Diversity of Pectobacteriaceae Species in Potato Growing Regions in Northern Morocco

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
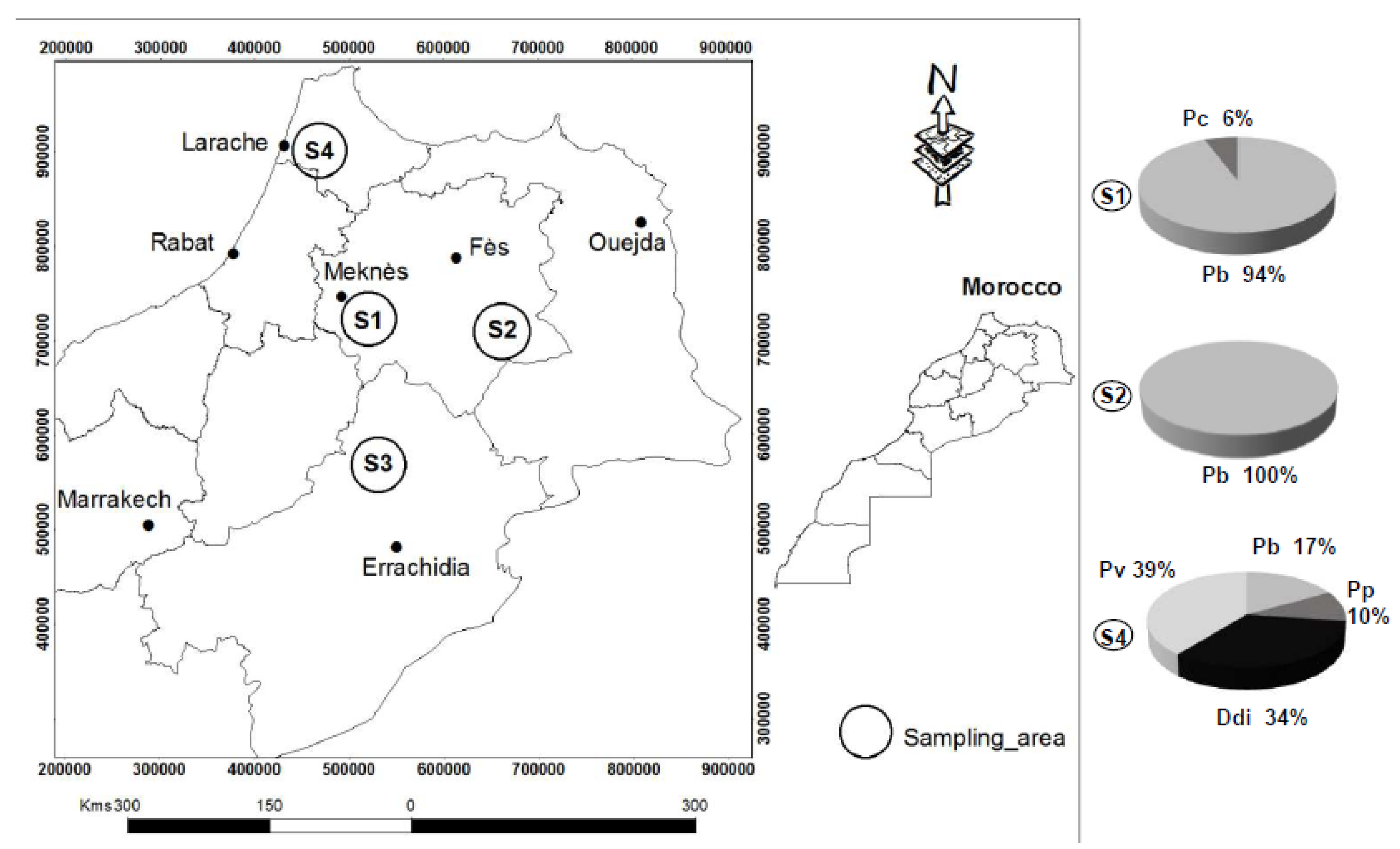
2.1. Sampling and Isolation of Pectinolytic Bacteria
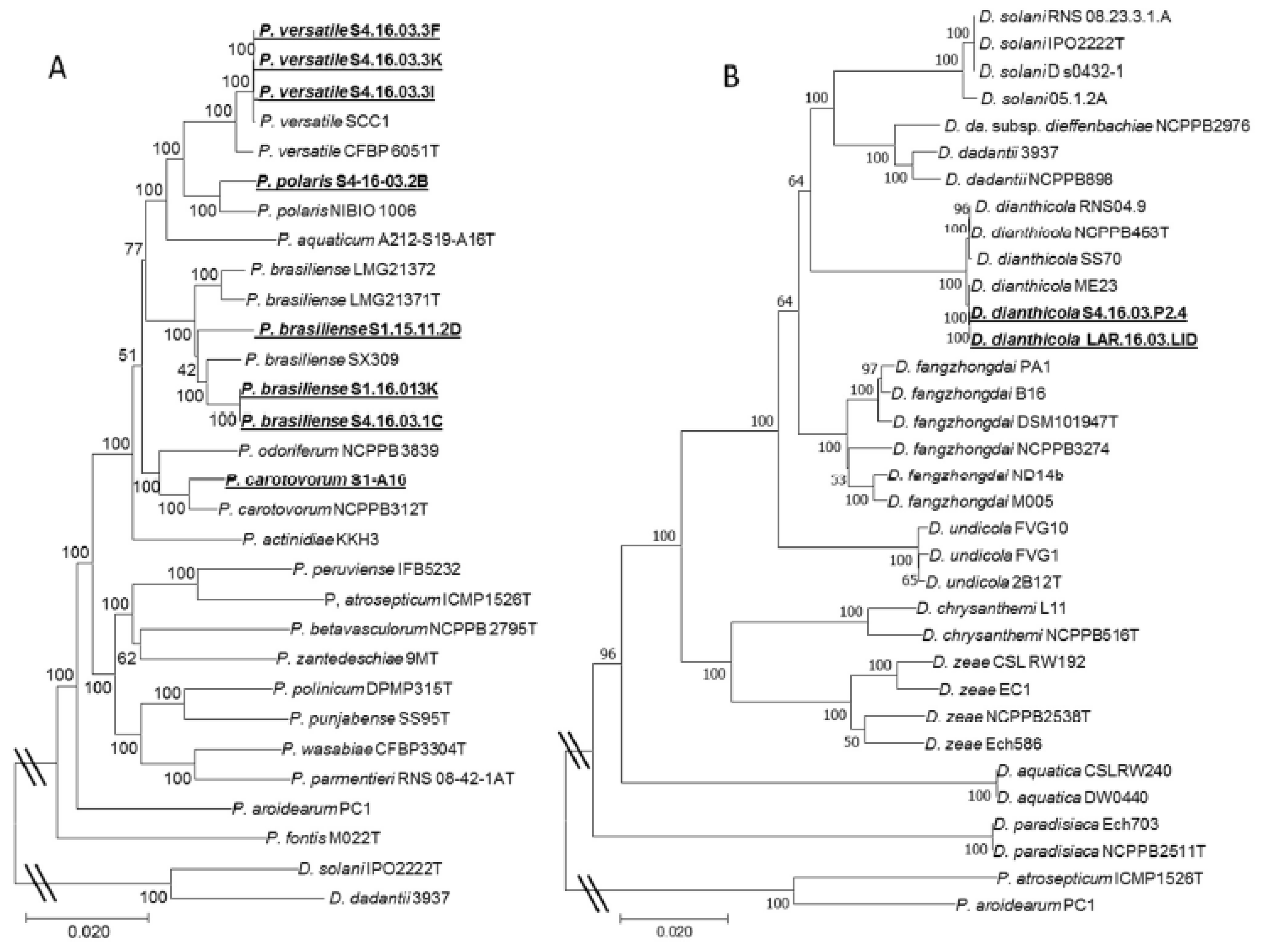
2.2. Molecular Characterization of Pectobacterium and Dickeya Isolates
2.3. Genome Sequencing
2.4. Genome Assembly
2.5. Genome Analysis
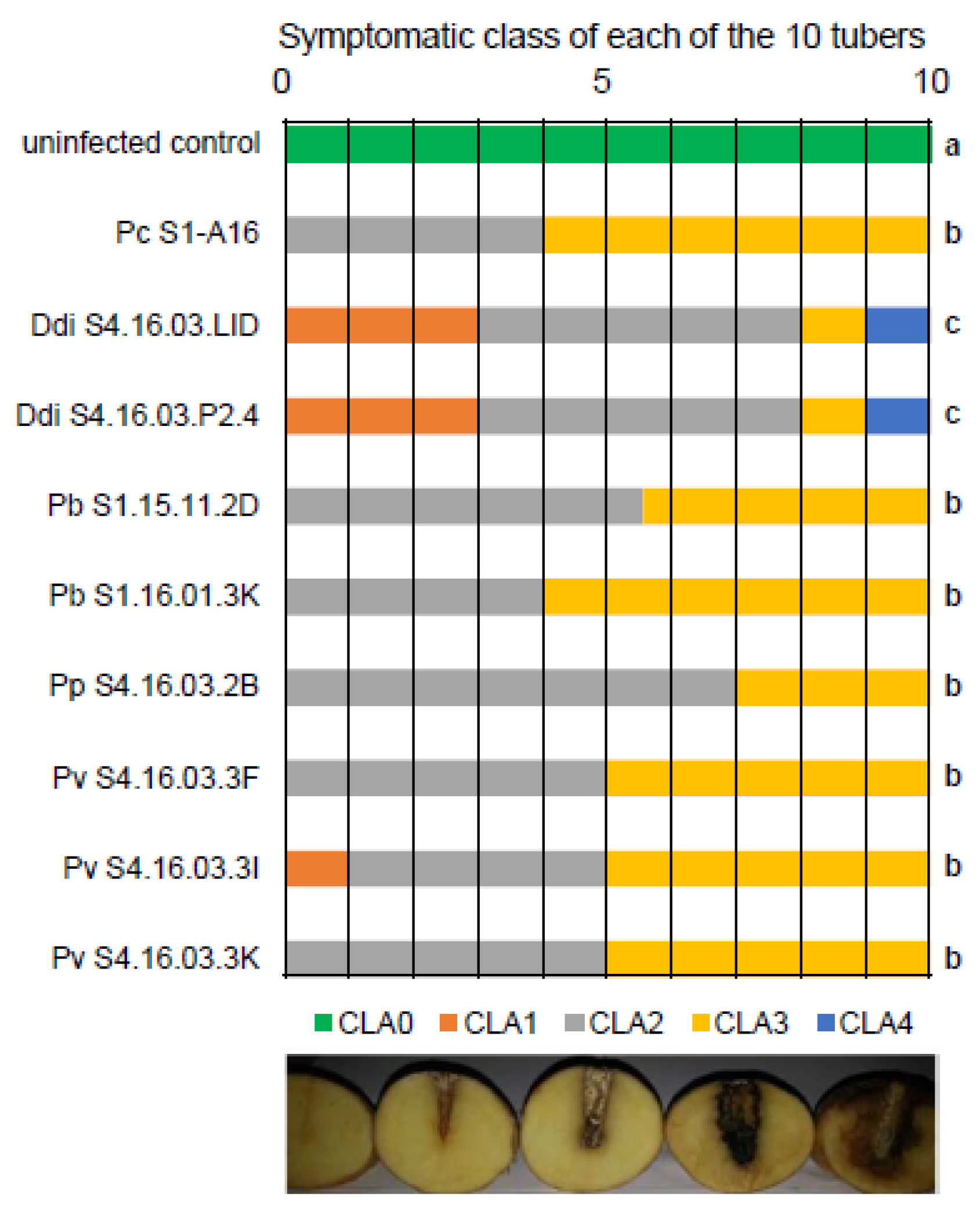
2.6. Potato Tuber Rotting Assays
3. Results
3.1. Diversity of the Pectinolytic Dickeya and Pectobacterium in Northern Morocco
3.2. Draft Genomes of 10 Pectinolytic Bacteria from Northern Morocco
3.3. Aggressiveness of the Pectinolytic Bacteria from Northern Morocco
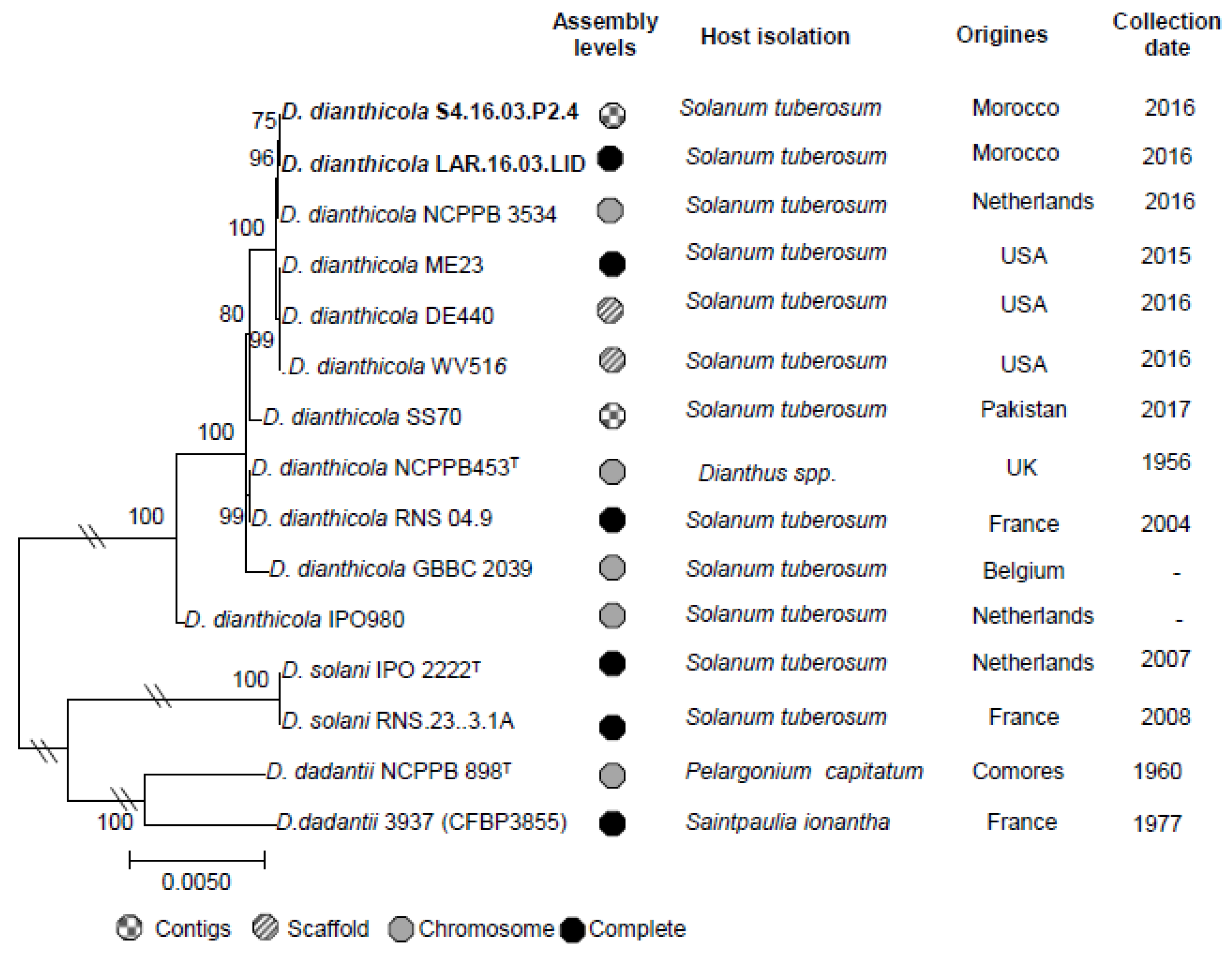
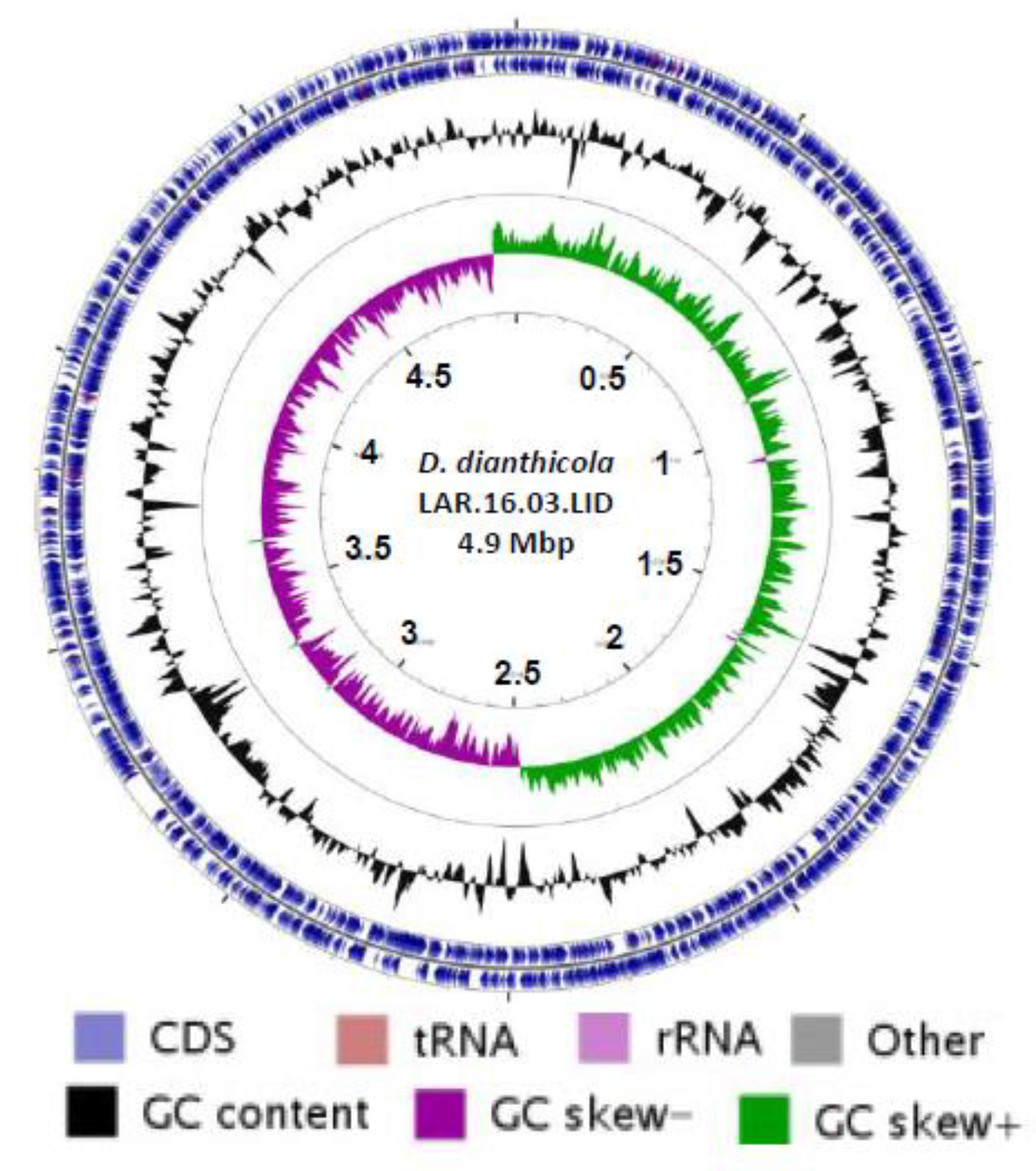
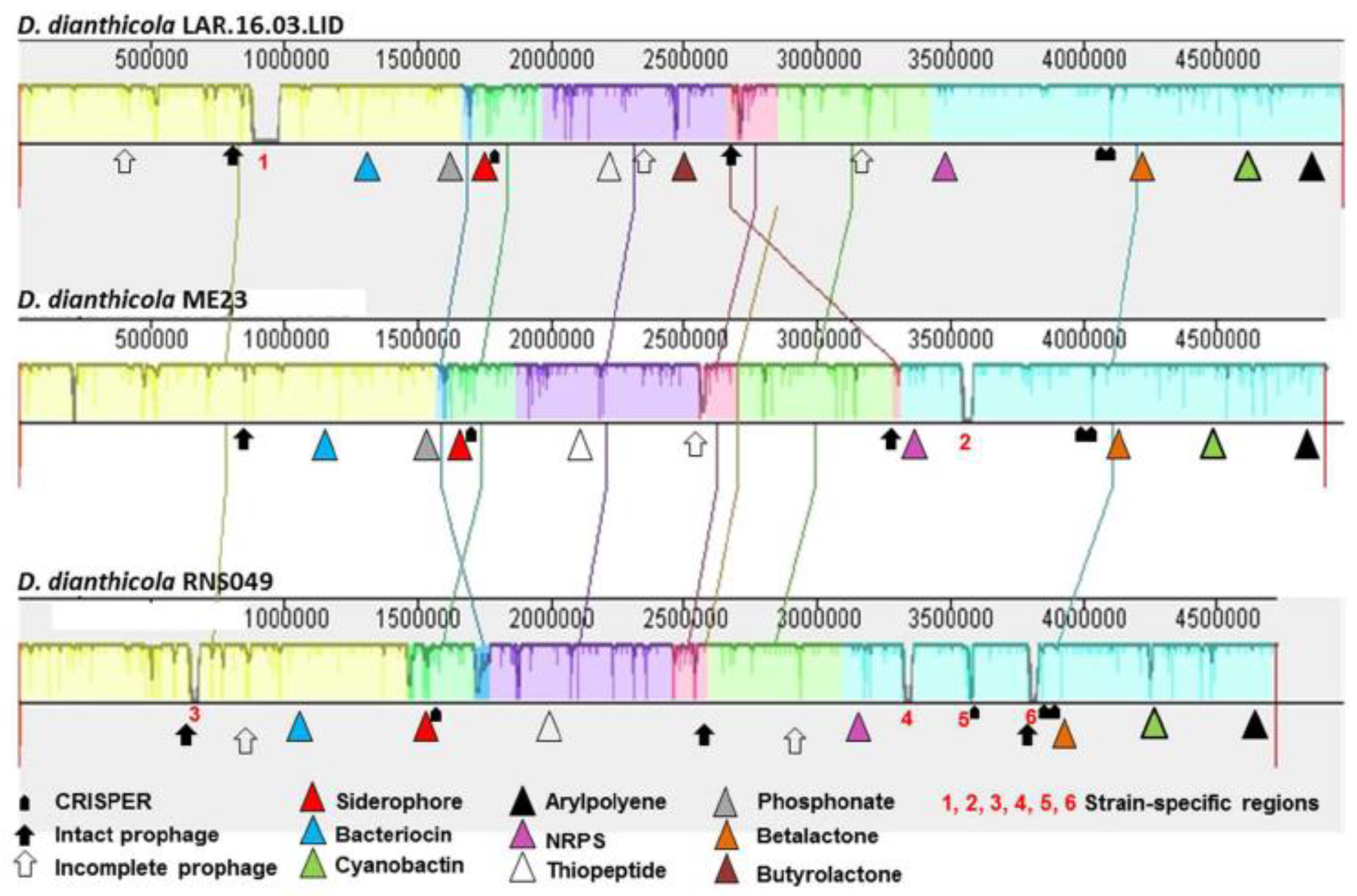
3.4. Complete Genome of D. dianthicola LAR.16.03.LID
