
Effects of Dietary Brazilian Palm Oil (Mauritia flexuosa L.) on Cholesterol Profile and Vitamin A and E Status of Rats

Abstract
:
1. Introduction
2. Results and Discussion
2.1. Assessment of Diet Intake, Weight Gain and Feed Efficiency Coefficient (FEC)

{kind=link}
{kind=link}
{kind=link}
| Variables | Animal Groups | ||
|---|---|---|---|
| CG (n = 10) | CBO (n = 10) | RBO (n = 10 ) | |
| Mass gain (g) | 104.98 ± 14.53 a | 91.37 ± 13.41 a | 110.36 ± 18.16 a |
| Diet intake (g) | 336.9 ± 36.72 a | 344.25 ± 25.67 a | 336.55 ± 21.04 a |
| FEC | 0.30 ± 0.07 ab | 0.26 ± 0.03 b | 0.34 ± 0.07 a |
2.2. Murine Anthropometric Parameters
| Murine Antropometric Data | Animal Groups | ||
|---|---|---|---|
| CG (n = 10) | CBO (n = 10) | RBO (n = 10 ) | |
| TC (cm) | 10.55 ± 1.23 a | 11.14 ± 0.45 a | 10.9 ± 0.46 a |
| AC (cm) | 10.86 ± 1.43 b | 12.41 ± 1.07 a | 12.25 ± 0.75 a |
| AC/TC | 1.03 ± 0.05 a | 1.11 ± 0.03 a | 1.12 ± 0.05 a |
| BL (cm) | 18.27 ± 1.42 a | 18.09 ± 1.04 a | 18.40 ± 0.97 a |
| BW (g) | 136.16 ± 31.38 a | 146.20 ± 20.09 a | 146.23 ± 24.05 a |
| BMI (g·cm−2) | 0.40 ± 0.05 a | 0.45 ± 0.04 a | 0.43 ± 0.04 a |
| Lee index | 0.28 ± 0.01 a | 0.29 ± 0.01 a | 0.26 ± 0.09 a |
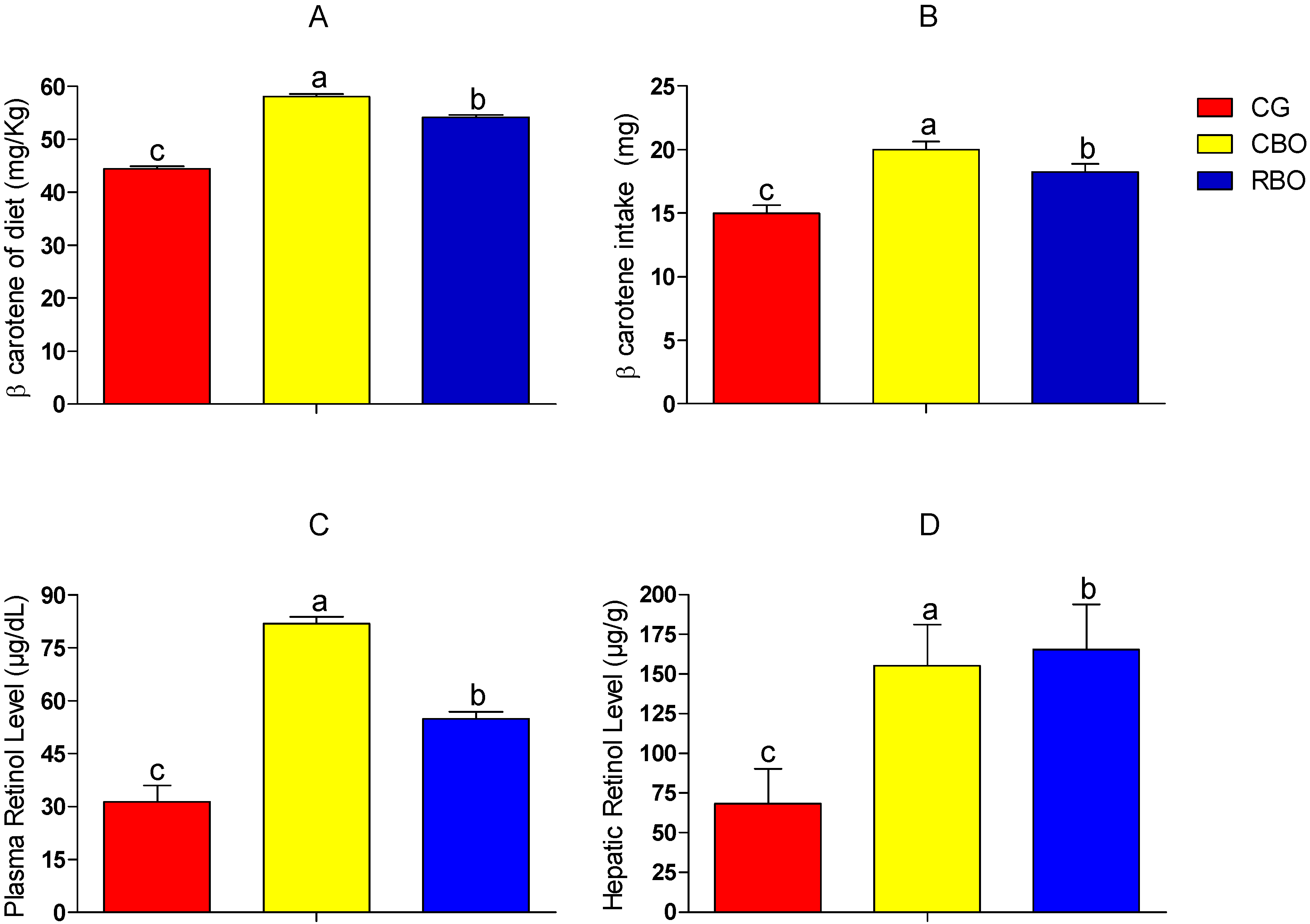
2.3. β-Carotene and Alpha Tocopherol in Diet, Blood and Liver of Rats


2.4. Lipid and Haematological Profiles
| Biochemical Data | Animals Groups | ||
|---|---|---|---|
| CG (n = 10) | CBO (n = 10) | RBO (n = 10 ) | |
| Hemoglobin (g/dL) | 10.55 ± 1.23 a | 11.14 ± 0.45 a | 10.9 ± 0.46 a |
| Hematocrit (%) | 41.38 ± 8.4 ab | 48.25 ± 3.11 a | 35.25 ± 10.24 b |
| TC (mg/dL) | 96.83 ± 13.97 b | 131.51 ± 15.52 a | 52.25 ± 6.88 c |
| HDL-C (mg/dL) | 24.38 ± 11.6 a | 23.64 ± 2.04 a | 13.41 ± 1.59 b |
| TC/HDL-C (mg/dL) | 3.91 ± 0.64 b | 5.44 ± 0.46 a | 3.96 ± 0.31 b |
| VLDL-C (mg/dL) | 16.18 ± 6.36 a | 19.88 ± 2.35 a | 8.86 ± 0.86 b |
| LDL-C (mg/dL) | 57.88 ± 3.86 b | 87.49 ± 5.28 a | 30.84 ± 5.24 c |
| TAG (mg/dL) | 81.50 ± 32.33 a | 99.38 ± 11.75 a | 44.25 ± 4.68 b |
| AST (IU/L) | 10.75 ± 1.58 ab | 12.75 ± 1.91 a | 10.00 ± 2.78 b |
| ALT (IU/L) | 12.13 ± 1.81 a | 14.38 ± 2.00 a | 11.75 ± 2.55 a |
3. Experimental Section
3.1. Oil Samples
| Composition | Vegetable Oils | ||
|---|---|---|---|
| Soybean Oil * | Crude Buriti Oil | Refined Buriti Oil | |
| β-Carotene (mg·kg−1) | 420.02 ± 2.80 c | 911.40 ± 2.40 a | 792.10 ± 4.54 b** |
| Tocopherol (mg·kg−1) | 305.20 ± 2.10 c | 810.00 ± 2.70 a | 709.00 ± 2.40 b*** |
| Fatty Acids (%) | |||
| Miristic acid-C 14:0 | 17.10 ± 0.02 a | 0.50 ± 0.04 b | 0.50 ± 0.02 c |
| Margaric acid-C 17:0 | 11.20 ± 0.03 b | 0.30 ± 0.02 a | 0.20 ± 0.01 c |
| Stearic acid-C18:0 | 3.40 ± 0.03 a | 2.30 ± 0.02 b | 3.90± 0.01 c |
| Total saturated fatty acids—SFA% | 31.70 ± 0.08 a | 3.10 ± 0.08 c | 4.6 ± 0.04 a |
| Palmitoleic acid-C16:1 | n.d. **** | 19.60 ± 0.02 | 19.40 ± 0.01 |
| Oleic acid-C18:1 | 23.60 ± 0,04 b | 72.70 ± 0.02 a | 72.20 ± 0.02 c |
