Treponema spp. Isolated from Bovine Digital Dermatitis Display Different Pathogenicity in a Murine Abscess Model

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Treponema spp. Inoculum
2.3. Necropsy
2.4. Bacterial Culture
2.5. Treponema spp. qPCR
2.6. Histopathology
2.7. Transcriptional Gene Expression of Innate Immune Factors in Murine Abscesses
2.8. Statistical Analyses
3. Results
3.1. DD-Associated Treponema spp. Induce Abscesses of Variable Sizes
3.2. Viable Treponema spp. Detected by Culture
3.3. Variable Numbers of Treponema spp. Detected
3.4. Differences in Treponema spp. Induced Lesion
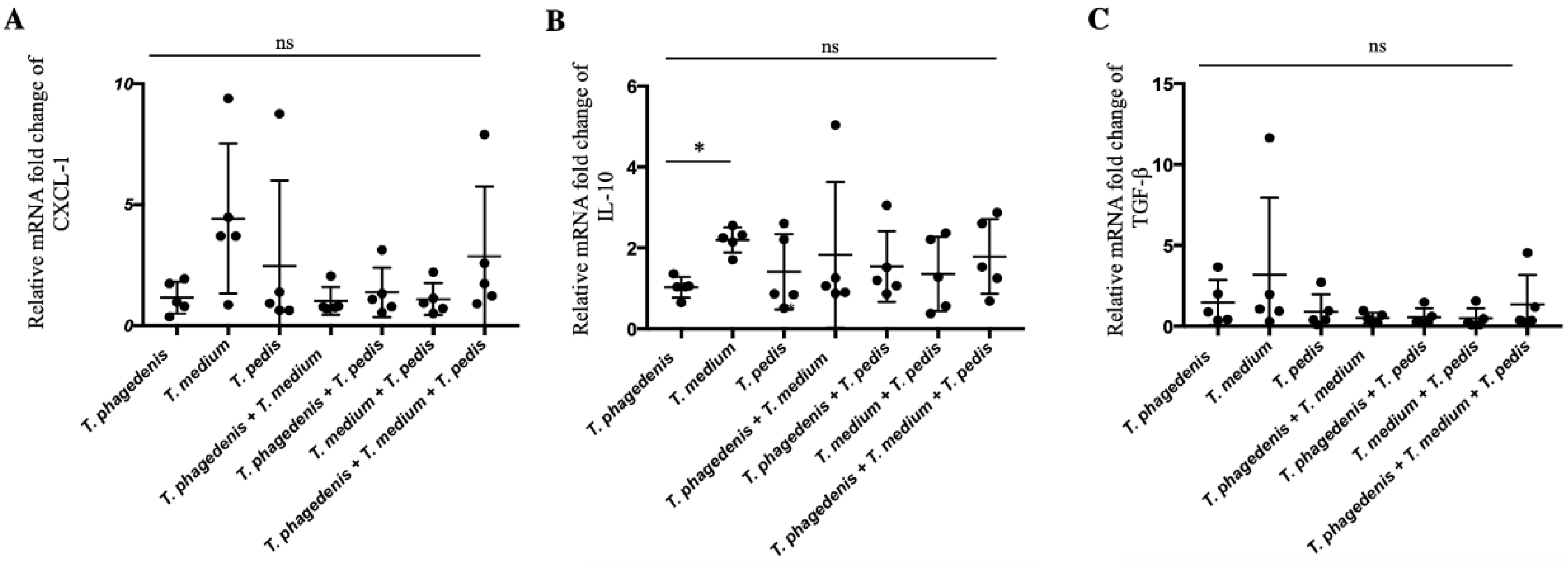
3.5. Treponema Induced Differential Expression of Innate Immune Factors
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Holzhauer, M.; Bartels, C.J.M.; Döpfer, D.; van Schaik, G. Clinical course of digital dermatitis lesions in an endemically infected herd without preventive herd strategies. Vet. J. 2008, 177, 222–230. [Google Scholar] [CrossRef]
- Berry, S.L. Diseases of the digital soft tissues. Vet. Clin. N. Am. Food Anim. Pract. 2001, 17, 129–142. [Google Scholar] [CrossRef]
- Losinger, W.C. Economic impacts of reduced milk production associated with papillomatous digital dermatitis in dairy cows in the USA. J. Dairy Res. 2006, 73, 244–256. [Google Scholar] [CrossRef] [PubMed]
- Cha, E.; Hertl, J.A.; Bar, D.; Gröhn, Y.T. The cost of different types of lameness in dairy cows calculated by dynamic programming. Prev. Vet. Med. 2010, 97, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Bruijnis, M.R.N.; Beerda, B.; Hogeveen, H.; Stassen, E.N. Assessing the welfare impact of foot disorders in dairy cattle by a modeling approach. Animal 2012, 6, 962–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gotoh, Y.; Chiba, K.; Sekiyama, Y.; Okada, K.; Hayashi, T.; Misawa, N. 16S rRNA-based amplicon analysis of changes in the bacterial population in the lesions of papillomatous digital dermatitis in dairy cattle after topical treatment with allyl isothiocyanate. Microbiol. Immunol. 2020. [Google Scholar] [CrossRef]
- Nielsen, M.W.; Strube, M.L.; Isbrand, A.; Al-Medrasi, W.D.H.M.; Boye, M.; Jensen, T.K.; Klitgaard, K. Potential bacterial core species associated with digital dermatitis in cattle herds identified by molecular profiling of interdigital skin samples. Vet. Microbiol. 2016, 186, 139–149. [Google Scholar] [CrossRef] [Green Version]
- Krull, A.C.; Shearer, J.K.; Gorden, P.J.; Cooper, V.L.; Phillips, G.J.; Plummer, P.J. Deep sequencing analysis reveals temporal microbiota changes associated with development of bovine digital dermatitis. Infect. Immun. 2014, 82, 3359–3373. [Google Scholar] [CrossRef] [Green Version]
- Evans, N.J.; Murray, R.D.; Carter, S.D. Bovine digital dermatitis: Current concepts from laboratory to farm. Vet. J. 2016, 211, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Evans, N.J.; Brown, J.M.; Demirkan, I.; Singh, P.; Getty, B.; Timofte, D.; Vink, W.D.; Murray, R.D.; Blowey, R.W.; Birtles, R.J.; et al. Association of unique, isolated treponemes with bovine digital dermatitis lesions. J. Clin. Microbiol. 2009, 47, 689–696. [Google Scholar] [CrossRef] [Green Version]
