
Coordinated Expression of HPV-6 Genes with Predominant E4 and E5 Expression in Laryngeal Papilloma

 ,
, 
 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Detection of Human Papillomavirus (HPV) DNA and Quantitative Polymerase Chain Reaction (PCR) Analysis of HPV-6 Viral Load
2.2.1. Extraction of Genomic DNA from Laryngeal Papilloma (LP) and PCR Conditions
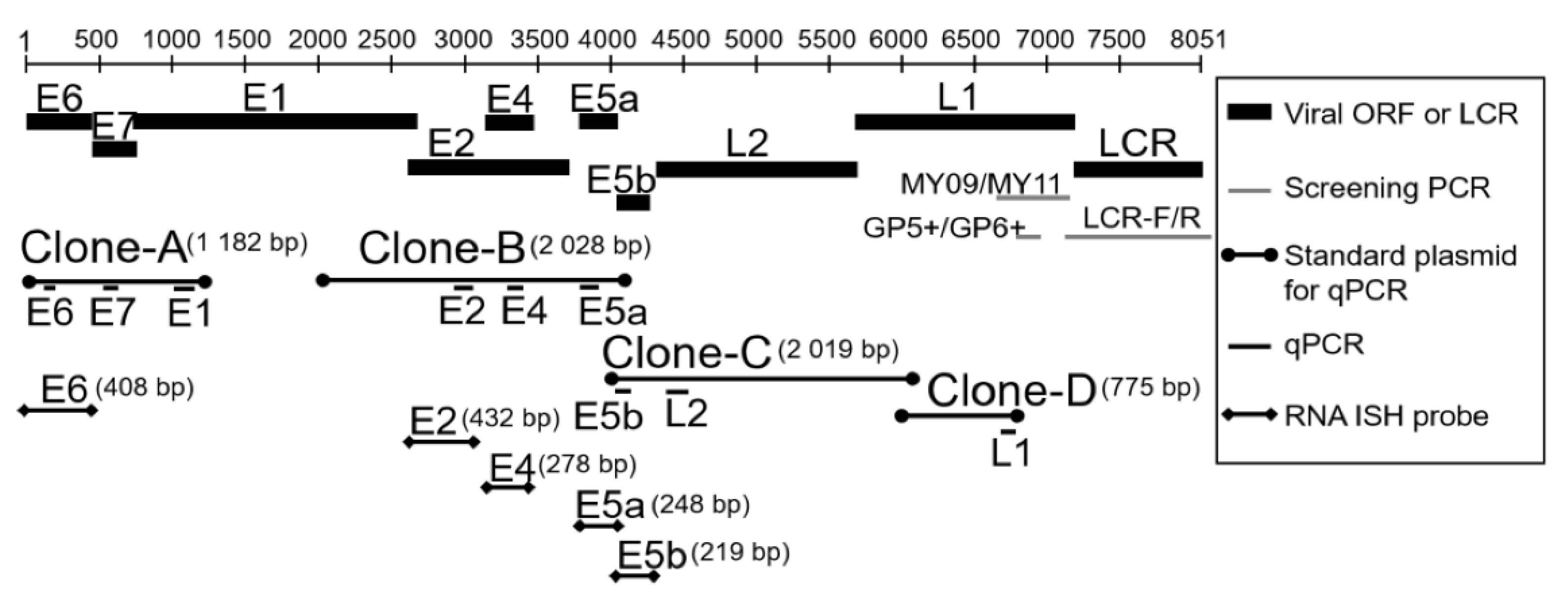
2.2.2. Partial Cloning of the HPV-6-LCR Region and Identification of HPV-6 Subtypes
2.2.3. Quantitative PCR Analysis of HPV-6 Viral Load in LP
2.3. Measurement of Viral mRNA Expression in HPV-6-infected LP by Quantitative Real-Time PCR
2.4. In Situ Hybridization (ISH) with HPV DNA Probes
2.5. RNA-ISH with HPV-6 E6, E2, E4, E5a, and E5b Digoxigenin RNA Probes
2.6. Double Fluorescence RNA-ISH
2.7. Immunohistochemistry for HPV-6 E4
2.7.1. Preparation of an Anti-HPV-6 E4 Monoclonal Antibody
2.7.2. Evaluation of Antibody Specificity
2.7.3. Immunohistochemistry for HPV-6 E1^E4
2.8. Statistical Analysis
2.9. Ethics Approval and Consent to Participate
3. Results
3.1. HPV DNA Distribution, Subtypes, and Viral Load in LP
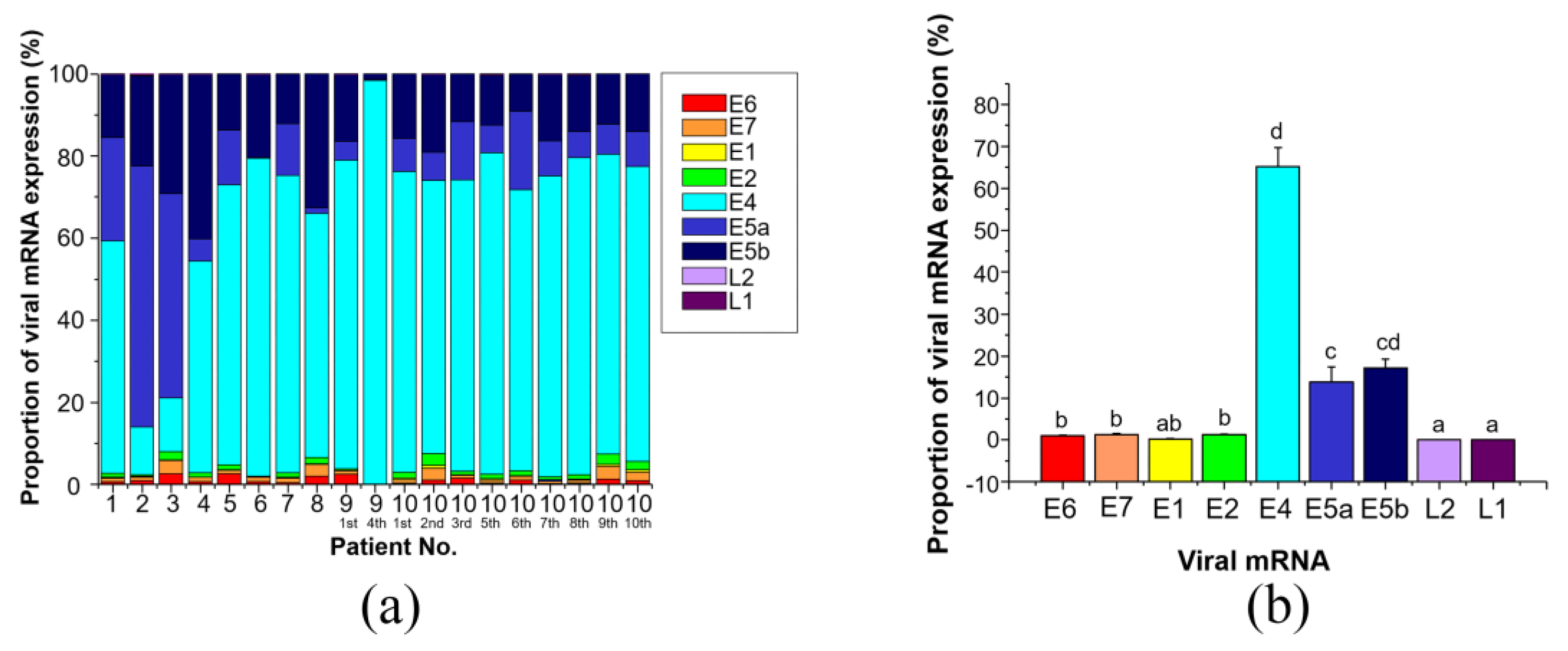
3.2. Expression of HPV-6 mRNAs in LP
3.2.1. Expression Levels of the 9 Viral mRNAs
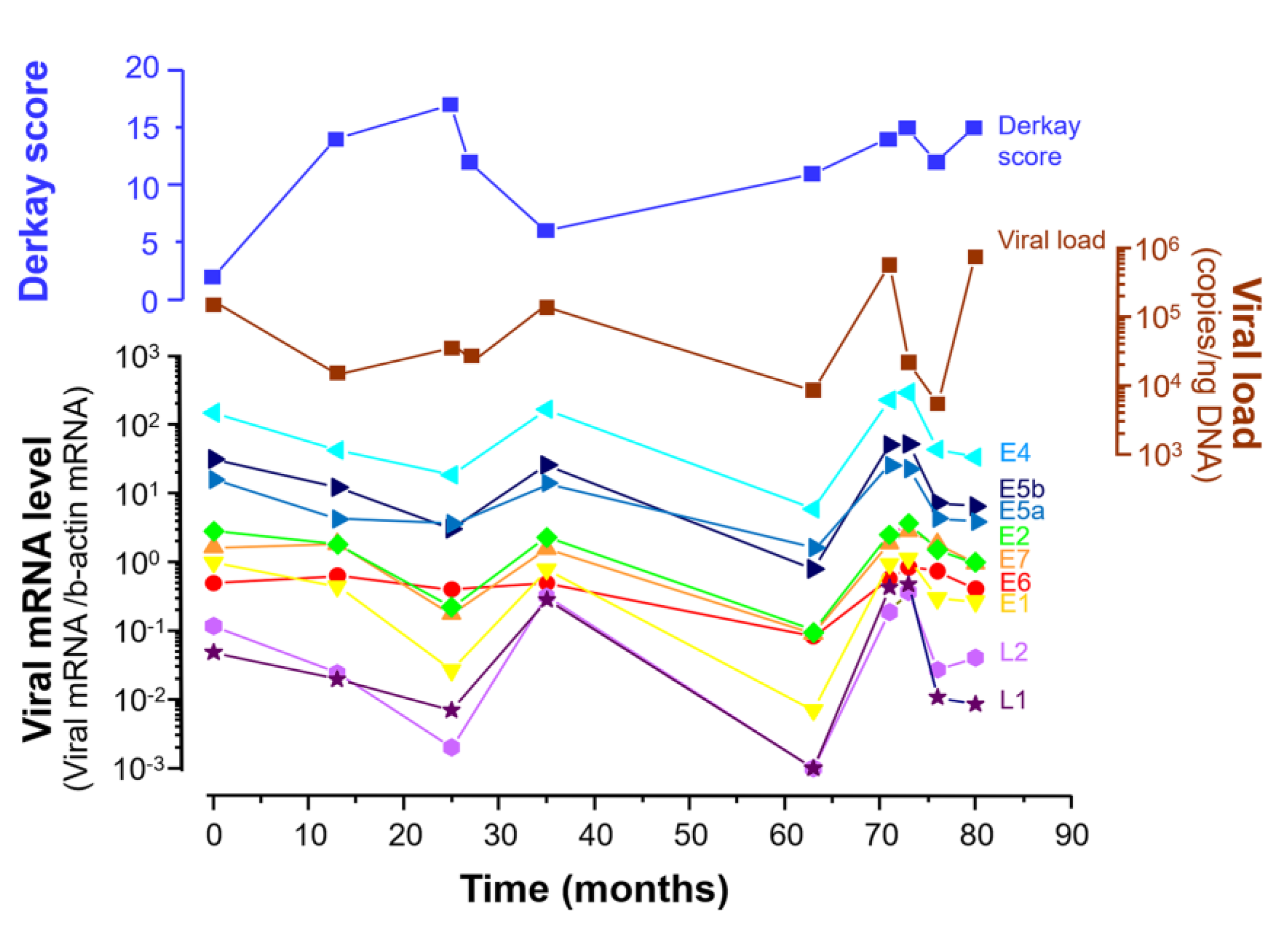
3.2.2. Viral Load Alteration and Levels of the 9 Viral mRNAs in LP during 80 Months
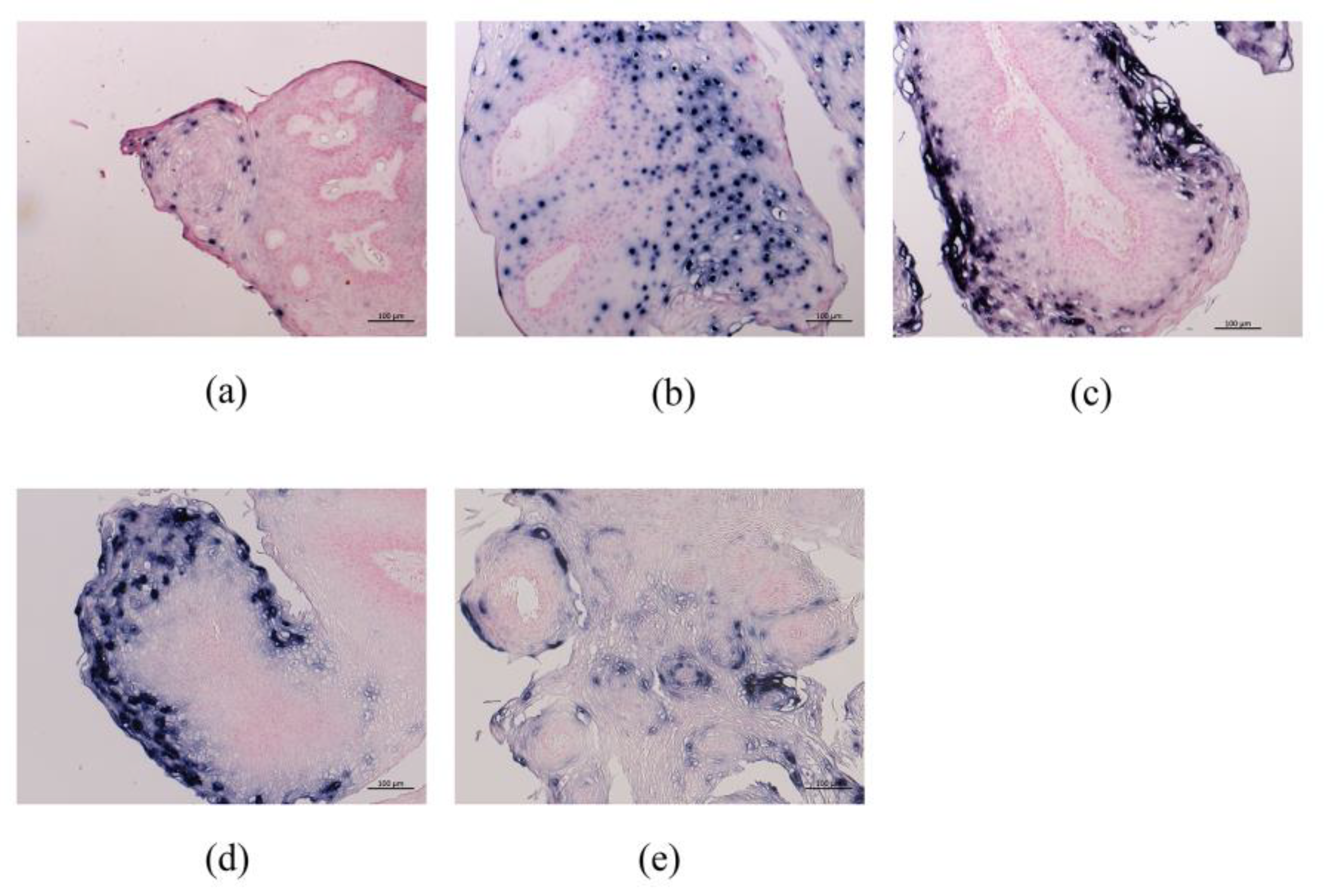
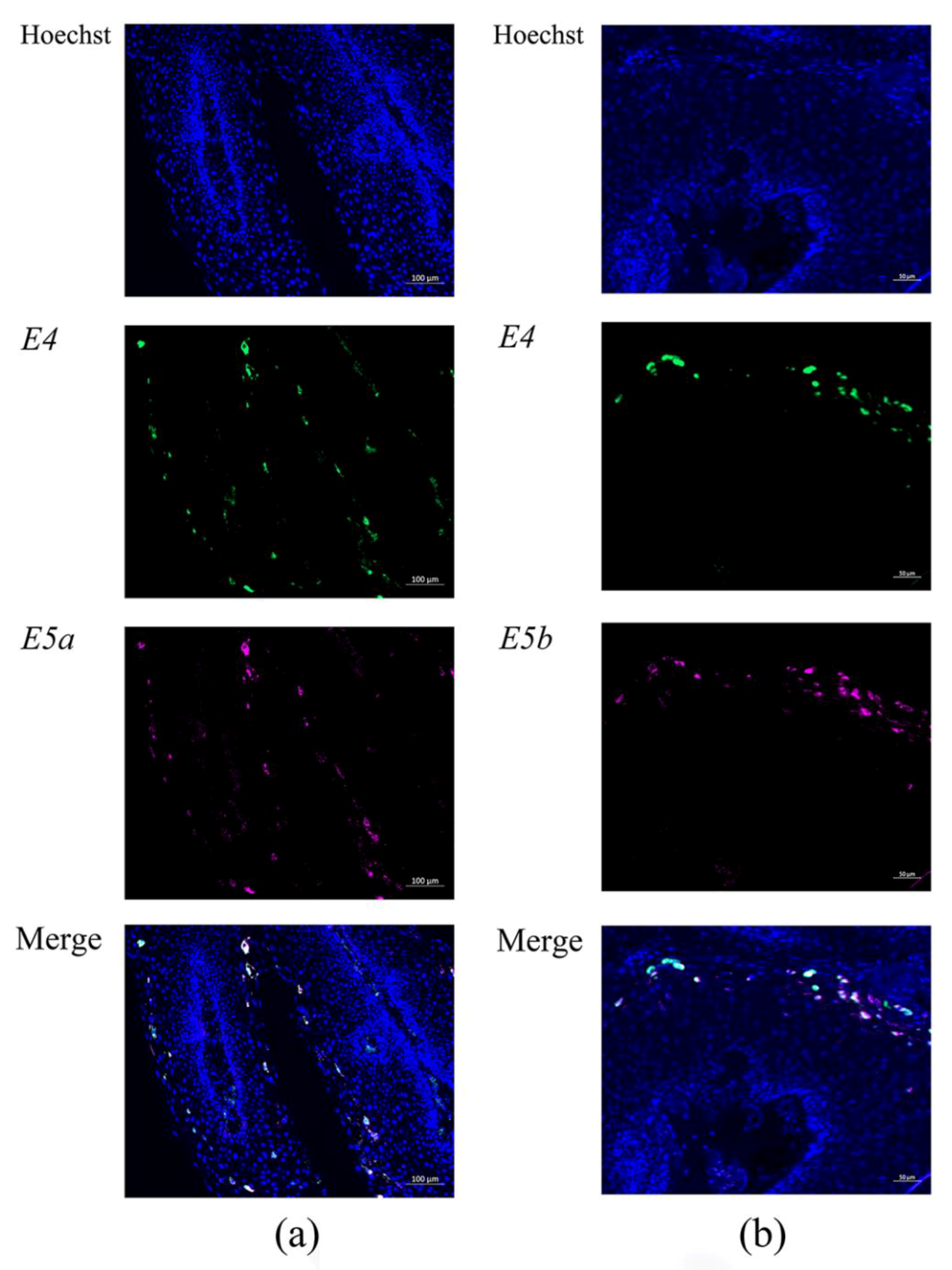
3.2.3. HPV-6 E6, E2, E4, E5a, and E5b mRNA Expression in LP Detected by RNA-ISH
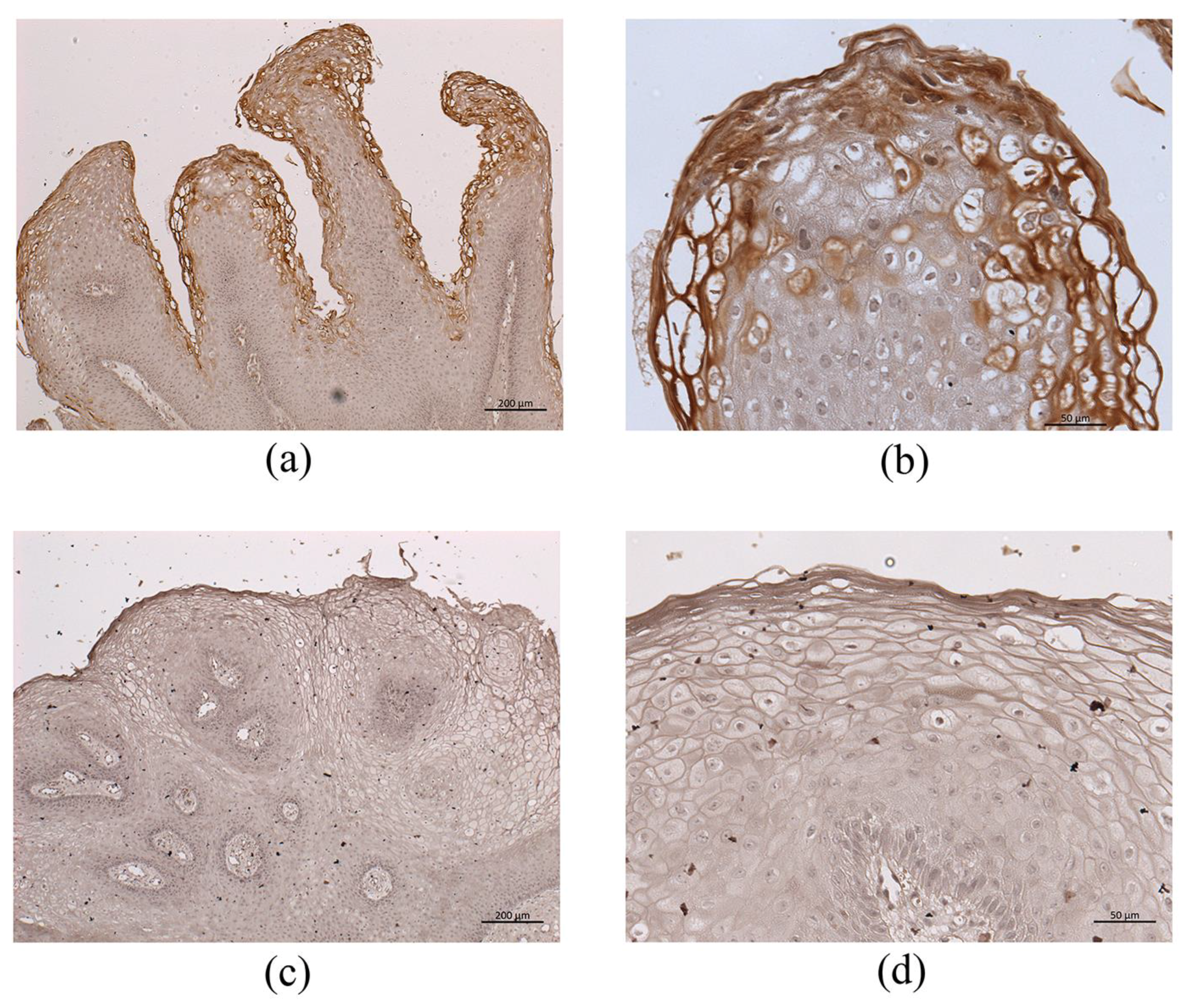
3.3. Immunohistochemistry for HPV-6 E1^E4
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fortes:, H.R.; von Ranke, F.M.; Escuissato, D.L.; Araujo Neto, C.A.; Zanetti, G.; Hochhegger, B.; Souza, C.A.; Marchiori, E. Recurrent respiratory papillomatosis: A state-of-the-art review. Respir. Med. 2017, 126, 116–121. [Google Scholar] [CrossRef] [Green Version]
