Hedgehogs, Squirrels, and Blackbirds as Sentinel Hosts for Active Surveillance of Borrelia miyamotoi and Borrelia burgdorferi Complex in Urban and Rural Environments

 , , , , , , , and
, , , , , , , and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Hosts, Dissections, Sampling
2.2. DNA Isolation
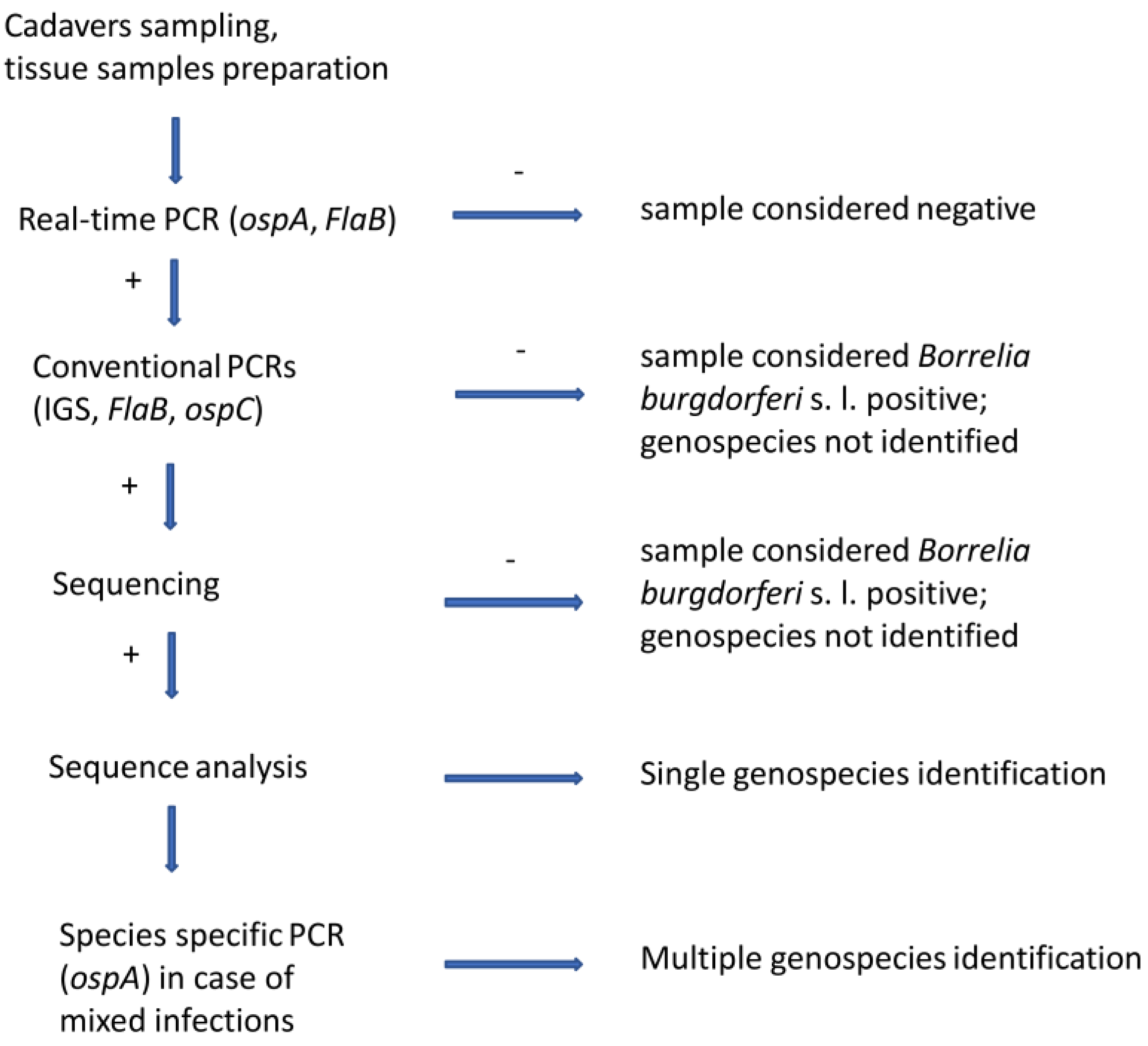
2.3. Screening of the Samples through Multiplex Real-Time PCR
2.4. Conventional PCRs and Sequencing
2.5. Species-Specific PCR
2.6. Statistical Analyses
3. Results
3.1. Prevalence of B. miyamotoi
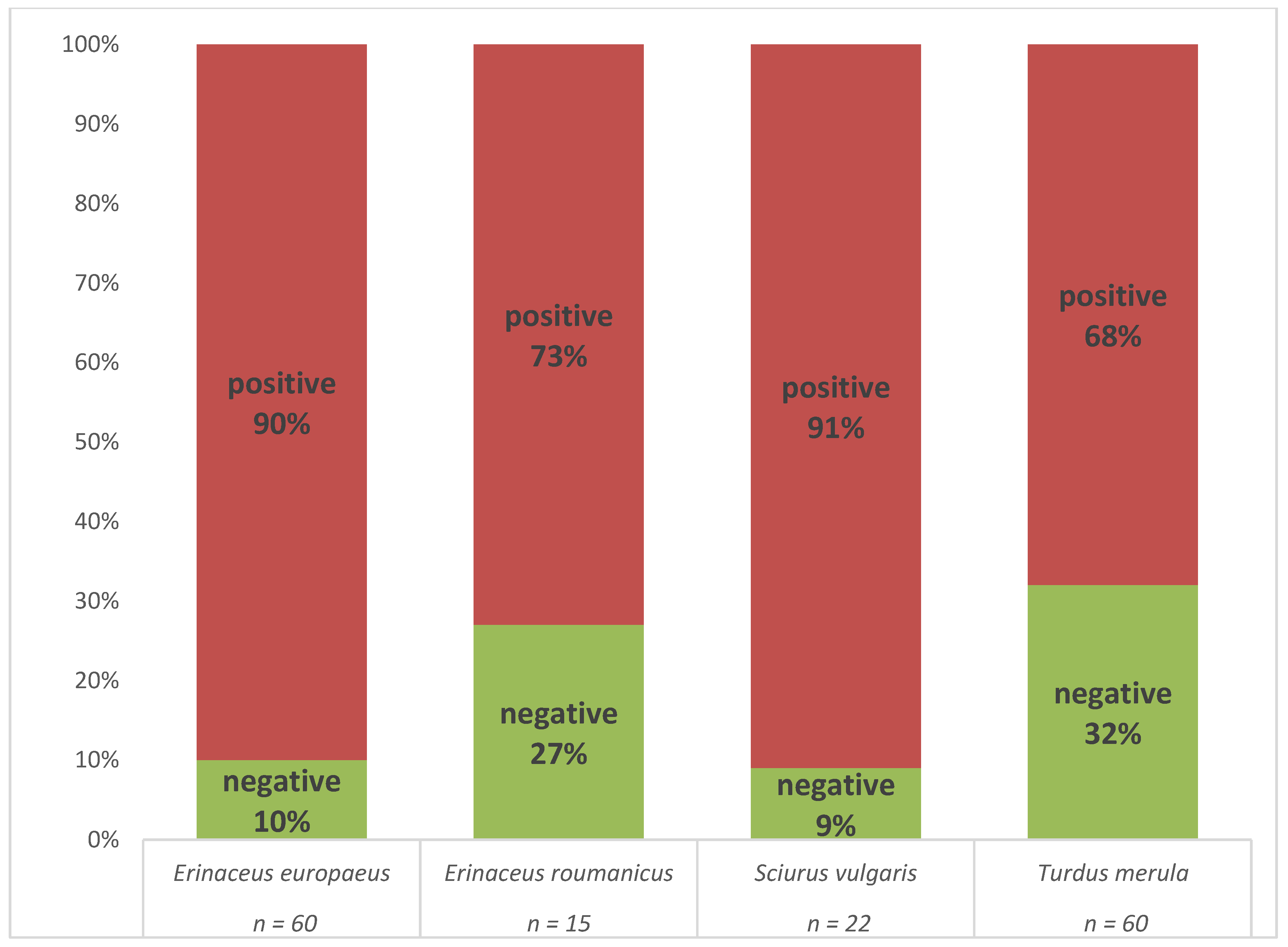
3.2. Prevalence of B. burgdorferi s.l.
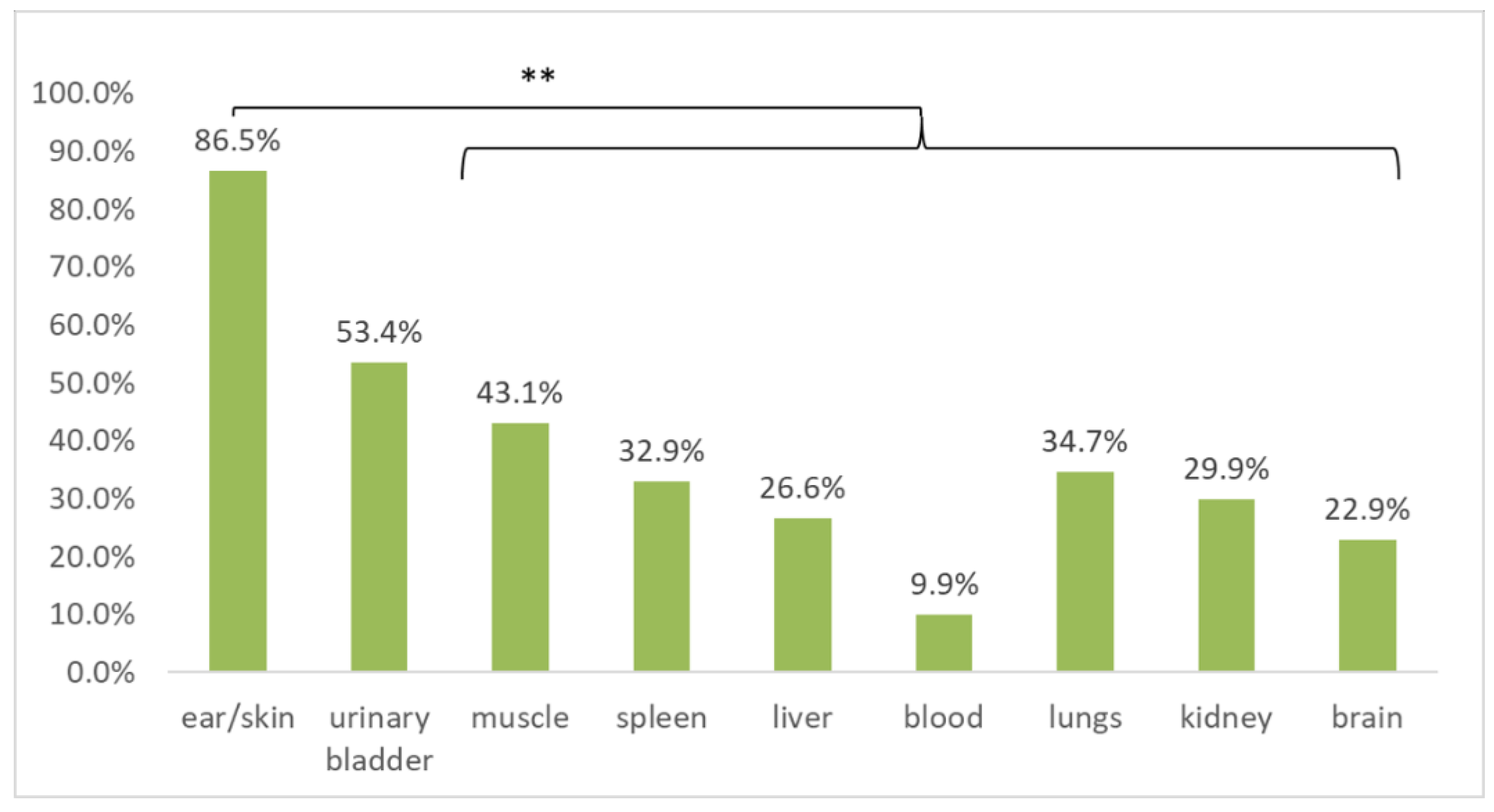
3.3. Occurrence of Borrelia burgdorferi s.l. in Specific Tissues
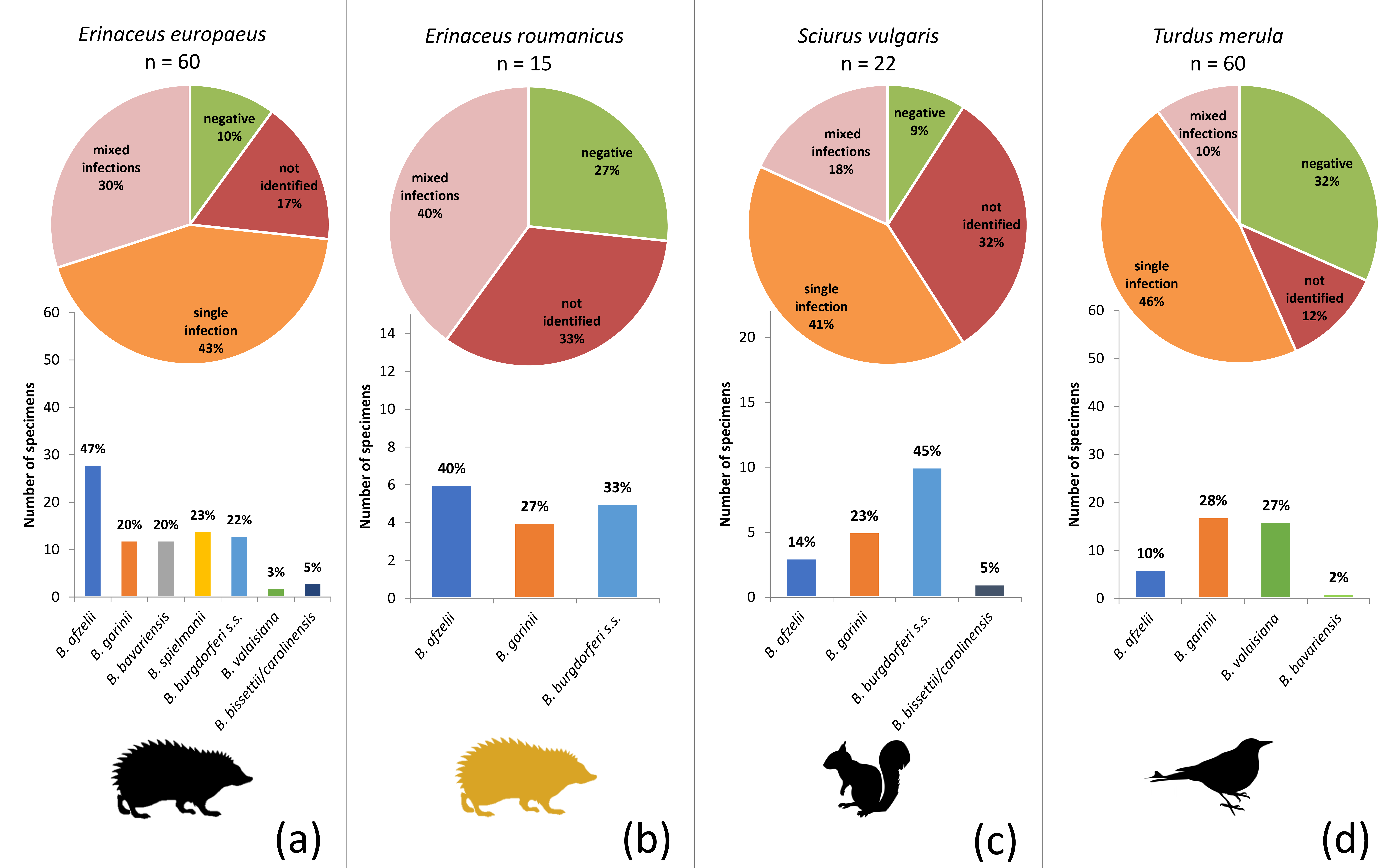
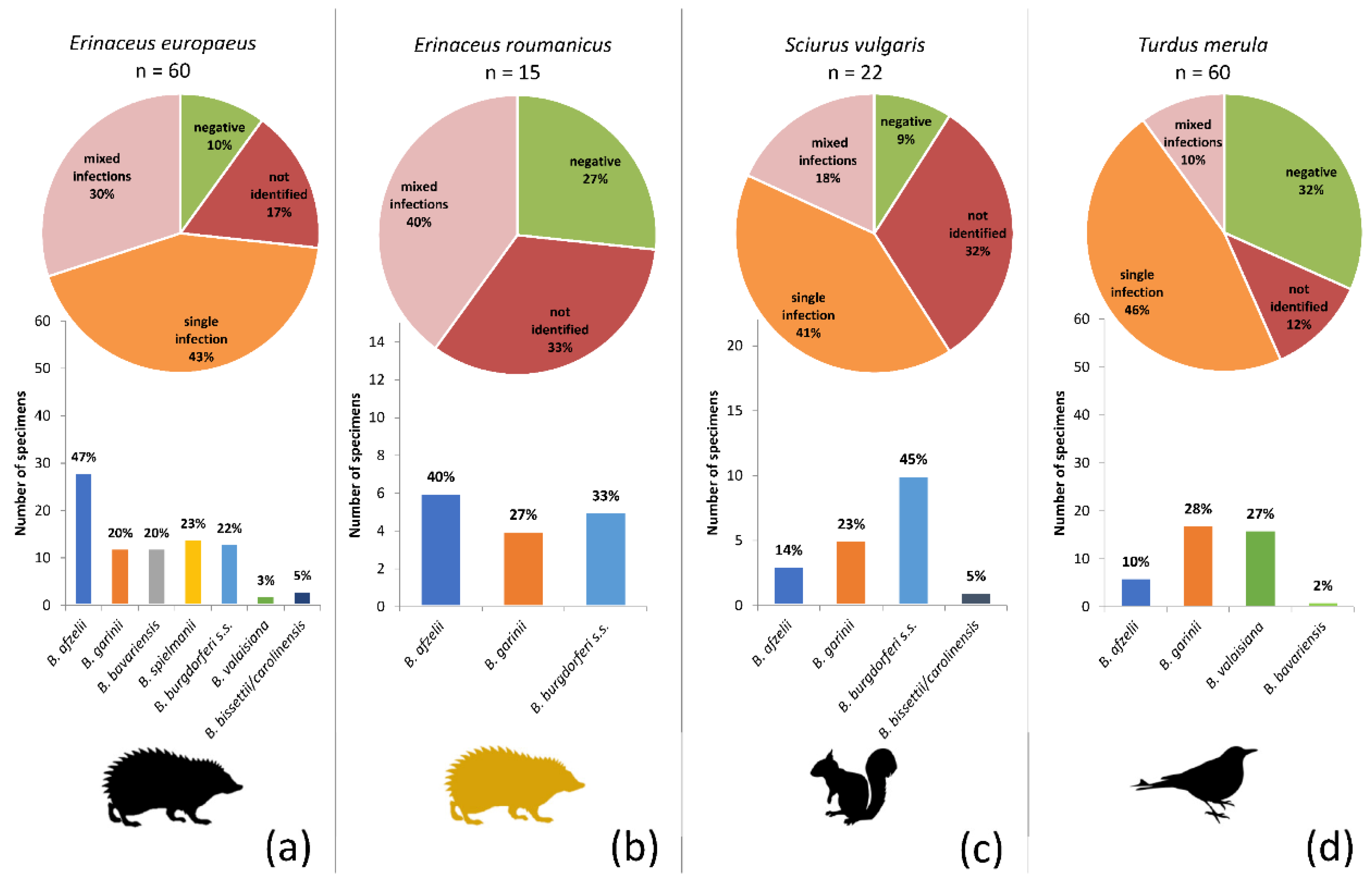
3.4. Identification of Borrelia burgdorferi s.l. Genospecies
3.5. Comparison of Occurrence of Borrelia burgdorferi s.l. Genospecies in Investigated Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lindgren, E.; Jaenson, T. Lyme Borreliosis in Europe: Influences of Climate and Climate Change, Epidemiology, Ecology and Adaptation Measures; WHO Regional Office for Europe: Copenhagen, Denmark, 2006. [Google Scholar]
- Hubálek, Z. Epidemiology of lyme borreliosis. Curr. Probl. Dermatol. 2009, 37, 31–50. [Google Scholar] [PubMed]
- Sykes, R.A.; Makiello, P. An estimate of Lyme borreliosis incidence in Western Europe. J. Public Health 2016, 39, 74–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Petrulioniené, A.; Radzišauskienè, D.; Ambrozaitis, A.; Čaplinskas, S.; Paulauskas, A.; Venalis, A. Epidemiology of Lyme Disease in a highly endemic European zone. Medicina 2020, 56, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stanek, G.; Strle, F. Lyme borreliosis–from tick bite to diagnosis and treatment. FEMS Microbiol. Rev. 2018, 42, 233–258. [Google Scholar] [CrossRef] [Green Version]
- Canica, M.M.; Nato, F.; du Merle, L.; Mazie, J.C.; Baranton, G.; Postic, D. Monoclonal antibodies for identification of Borrelia afzelii sp. nov. associated with late cutaneous manifestations of Lyme borreliosis. Scand. J. Infect. Dis. 1993, 25, 441–448. [Google Scholar] [CrossRef]
- Margos, G.; Vollmer, S.A.; Cornet, M.; Garnier, M.; Fingerle, V.; Wilske, B.; Bormane, A.; Vitorino, L.; Collares-Pereira, M.; Drancourt, M.; et al. A new Borrelia species defined by multilocus sequence analysis of housekeeping genes. Appl. Environ. Microbiol. 2009, 75, 5410–5416. [Google Scholar] [CrossRef] [Green Version]
- Postic, D.; Ras, N.M.; Lane, R.S.; Hendson, M.; Baranton, G. Expanded diversity among Californian Borrelia isolates and description of Borrelia bissettii sp. nov. (formerly Borrelia group DN127). J. Clin. Microbiol. 1998, 36, 3497–3504. [Google Scholar] [CrossRef] [Green Version]
- Baranton, G.; Postic, D.; Saint Girons, I.; Boerlin, P.; Piffaretti, J.C.; Assous, M.; Grimont, P.A. Delineation of Borrelia burgdorferi sensu stricto, Borrelia garinii sp. nov., and group VS461 associated with Lyme borreliosis. Int. J. Syst. Bacteriol. 1992, 42, 378–383. [Google Scholar] [CrossRef] [Green Version]
- Margos, G.; Hojgaard, A.; Lane, R.S.; Cornet, M.; Fingerle, V.; Rudenko, N.; Ogden, N.; Aanensen, D.M.; Fish, D.; Piesman, J. Multilocus sequence analysis of Borrelia bissettii strains from North America reveals a new Borrelia species, Borrelia kurtenbachii. Ticks Tick Borne Dis. 2010, 1, 151–158. [Google Scholar] [CrossRef] [Green Version]
- Le Fleche, A.; Postic, D.; Girardet, K.; Peter, O.; Baranton, G. Characterization of Borrelia lusitaniae sp. nov. by 16S ribosomal DNA sequence analysis. Int. J. Syst. Bacteriol. 1997, 47, 921–925. [Google Scholar] [CrossRef] [Green Version]