| Total monounsaturated fatty acids—MUFA% | 23.60 ± 0.04 b | 92.30 ± 0.04 a | 91.60 ± 0.03 c |
| Linoleic acid-C 18:2 | 39.90 ± 0.02 a | 2.60 ± 0.04 b | 2.30 ± 0.01 c |
| Linolenic acid-C 18:3 | 4.80 ± 0.02 a | 2.00 ± 0.01 b | 1.50 ± 0.01 c |
| Total polyunsaturated fatty acids—PUFA% | 44.70 ± 0.04 ª | 4.60 ± 0.05 b | 3.80 ± 0.01 c |
3.2. Animals and Diet
| Ingredients * | Amounts (g/100 g) | CS | CBO | RBO |
|---|---|---|---|---|
| Energy (Kcal) | Energy(Kcal) | Energy(Kcal) | ||
| Corn starch | 52.9 | 186.10 | 186.10 | 186.10 |
| Casein | 20.0 | 68.00 | 68.0 | 68.0 |
| Sucrose | 10.0 | 40.00 | 40.00 | 40.0 |
| Fiber | 5.0 | - | - | - |
| Soybean oil or buriti oil | 7.0 | 63.0 | 63.0 | 63.0 |
| Mix of minerals | 3.5 | - | - | - |
| Mix of vitamins | 1.0 | - | - | - |
| D.L-Methionine | 0.3 | - | - | - |
| Choline Bitartrate | 0.3 | - | - | - |
| Total | 100 | 357.10 | 357.10 | 357.10 |
3.3. Murine Anthropometric Parameters
3.4. Determination of Vitamins A and E in Diets and Intake of β-Carotene and Alpha Tocopherol by Rats
3.5. Determination of Serum and Hepatic Retinol and Alpha Tocopherol
3.6. Lipid and Haematological Profiles
3.7. Statistical Analyses
4. Conclusions
Acknowledgments
Author Contributions
Abbreviations
| AC | Abdominal circumference |
| AIN | American Institute of Nutrition |
| ALT | Alanine transaminase |
| AST | Aspartate transaminase |
| BMI | Body Mass Index |
| BHT | Butylated Hydroxytoluene |
| CBO | Experimental group that consumed diet added of casein and crude buriti oil |
| CG | Control group that consumed diet added of casein and soybean oil |
| CC | Chest circumference |
| CEPA | Ethics Committee on Animal Research |
| COBEA | Brazilian College of Animal Experimentation |
| DAD | Diode Array Detector |
| DRIs | Dietary Reference Intakes |
| FEC | Feed Efficiency Coefficient |
| HDL-C | High density lipoprotein |
| HPLC | High Performance Liquid Chromatography |
| IU | International Unit (vitamins) |
| LI | Lee index |
| LDL-C | Low density lipoprotein |
| RBO | Experimental group that consumed diet containing casein and refined buriti oil |
| RE | Retinol equivalents |
| TAG | triglycerides |
| UFPE | Federal University of Pernambuco |
| VLDL-C | Very low density lipoprotein |
Conflicts of Interest
References
- Wawrzyniaka, A.; Górnicka, M.; Hamułka, J.; Gajewska, M.; Drywieńa, M.; Pierzynowska, J.; Gronowska-Senger, A. α-Tocopherol, ascorbic acid, and β-carotene protect against oxidative stress but reveal no direct influence on p53 expression in rats subjected to stress. Nutr. Res. 2013, 33, 868–875. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Komaru, T.; Takeda, S.; Takeda, M.; Koshida, R.; Nakayama, M.; Kokusho, Y.; Kawakami, Y.; Yamaguchi, N.; Miyazawa, T.; et al. Gamma-tocopherol, but not α-tocopherol, potently inhibits neointimal formation induced by vascular injury in insulin resistant rats. J. Mol. Cell. Cardiol. 2006, 41, 544–554. [Google Scholar] [CrossRef] [PubMed]
