
Identification of Homologous Polyprenols from Thermophilic Bacteria
by
 , , and
, , and
Lucia Gharwalová
1 ,
,
Andrea Palyzová
2,
Helena Marešová
2,
Irena Kolouchová
1,
Lucie Kyselová
3 and
Tomáš Řezanka
2,* 1
Department of Biotechnology, Faculty of Food and Biochemical Technology, University of Chemistry and Technology Prague, 166 28 Prague, Czech Republic
2
Institute of Microbiology, The Czech Academy of Sciences, 142 20 Prague, Czech Republic
3
Research Institute of Brewing and Malting, 120 44 Prague, Czech Republic
*
Author to whom correspondence should be addressed.
Microorganisms 2021, 9(6), 1168; https://doi.org/10.3390/microorganisms9061168
Submission received: 29 April 2021
/
Revised: 25 May 2021
/
Accepted: 27 May 2021
/
Published: 28 May 2021
(This article belongs to the Special Issue Microbial Extremophiles as Life Pioneers and Wellsprings of Valuable Molecules)
Abstract
:Sixteen strains of five genera of thermophilic bacteria, i.e., Alicyclobacillus, Brevibacillus, Geobacillus, Meiothermus, and Thermus, were cultivated at a temperature from 42 to 70 °C. Twelve strains were obtained from the Czech Collection of Microorganisms, while four were directly isolated and identified by 16S rRNA gene sequencing from the hot springs of the world-famous Carlsbad spa (Czech Republic). Polyprenol homologs from C40 to C65 as well as free undecaprenol (C55), undecaprenyl phosphate, and undecaprenyl diphosphate were identified by shotgun analysis and RP-HPLC/MS-ESI+ (reverse phase high-performance liquid chromatography–high-resolution positive electrospray ionization mass spectrometry). The limit of detection (50 pM) was determined for individual homologs and free polyprenols and their phosphates. Thus, it has been shown that at least some thermophilic bacteria produce not just the major C55 polyprenol as previously described, but a mixture of homologs.
1. Introduction
The Eger Graben, called after the river Eger, is a geographic unit in the Czech Republic. It runs SW–NE along the Erzgebirge (Ore Mountains), is part of the European Cenozoic Rift System, and forms an extensive asymmetric tectonic moat. Its origin is related to the response of Alpine Orogeny in the Bohemian Massif, the manifestation of which was volcanic activity that began in the Late Cretaceous but reached its peak in the Tertiary times. This volcanic activity was related to the formation of thermal springs in the world-famous Carlsbad spa area (Czech Republic).
The “Vřídlo” is the official name for the largest spring of thermal water in Carlsbad spa, which has a temperature of 73 °C and contains gasified thermal water with mineralization typical of its complex deep origin, see Table S1. An average of 2000 L of thermal water is spewed from “Vřídlo” springs every minute, whose water is 18,000 years old [1]. All springs in Carlsbad spa are characterized by a uniform chemistry and are only partial sub-branches from the “Vřídlo” mainstream, which differ in temperature and, depending on it, in the amount of dissolved carbon dioxide. Microbial life in these waters very often remains largely unexplored. Despite being known worldwide, only one microbiological study has been devoted to Carlsbad’s springs, reporting the isolation and characterization of a bacterium of the genus Thermus from the spring “Vřídlo” [2].
Distribution, diversity, and activity of thermophilic (or thermotolerant) microorganisms have been analyzed mostly in high-temperature spring waters [3,4,5]. A number of different definitions have been used to describe microorganisms that grow at high temperatures, but commonly used definitions are based on optimal growth temperature [6]. Thermophiles have an optimal growth temperature in the range of 45 to <80 °C [3]. Bacteria isolated by us directly from hot springs are more like thermotolerant strains. Similarly, Hippchen et al. [7] identified, as early as 1981, bacteria related to Bacillus acidocaldarius (now Alicyclobacillus acidocaldarius [8]) in soil samples that were pasteurized for 10 min at 80 °C.
Polyisoprenoid alcohols occur from Archaea through bacteria to higher plants and animals. In bacteria, polyprenols serve primarily as specific membrane-bound carriers of glycan biosynthetic pathways responsible for the production of cellular structures such as N-linked protein glycans and bacterial peptidoglycans. In Gram-negative bacteria, the major accumulated forms are undecaprenyl phosphate (C55-P) and undecaprenyl diphosphate (C55-PP), but not free undecaprenol (C55-OH). In contrast, free undecaprenol is commonly identified in Gram-positive bacteria in addition to the two phosphates [9]. Molecules of polyprenols consist of repeating isoprenoid units linked head-tail with various structural modifications. These include cis-trans (E–Z) configurations of double bonds, their saturation (e.g., dolichols) and of course the number of repeating units. Plant polyprenols show the highest diversity in chain length [10] while, with exceptions, only polyprenol C55 (bactoprenol) has been identified in bacteria.
Many published articles [11,12,13] state that “An interesting feature of polyprenols is their occurrence as the homologous mixtures. Unique in this context are bacteria possessing a single polyprenol (undecaprenol)”. A complete exception are cyanobacteria, some of which have been shown to produce C35–C45 polyprenols [14]. Streptococcus mutans could be a certain exception, but evidence of homologs of polyprenols was only based by gas chromatography on the identity of retention times with standards [15]. Furthermore, decaprenyl-phosphate-arabinose and octahydro-heptaprenyl-phosphate-arabinose have been identified in Mycobacterium smegmatis [16]. Alicyclobacillus acidocaldarius, formerly Bacillus acidocaldarius, is a species of acidophilic, thermophilic Gram-positive bacterium [8]. De Rosa et al. [17] describe a TLC band with retention factor 0.35 which, according to MS, shows a series of molecular ions corresponding to a mixture of C45–C60 polyprenols. From the above overview of already published papers, it is clear that a number of polyprenol homologs also occur in some bacteria, which was confirmed by the data presented in the Results.
The analysis of polyprenols has been described many times, whether in reviews [10,12] or more recently in various articles, see below. Basically, the only method currently used is LC-MS with soft ionization techniques, e.g., electrospray ionization (ESI) [18], atmospheric pressure photoionization [19] or atmospheric pressure chemical ionization [20].
High-performance liquid chromatography (HPLC) combined with electrospray ionization tandem mass spectrometry (ESI-MS/MS) is one of the most suitable methods for analyzing and determining the structure of both free polyprenols and their phosphorylated derivatives. This paper presents two examples of the analysis of polyprenols and their derivatives in thermophilic bacteria, either obtained from the collection of microorganisms (CCM-Brno, Brno, Czech Republic) or directly isolated from the hot springs. The first is the use of shotgun analysis, which has several advantages over HPLC-ESI-MS/MS. In particular, it is a fast method which makes it possible to analyze a sample in the order of minutes; another advantage of shotgun analysis over LC-MS analysis is that during direct infusion the mass spectrum displays protonated molecules at constant concentrations, allowing the scanning of precursor-ion scans and/or neutral loss scans. Conversely, the use of LC-MS analysis allows the separation of individual homologs, in this case polyprenols, especially when using reverse phase HPLC (RP-HPLC). Further analysis by tandem MS, thus, allows the exact structure of each of the polyprenol homologs to be determined. This paper focused on the analysis of homologs of polyprenols obtained by saponification of total lipids of sixteen strains of thermophilic bacteria, whether obtained from the collection or directly isolated from the hot springs of the world-famous spa, Carlsbad (Czech Republic).