- Kanazawa, T.; Fukushima, N.; Imayoshi, S.; Nagamoto, T.; Kawada, K.; Nishino, H.; Ichimura, K. Rare case of malignant transformation of recurrent respiratory papillomatosis associated with human papillomavirus type 6 infection and p53 overexpression. SpringerPlus 2013, 2, 153. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schraff, S.; Derkay, C.S.; Burke, B.; Lawson, L. American society of pediatric otolaryngology members’ experience with recurrent respiratory papillomatosis and the use of adjuvant therapy. Arch. Otolaryngol. Head Neck Surg. 2004, 130, 1039–1042. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baumann, J.L.; Cohen, S.; Evjen, A.N.; Law, J.H.; Vadivelu, S.; Attia, A.; Schindler, J.S.; Chung, C.H.; Wirth, P.S.; Meijer, C.J.L.M.; et al. Human papillomavirus in early laryngeal carcinoma. Laryngoscope 2009, 119, 1531–1537. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J.; Quint, W.; Banks, L.; Bravo, I.G.; Stoler, M.; Broker, T.R.; Stanley, M.A. The biology and life-cycle of human papillomaviruses. Vaccine 2012, 30 (Suppl. 5), F55–F70. [Google Scholar] [CrossRef]
- Woodman, C.B.; Collins, S.I.; Young, L.S. The natural history of cervical hpv infection: Unresolved issues. Nat. Rev. Cancer 2007, 7, 11–22. [Google Scholar] [CrossRef]
- Ribeiro, A.L.; Caodaglio, A.S.; Sichero, L. Regulation of hpv transcription. Clinics (Sao Paulo) 2018, 73, e486s. [Google Scholar] [CrossRef]
- Graham, S.V. The human papillomavirus replication cycle, and its links to cancer progression: A comprehensive review. Clin. Sci. 2017, 131, 2201–2221. [Google Scholar] [CrossRef] [Green Version]
- Egawa, N.; Doorbar, J. The low-risk papillomaviruses. Virus Res. 2017, 231, 119–127. [Google Scholar] [CrossRef]
- Doorbar, J. The e4 protein; structure, function and patterns of expression. Virology 2013, 445, 80–98. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Xue, Y.; Poidinger, M.; Lim, T.; Chew, S.H.; Pang, C.L.; Abastado, J.P.; Thierry, F. Mapping of hpv transcripts in four human cervical lesions using rnaseq suggests quantitative rearrangements during carcinogenic progression. Virology 2014, 462–463, 14–24. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsai, T.C.; Chen, S.L. The biochemical and biological functions of human papillomavirus type 16 e5 protein. Arch. Virol. 2003, 148, 1445–1453. [Google Scholar] [CrossRef]