- Pritt, B.S.; Respicio-Kingry, L.B.; Sloan, L.M.; Schriefer, M.E.; Replogle, A.J.; Bjork, J.; Liu, G.; Kingry, L.C.; Mead, P.S.; Neitzel, D.F.; et al. Borrelia mayonii sp. nov., a member of the Borrelia burgdorferi sensu lato complex, detected in patients and ticks in the upper midwestern United States. Int. J. Syst. Evol. Microbiol. 2016, 66, 4878–4880. [Google Scholar] [CrossRef]
- Richter, D.; Postic, D.; Sertour, N.; Livey, I.; Matuschka, F.R.; Baranton, G. Delineation of Borrelia burgdorferi sensu lato species by multilocus sequence analysis and confirmation of the delineation of Borrelia spielmanii sp. nov. Int. J. Syst. Evol. Microbiol. 2006, 56, 873–881. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; van Dam, A.P.; Le Fleche, A.; Postic, D.; Peter, O.; Baranton, G.; de Boer, R.; Spanjaard, L.; Dankert, J. Genetic and phenotypic analysis of Borrelia valaisiana sp. nov. (Borrelia genomic groups VS116 and M19). Int. J. Syst. Bacteriol. 1997, 47, 926–932. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, N.; Golovchenko, M.; Lin, T.; Gao, L.; Grubhoffer, L.; Oliver, J.H., Jr. Delineation of a new species of the Borrelia burgdorferi Sensu Lato Complex, Borrelia americana sp. nov. J. Clin. Microbiol. 2009, 47, 3875–3880. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marconi, R.T.; Liveris, D.; Schwartz, I. Identification of novel insertion elements, restriction fragment length polymorphism patterns, and discontinuous 23S rRNA in Lyme disease spirochetes: Phylogenetic analyses of rRNA genes and their intergenic spacers in Borrelia japonica sp. nov. and genomic group 21038 (Borrelia andersonii sp. nov.) isolates. J. Clin. Microbiol. 1995, 33, 2427–2434. [Google Scholar] [PubMed]
- Postic, D.; Garnier, M.; Baranton, G. Multilocus sequence analysis of atypical Borrelia burgdorferi sensu lato isolates--description of Borrelia californiensis sp. nov., and genomospecies 1 and 2. Int. J. Med. Microbiol. 2007, 297, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H., Jr. Borrelia carolinensis sp. nov., a new (14th) member of the Borrelia burgdorferi Sensu Lato complex from the southeastern region of the United States. J Clin. Microbiol. 2009, 47, 134–141. [Google Scholar] [CrossRef] [Green Version]
- Ivanova, L.B.; Tomova, A.; González-Acuña, D.; Murúa, R.; Moreno, C.X.; Hernández, C.; Cabello, J.; Cabello, C.; Daniels, T.J.; Godfrey, H.P.; et al. Borrelia chilensis, a new member of the Borrelia burgdorferi sensu lato complex that extends the range of this genospecies in the Southern Hemisphere. Environ Microbiol. 2014, 16, 1069–1080. [Google Scholar] [CrossRef] [Green Version]
- Casjens, S.R.; Fraser-Liggett, C.M.; Mongodin, E.F.; Qiu, W.G.; Dunn, J.J.; Luft, B.J.; Schutzer, S.E. Whole genome sequence of an unusual Borrelia burgdorferi sensu lato isolate. J/ Bacteriol. 2011, 193, 1489–1490. [Google Scholar] [CrossRef] [Green Version]
- Kawabata, H.; Masuzawa, T.; Yanagihara, Y. Genomic analysis of Borrelia japonica sp. nov. isolated from Ixodes ovatus in Japan. Microbiol/ Immunol. 1993, 37, 843–848. [Google Scholar] [CrossRef]
- Margos, G.; Fedorova, N.; Kleinjan, J.E.; Hartberger, C.; Schwan, T.G.; Sing, A.; Fingerle, V. Borrelia lanei sp. nov. extends the diversity of Borrelia species in California. Int. J. Syst. Evol. Microbiol. 2017, 67, 3872–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukunaga, M.; Hamase, A.; Okada, K.; Nakao, M. Borrelia tanukii sp. nov. and Borrelia turdae sp. nov. found from ixodid ticks in Japan: Rapid species identification by 16S rRNA gene-targeted PCR analysis. Microbiol Immunol. 1996, 40, 877–881. [Google Scholar] [CrossRef] [PubMed]
