Effects of Thai Medicinal Herb Extracts with Anti-Psoriatic Activity on the Expression on NF-κB Signaling Biomarkers in HaCaT Keratinocytes



Abstract
:1. Introduction
2. Results and Discussion
2.1. Effect of Thai Herbal Extracts on NF-κB Signaling Network Biomarkers Expression in HaCaT Cells



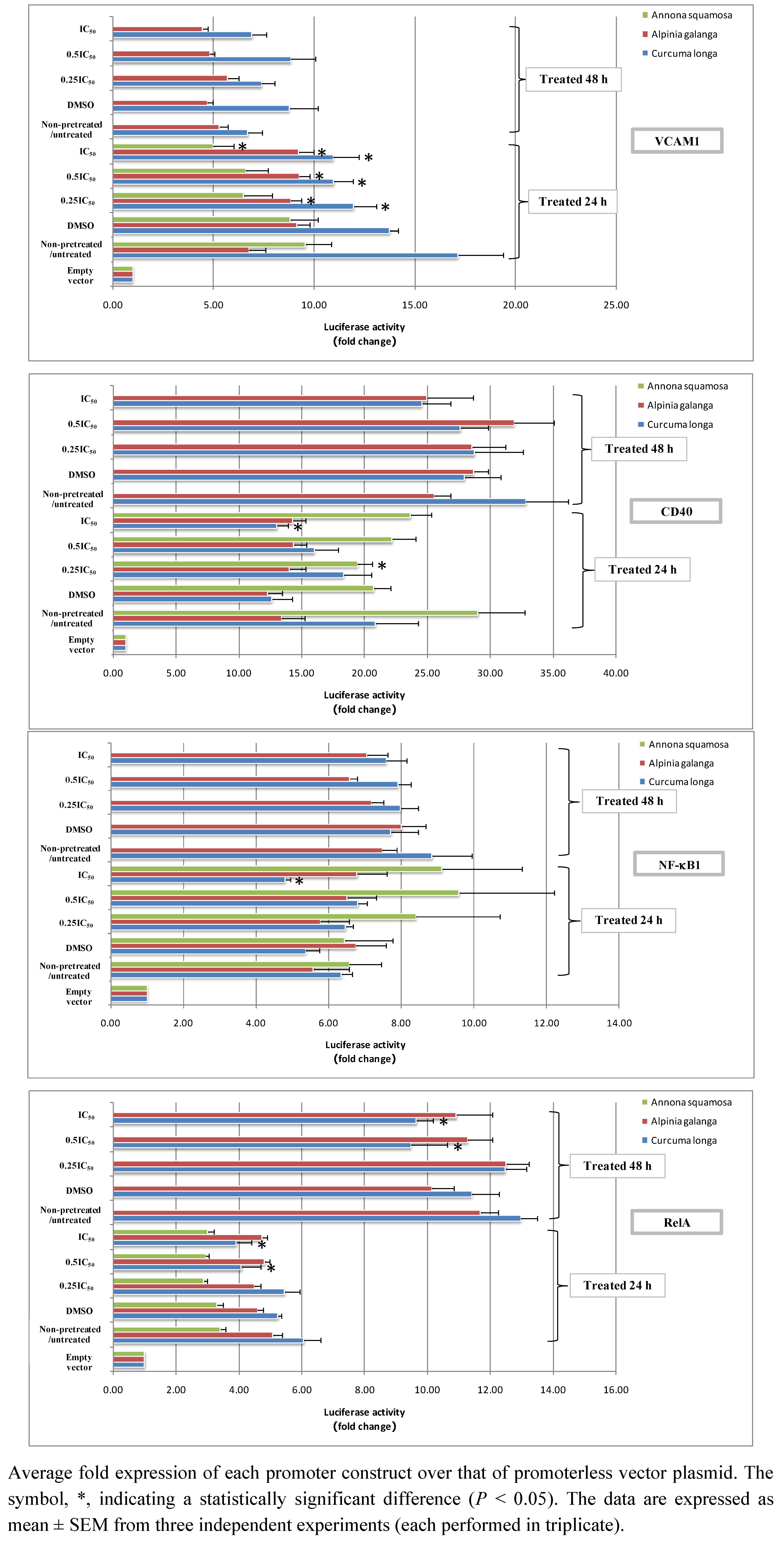
2.2. Effect of Thai Medicinal Herb Extracts on Functional Promoters of NF-κB Signaling Network Biomarkers in HaCaT Cells


3. Experimental
3.1. Plant Materials and Preparation of Herbal Extracts [16]
3.2. Cell Culture
3.3. Treatment of HaCaT Cells Using Thai Medicinal Herb Extracts
3.4. Semi-quantitative Reverse Transcriptase-polymerase Chain Reaction (RT-PCR)

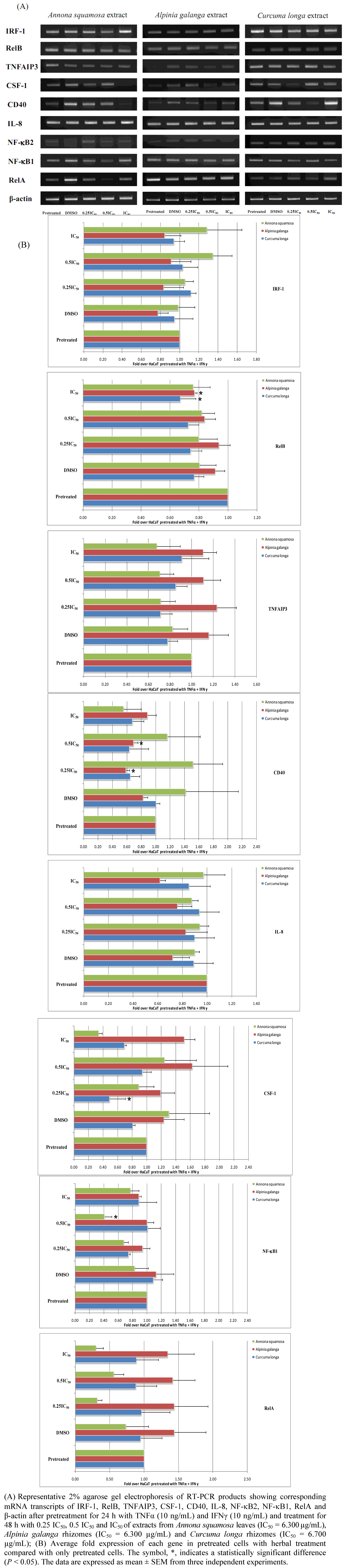{kind=link}
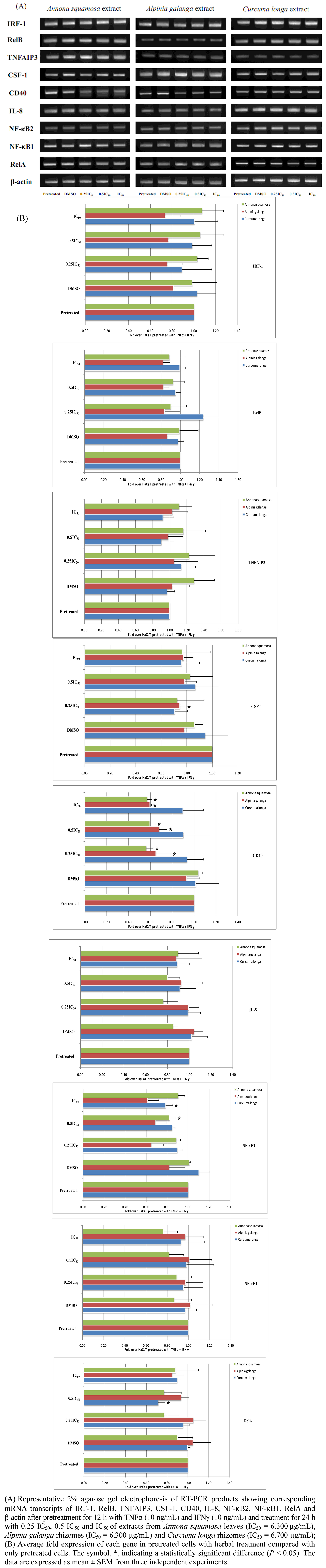{kind=link}
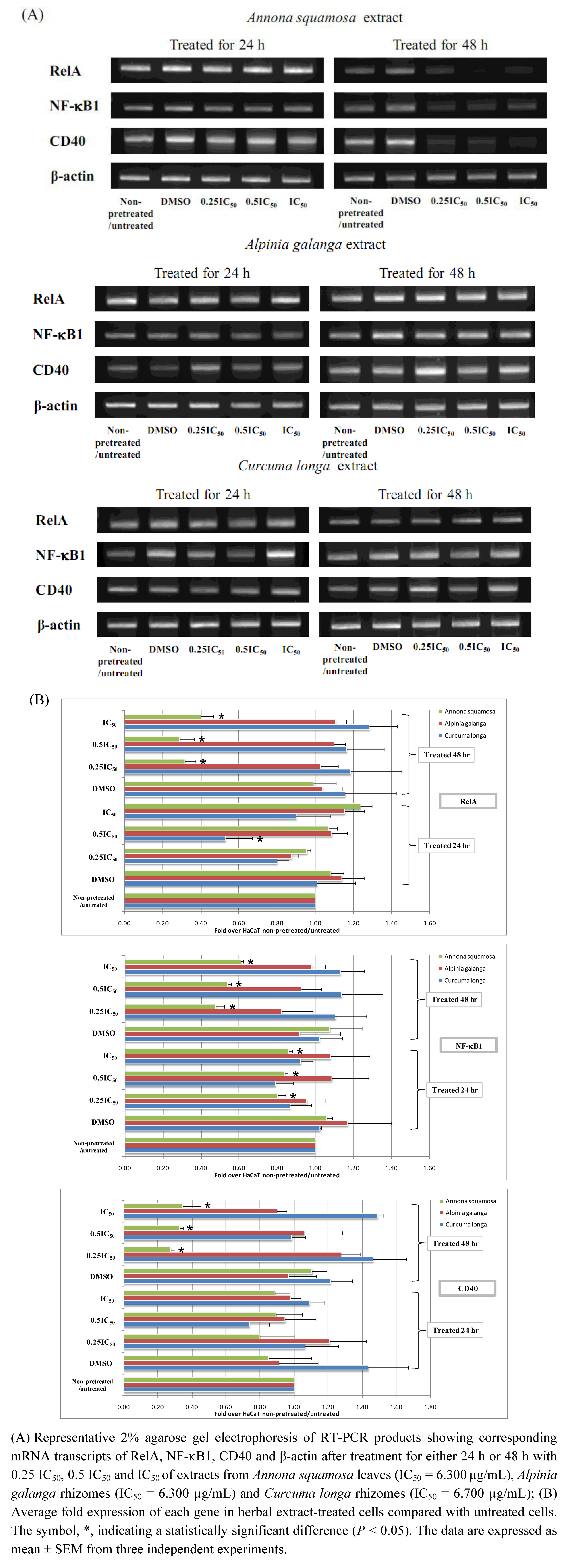{kind=link}
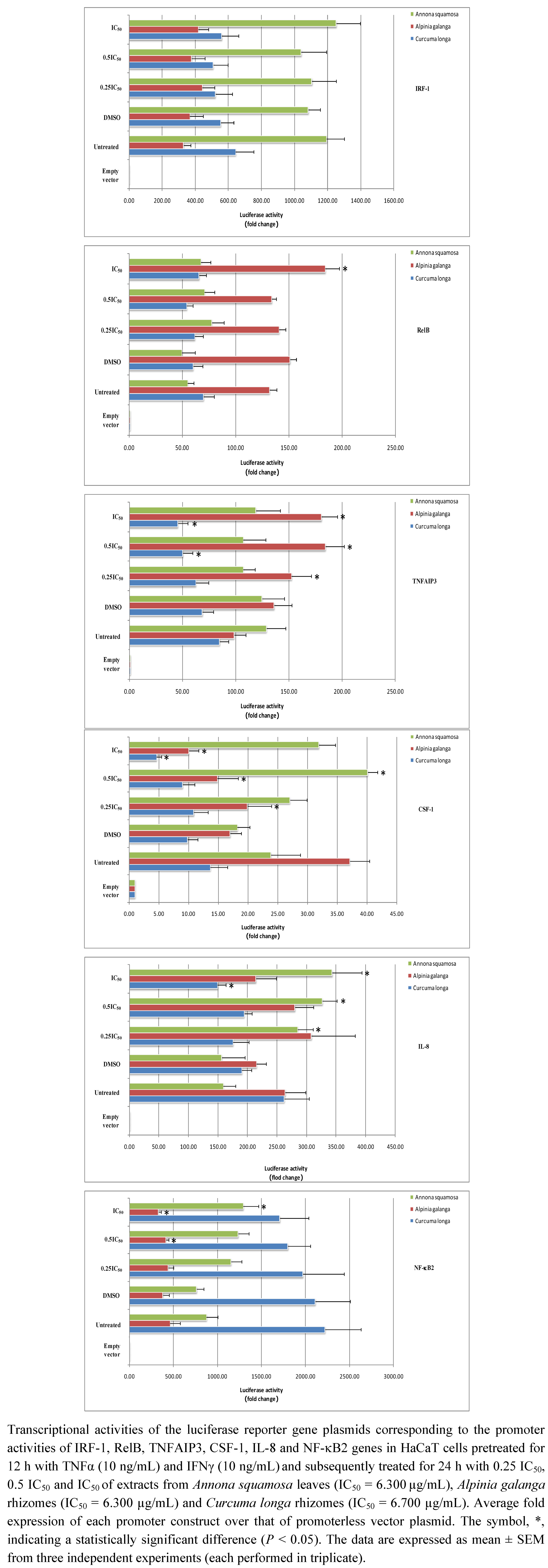{kind=link}
{kind=link}
| Name | Sequences | PCR product lengths | Ref. | |
|---|---|---|---|---|
| β-actin | forward | 5’ ACG GGT CAC CCA CAC TGT GC 3’ | 656 bp | [30] |
| reverse | 5’ CTA GAA GCA TTT GCG GTG GAC GAT 3’ | |||
| IRF-1 | forward | 5’ AAC AAG GGC AGC TCA GCT GT 3’ | 450 bp | [31] |
| reverse | 5’ TGT TGG CTG CCA CTC CGA CT 3’ | |||
| VCAM-1 | forward | 5’ AGT CAG GAA TTT CTG GAG GAT GC 3’ | 229 bp | [32] |
| reverse | 5’ GCA GCT TTG TGG ATG GAT TCA 3’ | |||
| RelB | forward | 5’ TCC CAA CCA GGA TGT CTA GC 3’ | 160 bp | [33] |
| reverse | 5’ AGC CAT GTC CCT TTT CCT CT 3’ | |||
| TNFAIP3 | forward | 5’ TTC AAG CAG ATG TAT GGC TAA CC 3’ | 267 bp | [34] |
| reverse | 5’ CCT TGG GCT GAA TCT GAC AT 3’ | |||
| CSF-1 | forward | 5’ ATG ACA GAC AGG TGG AAC TGC CAG 3’ | 438 bp | [35] |