- Ilahi, N.E.; Bhatti, A. Impact of hpv e5 on viral life cycle via egfr signaling. Microb. Pathog. 2020, 139, 103923. [Google Scholar] [CrossRef]
- Venuti, A.; Paolini, F.; Nasir, L.; Corteggio, A.; Roperto, S.; Campo, M.; Borzacchiello, G. Papillomavirus E5: The smallest oncoprotein with many functions. Mol. Cancer 2011, 10, 140. [Google Scholar] [CrossRef] [Green Version]
- Ikegami, T.; Uehara, T.; Deng, Z.; Kondo, S.; Maeda, H.; Kiyuna, A.; Agena, S.; Hirakawa, H.; Yamashita, Y.; Ganaha, A.; et al. Detection of human papillomavirus in branchial cleft cysts. Oncol. Lett. 2018, 16, 1571–1578. [Google Scholar] [CrossRef] [Green Version]
- Deng, Z.; Hasegawa, M.; Matayoshi, S.; Kiyuna, A.; Yamashita, Y.; Maeda, H.; Suzuki, M. Prevalence and clinical features of human papillomavirus in head and neck squamous cell carcinoma in okinawa, southern japan. Eur. Arch. Otorhinolaryngol. 2011, 268, 1625–1631. [Google Scholar] [CrossRef] [PubMed]
- Deng, Z.; Ikegami, T.; Kiyuna, A.; Zhang, C.; Zhang, T.; Matayoshi, S.; Uehara, T.; Maeda, H.; Suzuki, M.; Ganaha, A. Methylation of cpg sites in the upstream regulatory region, physical status and mrna expression of hpv-6 in adult-onset laryngeal papilloma. Oncotarget 2017, 8, 85368–85377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakayama, H.; Murakami, A.; Yoshida, M.; Muraoka, J.; Wakai, J.; Kenjyou, N.; Ito, Y. Characterization and selection of 3-(1-naphthoyl)-indole derivative-specific alpaca vhh antibodies using a phage display library. Monoclon. Antib. Immunodiagn Immunother 2016, 35, 231–234. [Google Scholar] [CrossRef]