- Masuzawa, T.; Takada, N.; Kudeken, M.; Fukui, T.; Yano, Y.; Ishiguro, F.; Kawamura, Y.; Imai, Y.; Ezaki, T. Borrelia sinica sp. nov., a lyme disease-related Borrelia species isolated in China. Int. J. Syst. Evol. Microbiol. 2001, 51, 1817–1824. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, C.Y.; Liu, W.; Jiang, B.G.; Wang, D.M.; Jiang, W.J.; Zhao, Q.M.; Zhang, P.H.; Wang, Z.X.; Tang, G.P.; Yang, H.; et al. Novel genospecies of Borrelia burgdorferi sensu lato from rodents and ticks in southwestern China. J. Clin. Microbiol. 2008, 46, 3130–3133. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, N.; Golovchenko, M.; Mokráček, A.; Piskunová, N.; Růžek, D.; Mallatová, N.; Grubhoffer, L. Detection of Borrelia bissettii in cardiac valve tissue of a patient with endocarditis and aortic valve stenosis in the Czech Republic. J. Clin. Microbiol. 2008, 46, 3540–3543. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, N.; Golovchenko, M.; Grubhoffer, L.; Oliver, J.H., Jr. Updates on Borrelia burgdorferi sensu lato complex with respect to public health. Ticks Tick Borne Dis. 2011, 2, 123–128. [Google Scholar] [CrossRef] [Green Version]
- Rudenko, N.; Golovchenko, M.; Vancová, M.; Clark, K.; Grubhoffer, L.; Oliver, J.H., Jr. Isolation of live Borrelia burgdorferi sensu lato spirochaetes from patients with undefined disorders and symptoms not typical for Lyme borreliosis. Clin. Microbiol. Infect. 2016, 22, 267.e9–267.e15. [Google Scholar] [CrossRef] [Green Version]
- Stanek, G.; Reiter, M. The expanding Lyme Borrelia complex--clinical significance of genomic species? Clin. Microbiol. Infect. 2011, 17, 487–493. [Google Scholar] [CrossRef] [Green Version]
- Gern, L. Borrelia burgdorferi sensu lato, the agent of lyme borreliosis: Life in the wilds. Parasite 2008, 15, 244–247. [Google Scholar] [CrossRef] [Green Version]
- Estrada- Peña, A.; Jongejan, F. Ticks feeding on humans: A review of records on human-biting Ixodoidea with special reference to pathogen transmission. Exp. Appl. Acarol. 1999, 23, 685–715. [Google Scholar] [CrossRef]
- Gern, L.; Rouvinez, E.; Toutoungi, L.N.; and Godfroid, E. Transmission cycles of Borrelia burgdorferi sensu lato involving Ixodes ricinus and/or I. hexagonus ticks and the European hedgehog, Erinaceus europaeus, in suburban and urban areas in Switzerland. Folia Parasitol 1997, 44, 309–314. [Google Scholar]
- Kybicová, K.; Baštová, K.; Malý, M. Detection of Borrelia burgdorferi sensu lato and Anaplasma phagocytophilum in questing ticks Ixodes ricinus from the Czech Republic. Ticks Tick Borne Dis. 2017, 8, 483–487. [Google Scholar] [CrossRef]
- Rizzoli, A.; Silaghi, C.; Obiegala, A.; Rudolf, I.; Hubálek, Z.; Földvári, G.; Plantard, O.; Vayssier-Taussat, M.; Bonnet, S.; Špitalská, E.; et al. Ixodes ricinus and its transmitted pathogens in urban and peri-urban areas in Europe: New hazards and relevance for public health. Front. Public Health 2014, 2, 251. [Google Scholar] [CrossRef] [PubMed]
- Gassner, F.; Hansford, K.M.; Medlock, J.M. Greener cities, a wild card for ticks? In Ecology and Prevention of Lyme Borreliosis. Ecology and Control of Vector-Borne Diseases Volume 4; Braks, M.A.H., van Wieren, S.E., Takken, W., Sprong, H., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; pp. 187–203. [Google Scholar]
- Uspensky, I. Tick pests and vectors (Acari: Ixodoidea) in European towns: Introduction, persistence and management. Ticks Tick Borne Dis 2014, 5, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Földvári, G.; Rigó, K.; Jablonszky, M.; Biró, N.; Majoros, G.; Molnár, V.; Tóth, M. Ticks and the city: Ectoparasites of the Northern white-breasted hedgehog (Erinaceus roumanicus) in an urban park. Ticks Tick Borne Dis. 2011, 2, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Heylen, D.; Lasters, R.; Adriaensen, F.; Fonville, M.; Sprong, H.; Matthysen, E. Ticks and tick-borne diseases in the city: Role of landscape connectivity and green space characteristics in a metropolitan area. Sci. Total Environ. 2019, 670, 941–949. [Google Scholar] [CrossRef]