2. Materials and Methods
2.1. Chemicals and Standards
Undecaprenol, undecaprenyl-phospate diammonium salt, and undecaprenyl-diphosphate triammonium salt were purchased from Larodan (Malmö, Sweden). All other chemicals were purchased from Merck (Darmstadt, Germany).
2.2. Isolation of Thermophilic Bacteria from Hot Springs
Samples were collected from four thermal springs (Štěpánka, Sadový, Mlýnský, and Vřídlo) in Carlsbad, Czech Republic. Sterile 50 mL Falcon tubes were used for the sampling. Each tube was either empty or contained 1.5 mL of sterile yeast extract-peptone-dextrose broth (YPD, in g/L: dextrose 20, peptone 20, yeast extract 10, pH 7.0). The spring water was filled to 15 mL in order to achieve a 10-fold dilution of the media. The samples were then transported back to laboratory in thermos flasks. The Falcon tubes were incubated on rotary shakers (150 rpm) at temperatures matching the temperatures of the given springs, i.e., 58 °C (Štěpánka, Mlýnský and Vřídlo) or 42 °C (Sadový). After 30 days of cultivation in Falcon tubes, each sample was diluted to achieve a final cell concentration of 103 cells/mL. A 0.1 mL aliquot from each of the diluted sample was inoculated to the surface of the agar medium. The agar media used in this study were prepared from the respective micro filtered spring water. The agar media were: Reasoner’s 2A (R2A, HiMedia, Brno, Czech Republic) agar and Thermus 162 agar (HiMedia, Brno, Czech Republic). The plates were incubated at temperatures of the springs (58 °C or 42 °C) for 1–3 weeks.
Several distinct colonies with different morphologies were taken from the above-mentioned agar plates to plate count agar (PCA; HiMedia, Brno, Czech Republic) plates and incubated at 58 °C or 42 °C for 48 h for further isolation and purification.
2.3. Molecular Identification of Four Strains of Thermophilic Bacteria from Hot Springs
Molecular identification was done by sequencing 16S rRNA gene amplified by polymerase chain reaction (PCR) using 16S rRNA gene primers Fwd27 and Rev1492 [21]. PCR amplicons were purified using a High Pure PCR Product Purification Kit (Roche, Basel, Switzerland) following the manufacturer’s protocol. The PCR amplicons were sequenced on an ABI PRISM 3130xl Genetic Analyzer (Applied Biosystems, Waltham, MA, USA). The sequences so obtained were edited by Chromas Lite software (Technelysium Pty Ltd., Brisbane, Australia) and assembled using SeqMan (DNASTAR, Inc., Madison, WI, USA). Searching for 16S rRNA gene sequence similarity was performed at the GenBank data library using the BLASTN program (NCBI, Bethesda, MD, USA).
2.4. Cultivation of Four Strains of Thermophilic Bacteria
For the pre-inoculum, four bacterial isolates from hot springs (designated Geobacillus stearothermophilus ST-YPD, Brevibacillus agri SA-1, Geobacillus kaustophilus ML-1, and Geobacillus stearothermophilus VR-1) were cultivated in Luria-Bertani broth (LB; 1% tryptone, 0.5% yeast extract, 1% NaCl, pH 7.0) on orbital shaker (150 rpm) for 72 h. For the inoculum, 100 mL of LB was inoculated with 10 mL of pre-culture to a final concentration of OD600 0.2 and incubated on a rotary shaker at 150 rpm for 24 h. For lipid production, 200 mL of LB was inoculated with 10 mL of pre-culture to a final concentration of OD600 0.2 and incubated on an orbital shaker at 150 rpm for 48 h to the stationary phase. The cultivation temperatures for isolate Geobacillus stearothermophilus ST-YPD was 42 °C and for the remaining three isolates 58 °C (corresponding to the original spring temperature). After cultivation, the cells were centrifuged (10,000× g, 10 min, 4 °C) and washed twice. Biomass yield was determined as dry weight after lyophilization. Cell mass was frozen at −70 °C and lyophilized. An overview of the cultivated bacterial isolates is given in Table 1.
2.5. Extraction and Isolation of Polyprenols
The extraction procedure was based on the method of Bligh and Dyer [22]. Briefly, the lyophilized cells (approximately 10 mg) were suspended in a chloroform-methanol mixture (2:1) (~1 mL) for 30 min at 20 °C with stirring, after which, chloroform and water were added and the insoluble material was separated by centrifugation. The aqueous phase was aspirated off and the chloroform phase was evaporated to dryness under reduced pressure. Part of extract was used for shotgun mass spectrometry and another part for saponification.
This latter part of extract was heated to 95 °C for 1 h with 3 mL of 15 M KOH aqueous solution. Non-saponifiable lipids were then extracted three times with hexane, the combined extracts were evaporated to dryness under a stream of nitrogen and analyzed by LC-MS.
2.6. Analysis of Polyprenols by Shotgun Mass Spectrometry
An LTQ-Orbitrap Velos mass spectrometer (Thermo Fisher Scientific, San Jose, CA, USA) equipped with a heated electrospray interface (HESI) was operated in positive and negative ionization mode. The MS scan range was performed in the FT cell and recorded within a window between 150 and 1500 Da. The mass resolution was set to 100.000, and the ion spray voltage was set at 3.4 kV (in the positive ionization mode) and −2.6 kV (in the negative ionization mode). Both ionization modes used the following parameters: sheath gas flow, 19 arbitrary units (AU); auxiliary gas flow, 8 AU; ion source temperature, 270 °C; capillary temperature, 240 °C; capillary voltage, 55 V; and tube lens voltage, 165 V. Helium was used as a collision gas for collision induced dissociation (CID) experiments. The CID normalization energy of 33% was used for the fragmentation of parent ions. Flow Injection Analysis was used for sample introduction into the heated ESI-MS (H-ESI-MS) ion source. Hexane/propan-2-ol (50/50 v/v) was used at the flow rate of 125 μL/min. The m/z value of the molecular weight-related ion of C55 polyprenol was measured at 765.6917 by FT mode and the mass accuracy of C55 polyprenol was +0.2 ppm, compared with the theoretical value of 765.6919 (Table S2). The differences between the measured and calculated values for all analyzed compounds, whether in positive or negative ESI, do not exceed 0.3 ppm (0.0003 Da).
2.7. LC-MS Analysis of Saponified Polyprenols
The HPLC equipment consisted of a 1090 Win system, PV5 ternary pump and automatic injector (HP 1090 series, Hewlett Packard, Palo Alto, CA, USA), and Ascentis Express column HIRPB-250AM (Hichrom Limited, Berkshire, UK), 250 × 2.1 mm ID, 5 μm particle size. LC was performed at a flow rate of 350 μL/min, with a linear gradient from mobile phase containing propan-2-ol/methanol/aqueous 1 mM ammonium acetate/lithium acetate in methanol (1 µM/mL) (50:40:10:10, v/v/v/v) to hexane/propan-2-ol/methanol/aqueous 1 mM ammonium acetate/lithium acetate in methanol (1 µM/mL) (10:70:10:10, v/v/v/v)/ with 30 min and held for 10 min. The composition was returned to the initial conditions over 10 min. The whole HPLC flow was introduced into the ESI source without any splitting. The performance was measured by undecaprenol as an internal standard under the conditions given above.