- Miyazaki, A.; Yogosawa, S.; Murakami, A.; Kitamura, D. Identification of cmtm7 as a transmembrane linker of blnk and the b-cell receptor. PLoS ONE 2012, 7, e31829. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Derkay, C.S.; Hester, R.P.; Burke, B.; Carron, J.; Lawson, L. Analysis of a staging assessment system for prediction of surgical interval in recurrent respiratory papillomatosis. Int. J. Pediatr. Otorhinolaryngol. 2004, 68, 1493–1498. [Google Scholar] [CrossRef] [PubMed]
- Jelen, M.M.; Chen, Z.; Kocjan, B.J.; Burt, F.J.; Chan, P.K.; Chouhy, D.; Combrinck, C.E.; Coutlée, F.; Estrade, C.; Ferenczy, A.; et al. Global genomic diversity of human papillomavirus 6 based on 724 isolates and 190 complete genome sequences. J. Virol. 2014, 88, 7307–7316. [Google Scholar] [CrossRef] [Green Version]
- Szinai, M.; Nagy, Z.; Máté, P.; Kovács, D.; Laczkó, L.; Kardos, G.; Sápy, T.; Szűcs, A.; Szarka, K. Comparative analysis of human papillomavirus type 6 complete genomes originated from head and neck and anogenital disorders. Infect. Genet. Evol. 2019, 71, 140–150. [Google Scholar] [CrossRef]
- Kocjan, B.J.; Gale, N.; Hočevar Boltežar, I.; Seme, K.; Fujs Komloš, K.; Hošnjak, L.; Maver, P.J.; Jelen, M.M.; Zupanič Pajnič, I.; Balažic, J.; et al. Identical human papillomavirus (HPV) genomic variants persist in recurrent respiratory papillomatosis for up to 22 years. J. Infect. Dis. 2013, 207, 583–587. [Google Scholar] [CrossRef] [Green Version]
- Doorbar, J.; Egawa, N.; Griffin, H.; Kranjec, C.; Murakami, I. Human papillomavirus molecular biology and disease association. Rev. Med. Virol. 2015, 25 (Suppl. 1), 2–23. [Google Scholar] [CrossRef] [Green Version]
- Bedard, M.C.; Brusadelli, M.G.; Carlile, A.; Ruiz-Torres, S.; Lodin, H.; Lee, D.; Kofron, M.; Lambert, P.F.; Lane, A.; Ameziane, N.; et al. Patient-derived organotypic epithelial rafts model phenotypes in juvenile-onset recurrent respiratory papillomatosis. Viruses 2021, 13, 68. [Google Scholar] [CrossRef] [PubMed]
- Doorbar, J. Host control of human papillomavirus infection and disease. Best Pract. Res. Clin. Obstet. Gynaecol. 2018, 47, 27–41. [Google Scholar] [CrossRef] [Green Version]
- DiMaio, D.; Petti, L.M. The E5 proteins. Virology 2013, 445, 99–114. [Google Scholar] [CrossRef]