| reverse | 5’ TCA CAC AAC TTC AGT AGG TTC AGG 3’ | |||
| CD40 | forward | 5’ AGA GTT CAC TGA AAC GGA ATG CC 3’ | 461 bp | [36] |
| reverse | 5’ ACA GGA TCC CGA AGA TGA TGG 3’ | |||
| IL-8 | forward | 5’ CTG CGC CAA CAC AGA AAT TA 3’ | 238 bp | [37] |
| reverse | 5’ ATT GCA TCT GGC AAC CCT AC 3’ | |||
| NF-κB2 | forward | 5’ CAG TGA GAA GGG CCG AAA GAC 3’ | 421 bp | This study |
| reverse | 5’ CAG GGG CAG GGA GAA GGA G 3’ | |||
| NF-κB1 | forward | 5’ AGC CCC CAA TGC ATC CAA CTT 3’ | 402 bp | [7] |
| reverse | 5’ CAA CCG CCG AAA CTA TCC GAA AAA 3’ | |||
| RelA | forward | 5’ AGC GCA TCC AGA CCA ACA ACA ACC 3’ | 419 bp | [7] |
| reverse | 5’ CCG CCG CAG CTG CAT GGA GAC AC 3’ | |||
3.5. Plasmids
3.6. Construction of Promoter Reporter Plasmids
3.7. Reporter Gene Assay
3.8. Statistical Analysis
4. Conclusions
Acknowledgements
References and Notes
- Lowes, M.A.; Bowcock, A.M.; Krueger, J.G. Pathogenesis and therapy of psoriasis. Nature 2007, 445, 866–873. [Google Scholar] [CrossRef]
- Schon, M.; Boehncke, W. Medical progress. Psoriasis. N. Engl. J. Med. 2005, 352, 1899–1912. [Google Scholar] [CrossRef]
- Ronpirin, C.; Tencomnao, T. Psoriasis: A review of the role of serotonergic system. Afr. J. Biotechnol. 2010, 9, 1528–1534. [Google Scholar]
- Shenying, F.; Xiangzhong, F.; Momiao, X. Psoriasis prediction from genome-wide SNP profiles. BMC Dermatol. 2011, 11, 1. [Google Scholar] [CrossRef]
- Nair, R.P.; Duffin, K.C.; Helms, C.; Ding, J.; Stuart, P.E.; Goldgar, D.; Gudjonsson, J.E.; Li, Y.; Tejasvi, T.; Feng, B.J.; et al. Genome-wide scan reveals association of psoriasis with IL-23 and NF-kappaB pathways. Nat. Genet. 2009, 41, 199–204. [Google Scholar]