2.8. Calibration
The polyprenol standard (commercially obtained C55 polyprenol from Larodan (Malmö, Sweden)) was analyzed separately and SIM at m/z 773.7146 ([M+Li]+) of C55 polyprenol was obtained. A calibration curve was generated using the area of peak of the SIM corresponding to the different concentrations of standard (1 pM, 10 pM, 50 pM, 100 pM, 1 nM, 10 nM, 100 nM, and 1 μM) and the linear regression of the calibration curve, as well as the signal/noise (S/N) ratio was calculated.
2.9. Statistical Analysis
The statistical analysis was performed using the IBM SPSS Statistics (Statistical Package for the Social Sciences; IBM Corp, 2013) Statistics software, version 26 (IBM® Corporation, Armonk, NY, USA).
3. Results and Discussion
3.1. Characterization and Identification of Bacterial Isolates
The molecular classification of four strains (SA-1, ML-1, ST-YPD, and VR-1) was carried out by the 16S rRNA gene sequence analysis of an approximately 1.5 kb 16S rRNA fragment amplified from the total DNA of these isolates. The BLAST search showed that these strains belong to the order Bacillales, namely to the genus Brevibacillus or Geobacillus. The partial nucleotide sequence of 16S rRNA gene of these strains (about 1355 nucleotides) showed the maximum nucleotide identity (over 99%) with the strains Brevibacillus agri DSM 6348, Geobacillus kaustophilus BGSC 90A1, and Geobacillus stearothermophilus BGSC 9A20. The conventional physiological and biochemical characterization was performed at the Czech Collection of Microorganisms, Masaryk University, Brno. All strains were defined as Gram-positive, rod-shaped in singlets (or in pairs), aerobic and thermophilic organisms, and spore-forming bacteria (ellipsoidal or oval spores are located at a terminal or subterminal position). The temperature ranges for growth: 40.0–75.0 °C with the optimum 42–58 °C; pH range for growth: 5.0–10.0 with the optimum of 6.0–7.0. On nutrient R2A, the isolate SA-1 formed white, flat colonies with irregular borders. The isolates ML-1, ST-YPD, and VR-1 were characterized by circular umbonate colonies of light orange color with transparent margins. Positive results for the strains ML-1, ST-YPD, and VR-1: catalase, hydrolysis of aesculin, and DNA, nitrite reduction, acids from glucose, mannitol, cellobiose, and fructose. Negative results: hydrolysis of gelatin starch, casein, Tween 80, tyrosine, production of acetoin, acids from xylose, lactose and inositol, growth in Simmons citrate, and arginine dihydrolase. Moreover, the comparison of the two strains ST-YPD and VR-1 was performed; they differed in hydrolysis of o-nitrophenyl-β-galactoside (positive result for the strain VR-1), growth in the presence of 10% NaCl and acids from xylose and mannitol (positive results for the strain ST-YPD). Phenotypic characterization of the strain SA-1 revealed that the isolate tested positive for catalase, urease, hydrolysis of aesculin and casein, growth in glucose, indole, and H2S production. The isolate was also capable of forming ellipsoidal spores in swollen sporangia. The isolate tested negative for oxidase, nitrite reduction, Methyl Red/Voges-Proskauer (MR/VP) test, and citrate utilization. Since the results of the molecular analysis were consistent with the phenotypic traits, we designated strains as the Brevibacillus agri SA-1, Geobacillus kaustophilus ML-1, Geobacillus stearothermophilus ST-YPD, and Geobacillus stearothermophilus VR-1.
3.2. Shotgun Analysis of Polyprenols
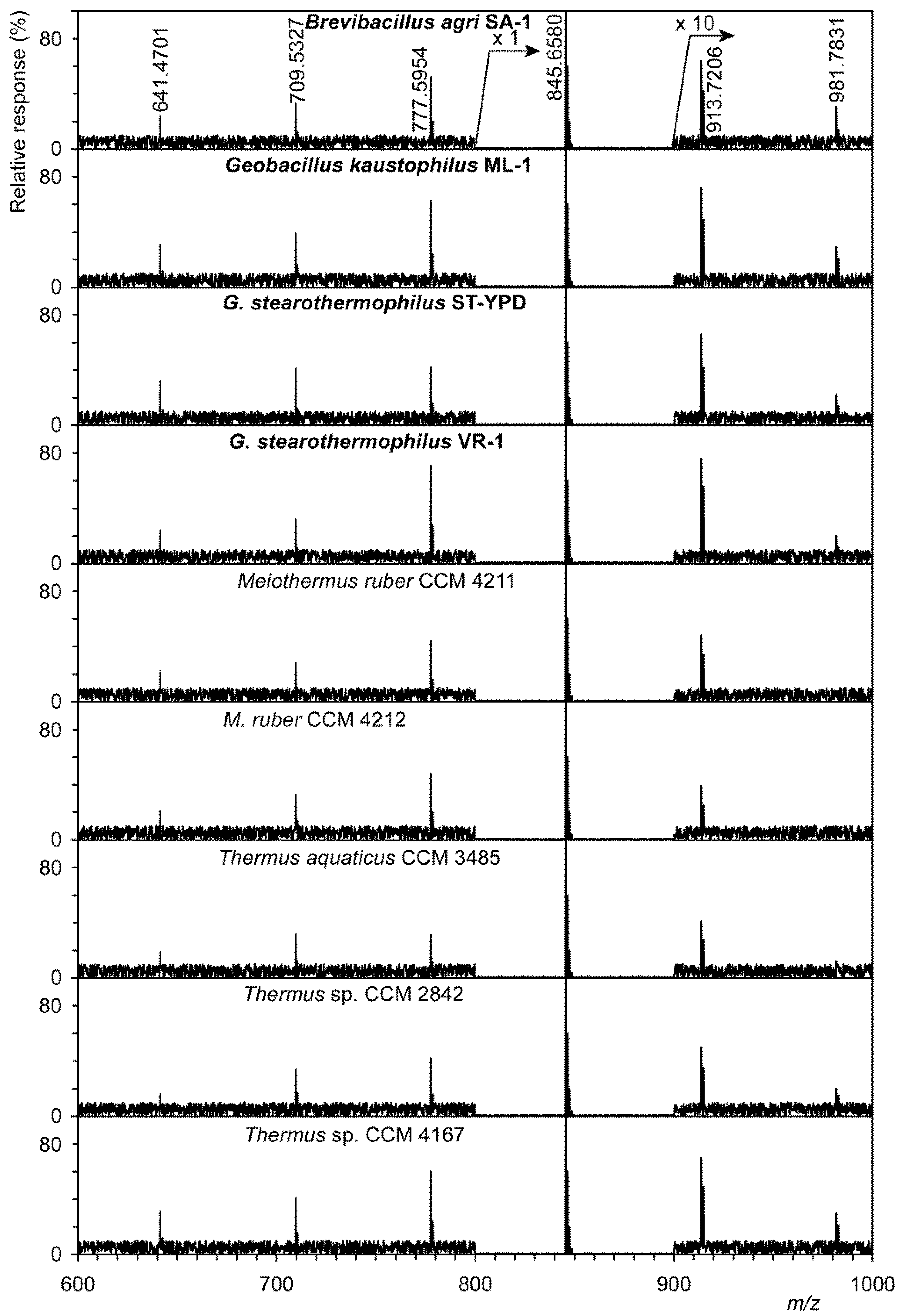
A shotgun analysis display of polyprenol phosphates is shown in Figure 1. Using a precursor ion scan at m/z 163.0165 (the structure of this ion is shown in the upper right corner of Figure 1), it was possible to obtain preliminary information on homologs of polyprenol phosphates. This image shows only those strains that originate from hot springs, regardless of whether they were spring isolates or strains from the CCM collection. In all strains we identified, albeit sometimes as very minor, homologs were from C40 to C65. Values for [M–H]− both measured and calculated, including summary formulas are given in Table S2. This table shows a clear agreement between the measured and calculated values for [M–H]− ions, which does not exceed 0.3 ppm (0.0003 Da). Full confirmation of the structure was performed by tandem MS in negative mode for all six polyprenol phosphates. A description of the tandem MS for the lowest identified C40-P is given as an example. A homologous series was identified from the ion at m/z 163.0164 (C5H8O4P−) to the ion at m/z 571.3919 (C35H48O4P−, for the structure see Figure S1). These values are in good agreement with already published data [24].