- Faulkner, G.; Banks, L. The human papillomavirus (HPV)-6 and HPV-16 E5 proteins co-operate with HPV-16 E7 in the transformation of primary rodent cells. J. Gen. Virol. 1995, 76, 1239–1245. [Google Scholar]
- Wlazlo, A.P.; Sparkowski, J.J.; Jenson, A.B.; Schlegel, R. Expression of the bovine papillomavirus type 1 E5b gene reveals a protein–protein interaction of the E5a and E5b gene products. Virology 2003, 307, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Griffin, H.; Soneji, Y.; Van Baars, R.; Arora, R.; Jenkins, D.; van de Sandt, M.; Wu, Z.; Quint, W.; Jach, R.; Okon, K.; et al. Stratification of HPV-induced cervical pathology using the virally encoded molecular marker E4 in combination with p16 or MCM. Mod. Pathol. 2015, 28, 977–993. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Case | Age (Years) | Sex | Surgery | No. of Tumors | Subsite | Derkay Score | HPV Subtype |
|---|---|---|---|---|---|---|---|
| 1 | 27 | M | 3rd | Multiple | B, TVC; B, FVC | 15 | HPV-6a |
| 2 | 43 | M | 1st | multiple | B, TVC; L, FVC | 13 | HPV-6b |
| 3 | 34 | M | 2nd | multiple | B, FVC | 6 | HPV-6b |
| 4 | 41 | M | 1st | multiple | B, TVC | 4 | HPV-6vc |
| 5 | 41 | M | 1st | single | R, TVC | 2 | HPV-6vc |
| 6 | 17 | F | 1st | multiple | B, TVC | 6 | HPV-6vc |
| 7 | 28 | M | 1st | multiple | B, TVC | 2 | HPV6-a |
| 8 | 4 | F | 1st | multiple | B, TVC | 6 | HPV-6vc |
| 9 | 59 | M | 1st | multiple | B, TVC | 6 | HPV6-vc |
| 61 | 4th | multiple | B, TVC | 3 | |||
| 10 | 67 | M | 1st | multiple | B, TVC | 2 | HPV6-vc |
| 68 | 2nd | multiple | B, TVC | 14 | |||
| 69 | 3rd | multiple | L, TVC | 17 | |||
| 69 | 4th | multiple | B, TVC | 12 | |||
| 71 | 5th | multiple | B, TVC | 6 | |||
| 72 | 6th | multiple | B, TVC | 11 | |||
| 73 | 7th | multiple | B, TVC | 14 | |||
| 73 | 8th | multiple | B, TVC | 15 | |||
| 73 | 9th | multiple | B, TVC | 12 | |||
| 74 | 10th | multiple | B, TVC | 15 | |||
| 11 | 45 | M | 1st | multiple | B, TVC | 6 | HPV-6 |
| 12 | 44 | M | 1st | single | R, TVC | 3 | HPV-6a |