- Klitgaard, K.; Boye, M.; Capion, N.; Jensen, T.K. Evidence of multiple Treponema phylotypes involved in bovine digital dermatitis as shown by 16S rRNA gene analysis and fluorescence in situ hybridization. J. Clin. Microbiol. 2008, 46, 3012–3020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nordhoff, M.; Moter, A.; Schrank, K.; Wieler, L.H. High prevalence of treponemes in bovine digital dermatitis-A molecular epidemiology. Vet. Microbiol. 2008, 131, 293–300. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orsel, K.; Plummer, P.; Shearer, J.; De Buck, J.; Carter, S.D.; Guatteo, R.; Barkema, H.W. Missing pieces of the puzzle to effectively control digital dermatitis. Transbound. Emerg. Dis. 2018, 65, 186–198. [Google Scholar] [CrossRef]
- Beninger, C.; Naqvi, S.A.; Naushad, S.; Orsel, K.; Luby, C.; Derakhshani, H.; Khafipour, E.; De Buck, J. Associations between digital dermatitis lesion grades in dairy cattle and the quantities of four Treponema species. Vet. Res. 2018, 49, 111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demirkan, I.; Erdoğan, M.; Demirkan, A.Ç.; Bozkurt, F.; Altındiş, M.; Navruz, F.Z.; Köse, Z. Isolation and identification of Treponema pedis and Treponema phagedenis-like organisms from bovine digital dermatitis lesions found in dairy cattle in Turkey. J. Dairy Sci. 2018, 101, 10317–10326. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, L.E.; Clegg, S.R.; Angell, J.W.; Newbrook, K.; Blowey, R.W.; Carter, S.D.; Bell, J.; Duncan, J.S.; Grove-White, D.H.; Murray, R.D.; et al. High-level association of bovine digital dermatitis Treponema spp. with contagious ovine digital dermatitis lesions and presence of Fusobacterium necrophorum and Dichelobacter nodosus. J. Clin. Microbiol. 2015, 53, 1628–1638. [Google Scholar] [CrossRef] [Green Version]
- Han, S.; Mansfield, K.G.; Bradway, D.S.; Besser, T.E.; Read, D.H.; Haldorson, G.J.; Alt, D.P.; Wilson-Welder, J.H. Treponeme-associated hoof disease of free-ranging elk (Cervus elaphus) in Southwestern Washington State, USA. Vet. Pathol. 2019, 56, 118–132. [Google Scholar] [CrossRef] [Green Version]
- Clegg, S.R.; Carter, S.D.; Stewart, J.P.; Amin, D.M.; Blowey, R.W.; Evans, N.J. Bovine ischaemic teat necrosis: A further potential role for digital dermatitis treponemes. Vet. Rec. 2016, 178, 71. [Google Scholar] [CrossRef]
- Clegg, S.R.; Sullivan, L.E.; Bell, J.; Blowey, R.W.; Carter, S.D.; Evans, N.J. Detection and isolation of digital dermatitis treponemes from skin and tail lesions in pigs. Res. Vet. Sci. 2016, 104, 64–70. [Google Scholar] [CrossRef]
- Lamont, R.J.; Hajishengallis, G. Polymicrobial synergy and dysbiosis in inflammatory disease. Trends Mol. Med. 2015, 21, 172–183. [Google Scholar] [CrossRef] [Green Version]
- Tan, K.H.; Seers, C.A.; Dashper, S.G.; Mitchell, H.L.; Pyke, J.S.; Meuric, V.; Slakeski, N.; Cleal, S.M.; Chambers, J.L.; McConville, M.J.; et al. Porphyromonas gingivalis and Treponema denticola exhibit metabolic symbioses. PLoS Pathog. 2014, 10, e1003955. [Google Scholar] [CrossRef] [PubMed]
- Grenier, D. Nutritional interactions between two suspected periodontopathogens, Treponema denticola and Porphyromonas gingivalis. Infect. Immun. 1992, 60, 5298–5301. [Google Scholar] [CrossRef] [Green Version]