- Li, Q.; Verma, I.M. NF-κB regulation in the immune system. Nat. Rev. Immunol. 2002, 2, 725–734. [Google Scholar] [CrossRef]
- Qin, J.Z.; Chaturvedi, V.; Denning, M.F.; Choubey, D.; Diaz, M.O.; Nickoloff, B.J. Role of NF-κB in the apoptotic-resistant phenotype of keratinocytes. J. Biol. Chem. 1999, 274, 37957. [Google Scholar]
- Tak, P.P.; Firestein, G.S. NF-kappaB: A key role in inflammatory diseases. J. Clini. Invest. 2001, 107, 7–12. [Google Scholar] [CrossRef]
- Takao, J.; Yudate, T.; Das, A.; Shikano, S.; Bonkobara, M.; Ariizumi, K.; Cruz, J. Expression of NF-κB in epidermis and the relationship between NF-κB activation and inhibition of keratinocyte growth. Br. J. Dermatol. 2003, 148, 680–688. [Google Scholar] [CrossRef]
- Lizzul, P.; Aphale, A.; Malaviya, R.; Sun, Y.; Masud, S.; Dombrovskiy, V.; Gottlieb, A. Differential Expression of Phosphorylated NF-κB/RelA in Normal and Psoriatic Epidermis and Downregulation of NF-κB in Response to Treatment with Etanercept. J. Invest. Dermatol. 2005, 124, 1275. [Google Scholar] [CrossRef]
- Johansen, C.; Flindt, E.; Kragballe, K.; Henningsen, J.; Westergaard, M.; Kristiansen, K.; Iversen, L. Inverse regulation of the nuclear factor-kappaB binding to the p53 and interleukin-8 kappaB response elements in lesional psoriatic skin. J. Invest. Dermatol. 2005, 124, 1284–1292. [Google Scholar] [CrossRef]
- Gerondakis, S.; Grossmann, M.; Nakamura, Y.; Pohl, T.; Grumont, R. Genetic approaches in mice to understand Rel/NF-kappaB and IkappaB function: Transgenics and knockouts. Oncogene 1999, 18, 6888–6895. [Google Scholar] [CrossRef]