To exclude other homologs (e.g., partially hydrogenated dolichol phosphates, i.e., C40-P, C45-P, and C50-P), that were identified, e.g., in the thermoacidophilic archaeon Sulfolobus acidocaldarius [25], we used the shotgun analysis in which both shorter and more saturated compounds would appear in the mass spectra.
3.3. LC-MS Analysis of Saponified Polyprenols
Based on these preliminary analyses, see Figure 1, the total lipids were saponified with KOH, see Materials and Methods and unsaponifiable mixture was extracted with hexane and used for LC-MS analysis. Based on both previous good experience [26,27,28] and published data [18], the analyses were performed in positive high-resolution ESI with the addition of Li+ ions. The use of SIM for [M+Li]+ ions made it possible to identify individual polyprenol homologs. The results of the SIM analysis are shown in Table 2. Table S2 lists the detected and theoretical masses of [M+Li]+ ions used for SIM analysis. The data from this table confirm that the total lipid extract of each of 16 strains showed the presence of more than one major homologue of C55 (undecaprenol).
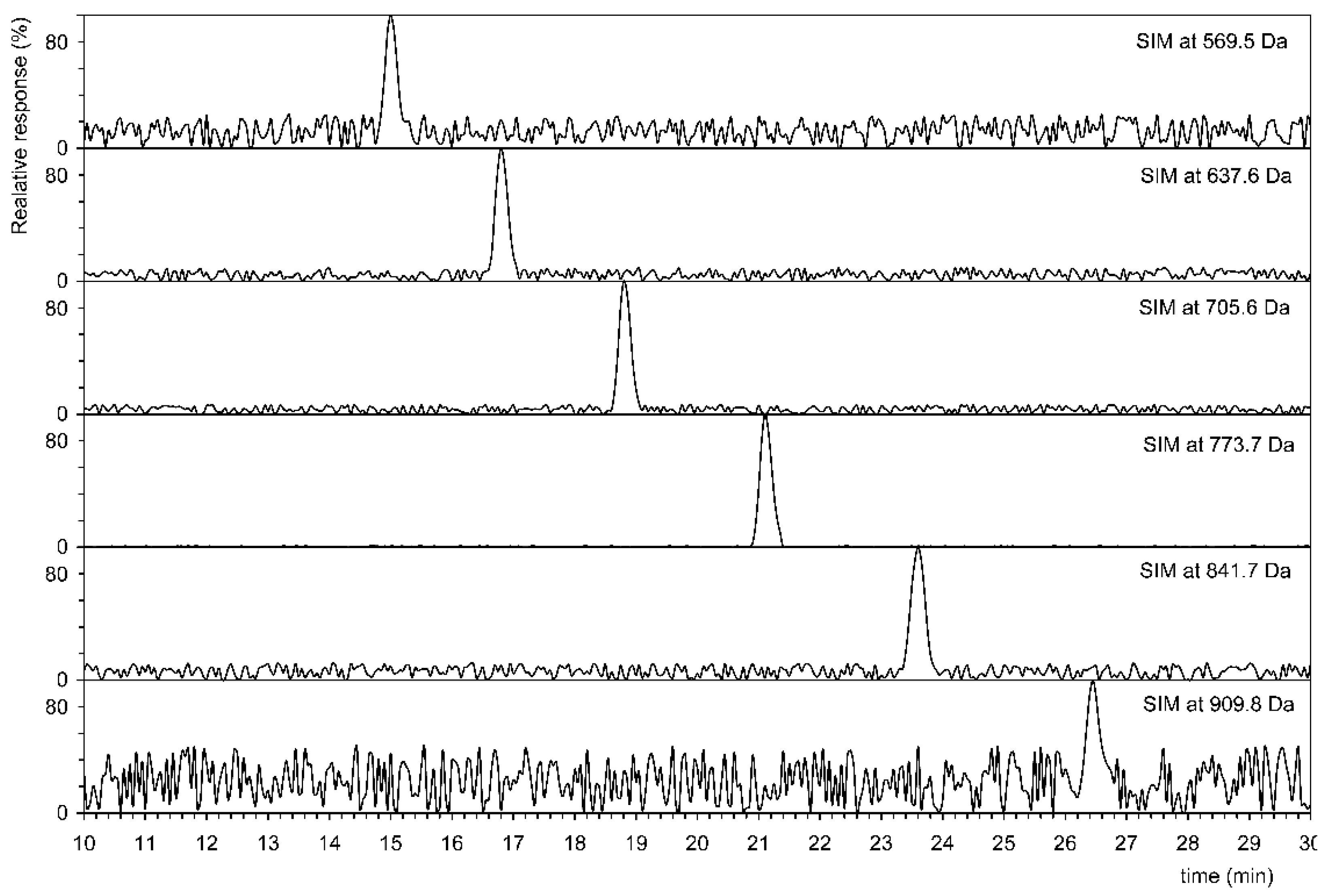
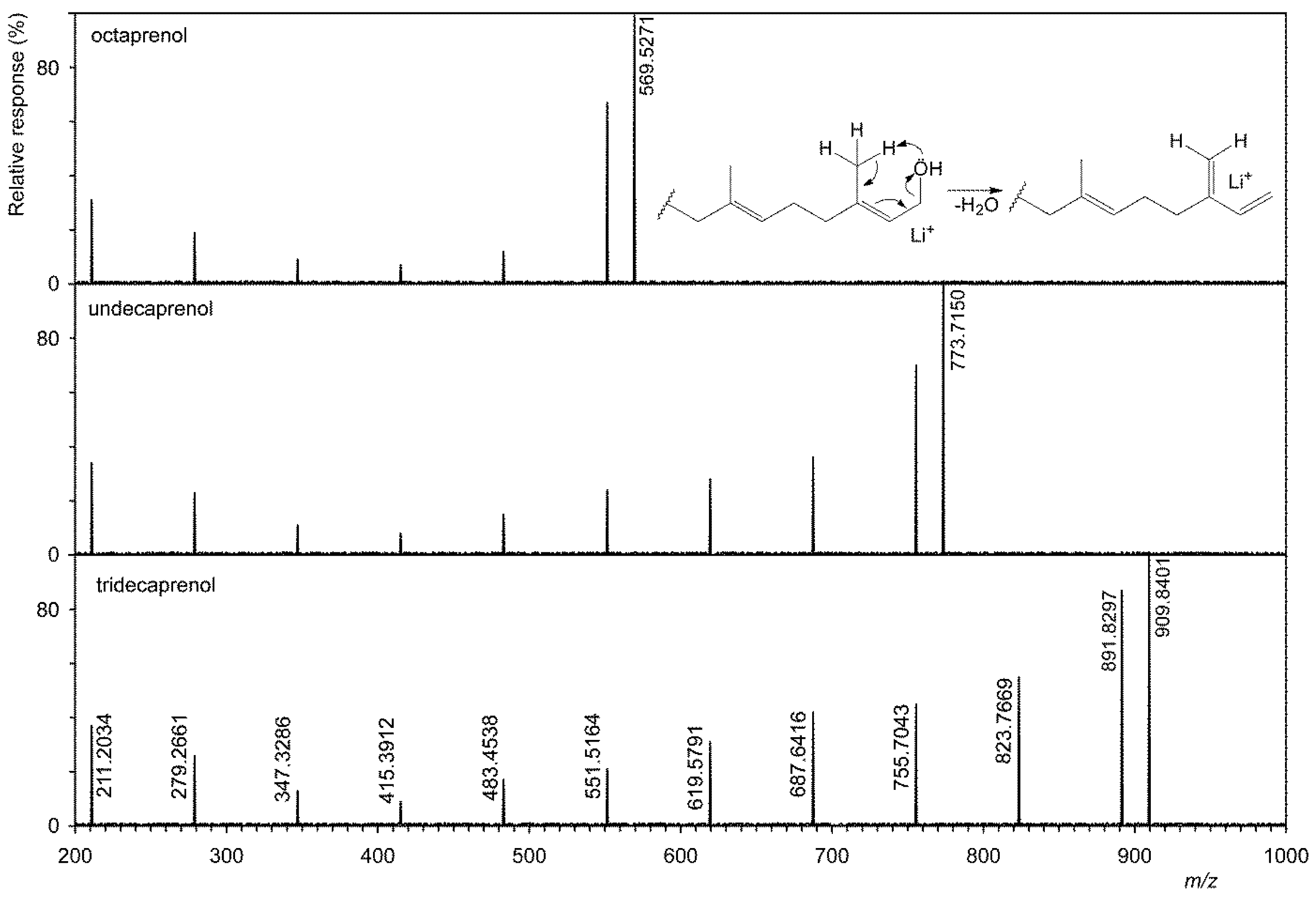
Figure 2 shows that six homologs have been identified in G. stearothermophilus VR-1, and their abundances are shown in Table 2. This table also shows the results of analysis of all other strains. The tandem MS of three homologs is showed on Figure 3, i.e., the shortest, longest, and most abundant homologs that have been identified. Tandem MS are relatively simple, the base peak is always an [M+Li]+ type ion (e.g., 909.8401 Da), and an abundant ion [M+Li-H2O]+ (e.g., 891.8297 Da) is also present. Furthermore, abundant ions of the type [M+Li-H2O-C5H8]+ were obtained. Polyprenols, due to the double bond in the allylic position to the hydroxyl, very easily lose water to form a conjugate system, see the insertion structure in Figure 3. This formation is facilitated by the transient formation of a six-member ring in which the H atom is transferred to the OH group. In contrast to D’Alexandri et al. [18], where ion formation [M+Li-CH2O]+ of m/z 743.7040 was observed, this ion was not found in polyprenol spectrum, see Figure 3.
The calibration curve was determined based on various concentrations of commercially available C55 polyprenol, see Materials and Methods. The detection limit, i.e., 50 pM (38.6 pg/μL) (defined as the concentration of the sample with an S/N ratio > 3) was obtained. This value is fully consistent with previously published concentrations; e.g., Skorupinska Tudek et al. [29] indicate an interval of 40 pg to 10 ng of the injected standard, or e.g., D’Alexandri et al. [18] where 0.10 nM C55 polyprenol is given.
As described above, polyprenol homologs other than C55 have been only rarely identified. However, there are several references that shorter and longer homologs than C55 exist. In particular, homologues from C35 to C45 have been identified in the above-mentioned cyanobacteria [14]. These include A. acidocaldarius [15] or S. mutans [17], where C45–C60 polyprenols have been found. Additionally, the presence of decaprenyl and/or nonaprenyl (C50 and C45) phosphate [30] was proved by mass spectrometry in three genera of Gram-positive bacteria of the genus Romboutsia. Four homologs of polyprenols (C45–C60) have been identified in Streptococcus faecalis, C55 being predominant [31].