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Czajkowski, R.; Pérombelon, M.C.M.; Van Veen, J.A.; Van der Wolf, J.M. Control of blackleg and tuber soft rot of potato caused by Pectobacterium and Dickeya species: A review. Plant Pathol. 2011, 60, 999–1013. [Google Scholar] [CrossRef]
- Mansfield, J.; Genin, S.; Magori, S.; Citovsky, V.; Sriariyanum, M.; Ronald, P.; Dow, M.; Verdier, V.; Beer, S.V.; Machado, M.A.; et al. Top 10 plant pathogenic bacteria in molecular plant pathology. Mol. Plant Pathol. 2012, 13, 614–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leonard, S.; Hommais, F.; Nasser, W.; Reverchon, S. Plant–phytopathogen interactions: Bacterial responses to environmental and plant stimuli. Environ. Microbiol. 2017, 19, 1689–1716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérombelon, M.C.M. Potato diseases caused by soft rot erwinias: An overview of pathogenesis. Plant Pathol. 2002, 51, 1–12. [Google Scholar] [CrossRef]
- De Boer, S.H.; Verdonck, L.; Vruggink, H.; Harju, P.; Bång, H.O.; Ley, J. De Serological and biochemical variation among potato strains of Erwinia carotovora subsp. atroseptica and their taxonomic relationship to other E. carotovora strains. J. Appl. Bacteriol. 1987, 63, 487–495. [Google Scholar] [CrossRef]
