
Fecal Microbiota Perspective for Evaluation of Prebiotic Potential of Bamboo Hemicellulose Hydrolysate in Mice: A Preliminary Study


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
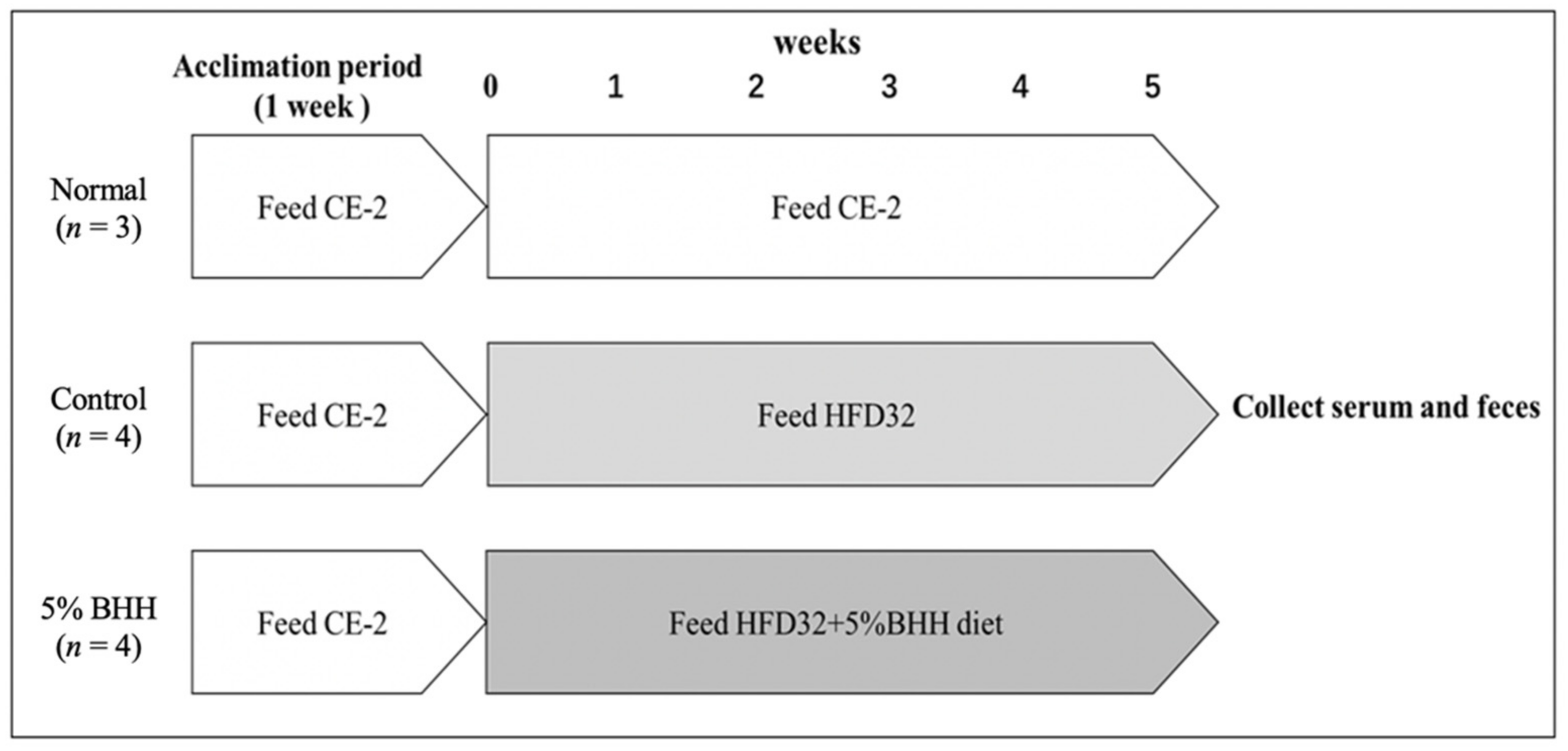
2.1. Animals and Experimental Design
2.2. Chemical Analysis
2.3. DNA Extraction from Feces
2.4. Microbiota Analysis by 16S rRNA Gene Sequencing
2.5. Statistical Analysis
3. Results
3.1. Effects of BHH on Body Weight, Food Intake, and Water Intake
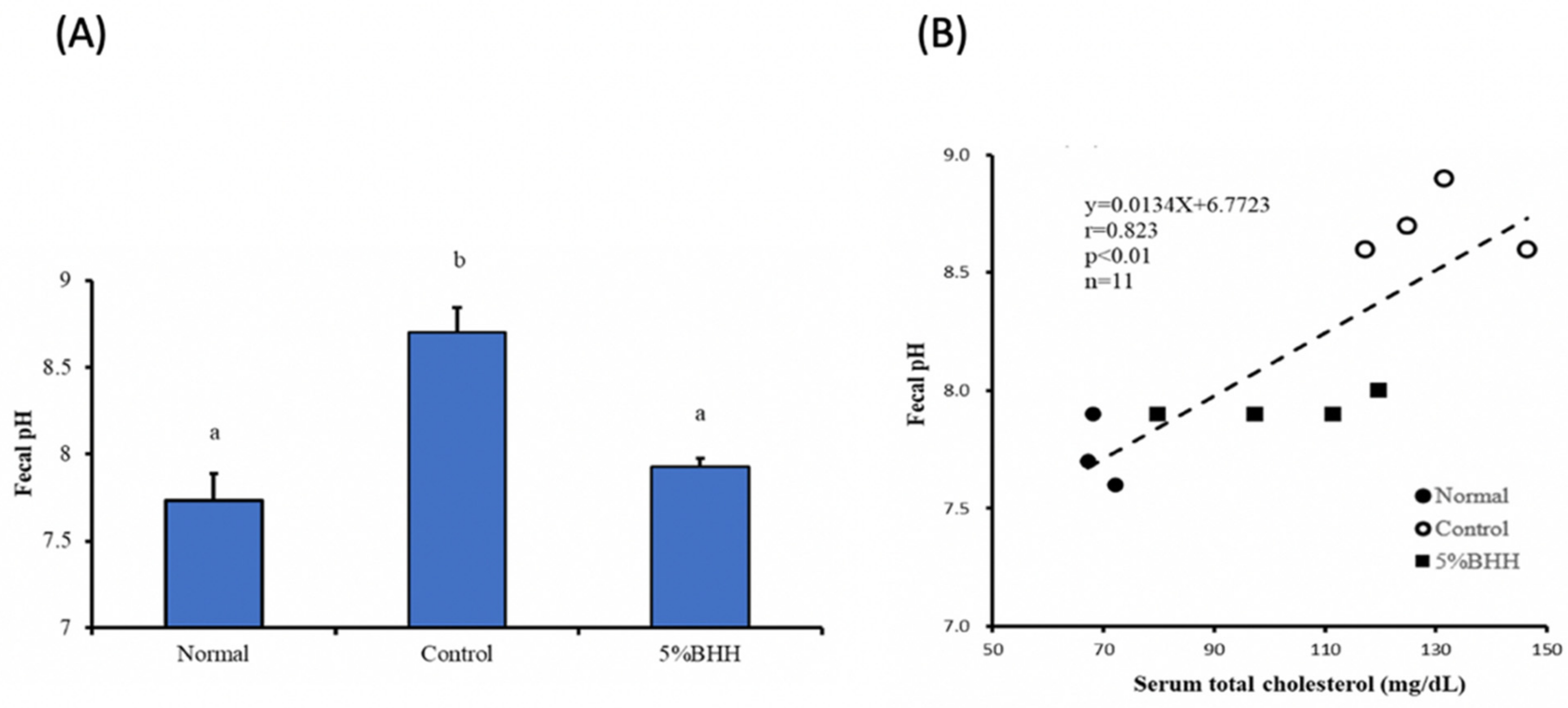
3.2. Effects of BHH on Serum Total Cholesterol, Triglycerides, Malondialdehydes (MDA), and Fecal pH
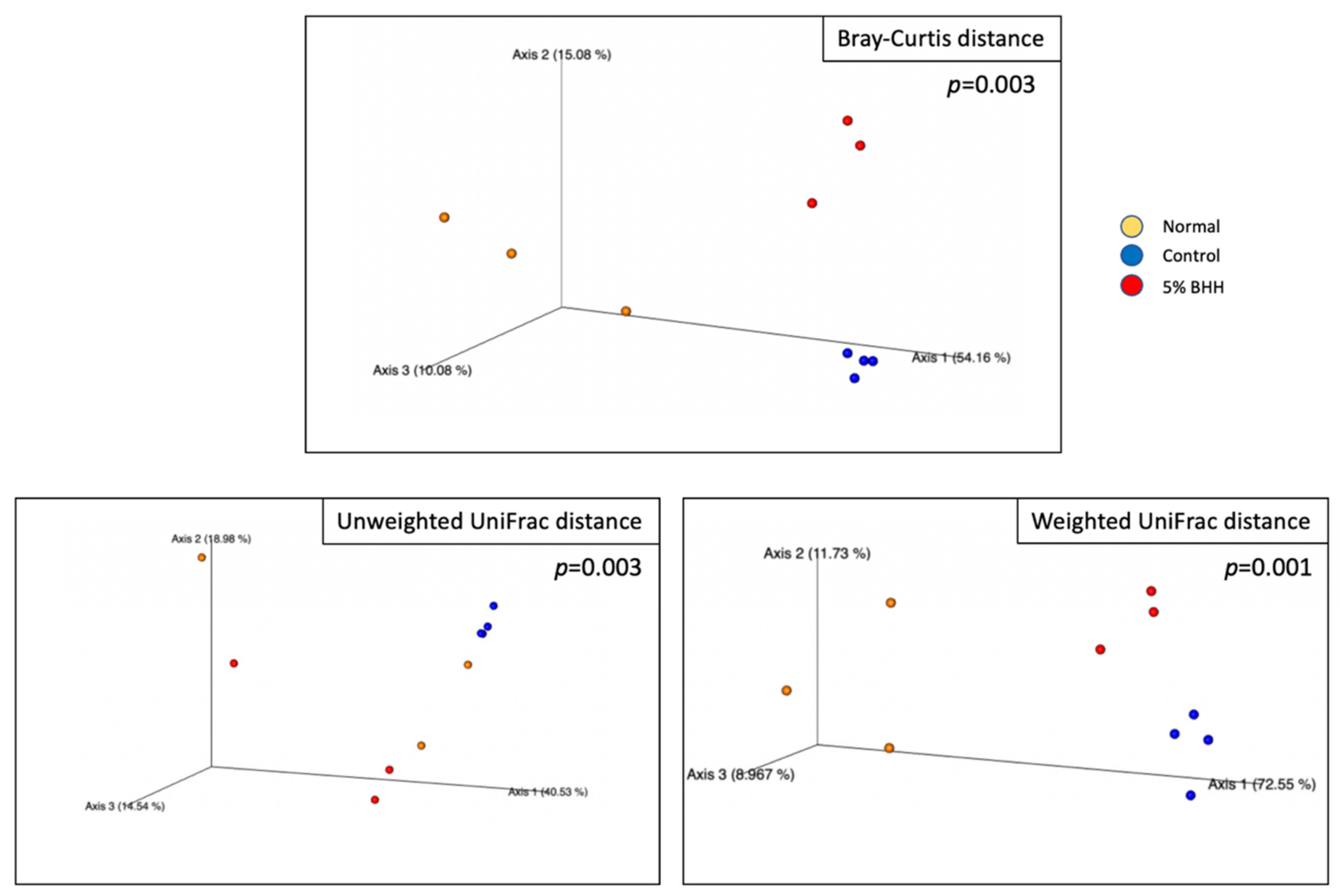
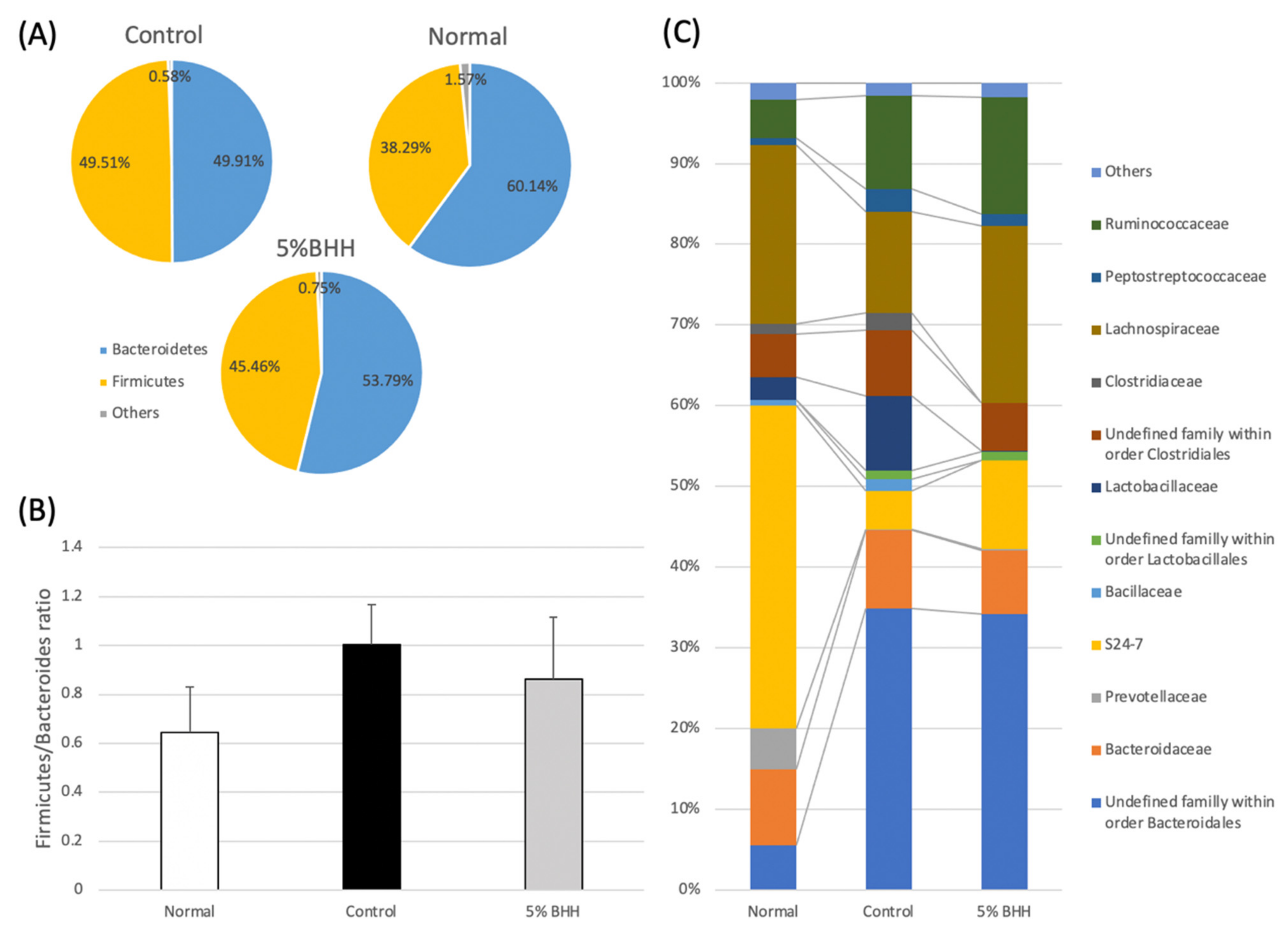
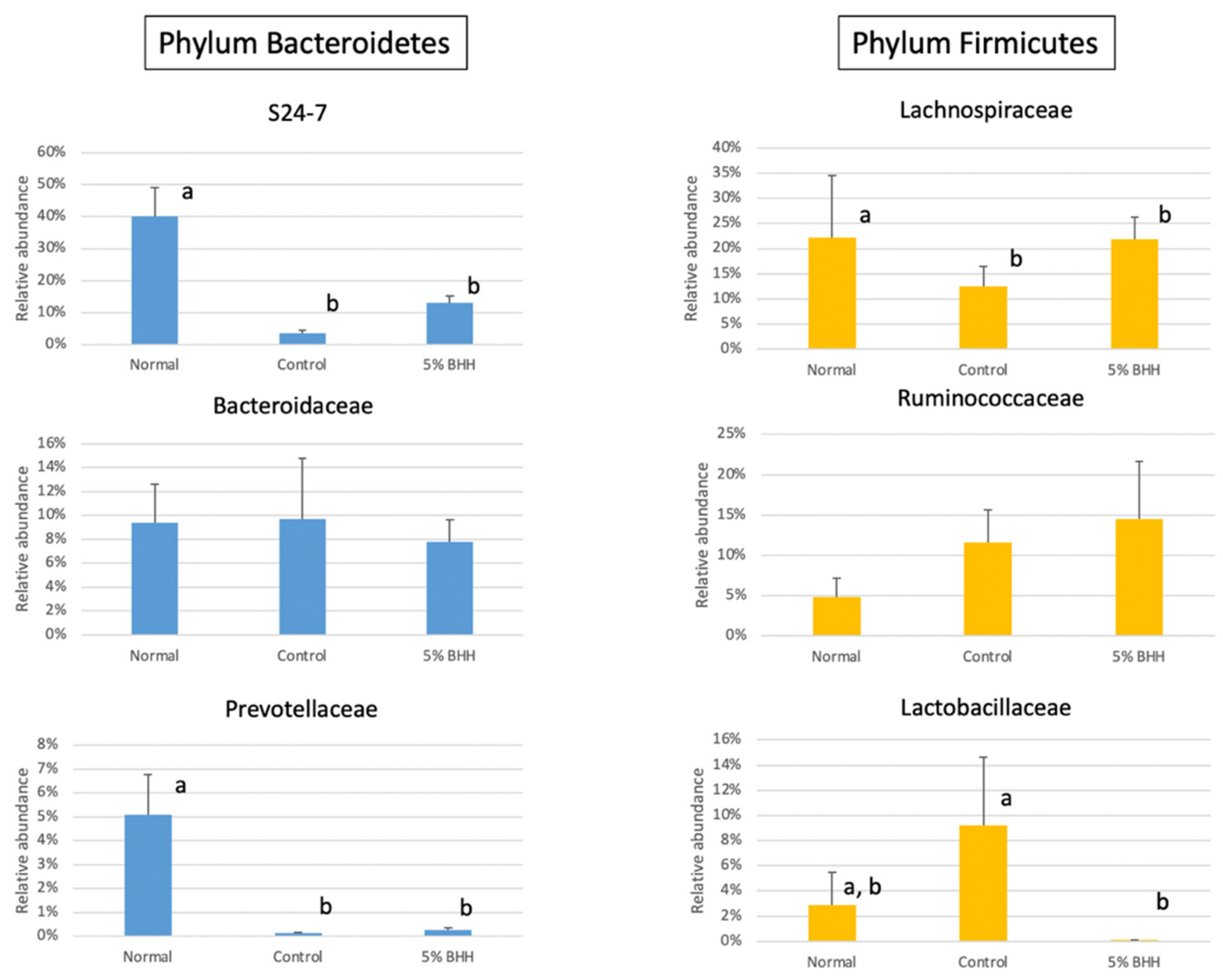
3.3. Effect of BHH on Fecal Microbiota Composition
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goyal, A.K.; Kar, P.; Sen, A. Advancement of bamboo taxonomy in the era of molecular biology: A review. In Biology of Useful Plants and Microbes; Sen, A., Ed.; Narosa Publication House: New Delhi, India, 2013; pp. 197–208. [Google Scholar]
- Goyal, A.K.; Brahma, B.K. Antioxidant and nutraceutical potential of bamboo: An overview. Int. J. Fund. Appl. Sci. 2014, 3, 2–10. [Google Scholar]
- Panee, J. Potential medicinal application and toxicity evaluation of extracts from bamboo plants. J. Med. Plant Res. 2015, 9, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Guo, J.; Ji, K.; Zhang, P. Bamboo shoot fiber prevents obesity in mice by modulating the gut microbiota. Sci. Rep. 2016, 6, 32953. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Duncan, S.H.; Louis, P.; Forano, E. Microbial degradation of complex carbohydrates in the gut. Gut Microbes 2012, 3, 289–306. [Google Scholar] [CrossRef] [Green Version]
- Koh, A.; De Vadder, F.; Kovatcheva-Datchary, P.; Bäckhed, F. From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites. Cell 2016, 165, 1332–1345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holscher, H.D. Dietary fiber and prebiotics and the gastrointestinal microbiota. Gut Microbes 2017, 8, 172–184. [Google Scholar] [CrossRef]
- Makki, K.; Deehan, E.C.; Walter, J.; Bäckhed, F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe 2018, 23, 705–715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maemura, M.; Horiuchi, M.; Abe, T.; Shiiba, K. Preparation of bamboo hemicellulose hydrolysate possessing anti-oxidative properties and their effects on mice plasma cholesterol. Food Sci. Technol. Res. 2016, 22, 537–543. [Google Scholar] [CrossRef] [Green Version]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16S rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [Green Version]
- Vazquez-Baeza, Y.; Pirrung, M.; Gonzalez, A.; Knight, R. EMPeror: A tool for visualizing high-throughput microbial community data. Gigascience 2013, 2, 16. [Google Scholar] [CrossRef] [Green Version]
- Tsikas, D. Assessment of lipid peroxidation by measuring malondialdehyde (MDA) and relatives in biological samples: Analytical and biological challenges. Anal. Biochem. 2017, 524, 13–30. [Google Scholar] [CrossRef]