- Canas, J.A.; Damaso, L.; Altomare, A.; Killen, K.; Hossain, J.; Balagopal, B.P. Insulin resistance and adiposity in relation to serum β-carotene levels. J. Pediatr. 2012, 161, 58–64. [Google Scholar] [CrossRef] [PubMed]
- López-Miranda, J.; Pérez-Jiménez, F.; Ros, E.; De Caterina, R.; Badimón, L.; Covas, M.I.; Escrich, E.; Ordovás, J.M.; Sorique, F.; Abiá, R.; et al. Olive oil and health: Summary of the II international conference on olive oil and health consensus report, Jaén and Córdoba (Spain) 2008. Nutr. Metab. Cardiovasc. Dis. 2010, 20, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Widmer, R.J.; Flammer, A.J.; Lermanl, O.; Lerman, A. The Mediterranean diet, its components and cardiovascular disease. Am. J. Med. 2015, 128, 229–238. [Google Scholar] [CrossRef] [PubMed]
- Gunstone, F.D. Vegetable Oils in Food Technology: Composition, Properties and Uses; Blackwell Publishing: Oxford, UK, 2002. [Google Scholar]
- Aquino, J.S.; Pontes Pessoa, D.C.N.; Araújo, K.L.G.V.; Epaminondas, P.S.; Schuler, A.R.P.; Souza, A.G.; Stamford, T.L.M. Refining of Buriti oil (Mauritia flexuosa) originated from the Brazilian Cerrado: Physicochemical, thermal-oxidative and nutritional implications. J. Braz. Chem. Soc. 2012, 23, 2012–2019. [Google Scholar]
- Szydłowska-Czerniaka, A.; Trokowski, K.; Karlovits, G.; Szłyk, E. Effect of refining processes on antioxidant capacity, total contents of phenolics and carotenoids in palm oils. Food Chem. 2011, 129, 1187–1192. [Google Scholar] [CrossRef] [PubMed]
- Landucci, G.; Pannocchia, G.; Pelagagge, L.; Nicolella, C. Analysis and simulation of an industrial vegetable oil refining process. J. Food Eng. 2013, 116, 840–851. [Google Scholar] [CrossRef]
- Oterhals, A.; Vogt, G. Impact of Extraction, Refining and Concentration Stages on the Stability of Fish Oil. Food Enrichment with Omega-3 Fatty Acids; Woodhead Publishing: Cambridge, UK, 2013. [Google Scholar]
- Asadi, F.; Shahriari, A.; Chahardah-Cheric, M. Effect of long-term optional ingestion of canola oil, grape seed oil, corn oil and yogurt butter on serum, muscle and liver cholesterol status in rats. Food Chem. Toxicol. 2010, 48, 2454–2457. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Qianchun, D.; Jiqu, X.; Fenghong, H.; Qingde, H.; Zhihua, Y.; Jine, Y. Effects of cold-pressed and vitamin E-enriched flaxseed oils on lipid profile and antioxidant status in high-fat fed rats. Eur. J. Lipid Sci. Technol. 2012, 114, 461–468. [Google Scholar] [CrossRef]
- Dhavamani, S.; Rao, Y.P.C.; Lokesh, B.R. Total antioxidant activity of selected vegetable oils and their influence on total antioxidant values in vivo: A photochemiluminescence based analysis. Food Chem. 2014, 164, 551–555. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, B.D.; Coelho, M.A.Z.; Barreto, D.W. Production of concentrated beta-carotene from buriti (Mauritia vinífera) oil by enzymatic hydrolysis. Food Bioprod. Process 2012, 90, 141–147. [Google Scholar] [CrossRef]