- Saurat, J.H.; Stingl, G.; Dubertret, L.; Papp, K.; Langley, R.G.; Ortonne, J.P.; Unnebrink, K.; Kaul, M.; Camez, A. Efficacy and safety results from the randomized controlled comparative study of adalimumab vs. methotrexate vs. placebo in patients with psoriasis (CHAMPION). Br. J. Dermatol. 2008, 158, 558–566. [Google Scholar]
- Burden, A.D. Health economics and the modern management of psoriasis. Br. J. Dermatol. 2010, 163, 670–671. [Google Scholar] [CrossRef]
- Gelfand, J.M.; Feldman, S.R.; Stern, R.S.; Thomas, J.; Rolstad, T.; Margolis, D.J. Determinants of quality of life in patients with psoriasis: A study from the US population. J. Am. Acad. Dermatol. 2004, 51, 704–708. [Google Scholar] [CrossRef]
- Thongrakard, V. Screening of Thai medicinal plant extracts for in vitro anti-psoriatic activity. Master’s degree thesis, Chulalongkron University, Bangkok, Thailand, 2009. [Google Scholar]
- Nickoloff, B.J.; Karabin, G.D.; Barker, J.N.; Griffiths, C.E.; Sarma, V.; Mitra, R.S.; Elder, J.T.; Kunkel, S.L.; Dixit, V.M. Cellular localization of interleukin-8 and its inducer, tumor necrosis factor-alpha in psoriasis. Am. J. Pathol. 1991, 138, 129–140. [Google Scholar]
- Boukamp, P.; Petrussevska, R.T.; Breitkreutz, D.; Hornung, J.; Markham, A.; Fusenig, N.E. Normal keratinization in a spontaneously immortalized aneuploid human keratinocyte cell line. J. Cell Biol. 1988, 106, 761–771. [Google Scholar] [CrossRef]
- Tencomnao, T.; Ronpirin, C.; Prasansuklab, A.; Poovorawan, Y. Decreased EGFR mRNA expression in response to antipsoriatic drug dithranol in vitro. Afr. J. Biotechnol. 2009, 8, 3141–3146. [Google Scholar]
- Tse, W.P.; Che, C.T.; Liu, K.; Lin, Z.X. Evaluation of the anti-proliferative properties of selected psoriasis-treating Chinese medicines on cultured HaCaT cells. J. Ethnopharmacol. 2006, 108, 133–141. [Google Scholar] [CrossRef]