- Ley, R.E.; Turnbaugh, P.J.; Klein, S.; Gordon, J.I. Microbial ecology: Human gut microbes associated with obesity. Nature 2006, 444, 1022–1023. [Google Scholar] [CrossRef]
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of high-fat diet on gut microbiota: A driving force for chronic disease risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515–520. [Google Scholar] [CrossRef]
- Lagkouvardos, I.; Lesker, T.R.; Hitch, T.C.A.; Gálvez, E.J.C.; Smit, N.; Neuhaus, K.; Wang, J.; Baines, J.F.; Abt, B.; Stecher, B.; et al. Sequence and cultivation study of Muribaculaceae reveals novel species, host preference, and functional potential of this yet undescribed family. Microbiome 2019, 7, 28. [Google Scholar] [CrossRef] [Green Version]
- Kodama, T.; Shiiba, K.; Tsuji, K. Suppressive effect of wheat- bran hemicellulose on blood pressure in spontaneously hypertensive rat. J. Jpn. Soc. Nutr. Food 1996, 49, 101–105. [Google Scholar] [CrossRef] [Green Version]
- Lopez, H.W.; Levrat, M.A.; Guy, C.; Messager, A.; Demigné, C.; Rémésy, C. Effects of soluble corn bran arabinoxylans on cecal digestion, lipid metabolism, and mineral balance (Ca, Mg) in rats. J. Nutr. Biochem. 1999, 10, 500–509. [Google Scholar] [CrossRef]
- Broekaert, W.F.; Courtin, C.M.; Verbeke, K.; Van de Wiele, T.; Verstraete, W.; Delcour, J.A. Prebiotic and other health-related effects of cereal-derived arabinoxylans, arabinoxylan-oligosaccharides, and xylooligosaccharides. Crit. Rev. Food Sci. Nutr. 2011, 51, 178–194. [Google Scholar] [CrossRef]