| 13 | 44 | M | 1st | single | L, TVC | 2 | HPV-6 |
| Case | Surgery | Viral Load (Copies/ng DNA) | HPV mRNA Expression/β-actin | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| E6 | E7 | E1 | E2 | E4 | E5a | E5b | L2 | L1 | |||
| 1 | 3rd | 2,268,452 | 0.110 | 0.179 | 0.026 | 0.185 | 10.675 | 4.766 | 2.900 | 0.010 | 0.012 |
| 2 | 1st | 649,319 | 0.335 | 0.312 | 0.105 | 0.159 | 4.496 | 24.633 | 8.496 | 0.062 | 0.134 |
| 3 | 2nd | 436 | 0.134 | 0.163 | 0.010 | 0.109 | 0.677 | 2.585 | 1.513 | 0.002 | 0.006 |
| 4 | 1st | 21,372 | 0.167 | 0.317 | 0.000 | 0.317 | 14.643 | 1.524 | 11.429 | 0.000 | 0.017 |
| 5 | 1st | 25,347 | 0.425 | 0.153 | 0.010 | 0.213 | 11.474 | 2.241 | 2.309 | 0.001 | 0.002 |
| 6 | 1st | 4739 | 0.135 | 0.218 | 0.003 | 0.059 | 16.934 | 0.004 | 4.460 | 0.000 | 0.010 |
| 7 | 1st | 14,170 | 0.032 | 0.100 | 0.006 | 0.099 | 6.175 | 1.068 | 1.035 | 0.000 | 0.001 |
| 8 | 1st | 36,408 | 0.394 | 0.554 | 0.018 | 0.322 | 11.688 | 0.278 | 6.403 | 0.002 | 0.004 |
| 9 | 1st | 2,199,081 | 0.587 | 0.208 | 0.019 | 0.122 | 18.409 | 1.108 | 4.032 | 0.002 | 0.008 |
| 4th | 2,918,599 | 0.542 | 2.799 | 0.693 | 5.824 | 20,828.885 | 74.591 | 256.091 | 6.180 | 0.775 | |
| 10 | 1st | 166,859 | 0.501 | 1.627 | 0.991 | 2.838 | 149.595 | 16.359 | 32.002 | 0.118 | 0.048 |
| 2nd | 17,173 | 0.634 | 1.841 | 0.448 | 1.806 | 42.551 | 4.378 | 12.183 | 0.025 | 0.020 | |
| 3rd | 39,002 | 0.401 | 0.177 | 0.027 | 0.223 | 18.655 | 3.723 | 3.061 | 0.002 | 0.007 | |
| 4th | 28,595 | NA | NA | NA | NA | NA | NA | NA | NA | NA | |
| 5th | 153,164 | 0.500 | 1.543 | 0.790 | 2.312 | 165.797 | 14.312 | 25.824 | 0.318 | 0.283 | |
| 6th | 9551 | 0.083 | 0.092 | 0.007 | 0.103 | 5.951 | 1.657 | 0.796 | 0.001 | 0.001 | |
| 7th | 645,354 | 0.566 | 1.893 | 0.946 | 2.516 | 231.081 | 26.454 | 51.181 | 0.190 | 0.423 | |
| 8th | 24,404 | 0.849 | 2.830 | 1.132 | 3.716 | 290.630 | 23.532 | 52.166 | 0.373 | 0.477 | |
| 9th | 6078 | 0.745 | 1.858 | 0.298 | 1.524 | 44.088 | 4.409 | 7.366 | 0.027 | 0.011 | |
| 10th | 838,013 | 0.406 | 0.960 | 0.269 | 0.947 | 33.520 | 3.924 | 6.529 | 0.040 | 0.009 | |
