular modes of toxic action in cells, from microorganisms to mammals. Environ. Microbiol. 2001, 3, 81–91. [Google Scholar] [CrossRef] [PubMed]
- Klionsky, D.J.; Abdalla, F.C.; Abeliovich, H.; Abraham, R.T.; Acevedo-Arozena, A.; Adeli, K.; Agholme, L.; Agnello, M.; Agostinis, P.; Aguirre-Ghiso, J.A. Guidelines for the use and interpretation of assays for monitoring autophagy. Autophagy 2012, 8, 445–544. [Google Scholar] [CrossRef] [PubMed]
- Yorimitsu, T.; Klionsky, D.J. Autophagy: Molecular machinery for self-eating. Cell Death Differ. 2005, 12, 1542–1552. [Google Scholar] [CrossRef] [PubMed]
- Dorsey, C.H.; Cousin, C.E.; Lewis, F.A.; Stirewalt, M.A. Ultrastructure of the Schistosoma mansoni cercaria. Micron 2002, 33, 279–323. [Google Scholar] [CrossRef]
- Gutierrez, M.G.; Master, S.S.; Singh, S.B.; Taylor, G.A.; Colombo, M.I.; Deretic, V. Autophagy is a defense mechanism inhibiting BCG and Mycobacterium tuberculosis survival in infected macrophages. Cell 2004, 119, 753–766. [Google Scholar] [CrossRef] [PubMed]
- Deretic, V. Autophagy as an immune defense mechanism. Curr. Opin. Immunol. 2006, 18, 375–382. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.J.; Chen, S.; Huang, K.X.; Le, W.D. Why should autophagic flux be assessed? Acta Pharmacol. Sin. 2013, 34, 595–599. [Google Scholar] [CrossRef] [PubMed]
- Bogitsh, B.J. Cytochemistry of gastrodermal autophagy following starvation in Schistosoma mansoni. J. Parasitol. 1975, 61, 237–248. [Google Scholar] [CrossRef] [PubMed]
- Al-Adhami, B.; Noble, C.; Sharaf, O.; Thornhill, J.; Doenhoff, M.; Kusel, J. The role of acidic organelles in the development of schistosomula of Schistosoma mansoni and their response to signalling molecules. Parasitology 2005, 130, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Carneiro-Santos, P.; Thornhill, J.; Doenhoff, M.; Hagan, P.; Kusel, J. Acidic vesicles of Schistosoma mansoni. Parasitol. Res. 2001, 87, 1001–1006. [Google Scholar] [PubMed]
- Biederbick, A.; Kern, H.; Elsässer, H. Monodansylcadaverine (MDC) is a specific in vivo marker for autophagic vacuoles. Eur. J. Cell Biol. 1995, 66, 3–14. [Google Scholar] [PubMed]
- Clarkson, J.; Erasmus, D. Schistosoma mansoni: An in vivo study of drug-induced autophagy in the gastrodermis. J. Helminthol. 1984, 58, 59–68. [Google Scholar] [CrossRef] [PubMed]
- Rao, A.; Zhang, Y.; Muend, S.; Rao, R. Mechanism of antifungal activity of terpenoid phenols resembles calcium stress and inhibition of the TOR pathway. Antimicrob. Agents Chemother. 2010, 54, 5062–5069. [Google Scholar] [CrossRef] [PubMed]
- Pei, R.; Zhou, F.; Ji, B.; Xu, J. Evaluation of combined antibacterial effects of eugenol, cinnamaldehyde, thymol, and carvacrol against E. coli with an improved method. J. Food Sci. 2009, 74, M379–M383. [Google Scholar] [CrossRef] [PubMed]
- Didry, N.; Dubreuil, L.; Pinkas, M. Activity of thymol, carvacrol, cinnamaldehyde and eugenol on oral bacteria. Pharm. Acta Helv. 1994, 69, 25–28. [Google Scholar] [CrossRef]
- Dai, J.P.; Zhao, X.F.; Zeng, J.; Wan, Q.Y.; Yang, J.C.; Li, W.Z.; Chen, X.X.; Wang, G.F.; Li, K.S. Drug screening for autophagy inhibitors based on the dissociation of beclin1-bcl2 complex using BiFC technique and mechanism of eugenol on anti-influenza A virus activity. PLoS ONE 2013. [Google Scholar] [CrossRef] [PubMed]
- NCBI Protein Database. Available online: http://www.ncbi.nlm.nih.gov/protein/CAJ19743.1 (accessed on 5 May 2015).
- Bringmann, G.; Thomale, K.; Bischof, S.; Schneider, C.; Schultheis, M.; Schwarz, T.; Moll, H.; Schurigt, U. A novel Leishmania major amastigote assay in 96-well format for rapid drug screening and its use for discovery and evaluation of a new class of leishmanicidal quinolinium salts. Antimicrob. Agents Chemother. 2013, 57, 3003–3011. [Google Scholar] [CrossRef] [PubMed]
- Sample Availability: Samples of all synthesized compounds are available from the authors.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaser, J.; Schurigt, U.; Suzuki, B.M.; Caffrey, C.R.; Holzgrabe, U. Anti-Schistosomal Activity of Cinnamic Acid Esters: Eugenyl and Thymyl Cinnamate Induce Cytoplasmic Vacuoles and Death in Schistosomula of Schistosoma mansoni. Molecules 2015, 20, 10873-10883. https://doi.org/10.3390/molecules200610873
Glaser J, Schurigt U, Suzuki BM, Caffrey CR, Holzgrabe U. Anti-Schistosomal Activity of Cinnamic Acid Esters: Eugenyl and Thymyl Cinnamate Induce Cytoplasmic Vacuoles and Death in Schistosomula of Schistosoma mansoni. Molecules. 2015; 20(6):10873-10883. https://doi.org/10.3390/molecules200610873
Chicago/Turabian StyleGlaser, Jan, Uta Schurigt, Brian M. Suzuki, Conor R. Caffrey, and Ulrike Holzgrabe. 2015. "Anti-Schistosomal Activity of Cinnamic Acid Esters: Eugenyl and Thymyl Cinnamate Induce Cytoplasmic Vacuoles and Death in Schistosomula of Schistosoma mansoni" Molecules 20, no. 6: 10873-10883. https://doi.org/10.3390/molecules200610873