- Hornok, S.; Meli, M.L.; Gönczi, E.; Halász, E.; Takács, N.; Farkas, R.; Hofmann-Lehmann, R. Occurrence of ticks and prevalence of Anaplasma phagocytophilum and Borrelia burgdorferi s.l. in three types of urban biotopes: Forests, parks and cemeteries. Ticks Tick Borne Dis. 2014, 5, 785–789. [Google Scholar] [CrossRef]
- Kubiak, K.; Sielawa, H.; Dziekońska-Rynko, J.; Kubiak, D.; Rydzewska, M.; Dzika, E. Dermacentor reticulatus ticks (Acari: Ixodidae) distribution in north-eastern Poland: An endemic area of tick-borne diseases. Exp. Appl. Acarol. 2018, 75, 289–298. [Google Scholar] [CrossRef] [Green Version]
- Kubiak, K.; Dziekońska-Rynko, J.; Szymańska, H.; Kubiak, D.; Dmitryjuk, M.; Dzika, E. Questing Ixodes ricinus ticks (Acari, Ixodidae) as a vector of Borrelia burgdorferi sensu lato and Borrelia miyamotoi in an urban area of north-eastern Poland. Exp. Appl. Acarol. 2019, 78, 113–126. [Google Scholar] [CrossRef] [Green Version]
- Maetzel, D.; Maier, W.A.; Kampen, H. Borrelia burgdorferi infection prevalences in questing Ixodes ricinus ticks (Acari: Ixodidae) in urban and suburban Bonn, western Germany. Parasitol. Res. 2005, 95, 5–12. [Google Scholar] [CrossRef]
- Mancini, F.; Di Luca, M.; Toma, L.; Vescio, F.; Bianchi, R.; Khoury, C.; Marini, L.; Rezza, G.; and Ciervo, A. Prevalence of tick-borne pathogens in an urban park in Rome, Italy. Ann. Agric. Environ. Med. 2014, 21, 723–727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szekeres, S.; van Leeuwen, A.D.; Tóth, E.; Majoros, G.; Sprong, H.; Földvári, G. Road-killed mammals provide insight into tick-borne bacterial pathogen communities within urban habitats. Transbound. Emerg. Dis. 2019, 66, 277–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venclíková, K.; Betášová, L.; Sikutová, S.; Jedličková, P.; Hubálek, Z.; Rudolf, I. Human pathogenic borreliae in Ixodes ricinus ticks in natural and urban ecosystem (Czech Republic). Acta Parasitol. 2014, 59, 717–720. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, K.L.; Gaston, K.J.; Frantz, A.C.; Dimon, M.T.; Sharp, S.P.; McGowan, A.; Dawson, D.A.; Walasz, K.; Partecke, J.; Burke, T.; et al. Independent colonization of multiple urban centres by a formerly forest specialist bird species. Proc. Biol. Sci. 2009, 276, 2403–2410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gray, J.S.; Kahl, O.; Janetzki-Mittman, C.; Stein, J.; Guy, E. Acquisition of Borrelia burgdorferi by Ixodes ricinus ticks fed on the European hedgehog, Erinaceus europaeus L. Exp. Appl. Acarol. 1994, 18, 485–491. [Google Scholar] [CrossRef]
- Hubert, P.; Julliard, R.; Biagianti, S.; Poulle, M.L. Ecological factors driving the higher hedgehog (Erinaceus europeaus) density in an urban area compared to the adjacent rural area. Landsc. Urban Plan 2011, 103, 34–43. [Google Scholar] [CrossRef]
- Humair, P.F.; Gern, L.E. Relationship between Borrelia burgdorferi sensu lato species, Eurasian red squirrels (Sciurus vulgaris) and Ixodes ricinus in enzootic areas in Switzerland. Acta Trop. 1998, 69, 213–227. [Google Scholar] [CrossRef] [Green Version]
- Humair, P.F.; Postic, D.; Wallich, R.; Gern, L. An avian reservoir (Turdus merula) of the Lyme borreliosis spirochetes. Zentralbl. Bakteriol. 1998, 287, 521–538. [Google Scholar] [CrossRef]
- Skuballa, J.; Oehme, R.; Hartelt, K.; Petney, T.; Bücher, T.; Kimmig, P.; Taraschewski, H. European hedgehogs as hosts for Borrelia spp., Germany. Emerg. Infect Dis. 2007, 13, 952–953. [Google Scholar] [CrossRef] [PubMed]