- Ribeiro, B.D.; Nascimento, R.F.; Barreto, D.W.; Coelho, M.A.S.; Freitas, S.P. An ethanol-based process to simultaneously extract and fractionate carotenoids from Mauritia flexuosa pulp. Rev. Bras. Frutic. 2010, 32, 657–663. [Google Scholar] [CrossRef]
- Albuquerque, M.L.S.; Guedes, I.; Alcantara Junior, P.; Moreira, S.G.C.; Barbosa Neto, N.M.; Correa, D.S.; Zilio, S.C. Characterization of Buriti (Mauritia flexuosa L.) Oil by Absorption and Emission Spectroscopies. J. Braz. Chem. Soc. 2005, 16, 1113–1117. [Google Scholar] [CrossRef]
- Berger, A.; Monnard, I.; Dionisi, F.; Gumy, D.; Hayes, KC.; Lambelet, P. Cholesterol-lowering properties of amaranth flakes, crude and refined oils in hamsters. Food Chem. 2003, 81, 119–124. [Google Scholar] [CrossRef]
- Hama, S.; Yoshida, A.; Tamadani, N.; Noda, H.A.; Kondo, A. Enzymatic production of biodiesel from waste cooking oil in a packed-bed reactor: An engineering approach to separation of hydrophilic impurities. Bioresour. Technol. 2013, 135, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Novelli, L.B.; Diniz, Y.S.; Galhardi, C.M.; Ebaid, G.M.X.; Rodrigues, H.G.; Mani, F.; Fernandes, A.A.; Cicogna, A.C.; Novelli Filho, J.L.V.B. Anthropometrical parameters and markers of obesity in rats. Lab. Anim. 2007, 41, 111–119. [Google Scholar] [CrossRef] [PubMed]
- Santillán, M.E.; Vincenti, L.M.; Martini, A.C.; de Cuneo, M.F.; Ruiz, R.D.; Mangeaud, A.; Stutz, G. Developmental and neurobehavioral effects of perinatal exposure to diets with different ω-6:ω-3 ratios in mice. Nutrition 2010, 26, 423–431. [Google Scholar] [CrossRef] [PubMed]
- Ramos, M.I.L.; Siqueira, E.M.; Isomura, C.C.; Barbosa, A.M.; Arruda, S.F. Bocaiuva (Acrocomia aculeata (Jacq) Lodd) improved vitamin A status in rats. J. Agric. Food Chem. 2007, 22, 3186–3190. [Google Scholar] [CrossRef]
- Siqueira, E.M.A.; Arruda, S.F.; Vargas, R.M.; Souza, E.M.T. β-Carotene from cassava (Manihot esculenta Crantz) leaves improves vitamin A status in rats. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2007, 146, 235–240. [Google Scholar] [CrossRef] [PubMed]
- FAO/WHO. Human Vitamin and Mineral Requirements. In Proceedings of the 7th Joint FAO/OMS Expert Consultation, Bangkok, Thailand, 30 September 2001; p. 286.
- Institute of Medicine, Food and Nutrition Board. Dietary Reference Intakes for Vitamins C, E, Selenium and Carotenoids; National Academy Press: Washington, WA, USA, 1998. [Google Scholar]
- Yuyama, L.K.O.; Yonekura, L.; Aguiar, J.P.L.; Sousa, R.F.S. Biodisponibilidade dos carotenoides do Buriti (Mauritia flexuosa L.) em ratos. Acta Amaz. 1998, 28, 409–415. [Google Scholar]
- Rossi, M.; Alamprese, C.; Ratti, S. Tocopherols and tocotrienols as free radical-scavengers in refined vegetable oils and their stability during deep-fat frying. Food Chem. 2007, 102, 812–817. [Google Scholar] [CrossRef]