- Segata, N. Gut microbiome: Westernization and the disappearance of intestinal diversity. Curr. Biol. 2015, 25, R611–R613. [Google Scholar] [CrossRef] [Green Version]
- Liu, T.W.; Cephas, K.D.; Holscher, H.D.; Kerr, K.R.; Mangian, H.F.; Tappenden, K.A.; Swanson, K.S. Nondigestible fructans alter gastrointestinal barrier function, gene expression, histomorphology, and the microbiota profiles of diet-induced obese C57BL/6J mice. J. Nutr. 2016, 146, 949–956. [Google Scholar] [CrossRef]
- Ormerod, K.L.; Wood, D.L.; Lachner, N.; Gellatly, S.L.; Daly, J.N.; Parsons, J.D.; Dal’Molin, C.G.; Palfreyman, R.W.; Nielsen, L.K.; Cooper, M.A.; et al. Genomic characterization of the uncultured Bacteroidales family S24-7 inhabiting the guts of homeothermic animals. Microbiome 2016, 4, 36. [Google Scholar] [CrossRef] [Green Version]
- La Reau, A.J.; Suen, G. The Ruminococci: Key symbionts of the gut ecosystem. J. Microbiol. 2018, 56, 199–208. [Google Scholar] [CrossRef]
- Wolin, M.J.; Miller, T.L.; Collins, M.D.; Lawson, P.A. Formate-dependent growth and homoacetogenic fermentation by a bacterium from human feces: Description of Bryantella formatexigens gen. nov., sp. nov. Appl. Environ. Microbiol. 2003, 69, 6321–6326. [Google Scholar] [CrossRef] [Green Version]