- Heyninck, K.; Beyaert, R. A20 inhibits NF-kappaB activation by dual ubiquitin-editing functions. Trends Biochem. Sci. 2005, 30, 1–4. [Google Scholar] [CrossRef]
- Beinke, S.; Ley, S.C. Functions of NF-kappaB1 and NF-kappaB2 in immune cell biology. Biochem. J. 2004, 382, 393–409. [Google Scholar] [CrossRef]
- Abdou, A.G.; Hanout, H.M. Evaluation of survivin and NF-kappaB in psoriasis, an immunohistochemical study. J. Cutan. Pathol. 2008, 35, 445–451. [Google Scholar] [CrossRef]
- Beljaards, R.C.; Van Beek, P.; Nieboer, C.; Stoof, T.J.; Boorsma, D.M. The expression of interleukin-8 receptor in untreated and treated psoriasis. Arch. Dermatol. Res. 1997, 289, 440–443. [Google Scholar] [CrossRef]
- Ohta, Y.; Hamada, Y. In situ Expression of CD40 and CD40 ligand in psoriasis. Dermatology 2004, 209, 21–28. [Google Scholar] [CrossRef]
- Cabrijan, L.; Lipozencic, J.; Batinac, T.; Lenkovic, M.; Stanic Zgombic, Z. Influence of PUVA and UVB radiation on expression of ICAM-1 and VCAM-1 molecules in psoriasis vulgaris. Coll. Antropol. 2008, 32, 53–56. [Google Scholar]
- Boots, A.W.; Wilms, L.C.; Swennen, E.L.R.; Kleinjans, J.; Bast, A.; Haenen, G.R.M.M. In vitro and ex vivo anti-inflammatory activity of quercetin in healthy volunteers. Nutrition 2008, 24, 703–710. [Google Scholar] [CrossRef]
- Satoskar, R.R.; Shah, S.J.; Shenoy, S.G. Evaluation of anti-inflammatory property of curcumin (diferuloyl methane) in patients with postoperative inflammation. Int. J. Clin. Pharmacol. Ther. Toxicol. 1986, 24, 651–654. [Google Scholar]