- Khayi, S.; Cigna, J.; Chong, T.M.; Qu�tu-Laurent, A.; Chan, K.G.; Helias, V.; Faure, D. Transfer of the potato plant isolates of Pectobacterium wasabiae to Pectobacterium parmentieri sp. nov. Int. J. Syst. Evol. Microbiol. 2016, 66, 5379–5383. [Google Scholar] [CrossRef]
- Portier, P.; Pédron, J.; Taghouti, G.; Fischer-Le Saux, M.; Caullireau, E.; Bertrand, C.; Laurent, A.; Chawki, K.; Oulgazi, S.; Moumni, M.; et al. Elevation of Pectobacterium carotovorum subsp. odoriferum to species level as Pectobacterium odoriferum sp. nov., proposal of Pectobacterium brasiliense sp. nov. and Pectobacterium actinidiae sp. nov., emended description of Pectobacterium carotovorum. Int. J. Syst. Evol. Microbiol. 2019, 69, 3207–3216. [Google Scholar] [CrossRef]
- Smith, C. Variation in the Pathogenicity and Aggressiveness of Strains of Erwinia carotovora subsp. carotovora Isolated from Different Hosts. Plant Dis. 2007, 74, 505. [Google Scholar] [CrossRef]
- Dees, M.W.; Lysøe, E.; Rossmann, S.; Perminow, J.; Brurberg, M.B. Pectobacterium polaris sp. nov., isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2017, 67, 5222–5229. [Google Scholar] [CrossRef]
- Sarfraz, S.; Riaz, K.; Oulghazi, S.; Cigna, J.; Sahi, S.T.; Khan, S.H.; Faure, D. Pectobacterium punjabense sp. nov., isolated from blackleg symptoms of potato plants in Pakistan. Int. J. Syst. Evol. Microbiol. 2018, 68, 3551–3556. [Google Scholar] [CrossRef]
- Waleron, M.; Misztak, A.; Waleron, M.; Franczuk, M.; Wielgomas, B.; Waleron, K. Transfer of Pectobacterium carotovorum subsp. carotovorum strains isolated from potatoes grown at high altitudes to Pectobacterium peruviense sp. nov. Syst. Appl. Microbiol. 2018, 41, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Waleron, M.; Waleron, K.; Lojkowska, E. Characterization of Pectobacterium carotovorum subsp. odoriferum causing soft rot of stored vegetables. Eur. J. Plant Pathol. 2014, 139, 457–469. [Google Scholar] [CrossRef] [Green Version]