- Domitrovic, R.; Tota, M.; Milin, C. Differential effect of high dietary iron on α-tocopherol and retinol levels in the liver and serum of mice fed olive oil- and corn oil-enriched diets. Nutr. Res. 2008, 28, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Naziroglu, M.; Brandsch, C. Dietary hydrogenated soybean oil affects lipid and vitamin E metabolism in rats. J. Nutr. Sci. Vitaminol. 2006, 52, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Luostarinen, R.L.; Laasonen, K.; Calder, P.C. α-Tocopherol concentrations, lipid peroxidation and superoxide dismutase and glutathione peroxidase activities in rat heart and liver after feeding stabilized and unstabilized fish oil. Nutr. Res. 2001, 21, 1529–1544. [Google Scholar] [CrossRef]
- Odabasoglu, F.; Halici, Z.; Cakir, A.; Halici, M.; Aygun, H.; Suleyman, H.; Cadirci, E.; Atalay, F. Beneficial effects of vegetable oils (corn, olive and sunflower oils) and tocopherol on anti-inflammatory and gastrointestinal profiles of indomethacin in rats. Eur. J. Pharmacol. 2008, 591, 300–306. [Google Scholar] [CrossRef] [PubMed]
- Fiedor, J.; Burda, K. Potential Role of Carotenoids as Antioxidants in Human Health and Disease. Nutrients 2014, 6, 466–488. [Google Scholar] [CrossRef] [PubMed]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef] [PubMed]
- Denayer, T.; Stöhr, T.; van Roy, M. Animal models in translational medicine: Validation and prediction. New Horiz. Transl. Med. 2014, 2, 5–11. [Google Scholar] [CrossRef]
- Groenink, L.; Folkerts, G.; Schuurman, H.J. European Journal of Pharmacology, Special issue on translational value of animal models: Introduction. Eur. J. Pharmacol. 2015, 759, 1–2. [Google Scholar] [CrossRef] [PubMed]
- García-Casal, M.N. Carotenoids increase iron absorption from cereal-based food in the human. Nutr. Res. 2006, 26, 340–344. [Google Scholar] [CrossRef]
- Tzang, B.-S.; Yang, S.-F.; Fu, S.-G.; Yang, H.-C.; Sun, H.-L.; Chen, Y.-C. Effects of dietary flaxseed oil on cholesterol metabolism of hamsters. Food Chem. 2009, 114, 1450–1455. [Google Scholar] [CrossRef]
- Quiles, J.L.; Ocho, J.J.; Ramirez-Torotsa, C.; Bttino, M.; Huertas, J.R.; Martín, Y.; Mataix, J. Dietary fat type (virgin olive vs. sunflower oils) affects age-related changes in DNA double-strand-breaks, antioxidant capacity and blood lipids in rats. Exp. Geront. 2004, 39, 1189–1198. [Google Scholar] [CrossRef]
- Jacomelli, M.; Pitozzi, V.; Zaid, M.; Larrosa, M.; Toninib, G.; Martini, A.; Urbani, S.; Taticchi, A.; Servili, M.; Dolara, P.; et al. Dietary extra-virgin olive oil rich in phenolic antioxidants and the aging process: Long-term effects in the rat. J. Nutr. Biochem. 