- Ohata, T.; Fukuda, K.; Murakami, A.; Ohigashi, H.; Sugimura, T.; Wakabayashi, K. Inhibition by 1'-acetoxychavicol acetate of lipopolysaccharide-and interferon-gamma-induced nitric oxide production through suppression of inducible nitric oxide synthase gene expression in RAW264 cells. Carcinogenesis 1998, 19, 1007. [Google Scholar] [CrossRef]
- Fisker, S.; Hansen, B.; Fuglsang, J.; Kristensen, K.; Ovesen, P.; Orskov, H.; Jorgensen, J.O. Gene expression of the GH receptor in subcutaneous and intraabdominal fat in healthy females: Relationship to GH-binding protein. Eur. J. Endocrinol. 2004, 150, 773–777. [Google Scholar] [CrossRef]
- Sun, Q.H.; Peng, J.P.; Xia, H.F.; Yang, Y. IFN-gamma promotes apoptosis of the uterus and placenta in pregnant rat and human cytotrophoblast cells. J. Interferon Cytokine Res. 2007, 27, 567–578. [Google Scholar] [CrossRef]
- Schaumann, D.H.; Tuischer, J.; Ebell, W.; Manz, R.A.; Lauster, R. VCAM-1-positive stromal cells from human bone marrow producing cytokines for B lineage progenitors and for plasma cells: SDF-1, flt3L, and BAFF. Mol. Immunol. 2007, 44, 1606–1612. [Google Scholar] [CrossRef]
- Braam, B.; de Roos, R.; Dijk, A.; Boer, P.; Post, J.A.; Kemmeren, P.P.; Holstege, F.C.; Bluysen, H.A.; Koomans, H.A. Nitric oxide donor induces temporal and dose-dependent reduction of gene expression in human endothelial cells. Am. J. Physiol. Heart. Circ. Physiol. 2004, 287, H1977–H1986. [Google Scholar] [CrossRef]
- Zhang, D.; Li, Y.; Bi, Z.; Wang, C.; Ouyang, J.; Wang, B.; Bi, Y. Sodium ferulate modified gene expression profile of oxidized low-density lipoproteins-treated endothelial cells. FASEB J. 2009, 14, 302–313. [Google Scholar]