- Zhu, Y.; Dashper, S.G.; Chen, Y.Y.; Crawford, S.; Slakeski, N.; Reynolds, E.C. Porphyromonas gingivalis and Treponema denticola synergistic polymicrobial biofilm development. PLoS ONE 2013, 8, e71727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamada, M.; Ikegami, A.; Kuramitsu, H.K. Synergistic biofilm formation by Treponema denticola and Porphyromonas gingivalis. FEMS Microbiol. Lett. 2005, 250, 271–277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orth, R.K.-H.; O’Brien-Simpson, N.M.; Dashper, S.G.; Reynolds, E.C. Synergistic virulence of Porphyromonas gingivalis and Treponema denticola in a murine periodontitis model. Mol. Oral Microbiol. 2011, 26, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Kesavalu, L.; Holt, S.C.; Ebersole, J.L. Virulence of a polymicrobic complex, Treponema denticola and Porphyromonas gingivalis, in a murine model. Oral Microbiol. Immunol. 1998, 13, 373–377. [Google Scholar] [CrossRef]
- Bodet, C.; Chandad, F.; Grenier, D. Potentiel pathogénique de Porphyromonas gingivalis, Treponema denticola et Tannerella forsythia, le complexe bactérien rouge associé à la parodontite. Pathol. Biol. 2007, 55, 154–162. [Google Scholar] [CrossRef]
- Hajishengallis, G.; Lamont, R.J. Beyond the red complex and into more complexity: The polymicrobial synergy and dysbiosis (PSD) model of periodontal disease etiology. Mol. Oral Microbiol. 2012, 27, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Abe, T.; Hajishengallis, G. Optimization of the ligature-induced periodontitis model in mice. J. Immunol. Methods 2013, 394, 49–54. [Google Scholar] [CrossRef] [Green Version]
- Kesavalu, L.; Walker, S.G.; Holt, S.C.; Crawley, R.R.; Ebersole, J.L. Virulence characteristics of oral treponemes in a murine model. Infect. Immun. 1997, 65, 5096–5102. [Google Scholar] [CrossRef] [Green Version]
- Washizu, M.; Ishihara, K.; Honma, K.; Okuda, K. Effects of a mixed infection with Porphyromonas gingivalis and Treponema denticola on abscess formation and immune responses in mice. Bull. Tokyo Dent. Coll. 2003, 44, 141–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, M.K.; Alt, D.P.; Zuerner, R.L. Lesion formation and antibody response induced by papillomatous digital dermatitis-associated spirochetes in a murine abscess model. Infect. Immun. 2007, 75, 4400–4408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gomez, A.; Cook, N.B.; Bernardoni, N.D.; Rieman, J.; Dusick, A.F.; Hartshorn, R.; Socha, M.T.; Read, D.H.; Döpfer, D. An experimental infection model to induce digital dermatitis infection in cattle. J. Dairy Sci. 2012, 95, 1821–1830. [Google Scholar] [CrossRef] [PubMed]
- Krull, A.C.; Cooper, V.L.; Coatney, J.W.; Shearer, J.K.; Gorden, P.J.; Plummer, P.J. A Highly effective protocol for the rapid and consistent induction of digital dermatitis in holstein calves. PLoS ONE 2016, 11, e0154481. [Google Scholar] [CrossRef] [Green Version]
- Wilson-Welder, J.H.; Nally, J.E.; Alt, D.P.; Palmer, M.V.; Coatney, J.; Plummer, P. Experimental transmission of bovine digital dermatitis to sheep: Development of an infection model. Vet. Pathol. 2018, 55, 245–257. [Google Scholar] [CrossRef] [Green Version]
- Arrazuria, R.; Caddey, B.; Cobo, E.R.; Barkema, H.W.; DeBuck, J. Digital dermatitis Treponema spp. mixed community responds differently to culture media and serum supplementation. Anaerobe 2020. submitted. [Google Scholar]