2010, 21, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Condelli, N.; Caruso, M.C.; Galgano, F.; Russo, D.; Milella, L.; Favati, F. Prediction of the antioxidant activity of extra virgin olive oils produced in the Mediterranean area. Food Chem. 2015, 177, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Arbonés-Mainar, J.M.; Navarro, M.A.; Carnicer, R.; Guillén, N.; Surra, J.C.; Acín, S.; Guzmán, M.A.; Sarría, A.J.; Arnal, C.; Aguilera, M.P.; et al. Accelerated atherosclerosis in apolipoprotein E-deficient mice fed Western diets containing palm oil compared with extra virgin olive oils: A role for small, dense high-density lipoproteins. Atherosclerosis 2007, 194, 372–382. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, T.; Shimoda, K.; Horiguchi, S.; Domon, M.; Hagiwara, C.; Takiyama, M.; Kagawa, Y. Influences of stearidonic acid-enriched soybean oil on the blood and organ biochemical parameters in rats. Prostaglandins Leukot. Essent. Fatty Acids 2013, 88, 179–184. [Google Scholar] [CrossRef] [PubMed]
- Ohara, N.; Naito, Y.; Kasama, K.; Shindo, T.; Yoshida, H.; Nagata, T.; Okuyama, H. Similar changes in clinical and pathological parameters in Wistar Kyoto rats after a 13-week dietary intake of canola oil or a fatty acid composition-based interesterified canola oil mimic. Food Chem. Toxicol. 2009, 47, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Rueda-Clausen, C.F.; Silva, F.A.; Lindarte, M.A.; Villa-Roel, C.; Gomez, E.; Guitierrez, R.; Cure-Cure, C.; López-Jaramillo, P. Olive, soybean and palm oils intake have a similar acute detrimental effect over the endothelial function in healthy young subjects. Nutr. Metab. Cardiovas. 2007, 1, 50–57. [Google Scholar] [CrossRef]
- Maki, K.C.; Lawlwss, A.L.; Kelley, K.M.; Kaden, V.N.; Geiger, C.J.; Dicklin, M.R. Corn oil improves the plasma lipoprotein lipid profile compared with extra-virgin olive oil consumption in men and women with elevated cholesterol: Results from a randomized controlled feeding trial. J. Clin. Lipidol. 2015, 9, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Ng, C.-Y.; Leong, X.-F.; Namasbah, N.; Adam, S.K.; Kamisah, Y.; Jaarin, K. Heated vegetable oils and cardiovascular disease risk factors. Vascul. Pharmacol. 2014, 61, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yeh, Y.-H.; Lee, Y.-T.; Hsieh, H.S.; Hwang, D.-F. Effect of red yeast rice on toxicity of oxidized cholesterol and oxidized fish oil in rats. Eur. J. Clin. Nutr. Metab. 2010, 5, 230–237. [Google Scholar] [CrossRef]
- Sulli, K.C.; Sun, J.; Giraud, D.W.; Moxley, R.A.; Driskell, J.A. Effects of β-carotene and α-tocopherol on the levels of tissue cholesterol and triglyceride in hypercholesterolemic rabbits. J. Nutr. Biochem. 1998, 9, 344–350. [Google Scholar] [CrossRef]
- Kaulmann, A.; Bohn, T. Carotenoids, inflammation, and oxidative stress—Implications of cellular signaling pathways and relation to chronic disease prevention. Nutr. Res. 2014, 34, 907–929. [Google Scholar] [CrossRef] [PubMed]
- Plaa, G.L. Evaluation of hepatotoxicity: Physiological and biochemical measures of hepatic function in animals. Comprehensive Toxicol. 2010, 9, 129–140. [Google Scholar]