- Gill, K.; Kirma, N.; Gunna, V.S.; Santanam, N.; Parthasarathy, S.; Tekmal, R.R. Regulation of colony stimulating factor-1 (CSF-1) in endometrial cells: Glucocorticoids and oxidative stress regulate the expression of CSF-1 and its receptor c-fms in endometrial cells. Fertil. Steril. 2001, 76, 1005–1011. [Google Scholar] [CrossRef]
- Basok, A.; Shnaider, A.; Man, L.; Chaimovitz, C.; Douvdevani, A. CD40 is expressed on human peritoneal mesothelial cells and upregulates the production of interleukin-15 and RANTES. J. Am. Soc. Nephrol. 2001, 12, 695. [Google Scholar]
- Vogel, C.F.; Sciullo, E.; Li, W.; Wong, P.; Lazennec, G.; Matsumura, F. RelB, a new partner of aryl hydrocarbon receptor-mediated transcription. Mol. Endocrinol. 2007, 21, 2941–2955. [Google Scholar] [CrossRef]
- Sample Availability: Samples of the Thai plant extracts, Curcuma longa, Alpinia galanga and Annona squamosa, are available from the authors.
© 2011 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Saelee, C.; Thongrakard, V.; Tencomnao, T. Effects of Thai Medicinal Herb Extracts with Anti-Psoriatic Activity on the Expression on NF-κB Signaling Biomarkers in HaCaT Keratinocytes. Molecules 2011, 16, 3908-3932. https://doi.org/10.3390/molecules16053908
Saelee C, Thongrakard V, Tencomnao T. Effects of Thai Medicinal Herb Extracts with Anti-Psoriatic Activity on the Expression on NF-κB Signaling Biomarkers in HaCaT Keratinocytes. Molecules. 2011; 16(5):3908-3932. https://doi.org/10.3390/molecules16053908
Chicago/Turabian StyleSaelee, Chanachai, Visa Thongrakard, and Tewin Tencomnao. 2011. "Effects of Thai Medicinal Herb Extracts with Anti-Psoriatic Activity on the Expression on NF-κB Signaling Biomarkers in HaCaT Keratinocytes" Molecules 16, no. 5: 3908-3932. https://doi.org/10.3390/molecules16053908