| 11 | 1st | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| 12 | 1st | 19,850 | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| 13 | 1st | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| Case | DNA-ISH | HPV RNA-ISH | E4-IHC | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| E6 | E2 | E4 | E5a | E5b | |||||||||||||||||
| L | M | U | L | M | U | L | M | U | L | M | U | L | M | U | L | M | U | L | M | U | |
| 1 * | − | 1+ | 1+ | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA | NA |
| 2 | 1+ | 1+ | 3+ | − | − | − | − | − | − | − | − | 1+ | − | 2+ | 3+ | − | − | − | − | 1+ | 2+ |
| 3 | 3+ | 3+ | 3+ | − | − | 1+ | − | 1+ | − | − | 1+ | 1+ | − | 1+ | 2+ | − | 1+ | 2+ | − | − | 1+ |
| 4 | 3+ | 3+ | 3+ | − | − | − | − | − | 1+ | − | − | 1+ | − | − | − | − | − | − | − | 1+ | 1+ |
| 5 | 3+ | 3+ | 3+ | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | − | 1+ | 1+ |
| 6 | − | 2+ | 2+ | − | − | − | − | − | 1+ | − | − | 2+ | − | − | 1+ | − | − | − | − | 2+ | 2+ |
| 7 | 2+ | 2+ | 2+ | − | − | − | − | − | − | − | − | 2+ | − | − | − | − | − | − | − | 1+ | 2+ |
| 8 | 1+ | 3+ | 2+ | − | − | − | − | − | − | − | 1+ | 1+ | − | − | 1+ | − | − | − | − | 1+ | 1+ |
| 9 | − | 3+ | 3+ | − | − | 1+ | − | − | 1+ | − | 2+ | 3+ | − | 1+ | 2+ | − | − | 1+ | − | 2+ | 3+ |
| 10 | − | 3+ | 3+ | − | − | 1+ | − | − | − | − | 1+ | 2+ | − | 1+ | 2+ | − | 1+ | 2+ | − | 2+ | 3+ |
| 11 | 1+ | 2+ | 3+ | − | 2+ | 2+ | 2+ | 3+ | 2+ | − | 2+ | 3+ | − | 1+ | 1+ | − | 2+ | 3+ | − | 2+ | 3+ |
| 12 | 1+ | 1+ | 3+ | − | − | − | − | − | 1+ | − | − | 2+ | − | 2+ | 3+ | − | − | 1+ | − | 1+ | 2+ |
| 13 | 2+ | 3+ | 3+ | − | 1+ | 2+ | − | − | − | − | 1+ | 2+ | − | 1+ | 1+ | − | 1+ | 1+ | − | 1+ | 2+ |
| E6 | E7 | E1 | E2 | E4 | E5a | E5b | L2 | L1 | |
|---|---|---|---|---|---|---|---|---|---|
| E6 | - | 0.786 *** | 0.757 *** | 0.754 *** | 0.807 *** | 0.498 * | 0.705 *** | 0.650 *** | 0.577 *** |
| E7 | 0.786 *** | - | 0.794 *** | 0.882 *** | 0.858 *** | 0.649 ** | 0.942 *** | 0.789 *** | 0.838 *** |