- Clegg, S.R.; Carter, S.D.; Birtles, R.J.; Brown, J.M.; Anthony Hart, C.; Evans, N.J. Multilocus sequence typing of pathogenic treponemes isolated from cloven-hoofed animals and comparison to treponemes isolated from humans. Appl. Environ. Microbiol. 2016, 82, 4523–4536. [Google Scholar] [CrossRef] [Green Version]
- Willcox, R.R.; Guthe, T. Treponema pallidum. A bibliographical review of the morphology, culture and survival of T. pallidum and associated organisms: Introduction. Bull. World Health Organ. 1966, 35, 5–9. [Google Scholar]
- Moreira, T.F.; Facury Filho, E.J.; Carvalho, A.U.; Strube, M.L.; Nielsen, M.W.; Klitgaard, K.; Jensen, T.K. Pathology and bacteria related to digital dermatitis in dairy cattle in all year round grazing system in Brazil. PLoS ONE 2018, 13, e0193870. [Google Scholar] [CrossRef] [Green Version]
- Pringle, M.; Bergsten, C.; Fernström, L.-L.; Höök, H.; Johansson, K.-E. Isolation and characterization of Treponema phagedenis-like spirochetes from digital dermatitis lesions in Swedish dairy cattle. Acta Vet. Scand. 2008, 50, 40. [Google Scholar] [CrossRef] [Green Version]
- Wilson-Welder, J.H.; Elliott, M.K.; Zuerner, R.L.; Bayles, D.O.; Alt, D.P.; Stanton, T.B. Biochemical and molecular characterization of Treponema phagedenis-like spirochetes isolated from a bovine digital dermatitis lesion. BMC Microbiol. 2013, 13, 280. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klitgaard, K.; Bretó, A.F.; Boye, M.; Jensen, T.K. Targeting the treponemal microbiome of digital dermatitis infections by high-resolution phylogenetic analyses and comparison with fluorescent in situ hybridization. J. Clin. Microbiol. 2013, 51, 2212–2219. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zinicola, M.; Lima, F.; Lima, S.; Machado, V.; Gomez, M.; Döpfer, D.; Guard, C.; Bicalho, R. Altered microbiomes in bovine digital dermatitis lesions, and the gut as a pathogen reservoir. PLoS ONE 2015, 10, e0120504. [Google Scholar] [CrossRef] [PubMed]
- Angell, J.W.; Clegg, S.R.; Grove-White, D.H.; Blowey, R.W.; Carter, S.D.; Duncan, J.S.; Evans, N.J. Survival of contagious ovine digital dermatitis (CODD)-Associated treponemes on disposable gloves after handling CODD-Affected feet. Vet. Rec. 2017, 181, 89. [Google Scholar] [CrossRef] [Green Version]
- Siqueira, J.F.; Rocas, I.N. Treponema species associated with abscesses of endodontic origin. Oral Microbiol. Immunol. 2004, 19, 336–339. [Google Scholar] [CrossRef]
- Stamm, L.V.; Walker, R.L.; Read, D.H. Genetic diversity of bovine ulcerative mammary dermatitis-associated Treponema. Vet. Microbiol. 2009, 136, 192–196. [Google Scholar] [CrossRef]
- Foschi, F.; Izard, J.; Sasaki, H.; Sambri, V.; Prati, C.; Müller, R.; Stashenko, P. Treponema denticola in disseminating endodontic infections. J. Dent. Res. 2006, 85, 761–765. [Google Scholar] [CrossRef] [Green Version]
- Chukkapalli, S.S.; Rivera, M.F.; Velsko, I.M.; Lee, J.Y.; Chen, H.; Zheng, D.; Bhattacharyya, I.; Gangula, P.R.; Lucas, A.R.; Kesavalu, L. Invasion of oral and aortic tissues by oral spirochete Treponema denticola in ApoE-/- mice causally links periodontal disease and atherosclerosis. Infect. Immun. 2014, 82, 1959–1967. [Google Scholar] [CrossRef] [Green Version]
- Salazar, J.C.; Rathi, A.; Michael, N.L.; Radolf, J.D.; Jagodzinski, L.L. Assessment of the kinetics of Treponema pallidum dissemination into blood and tissues in experimental syphilis by real-time quantitative PCR. Infect. Immun. 2007, 75, 2954–2958. [Google Scholar] [CrossRef] [Green Version]