C50-P and trace C45-P were also identified. This again confirmed that although C55 is the major homolog, lower and higher homologs and those polyprenol phosphates are present. Polyprenols of three isolates, Romboutsia lituseburensis, R. ilealis, and Romboutsia sp. were determined and decaprenyl-phosphate (C50) was found to be the major one by mass spectrometry. Furthermore, the presence of decaprenyl phosphate and/or nonaprenyl phosphate as minor polyprenols has been demonstrated in all three species of bacteria of the genus Romboutsia [30].
Table 2 shows that in all 16 strains lower and higher homologs than C55 were identified in percent of majority polyprenol (C55). Prenols with less than 8 and more than 13 units were not detected, indicating that its concentration could be below the detection limit (LOD) for ESI+-MS (50 pM). For this reason, we cannot exclude the presence of other homologues in trace amounts.
Detection of undecaprenol (C55), undecaprenyl phosphate (C55-P), and undecaprenyl diphosphate (sometimes also called pyrophosphate) (C55-PP) by shotgun negative ESI was first performed on commercially obtained standards. In the tandem MS spectra, a major ion of the type [M–H]− was always identified at m/z 765.6919 (C55H89O−), 845.6582 (C55H90O4P−), and 925.6246 (C55H91O7P2−). To fully confirm the structure of the above polyprenols, a tandem electrospray MS in negative mode was performed. In undecaprenyl phosphate, the ion at m/z 78.9591 (PO3−) was identified as the majority, while in undecaprenyl diphosphate the two ions at m/z 78.9591 and the ion at 158.9254 (HP2O6−) were identified. An ion of this type has previously been identified, for example, in Chhonker et al. [32] in tandem MS of geranylgeranyl diphosphate. Unfortunately, commercially available standards contained about 5% lower and higher homologs, as evidenced by the negative ESI spectra. For example, two ions with m/z 777.5956 (C50-P) and m/z 913.7208 (C60-P), respectively, have been identified in undecaprenyl phosphate.
Biosynthesis of polyprenols in Gram-positive bacteria proceeds by successive head-to-tail additions of IPP (isopentenyl diphosphate) that generate longer-chain polyisoprenoid dophosphates. Both C55-P and/or C55-PP are loaded with glycans for the polymerization of cell wall components and can also be replenished during bacterial cell wall synthesis. The individual C55 compounds (C55-OH, C55-P, and C55-PP) can be converted into each other [33]. Based on shotgun analysis and by calibrating three commercially available standards, the ratio between them was determined (Table 3). This ratio is very similar to the published data [34], where polyprenols were found in Staphylococcus aureus ratio 113:43:70 nM/mg (dry weight) of cells, i.e., C55-PP:C55-P:C55-OH. Direct determination, e.g., by HPLC, is difficult because, due to the very different polarity, the individual compounds have different retention times [34].
4. Conclusions
All the above results of analyses of both sixteen strains and published analyses provide a conclusive evidence that at least thermophilic bacteria biosynthesize different homologs of polyprenols. In addition, various peptides catalyzing reactions in homologs of varying length, i.e., geranylgeranylglyceryl/heptaprenylglyceryl phosphate synthase (ATA58721), farnesyl diphosphate synthase (ALA71315), and trans-hexaprenyltranstransferase (ATA60441) are known due to the known sequence of the entire G. stearothermophilus genome (NZ_CP008934.1).