- Wolin, M.J.; Miller, T.L.; Lawson, P.A. Proposal to replace the illegitimate genus name Bryantella Wolin et al. 2004VP with the genus name Marvinbryantia gen. nov. and to replace the illegitimate combination Bryantella formatexigens Wolin et al. 2004VP with Marvinbryantia formatexigens comb. nov. Int. J. Syst. Evol. Microbiol. 2008, 58, 742–744. [Google Scholar] [CrossRef]
- Grech-Mora, I.; Fardeau, M.L.; Patel, B.K.C.; Ollivier, B.; Rimbault, A.; Prensier, G.; Garcia, J.L.; Garnier-Sillam, E. Isolation and characterization of Sporobacter termitidis gen. nov., sp. nov., from the digestive tract of the wood-feeding termite Nasutitermes lujae. Int. J. Syst. Bacteriol. 1996, 46, 512–518. [Google Scholar] [CrossRef] [Green Version]
- Liu, S.; Qin, P.; Wang, J. High-fat diet alters the intestinal microbiota in streptozotocin-induced type 2 diabetic mice. Microorganisms 2019, 7, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azad, M.; Sarker, M.; Li, T.; Yin, J. Probiotic species in the modulation of gut microbiota: An overview. Biomed. Res. Int. 2018, 2018, 9478630. [Google Scholar] [CrossRef] [Green Version]
- Armougom, F.; Henry, M.; Vialettes, B.; Raccah, D.; Raoult, D. Monitoring bacterial community of human gut microbiota reveals an increase in Lactobacillus in obese patients and Methanogens in anorexic patients. PLoS ONE 2009, 4, e7125. [Google Scholar] [CrossRef]
- Eckburg, P.B.; Bik, E.M.; Bernstein, C.N.; Purdom, E.; Dethlefsen, L.; Sargent, M.; Gill, S.R.; Nelson, K.E.; Relman, D.A. Diversity of the human intestinal microbial flora. Science 2005, 308, 1635–1638. [Google Scholar] [CrossRef] [Green Version]