- Sugiura, M.; Nakamura, M.; Ikoma, Y.; Yano, M.; Ogawa, K.; Matsumoto, K.; Kato, M.; Ohshima, M.; Nagao, A. Serum carotenoid concentrations are inversely associated with serum aminotransferases in hyperglycemic subjects. Diabetes Res. Clin. Pract. 2006, 71, 82–91. [Google Scholar] [CrossRef] [PubMed]
- Association Official Analytical Chemistis. Official Methods of Analysis of the Association Chemistis, 17th ed.; Association Official Analytical Chemistis: Washington, WA, USA, 2002; pp. 10–12. [Google Scholar]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C. AIN-93: Purified diets for laboratory rodents: final report of the American Institute of Nutrition and hoe writing comitee on the reformulation of the American Institute of Nutrition (AIN)-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [PubMed]
- Campbell, J.A. Method for determination of PER and NPR. In Food and Nutrition Board. Evaluation of Protein Quality; Committee on Protein Quality: Washington, WA, USA, 1963; pp. 31–32. [Google Scholar]
- Prado, J.P.S.; Cavalheiro, J.M.O.; Silva, F.V.G.; Queiroga Neto, V.; Bora, P.S.; Cavalheiro, T.B. Thermal stability of vitamins A and E in rations and vitamin premixes for shrimp. Cien. Rural 2011, 41, 544–549. [Google Scholar] [CrossRef]
- Furr, H.C.; Tranuminardjo, S.A.; Olson, J.A. Training Manual for Assessing Vitamin a Status by Use of the Modified Relative Dose Response and the Relative Dose Response Assays; Office of Nutrition Bureau for Research and Development: Washington, WA, USA, 1992. [Google Scholar]
- Stahl, W.; Sies, H. Antioxidant activity of carotenoids. Mol. Asp. Med. 2003, 24, 345–351. [Google Scholar] [CrossRef]
- SPSS for Windows Evaluation Version 14.0 [Computer program]; LEAD Technologies SPSS Inc.: Chicago, IL, USA, 2005.
- Sample Availability: Not available.
© 2015 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aquino, J.D.S.; Soares, J.K.B.; Magnani, M.; Stamford, T.C.M.; Mascarenhas, R.D.J.; Tavares, R.L.; Stamford, T.L.M. Effects of Dietary Brazilian Palm Oil (Mauritia flexuosa L.) on Cholesterol Profile and Vitamin A and E Status of Rats. Molecules 2015, 20, 9054-9070. https://doi.org/10.3390/molecules20059054
Aquino JDS, Soares JKB, Magnani M, Stamford TCM, Mascarenhas RDJ, Tavares RL, Stamford TLM. Effects of Dietary Brazilian Palm Oil (Mauritia flexuosa L.) on Cholesterol Profile and Vitamin A and E Status of Rats. Molecules. 2015; 20(5):9054-9070. https://doi.org/10.3390/molecules20059054
Chicago/Turabian StyleAquino, Jailane De Souza, Juliana Késsia Barbosa Soares, Marciane Magnani, Thayza Christina Montenegro Stamford, Robson De Jesus Mascarenhas, Renata Leite Tavares, and Tânia Lúcia Montenegro Stamford. 2015. "Effects of Dietary Brazilian Palm Oil (Mauritia flexuosa L.) on Cholesterol Profile and Vitamin A and E Status of Rats" Molecules 20, no. 5: 9054-9070. https://doi.org/10.3390/molecules20059054