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/microorganisms9061168/s1, Figure S1: The structure of the ion at m/z 163.0165 (C5H8O4P−), Table S1: Chemical composition of the spring “Vřídlo“, Table S2: Designation of compounds based on their length and number of carbon in the chain, detected and theoretical masses of ions [M–H]− and/or [M+Li]+ (Da), and molecular formulas, Table S3: The data of tandem mass spectrum of C55-PP.
Author Contributions
Conceptualization, A.P. and T.Ř.; Data curation, L.G.; Formal analysis, H.M.; Funding acquisition, I.K., L.K., and T.Ř.; Investigation, A.P., T.Ř., and L.K.; Project administration, T.Ř., L.G., and L.K.; Resources, A.P., and H.M.; Supervision, T.Ř.; Visualization, I.K.; Writing—original draft, T.Ř.; Writing—review & editing, T.Ř. and A.P. All authors have read and agreed to the published version of the manuscript.
Funding
The work was supported by Czech Science Foundation (GACR) project 18-00036S, by Institutional Research Concept RVO61388971 and by the Ministry of Agriculture of the Czech Republic (institutional support MZE-RO1918).
Data Availability Statement
The obtained sequences see Table 1 were submitted in GenBank.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Pačes, T.; Šmejkal, V. Magmatic and Fossil Components of Thermal and Mineral Waters in the Eger River Continental Rift (Bohemian Massif, Central Europe). In Water—Rock interaction, Wanty and Seal II; Taylor and Francis Group: London, UK, 2004; pp. 167–172. ISBN 90-5809-641-6. [Google Scholar]
- Peckova, M. Properties of a hyperthermophilic bacterium (Thermus sp.) isolated from a Carlsbad spring. Folia Microbiol. 1991, 36, 515–521. [Google Scholar] [CrossRef]
- Mehta, D.; Satyanarayana, T. Diversity of Hot Environments and Thermophilic Microbes. In Thermophilic Microbes in Environmental and Industrial Biotechnology; Springer Netherlands: Dordrecht, The Netherlands, 2013; pp. 3–60. ISBN 978-94-007-5898-8. [Google Scholar]
- Inskeep, W.P.; Jay, Z.J.; Tringe, S.G.; Herrgård, M.J.; Rusch, D.B.; Co, Y.M.P.S. The YNP metagenome project: Environmental parameters responsible for microbial distribution in the Yellowstone geothermal ecosystem. Front. Microbiol. 2013, 4, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inskeep, W.P.; Jay, Z.J.; Herrgard, M.J.; Kozubal, M.A.; Rusch, D.B.; Tringe, S.G.; Macur, R.E.; Jennings, R.D.; Boyd, E.S.; Spear, J.R.; et al. Phylogenetic and Functional Analysis of Metagenome Sequence from High-Temperature Archaeal Habitats Demonstrate Linkages between Metabolic Potential and Geochemistry. Front. Microbiol. 2013, 4, 95. [Google Scholar] [CrossRef] [Green Version]
- Stetter, K.O. Hyperthermophilic procaryotes. FEMS Microbiol. Rev. 1996, 18, 149–158. [Google Scholar] [CrossRef]
- Hippchen, B.; Röll, A.; Poralla, K. Occurrence in soil of thermo-acidophilic bacilli possessing ω-cyclohexane fatty acids and hopanoids. Arch. Microbiol. 1981, 129, 53–55. [Google Scholar] [CrossRef]
- Wisotzkey, J.D.; Jurtshuk, P.; Fox, G.E.; Deinhard, G.; Poralla, K. Comparative Sequence Analyses on the 16S rRNA (rDNA) of Bacillus acidocaldarius, Bacillus acidoterrestris, and Bacillus cycloheptanicus and Proposal for Creation of a New Genus, Alicyclobacillus gen. nov. Int. J. Syst. Bacteriol. 1992, 42, 263–269. [Google Scholar] [CrossRef]
- Hartley, M.D.; Imperiali, B. At the membrane frontier: A prospectus on the remarkable evolutionary conservation of polyprenols and polyprenyl-phosphates. Arch. Biochem. Biophys. 2012, 517, 83–97. [Google Scholar] [CrossRef] [Green Version]
- Řezanka, T.; Votruba, J. Chromatography of long chain alcohols (polyprenols) from animal and plant sources. J. Chromatogr. A 2001, 936, 95–110. [Google Scholar] [CrossRef]
- Surmacz, L.; Swiezewska, E. Polyisoprenoids—Secondary metabolites or physiologically important superlipids? Biochem. Biophys. Res. Commun. 2011, 407, 627–632. [Google Scholar] [CrossRef]
- Swiezewska, E.; Danikiewicz, W. Polyisoprenoids: Structure, biosynthesis and function. Prog. Lipid Res. 2005, 44, 235–258. [Google Scholar] [CrossRef]
- Yamaguchi, T.; Fujikawa, N.; Nimura, S.; Tokuoka, Y.; Tsuda, S.; Aiuchi, T.; Kato, R.; Obama, T.; Itabe, H. Characterization of lipid droplets in steroidogenic MLTC-1 Leydig cells: Protein profiles and the morphological change induced by hormone stimulation. Biochim. Biophys. Acta 2015, 1851, 1285–1295. [Google Scholar] [CrossRef]
- Bauersachs, T.; Schouten, S.; Compaoré, J.; Stal, L.J.; Damsté, J.S.S. Occurrence of C35–C45 polyprenols in filamentous and unicellular cyanobacteria. Org. Geochem. 2010, 41, 867–870. [Google Scholar] [CrossRef] [Green Version]
- Szabo, E.; Amdur, B.; Socransky, S. Lipid Composition of Streptococcus mutans. Caries Res. 1978, 12, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Wolucka, B.; McNeil, M.; de Hoffmann, E.; Chojnacki, T.; Brennan, P. Recognition of the lipid intermediate for arabinogalactan/arabinomannan biosynthesis and its relation to the mode of action of ethambutol on mycobacteria. J. Biol. Chem. 1994, 269, 23328–23335. [Google Scholar] [CrossRef]
- De Rosa, M.; Gambacorta, A.; Minale, L.; Bu’Lock, J.D. Isoprenoids of Bacillus acidocaldarius. Phytochemistry 1973, 12, 1117–1123. [Google Scholar] [CrossRef]
- D’Alexandri, F.L.; Gozzo, F.C.; Eberlin, M.N.; Katzin, A.M. Electrospray ionization mass spectrometry analysis of polyisoprenoid alcohols via Li+ cationization. Anal. Biochem. 2006, 355, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Kania, M.; Skorupinska-Tudek, K.; Swiezewska, E.; Danikiewicz, W. Atmospheric pressure photoionization mass spectrometry as a valuable method for the identification of polyisoprenoid alcohols. Rapid Commun. Mass Spectrom. 2012, 26, 1705–1710. [Google Scholar] [CrossRef] [PubMed]