- Singhal, P.; Bal, L.M.; Satya, S.; Sudhakar, P.; Naik, S.N. Bamboo shoots: A novel source of nutrition and medicine. Crit. Rev. Food Sci. Nutr. 2013, 5, 517–534. [Google Scholar] [CrossRef]
- Shen, M.; Xie, Z.; Jia, M.; Li, A.; Han, H.; Wang, T.; Zhang, L. Effect of bamboo leaf extract on antioxidant status and cholesterol metabolism in broiler chickens. Animals 2019, 9, 699. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.; Li, Z.; Cao, G.; Huang, S.; Yang, H. Bamboo leaf flavonoids extracts alleviate oxidative stress in Hepg2 cells via naturally modulating reactive oxygen species production and nrf2-mediated antioxidant defense responses. J. Food Sci. 2019, 84, 1609–1620. [Google Scholar] [CrossRef] [PubMed]
- Luo, X.; Wang, Q.; Zheng, B.; Lin, L.; Chen, B.; Zheng, Y.; Xiao, J. Hydration properties and binding capacities of dietary fibers from bamboo shoot shell and its hypolipidemic effects in mice. Food Chem. Toxicol. 2017, 109, 1003–1009. [Google Scholar] [CrossRef] [PubMed]
- Wu, W.; Hu, J.; Gao, H.; Chen, H.; Fang, X.; Mu, H.; Han, Y.; Liu, R. The potential cholesterol-lowering and prebiotic effects of bamboo shoot dietary fibers and their structural characteristics. Food Chem. 2020, 332, 127372. [Google Scholar] [CrossRef]
- Zhao, Z.; Egashira, Y.; Sanada, H. Ferulic acid sugar esters are recovered in rat plasma and urine mainly as the sulfoglucuronide of ferulic acid. J. Nutr. 2003, 133, 1355–1361. [Google Scholar] [CrossRef]