| E1 | 0.757 *** | 0.794 *** | - | 0.853 *** | 0.780 *** | 0.845 *** | 0.766 *** | 0.944 *** | 0.745 *** |
| E2 | 0.754 *** | 0.882 *** | 0.853 *** | - | 0.861 *** | 0.721 *** | 0.872 *** | 0.819 *** | 0.743 *** |
| E4 | 0.807 *** | 0.858 *** | 0.780 *** | 0.861 *** | - | 0.554 *** | 0.828 *** | 0.705 *** | 0.710 *** |
| E5a | 0.498 * | 0.649 ** | 0.845 *** | 0.721 *** | 0.554 *** | - | 0.6965 *** | 0.904 *** | 0.814 *** |
| E5b | 0.705 *** | 0.942 *** | 0.766 *** | 0.872 *** | 0.828 *** | 0.6965 *** | - | 0.781 *** | 0.922 *** |
| L2 | 0.650 *** | 0.789 *** | 0.944 *** | 0.819 *** | 0.705 *** | 0.904 *** | 0.781 *** | - | 0.806 *** |
| L1 | 0.577 *** | 0.838 *** | 0.745 *** | 0.743 *** | 0.710 *** | 0.814 *** | 0.922 *** | 0.806 *** | - |
| mRNA | r | p-Value |
|---|---|---|
| E6 | 0.237 | 0.3289 |
| E7 | 0.277 | 0.2506 |
| E1 | 0.449 | 0.0536 |
| E2 | 0.395 | 0.0944 |
| E4 | 0.323 | 0.1777 |
| E5a | 0.507 | 0.0267 * |
| E5b | 0.374 | 0.1150 |
| L2 | 0.542 | 0.0165 * |
| L1 | 0.436 | 0.0619 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ikegami, T.; Hirakawa, H.; Tsukahara, N.; Murakami, A.; Kise, N.; Kiyuna, A.; Kosugi, T.; Agena, S.; Kinjyo, H.; Hasegawa, N.; et al. Coordinated Expression of HPV-6 Genes with Predominant E4 and E5 Expression in Laryngeal Papilloma. Microorganisms 2021, 9, 520. https://doi.org/10.3390/microorganisms9030520
Ikegami T, Hirakawa H, Tsukahara N, Murakami A, Kise N, Kiyuna A, Kosugi T, Agena S, Kinjyo H, Hasegawa N, et al. Coordinated Expression of HPV-6 Genes with Predominant E4 and E5 Expression in Laryngeal Papilloma. Microorganisms. 2021; 9(3):520. https://doi.org/10.3390/microorganisms9030520
Chicago/Turabian StyleIkegami, Taro, Hitoshi Hirakawa, Narutoshi Tsukahara, Akikazu Murakami, Norimoto Kise, Asanori Kiyuna, Takayoshi Kosugi, Shinya Agena, Hidetoshi Kinjyo, Narumi Hasegawa, and et al. 2021. "Coordinated Expression of HPV-6 Genes with Predominant E4 and E5 Expression in Laryngeal Papilloma" Microorganisms 9, no. 3: 520. https://doi.org/10.3390/microorganisms9030520