- Skuballa, J.; Petney, T.; Pfäffle, M.; Oehme, R.; Hartelt, K.; Fingerle, V.; Kimmig, P.; Taraschewski, H. Occurrence of different Borrelia burgdorferi sensu lato genospecies including B. afzelii, B. bavariensis, and B. spielmanii in hedgehogs (Erinaceus spp.) in Europe. Ticks Tick Borne Dis. 2012, 3, 8–13. [Google Scholar] [CrossRef]
- Taragel’ová, V.; Koči, J.; Hanincová, K.; Kurtenbach, K.; Derdáková, M.; Ogden, N.H.; Literák, I.; Kocianová, E.; Labuda, M. Blackbirds and song thrushes constitute a key reservoir of Borrelia garinii, the causative agent of borreliosis in Central Europe. Appl. Environ. Microbiol. 2008, 74, 1289–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hudec, K.; Šťastný, K. Fauna ČR Ptáci III/I (pěvci), 2nd ed.; Academia: Prague, Czech Republic, 2011; pp. 278–289. [Google Scholar]
- Wilson, D.E.; Lacher, T.E.; Mittermeier, R.A. Handbook of the Mammals of the World. Volume 6: Lagomorphs and Rodents I; Lynx Edicions and IUCN: Barcelona, Spain, 2016; p. 987. [Google Scholar]
- Wilson, D.E.; Mittermeier, R.A. Handbook of the Mammals of the World. Volume 8: Insectivores, Sloths and Colugos; Lynx Edicions and IUCN: Barcelona, Spain, 2018; p. 710. [Google Scholar]
- Bolfíková, B.; Hulva, P. Microevolution of sympatry: Landscape genetics of hedgehogs Erinaceus europaeus and E. roumanicus in Central Europe. Heredity (Edinb) 2012, 108, 248–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Černá Bolfíková, B.; Eliášová, K.; Loudová, M.; Kryštufek, B.; Lymberakis, P.; Sándor, A.D.; Hulva, P. Glacial allopatry vs. postglacial parapatry and peripatry: The case of hedgehogs. PeerJ 2017, 5, e3163. [Google Scholar] [CrossRef] [PubMed]
- Hönig, V.; Palus, M.; Kašpar, T.; Zemanová, M.; Majerová, K.; Hofmannová, L.; Papežík, P.; Šikutová, S.; Rettich, F.; Hubálek, Z.; et al. Multiple Lineages of Usutu Virus (Flaviviridae, Flavivirus) in Blackbirds (Turdus merula) and Mosquitoes (Culex pipiens, Cx. modestus) in the Czech Republic (2016–2019). Microorganisms 2019, 7, 568. [Google Scholar]
- Kasahara, T.; Miyazaki, T.; Nitta, H.; Ono, A.; Miyagishima, T.; Nagao, T.; Urushidani, T. Evaluation of methods for duration of preservation of RNA quality in rat liver used for transcriptome analysis. J. Toxicol. Sci. 2006, 31, 509–519. [Google Scholar] [CrossRef] [Green Version]
- Heylen, D.J.A.; Tijsse, E.; Fonville, M.; Matthysen, E.; Sprong, H. Transmission dynamics of Borrelia burgdorferi s.l. in a bird tick community. Environ. Microbiol. 2013, 15, 663–673. [Google Scholar] [CrossRef]
- Demaerschalck, I.; Ben Messaoud, A.; De Kesel, M.; Hoyois, B.; Lobet, Y.; Hoet, P.; Bigaignon, G.; Bollen, A.; Godfroid, E. Simultaneous presence of different Borrelia burgdorferi genospecies in biological fluids of Lyme disease patients. J. Clin. Microbiol. 1995, 33, 602–608. [Google Scholar] [CrossRef] [Green Version]
- Agresti, A. Categorical Data Analysis, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Hothorn, T.; Hornik, K.; van de Wiel, M.A.; Zeileis, A. Implementing a class of permutation tests: The coin package. J. Stat. Softw. 2008, 28, 1–23. [Google Scholar] [CrossRef]
- Cutler, S.; Vayssier-Taussat, M.; Estrada-Peña, A.; Potkonjak, A.; Mihalca, A.D.; Zeller, H. A new Borrelia on the block: Borrelia miyamotoi—A human health risk? Euro Surveill 2019, 24, 1800170. [Google Scholar] [CrossRef] [Green Version]
- Crowder, C.D.; Carolan, H.E.; Rounds, M.A.; Hönig, V.; Mothes, B.; Haag, H.; Nolte, O.; Luft, B.J.; Grubhoffer, L.; Ecker, D.J.; et al. Prevalence of Borrelia miyamotoi in Ixodes Ticks in Europe and the United States. Emerg. Infect. Dis. 2014, 20, 1678–1682. [Google Scholar] [CrossRef]