- Schwarzova, K.; Ciznar, I.; Svihrova, V.; Hudeckova, H. Initial attachment of Borrelia burgdorferi spirochetes to Vero cells. Bratislava Med. J. 2019, 120, 872–875. [Google Scholar] [CrossRef] [Green Version]
- Chiu, S.-H.; Pan, M.-J. Detection and Treatment of Leptospirosis Kidney Disease. In Leptospirosis and the Kidney; S. Karger AG: Basel, Switzerland, 2019; Volume 7, pp. 37–46. [Google Scholar]
- Döpfer, D.; Anklam, K.; Mikheil, D.; Ladell, P. Growth curves and morphology of three Treponema subtypes isolated from digital dermatitis in cattle. Vet. J. 2012, 193, 685–693. [Google Scholar] [CrossRef] [PubMed]
- Miklossy, J.; Kasas, S.; Zurn, A.D.; McCall, S.; Yu, S.; McGeer, P.L. Persisting atypical and cystic forms of Borrelia burgdorferi and local inflammation in Lyme neuroborreliosis. J. Neuroinflammation 2008, 5, 40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saraiva, M.; O’Garra, A. The regulation of IL-10 production by immune cells. Nat. Rev. Immunol. 2010, 10, 170–181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watts, K.M.; Fodor, C.; Beninger, C.; Lahiri, P.; Arrazuria, R.; De Buck, J.; Knight, C.G.; Orsel, K.; Barkema, H.W.; Cobo, E.R. A differential innate immune response in active and chronic stages of bovine infectious digital dermatitis. Front. Microbiol. 2018, 9, 1586. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Culture (%) 2 | Average Size 1 (SD) | |
|---|---|---|---|
| Culture Positive | Culture Negative | ||
| T. phagedenis | 0 | - | 5.3 (1.89) |
| T. medium | 80 | 311.9 (77.92) | 228.0 (84.85) |
| T. pedis | 0 | - | 72.9 (17.62) |
| T. phagedenis + T. medium | 20 | 149.0 (7.07) | 40.6 (27.86) |
| T. phagedenis + T. pedis | 0 | - | 22.2 (18.12) |
| T. medium + T. pedis | 30 | 124.0 (3.46) | 57.0 (39.32) |
| T. phagedenis + T. medium + T. pedis | 50 | 173.8 (48.62) | 120.4 (26.84) |
| Group | T. phagedenis LGE/g Tissue (SD) | T. medium LGE/g Tissue (SD) | T. pedis LGE/g Tissue (SD) |
|---|---|---|---|
| T. phagedenis | 7.6 (0.72) | - | - |
| T. medium | - | 6.8 (0.30) | - |
| T. pedis | - | - | 8.2 (0.33) |
| T. phagedenis + T. medium | 8.0 (0.53) | 6.3 (0.37) | - |
| T. phagedenis + T. pedis | 8.3 (0.38) | - | 8.0 (0.48) |
| T. medium + T. pedis | - | 6.7 (0.38) | 7.7 (0.4) |
| T. phagedenis + T. medium + T. pedis | 8.3 (0.39) | 6.7 (0.47) | 8.1 (0.37) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arrazuria, R.; Knight, C.G.; Lahiri, P.; Cobo, E.R.; Barkema, H.W.; De Buck, J. Treponema spp. Isolated from Bovine Digital Dermatitis Display Different Pathogenicity in a Murine Abscess Model. Microorganisms 2020, 8, 1507. https://doi.org/10.3390/microorganisms8101507
Arrazuria R, Knight CG, Lahiri P, Cobo ER, Barkema HW, De Buck J. Treponema spp. Isolated from Bovine Digital Dermatitis Display Different Pathogenicity in a Murine Abscess Model. Microorganisms. 2020; 8(10):1507. https://doi.org/10.3390/microorganisms8101507
Chicago/Turabian StyleArrazuria, Rakel, Cameron G. Knight, Priyoshi Lahiri, Eduardo R. Cobo, Herman W. Barkema, and Jeroen De Buck. 2020. "Treponema spp. Isolated from Bovine Digital Dermatitis Display Different Pathogenicity in a Murine Abscess Model" Microorganisms 8, no. 10: 1507. https://doi.org/10.3390/microorganisms8101507