- Walker, A.W.; Duncan, S.H.; McWilliam Leitch, E.C.; Child, M.W.; Flint, H.J. pH and peptide supply can radically alter bacterial populations and short-chain fatty acid ratios within microbial communities from the human colon. Appl. Environ. Microbiol. 2005, 71, 3692–3700. [Google Scholar] [CrossRef] [Green Version]
- Duncan, S.H.; Belenguer, A.; Holtrop, G.; Johnstone, A.M.; Flint, H.J.; Lobley, G.E. Reduced dietary intake of carbohydrates by obese subjects results in decreased concentrations of butyrate and butyrate-producing bacteria in feces. Appl. Environ. Microbiol. 2007, 73, 1073–1078. [Google Scholar] [CrossRef] [Green Version]
- Ikeyama, N.; Murakami, T.; Toyoda, A.; Mori, H.; Iino, T.; Ohkuma, M.; Sakamoto, M. Microbial interaction between the succinate-utilizing bacterium Phascolarctobacterium faecium and the gut commensal Bacteroides thetaiotaomicron. Microbiologyopen 2020, 9, e1111. [Google Scholar] [CrossRef]
- Lin, L.; Zhang, J. Role of intestinal microbiota and metabolites on gut homeostasis and human diseases. BMC Immunol. 2017, 18, 2. [Google Scholar] [CrossRef] [Green Version]
- Hara, H.; Haga, S.; Aoyama, Y.; Kiriyama, S. Short-chain fatty acids suppress cholesterol synthesis in rat liver and intestine. J. Nutr. 1999, 129, 942–948. [Google Scholar] [CrossRef] [Green Version]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets | Final Body Weight (g) | Weight Gain (g/5 weeks) | Food Intake | Water Intake (mL/5 weeks) | |
|---|---|---|---|---|---|
| (g/5 weeks) | (kcal/5 weeks) | ||||