- Page, S.; Daschkin, C.; Anniko, S.; Krey, V.; Nicolaus, C.; Maxeiner, H.G. First report of Borrelia miyamotoi in an Ixodes ricinus tick in Augsburg, Germany. Exp. Appl. Acarol. 2018, 74, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Răileanu, C.; Tauchmann, O.; Vasić, A.; Wöhnke, E.; Silaghi, C. Borrelia miyamotoi and Borrelia burgdorferi (sensu lato) identification and survey of tick-borne encephalitis virus in ticks from north-eastern Germany. Parasit. Vectors 2020, 13, 106. [Google Scholar] [CrossRef] [PubMed]
- Morán Cadenas, F.; Rais, O.; Humair, P.F.; Douet, V.; Moret, J.; Gern, L. Identification of host bloodmeal source and Borrelia burgdorferi sensu lato in field-collected Ixodes ricinus ticks in Chaumont (Switzerland). J. Med. Entomol. 2007, 44, 1109–1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pisanu, B.; Chapuis, J.L.; Dozières, A.; Basset, F.; Poux, V.; Vourc´h, G. High prevalence of Borrelia burgdorferi s. l. in the European res squirrel in France. Ticks Tick Borne Dis. 2014, 5, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Ruyts, S.C.; Frazer-Mendelewska, E.; Van Den Berge, K.; Verheyen, K.; Sprong, H. Molecular detection of tick-borne pathogens Borrelia afzelii, Borrelia miyamotoi and Anaplasma phagocytophilum in Eurasian red squirrels (Sciurus vulgaris). Eur. J. Wildl. Res. 2017, 63, 43. [Google Scholar] [CrossRef]
- Mysterud, A.; Stigum, V.M.; Jaarsma, R.I.; Sprong, H. Genospecies of Borrelia burgdorferi s. l. detected in 16 mammal species and questing ticks from northern Europe. Sci. Rep. 2019, 9, 5088. [Google Scholar] [CrossRef]
- Krawczyk, A.I.; van Leeuwen, A.D.; Jacobs-Reitsma, W.; Wijnands, L.M.; Bouw, E.; Jahfari, S.; van Hoek, A.H.; van der Giessen, J.W.; Roelfsema, J.H.; Kroes, M.; et al. Presence of zoonotic agents in engorged ticks and hedgehog faeces from Erinaceus europaeus in (sub) urban areas. Parasit. Vectors 2015, 8, 210. [Google Scholar] [CrossRef] [Green Version]
- Szekeres, S.; Majláthová, V.; Majláth, I.; Földvári, G. Neglected hosts: The role of lacertid lizards and medium-sized mammals in the eco-epidemiology of Lyme borreliosis. In Ecology and Prevention of Lyme Borreliosis. Ecology and Control of Vector-Borne Diseases Volume 4; Braks, M.A.H., van Wieren, S.E., Takken, W., Sprong, H., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; pp. 103–126. [Google Scholar]
- Jahfari, S.; Ruyts, S.C.; Frazer-Mendelewska, E.; Jaarsma, R.; Verheyen, K.; Sprong, H. Melting pot of tick-borne zoonoses: The European hedgehog contributes to the maintenance of various tick-borne diseases in natural cycles urban and suburban areas. Parasit. Vectors 2017, 10, 134. [Google Scholar] [CrossRef] [Green Version]
- Dumitrache, M.O.; Paştiu, A.I.; Kalmár, Z.; Mircean, V.; Sándor, A.D.; Gherman, C.M.; Peştean, C.; Mihalca, A.D.; Cozma, V. Northern white-breasted hedgehogs Erinaceus roumanicus as hosts for ticks infected with Borrelia burgdorferi sensu lato and Anaplasma phagocytophilum in Romania. Ticks Tick Borne Dis. 2013, 4, 214–217. [Google Scholar] [CrossRef]
- Dziemian, S.; Sikora, B.; Pilacinska, P.; Michalik, J.; Zwolak, R. Ectoparasite loads in sympatric urban populations of the Northern white-breasted and the European hedgehog. Parasitol. Res. 2015, 114, 2317–2323. [Google Scholar] [CrossRef] [Green Version]
- Heylen, D.J.A. Ecological interactions between songbirds, ticks, and Borrelia burfdorferi s.l. In Europe. In Ecology and Prevention of Lyme Borreliosis. Ecology and Control of Vector-Borne Diseases Volume 4; Braks, M.A.H., van Wieren, S.E., Takken, W., Sprong, H., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2016; pp. 91–101. [Google Scholar]