- Yu, J.; Wang, Y.; Qian, H.; Zhao, Y.; Liu, B.; Fu, C. Polyprenols from the needles of Taxus chinensis var. mairei. Fitoterapia 2012, 83, 831–837. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.J. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; John Wiley & Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Bligh, E.; Dyer, W. A Rapid Method of Total Lipid Extraction and Purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siristova, L.; Melzoch, K.; Řezanka, T. Fatty acids, unusual glycophospholipids and DNA analyses of thermophilic bacteria isolated from hot springs. Extremophiles 2008, 13, 101–109. [Google Scholar] [CrossRef] [PubMed]
- Haeuptle, M.A.; Hülsmeier, A.J.; Hennet, T. HPLC and mass spectrometry analysis of dolichol-phosphates at the cell culture scale. Anal. Biochem. 2010, 396, 133–138. [Google Scholar] [CrossRef] [Green Version]
- Guan, Z.; Meyer, B.H.; Albers, S.-V.; Eichler, J. The thermoacidophilic archaeon Sulfolobus acidocaldarius contains an unsually short, highly reduced dolichyl phosphate. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2011, 1811, 607–616. [Google Scholar] [CrossRef] [Green Version]
- Řezanka, T.; Kambourova, M.; Derekova, A.; Kolouchová, I.; Sigler, K. LC–ESI–MS/MS Identification of Polar Lipids of Two Thermophilic Anoxybacillus Bacteria Containing a Unique Lipid Pattern. Lipids 2012, 47, 729–739. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Matoulkova, D.; Kyselová, L.; Sigler, K. Identification of plasmalogen cardiolipins from pectinatus by liquid chromatography–high resolution electrospray ionization tandem mass spectrometry. Lipids 2013, 48, 1237–1251. [Google Scholar] [CrossRef] [PubMed]
- Řezanka, T.; Matoulková, D.; Benada, O.; Sigler, K. Lipidomics as an important key for the identification of beer-spoilage bacteria. Lett. Appl. Microbiol. 2015, 60, 536–543. [Google Scholar] [CrossRef] [PubMed]
- Skorupińska-Tudek, K.; Bieńkowski, T.; Olszowska, O.; Furmanowa, M.; Chojnacki, T.; Danikiewicz, W.; Swiezewska, E. Divergent pattern of polyisoprenoid alcohols in the tissues of Coluria geoides: A new electrospray lonization MS approach. Lipids 2003, 38, 981–990. [Google Scholar] [CrossRef] [PubMed]
- Guan, Z.; Chen, L.; Gerritsen, J.; Smidt, H.; Goldfine, H. The cellular lipids of Romboutsia. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2016, 1861, 1076–1082. [Google Scholar] [CrossRef]
- Umbreit, J.N.; Stone, K.J.; Strominger, J.L. Isolation of Polyisoprenyl Alcohols from Streptococcus faecalis. J. Bacteriol. 1972, 112, 1302–1305. [Google Scholar] [CrossRef] [Green Version]
- Chhonker, Y.S.; Haney, S.L.; Bala, V.; Holstein, S.A.; Murry, D.J. Simultaneous Quantitation of Isoprenoid Pyrophosphates in Plasma and Cancer Cells Using LC-MS/MS. Molecules 2018, 23, 3275. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.-Y.; Wang, S.-C.; Cheng, T.-J.R.; Wong, C.-H. Undecaprenyl phosphate phosphatase activity of undecaprenol kinase regulates the lipid pool in gram-positive bacteria. Biochemistry 2017, 56, 5417–5427. [Google Scholar] [CrossRef]
- Barreteau, H.; Magnet, S.; El Ghachi, M.; Touzé, T.; Arthur, M.; Mengin-Lecreulx, D.; Blanot, D. Quantitative high-performance liquid chromatography analysis of the pool levels of undecaprenyl phosphate and its derivatives in bacterial membranes. J. Chromatogr. B 2009, 877, 213–220. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Shotgun analysis of a mixture of polyprenol phosphates from thermophilic bacteria obtained from the Czech Collection of Microorganisms (Brno, Czech Republic) and isolated from hot springs (marked in bold). Precursor ion scan at m/z 163.0165, see structure formula at the top right.
Figure 1.
Shotgun analysis of a mixture of polyprenol phosphates from thermophilic bacteria obtained from the Czech Collection of Microorganisms (Brno, Czech Republic) and isolated from hot springs (marked in bold). Precursor ion scan at m/z 163.0165, see structure formula at the top right.

Figure 2.
Selected ion monitoring (SIM) indicative of six homologs of polyprenols from C40 (569.5 Da) to C65 (909.8 Da) in Geobacillus stearothermophilus VR-1.
Figure 2.
Selected ion monitoring (SIM) indicative of six homologs of polyprenols from C40 (569.5 Da) to C65 (909.8 Da) in Geobacillus stearothermophilus VR-1.

Figure 3.
Tandem mass spectra of shortest (octaprenol, i.e., C40), most abundant (undecaprenol, i.e., C55), and longest (tridecaprenol, i.e., C65) polyprenol homologues.
Figure 3.
Tandem mass spectra of shortest (octaprenol, i.e., C40), most abundant (undecaprenol, i.e., C55), and longest (tridecaprenol, i.e., C65) polyprenol homologues.


{kind=link}
{kind=link}
{kind=link}
Table 1.
List of the cultivated bacterial isolates from four springs of Carlsbad (Karlovy Vary), Czech Republic.
Table 1.
List of the cultivated bacterial isolates from four springs of Carlsbad (Karlovy Vary), Czech Republic.
| Isolate | Hot Spring | Cultivation Temperature (°C) | GenBank Accession Numbers |
|---|---|---|---|
| Brevibacillus agri SA-1 | Sadový | 42 | MT251434 |
| Geobacillus kaustophilus ML-1 | Mlýnský | 58 | MT251494 |
| Geobacillus stearothermophilus ST-YPD | Štěpánka | 58 | MT251887 |
| Geobacillus stearothermophilus VR-1 | Vřídlo | 58 | MT251886 |
Table 2.
Content (relative %) of polyprenol homologs from 16 strains of thermophilic bacteria obtained after saponification using reverse-phase liquid chromatography-positive electrospray mass spectrometry.
Table 2.
Content (relative %) of polyprenol homologs from 16 strains of thermophilic bacteria obtained after saponification using reverse-phase liquid chromatography-positive electrospray mass spectrometry.