- Goto, M.; Matsumoto, K. Erwinia carotovora subsp. wasabiae subsp. nov. Isolated from Diseased Rhizomes and Fibrous Roots of Japanese Horseradish (Eutrema wasabi Maxim.). Int. J. Syst. Bacteriol. 1987, 37, 130–135. [Google Scholar] [CrossRef] [Green Version]
- Gardan, L.; Gouy, C.; Christen, R.; Samson, R. Elevation of three subspecies of Pectobacterium carotovorum to species level: Pectobacterium atrosepticum sp. nov., Pectobacterium betavasculorum sp. nov. and Pectobacterium wasabiae sp. nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 381–391. [Google Scholar] [CrossRef] [PubMed]
- Nabhan, S.; de Boer, S.H.; Maiss, E.; Wydra, K. Pectobacterium aroidearum sp. nov., a soft rot pathogen with preference for monocotyledonous plants. Int. J. Syst. Evol. Microbiol. 2013, 63, 2520–2525. [Google Scholar] [CrossRef] [Green Version]
- Waleron, M.; Misztak, A.; Waleron, M.; Franczuk, M.; Jońca, J.; Wielgomas, B.; Mikiciński, A.; Popović, T.; Waleron, K. Pectobacterium zantedeschiae sp. nov. a new species of a soft rot pathogen isolated from Calla lily (Zantedeschia spp.). Syst. Appl. Microbiol. 2019, 42, 275–283. [Google Scholar] [CrossRef]
- Koh, Y.J.; Kim, G.H.; Lee, Y.S.; Sohn, S.H.; Koh, H.S.; Kwon, S.; Heu, S.; Jung, J.S. Pectobacterium carotovorum subsp. actinidiae subsp. nov., a new bacterial pathogen causing canker-like symptoms in yellow kiwifruit, Actinidia chinensis. N. Z. J. Crop Hortic. Sci. 2012, 40, 269–279. [Google Scholar] [CrossRef] [Green Version]
- Oulghazi, S.; Cigna, J.; Lau, Y.Y.; Moumni, M.; Chan, K.G.; Faure, D. Transfer of the waterfall source isolate Pectobacterium carotovorum M022 to Pectobacterium fontis sp. nov., a deep-branching species within the genus Pectobacterium. Int. J. Syst. Evol. Microbiol. 2019, 69, 470–475. [Google Scholar] [CrossRef]
- Pédron, J.; Bertrand, C.; Taghouti, G.; Portier, P.; Barny, M.-A. Pectobacterium aquaticum sp. nov., isolated from waterways. Int. J. Syst. Evol. Microbiol. 2019, 69, 745–751. [Google Scholar] [CrossRef]