- Mtierová, Z.; Derdáková, M.; Chvostáč, M.; Didyk, Y.M.; Mangová, B.; Rusňáková Tarageľová, V.; Selyemová, D.; Šujanová, A.; Václav, R. Local population structure and seasonal variability of Borrelia garinii genotypes in Ixodes ricinus ticks, Slovakia. Int. J. Environ. Res. Public Health 2020, 17, 3607. [Google Scholar] [CrossRef] [PubMed]
- Norte, A.C.; Margos, G.; Becker, N.S.; Ramos, J.A.; Núncio, M.S.; Fingerle, V.; Araújo, P.M.; Adamík, P.; Alivizatos, H.; Barba, E.; et al. Host dispersal shapes the population structure of a tick-borne bacterial pathogen. Mol. Ecol. 2020, 29, 485–501. [Google Scholar] [CrossRef] [PubMed]
- Margos, G.; Fingerle, V.; Reynolds, S. Borrelia bavariensis: Vector switch, niche invasion, and geographical spread of a tick-borne bacterial parasite. Front. Ecol. Evol. 2019, 7, 401. [Google Scholar] [CrossRef] [Green Version]
- Heylen, D.J.A.; Sprong, H.; Krawczyk, A.; Van Houtte, N.; Genné, D.; Gomez-Chamorro, A.; van Oers, K.; Voordouw, M.J. Inefficient co-feeding transmission of Borrelia afzelii in two common European songbirds. Sci. Rep. 2017, 7, 39596. [Google Scholar] [CrossRef] [Green Version]
- Franke, J.; Moldenhauer, A.; Hildebrandt, A.; Dorn, W. Are birds reservoir hosts for Borrelia afzelii? Ticks Tick Borne Dis. 2010, 1, 109–112. [Google Scholar] [CrossRef]
- Tichá, L.; Golovchenko, M.; Oliver, J.H.; Grubhoffer, L.; Rudenko, N. Sensitivity of Lyme borreliosis spirochetes to serum complement of regular Zoo animals: Potential reservoir competence of some exotic vertebrates. Vector Borne Zoonotic Dis 2016, 16, 13–19. [Google Scholar] [CrossRef]
- Norte, A.C.; de Carvalho, I.L.; Núncio, M.S.; Araújo, P.M.; Matthysen, E.; Ramos, J.A.; Sprong, H.; Heylen, D. Getting under the bird´s skin-tissue tropism of Borrelia burgdorferi s.l. in naturally and experimentally infected avian hosts. Microb. Ecol. 2020, 79, 756–769. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Host Species (Number of Individuals) | Ear/Skin * | Muscle | Blood | Lungs | Liver | Spleen | Urinary Bladder | Kidney | Brain |
|---|---|---|---|---|---|---|---|---|---|
| Erinaceus europaeus (n = 54) | 87.0% | 63.5% | 17.8% | 38.3% | 44.4% | 40.0% | 60.9% | 40.0% | 33.3% |
| E. roumanicus (n = 11) | 72.7% | 54.5% | 12.5% | 22.2% | 22.2% | 20.0% | 45.5% | 22.2% | 37.5% |
| Sciurus vulgaris (n = 20) | 90.0% | 35.0% | 5.9% | 31.6% | 15.8% | 22.2% | 37.5% | 5.6% | 7.1% |
| Turdus merula (n = 41) | 87.8% | 17.5% | 0.0% | x | 11.1% | x | x | x | 13.2% |
| Ear/Skin | Muscle | Blood | Lungs | Liver | Spleen | Urinary Bladder | Kidney | Brain | |
|---|---|---|---|---|---|---|---|---|---|
| B. afzelii *** | 27.84% | 23.71% | 2.06% | 4.12% | 6.19% | 7.22% | 15.46% | 7.22% | 6.19% |
| B. garinii*** | 39.58% | 20.83% | 0.00% | 6.25% | 8.33% | 2.08% | 16.67% | 2.08% | 4.17% |
| B. bavariensis | 31.58% | 5.26% | 5.26% | 10.53% | 10.53% | 5.26% | 21.05% | 5.26% | 5.26% |
| B. burgdorferi s.s. | 12.73% | 12.73% | 10.91% | 16.36% | 5.45% | 7.27% | 18.18% | 5.45% | 10.91% |
| B. spielmanii | 26.09% | 30.43% | 0.00% | 4.35% | 4.35% | 8.70% | 13.04% | 13.04% | 0.00% |
| B. valaisiana *** | 83.33% | 5.56% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 11.11% |
| B.bissettii/carolinensis | 16.67% | 0.00% | 16.67% | 16.67% | 16.67% | 33.33% | 0.00% | 0.00% | 0.00% |
| E. europaeus | Total | Ear | Muscle | Blood | Lungs | Liver | Spleen | Urinary Bladder | Kidney | Brain |
|---|---|---|---|---|---|---|---|---|---|---|
| B. afzelii | 74 | 25.68% | 22.97% | 1.35% | 4.05% | 8.11% | 6.76% | 14.86% | 8.11% | 8.11% |
| B. garinii | 16 | 12.50% | 31.25% | 0.00% | 0.00% | 18.75% | 0.00% | 31.25% | 6.25% | 0.00% |
| B. bavariensis | 18 | 27.78% | 5.56% | 5.56% | 11.11% | 11.11% | 5.56% | 22.22% | 5.56% | 5.56% |