| Normal | 24.98 ± 1.40 a | 1.75 ± 1.35 a | 104.35 ± 2.25 | 355.26 ± 7.66 a | 266.0 ± 18.0 |
| Control | 30.95 ± 1.69 b | 7.60 ± 2.35 b | 91.30 ± 11.8 | 463.43 ± 59.89 b | 240.0 ± 12.0 |
| 5% BHH | 29.87 ± 2.31 b | 6.75 ± 1.35 b | 89.92 ± 15.85 | 442.60 ± 77.94 b | 245.0 ± 16.0 |
| Diets | Total Cholesterol (mg/mL) | Triglyceride (mg/mL) | MDA (µM) |
|---|---|---|---|
| Normal | 69.19 ± 2.68 a | 56.53 ± 6.11 | 28.48 ± 4.49 a |
| Control | 129.96 ± 12.40 b | 62.70 ± 4.12 | 36.32 ± 4.83 b |
| 5% BHH | 102.04 ± 17.52 c | 59.70 ± 21.56 | 34.08 ± 7.65 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikeyama, N.; Sakamoto, M.; Ohkuma, M.; Hiramoto, S.; Wang, J.; Tone, S.; Shiiba, K. Fecal Microbiota Perspective for Evaluation of Prebiotic Potential of Bamboo Hemicellulose Hydrolysate in Mice: A Preliminary Study. Microorganisms 2021, 9, 888. https://doi.org/10.3390/microorganisms9050888
Ikeyama N, Sakamoto M, Ohkuma M, Hiramoto S, Wang J, Tone S, Shiiba K. Fecal Microbiota Perspective for Evaluation of Prebiotic Potential of Bamboo Hemicellulose Hydrolysate in Mice: A Preliminary Study. Microorganisms. 2021; 9(5):888. https://doi.org/10.3390/microorganisms9050888
Chicago/Turabian StyleIkeyama, Nao, Mitsuo Sakamoto, Moriya Ohkuma, Shigeru Hiramoto, Jianpeng Wang, Shigenobu Tone, and Kiwamu Shiiba. 2021. "Fecal Microbiota Perspective for Evaluation of Prebiotic Potential of Bamboo Hemicellulose Hydrolysate in Mice: A Preliminary Study" Microorganisms 9, no. 5: 888. https://doi.org/10.3390/microorganisms9050888