| Strain | C40 | C45 | C50 | C55 | C60 | C65 |
|---|---|---|---|---|---|---|
| Alicyclobacillus acidoterrestris CCM a 4659 (Apple-grape-raspberry juice) b | 0.9 ± 0.2 c | 1.8 ± 0.5 | 5.3 ± 1.5 | 87.6 ± 2.1 | 3.5 ± 1.0 | 0.9 ± 0.2 |
| Alicyclobacillus acidoterrestris CCM 4660 (Apple-grape-raspberry juice) | 0.0 ± 0.0 | 1.8 ± 0.7 | 4.6 ± 1.7 | 90.9 ± 1.8 | 2.7 ± 0.9 | 0.0 ± 0.1 |
| Brevibacillus agri SA-1 (spring Sadový) d | 1.7 ± 0.3 | 2.5 ± 1.2 | 4.2 ± 0.9 | 84.8 ± 3.2 | 5.1 ± 0.6 | 1.7 ± 0.3 |
| Geobacillus kaustophilus ML-1 (spring Mlýnský) d | 2.4 ± 0.3 | 3.3 ± 1.8 | 5.1 ± 1.6 | 81.1 ± 1.4 | 5.7 ± 1.7 | 2.4 ± 0.4 |
| Geobacillus stearothermophilus CCM 2062 (no information about source) | 0.9 ± 0.4 | 2.6 ± 0.9 | 3.5 ± 1.0 | 87.0 ± 2.1 | 4.3 ± 2.1 | 1.7 ± 0.4 |
| Geobacillus stearothermophilus CCM 5965 (Evaporated milk) | 0.9 ± 0.2 | 0.9 ± 0.5 | 2.7 ± 0.8 | 89.2 ± 1.7 | 3.6 ± 1.6 | 2.7 ± 0.3 |
| Geobacillus stearothermophilus ST-YPD (spring Štěpánka) d | 2.5 ± 0.6 | 3.4 ± 1.4 | 3.4 ± 1.4 | 84.0 ± 3.4 | 5.0 ± 1.4 | 1.7 ± 0.5 |
| Geobacillus stearothermophilus VR-1 (spring Vřídlo) d | 1.6 ± 0.5 | 2.5 ± 1.0 | 5.7 ± 1.6 | 82.0 ± 2.8 | 6.6 ± 2.1 | 1.6 ± 0.4 |
| Geobacillus thermoglucosidasius CCM 3731 (soil, Japan, Shimogamo, Kyoto) | 0.9 ± 0.5 | 1.8 ± 0.8 | 3.5 ± 0.8 | 88.5 ± 2.2 | 4.4 ± 1.2 | 0.9 ± 0.2 |
| Geobacillus thermoglucosidasius CCM 3732 (soil, Japan, Shimogamo, Kyoto) | 0.9 ± 0.2 | 1.8 ± 0.4 | 2.7 ± 0.9 | 90.1 ± 1.9 | 3.6 ± 1.3 | 0.9 ± 0.5 |
| Meiothermus ruber CCM 4211 (Thermal pools, Hveragherti, Iceland) | 1.8 ± 0.4 | 2.6 ± 1.6 | 3.5 ± 1.4 | 87.7 ± 5.8 | 4.4 ± 1.8 | 0.0 ± 0.0 |
| Meiothermus ruber CCM 4212 (Thermal pools, Hveragherti, Iceland) | 1.7 ± 0.7 | 2.6 ± 1.1 | 4.3 ± 2.0 | 87.0 ± 2.6 | 3.5 ± 1.1 | 0.9 ± 0.4 |
| Thermus aquaticus CCM 3485 (received from R. A. D. Williams collection, strain DI) | 0.0 ± 0.0 | 0.9 ± 0.4 | 2.8 ± 0.7 | 91.7 ± 3.1 | 2.8 ± 0.9 | 1.8 ± 0.3 |
| Thermus aquaticus CCM 3488 (source: Thermally polluted river near Brussels, Belgium) | 1.8 ± 0.3 | 2.7 ± 0.9 | 2.7 ± 0.9 | 88.4 ± 2.9 | 3.5 ± 0.8 | 0.9 ± 0.2 |
| Thermus sp. CCM 2842 (Hot spring of Kamchatka, Kamchatka, Russia) | 0.0 ± 0.0 | 0.9 ± 0.3 | 3.6 ± 1.3 | 89.2 ± 1.7 | 4.5 ± 2.3 | 1.8 ± 0.6 |
| Thermus sp. CCM 4167 (Hot spring Vřídlo, Carlsbad, Czech Republic) | 2.4 ± 0.5 | 3.3 ± 1.3 | 4.9 ± 1.2 | 81.3 ± 3.6 | 5.7 ± 1.7 | 2.4 ± 0.4 |
a Czech Collection of Microorganisms (CCM), Brno, Czech Republic. b The source(s) from which the strains were isolated according to data from the CCM collection. c Mean ± S.D. of three independent analyses. d Isolated bacteria in this study are marked in bold, see Table 1.
Table 3.
Content (relative %) of polyprenol derivatives from 16 strains of thermophilic bacteria obtained after saponification using reverse-phase liquid chromatography-positive electrospray mass spectrometry, see Materials and Methods.
Table 3.
Content (relative %) of polyprenol derivatives from 16 strains of thermophilic bacteria obtained after saponification using reverse-phase liquid chromatography-positive electrospray mass spectrometry, see Materials and Methods.
| Strain (Source) | C55-OH | C55-P | C55-PP |
|---|---|---|---|
| Alicyclobacillus acidoterrestris CCM a 4659 (apple-grape-raspberry juice) | 41 ± 4 | 17 ± 3 | 46 ± 7 |
| Alicyclobacillus acidoterrestris CCM 4660 (apple-grape-raspberry juice) | 36 ± 4 | 14 ± 6 | 53 ± 6 |
| Brevibacillus agri SA-1 (spring Sadový) | 34 ± 7 | 27 ± 4 | 52 ± 8 |
| Geobacillus kaustophilus ML-1 (spring Mlýnský) | 44 ± 8 | 31 ± 7 | 41 ± 11 |
| Geobacillus stearothermophilus CCM 2062 (no information about source) | 41 ± 5 | 24 ± 3 | 35 ± 7 |
| Geobacillus stearothermophilus CCM 5965 (evaporated milk) | 50 ± 8 | 19 ± 5 | 34 ± 8 |
| Geobacillus stearothermophilus ST-YPD (spring Štěpánka) | 43 ± 6 | 25 ± 9 | 46 ± 10 |
| Geobacillus stearothermophilus VR-1 (spring Vřídlo) | 42 ± 10 | 48 ± 7 | 44 ± 9 |
| Geobacillus thermoglucosidasius CCM 3731 (soil, Japan, Shimogamo, Kyoto) | 36 ± 4 | 12 ± 3 | 49 ± 7 |
| Geobacillus thermoglucosidasius CCM 3732 (soil, Japan, Shimogamo, Kyoto) | 51 ± 4 | 14 ± 5 | 32 ± 6 |
| Meiothermus ruber CCM 4211 (thermal pools, Hveragherti, Iceland) | 46 ± 7 | 17 ± 2 | 31 ± 5 |
| Meiothermus ruber CCM 4212 (thermal pools, Hveragherti, Iceland) | 49 ± 8 | 16 ± 4 | 37 ± 4 |
| Thermus aquaticus CCM 3485 (received from R. A. D. Williams collection, strain DI) | 57 ± 10 | 19 ± 3 | 28 ± 8 |
| Thermus aquaticus CCM 3488 (source: Thermally polluted river near Brussels, Belgium) | 57 ± 7 | 20 ± 6 | 25 ± 5 |
| Thermus sp. CCM 2842 (hot spring of Kamchatka, Kamchatka, Russia) | 57 ± 4 | 15 ± 5 | 26 ± 5 |
| Thermus sp. CCM 4167 (hot spring Vřídlo, Carlsbad, Czech Republic) | 48 ± 3 | 11 ± 5 | 24 ± 5 |
a Mean ± S.D. of three independent analyses. Values in nM/mg lyophilized cells were obtained by means of the calibrating three commercially available standards, see Materials and Methods. Isolated strains in this study are marked in bold.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Gharwalová, L.; Palyzová, A.; Marešová, H.; Kolouchová, I.; Kyselová, L.; Řezanka, T. Identification of Homologous Polyprenols from Thermophilic Bacteria. Microorganisms 2021, 9, 1168. https://doi.org/10.3390/microorganisms9061168
AMA Style
Gharwalová L, Palyzová A, Marešová H, Kolouchová I, Kyselová L, Řezanka T. Identification of Homologous Polyprenols from Thermophilic Bacteria. Microorganisms. 2021; 9(6):1168. https://doi.org/10.3390/microorganisms9061168
Chicago/Turabian StyleGharwalová, Lucia, Andrea Palyzová, Helena Marešová, Irena Kolouchová, Lucie Kyselová, and Tomáš Řezanka. 2021. "Identification of Homologous Polyprenols from Thermophilic Bacteria" Microorganisms 9, no. 6: 1168. https://doi.org/10.3390/microorganisms9061168
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.