| B. burgdorferi s.s. | 23 | 4.35% | 4.35% | 17.39% | 21.74% | 4.35% | 8.70% | 21.74% | 4.35% | 13.04% |
| B. spielmanii | 23 | 26.09% | 30.43% | 0.00% | 4.35% | 4.35% | 8.70% | 13.04% | 13.04% | 0.00% |
| B. valaisiana | 2 | 0.00% | 50.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 50.00% |
| B. bissettii/carolinensis | 5 | 20.00% | 0.00% | 20.00% | 20.00% | 0.00% | 40.00% | 0.00% | 0.00% | 0.00% |
| E. roumanicus | Total | Ear | Muscle | Blood | Lungs | Liver | Spleen | Urinary Bladder | Kidney | Brain |
| B. afzelii | 12 | 16.67% | 33.33% | 0.00% | 0.00% | 0.00% | 16.67% | 25.00% | 8.33% | 0.00% |
| B. garinii | 4 | 0.00% | 50.00% | 0.00% | 0.00% | 0.00% | 0.00% | 50.00% | 0.00% | 0.00% |
| B. bavariensis | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| B. burgdorferi s.s. | 16 | 0.00% | 4.12% | 1.03% | 2.06% | 1.03% | 2.06% | 3.09% | 1.03% | 2.06% |
| B. spielmanii | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| B. valaisiana | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| B. bissettii/carolinensis | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| S. vulgaris | Total | Ear | Muscle | Blood | Lungs | Liver | Spleen | Urinary Bladder | Kidney | Brain |
| B. afzelii | 5 | 40.00% | 0.00% | 20.00% | 20.00% | 0.00% | 0.00% | 20.00% | 0.00% | 0.00% |
| B. garinii | 9 | 22.22% | 11.11% | 0.00% | 33.33% | 11.11% | 11.11% | 11.11% | 0.00% | 0.00% |
| B. bavariensis | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| B. burgdorferi s.s. | 16 | 37.50% | 12.50% | 6.25% | 12.50% | 6.25% | 0.00% | 12.50% | 6.25% | 6.25% |
| B. spielmanii | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| B. valaisiana | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| B. bissettii/carolinensis | 1 | 0.00% | 0.00% | 0.00% | 0.00% | 100.00% | 0.00% | 0.00% | 0.00% | 0.00% |
| T. merula | Total | Skin | Muscle | Blood | Liver | Brain | ||||
| B. afzelii | 6 | 66.67% | 33.33% | 0.00% | 0.00% | 0.00% | ||||
| B. garinii | 19 | 78.95% | 10.53% | 0.00% | 0.00% | 10.53% | ||||
| B. bavariensis | 1 | 100.00% | 0.00% | 0.00% | 0.00% | 0.00% | ||||
| B. burgdorferi s.s. | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | ||||
| B. spielmanii | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% | ||||
| B. valaisiana | 16 | 93.75% | 0.00% | 0.00% | 0.00% | 6.25% | ||||
| B. bissettii/carolinensis | 0 | 0.00% | 0.00% | 0.00% | 0.00% | 0.00% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Majerová, K.; Hönig, V.; Houda, M.; Papežík, P.; Fonville, M.; Sprong, H.; Rudenko, N.; Golovchenko, M.; Černá Bolfíková, B.; Hulva, P.; et al. Hedgehogs, Squirrels, and Blackbirds as Sentinel Hosts for Active Surveillance of Borrelia miyamotoi and Borrelia burgdorferi Complex in Urban and Rural Environments. Microorganisms 2020, 8, 1908. https://doi.org/10.3390/microorganisms8121908
Majerová K, Hönig V, Houda M, Papežík P, Fonville M, Sprong H, Rudenko N, Golovchenko M, Černá Bolfíková B, Hulva P, et al. Hedgehogs, Squirrels, and Blackbirds as Sentinel Hosts for Active Surveillance of Borrelia miyamotoi and Borrelia burgdorferi Complex in Urban and Rural Environments. Microorganisms. 2020; 8(12):1908. https://doi.org/10.3390/microorganisms8121908
Chicago/Turabian StyleMajerová, Karolina, Václav Hönig, Michal Houda, Petr Papežík, Manoj Fonville, Hein Sprong, Natalie Rudenko, Maryna Golovchenko, Barbora Černá Bolfíková, Pavel Hulva, and et al. 2020. "Hedgehogs, Squirrels, and Blackbirds as Sentinel Hosts for Active Surveillance of Borrelia miyamotoi and Borrelia burgdorferi Complex in Urban and Rural Environments" Microorganisms 8, no. 12: 1908. https://doi.org/10.3390/microorganisms8121908