- Toth, I.K.; van der Wolf, J.M.; Saddler, G.; Lojkowska, E.; Hélias, V.; Pirhonen, M.; Tsror (Lahkim), L.; Elphinstone, J.G. Dickeya species: An emerging problem for potato production in Europe. Plant Pathol. 2011, 60, 385–399. [Google Scholar] [CrossRef]
- Tsror, L.; Erlich, O.; Lebiush, S.; Hazanovsky, M.; Zig, U.; Slawiak, M.; Grabe, G.; Van Der Wolf, J.M.; Van De Haar, J.J. Assessment of recent outbreaks of Dickeya sp. (syn. Erwinia chrysanthemi) slow wilt in potato crops in Israel. Eur. J. Plant Pathol. 2009, 123, 311–320. [Google Scholar]
- Van der Wolf, J.M.; Nijhuis, E.H.; Kowalewska, M.J.; Saddler, G.S.; Parkinson, N.; Elphinstone, J.G.; Pritchard, L.; Toth, I.K.; Lojkowska, E.; Potrykus, M.; et al. Dickeya solani sp. nov., a pectinolytic plant-pathogenic bacterium isolated from potato (Solanum tuberosum). Int. J. Syst. Evol. Microbiol. 2014, 64, 768–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ignatov, A.; Karlov, A.; Dzhalilov, F. Spreading of the blackleg of potatoes in Russia caused by bacteria of Dickeya genus. Zaschita i Karantin Rastenij 2014, 11, 41–43. [Google Scholar]
- Ozturk, M.; Aksoy, H.M. First report of Dickeya solani associated with potato blackleg and soft rot in Turkey. J. Plant Pathol. 2017, 99, 298. [Google Scholar]
- Tsror [Lahkim], L.; Erlich, O.; Lebiush, S.; van der Wolf, J.; Czajkowski, R.; Mozes, G.; Sikharulidze, Z.; Ben Daniel, B. First report of potato blackleg caused by a biovar 3 Dickeya sp. in Georgia. New Dis. Rep. 2011, 23, 1. [Google Scholar] [CrossRef] [Green Version]
- Cardoza, Y.F.; Duarte, V.; Lopes, C.A. First report of blackleg of potato caused by Dickeya solani in Brazil. Plant Dis. 2017, 101, 243. [Google Scholar] [CrossRef]
- Nykyri, J.; Niemi, O.; Koskinen, P.; Nokso-Koivisto, J.; Pasanen, M.; Broberg, M.; Plyusnin, I.; Törönen, P.; Holm, L.; Pirhonen, M.; et al. Revised Phylogeny and Novel Horizontally Acquired Virulence Determinants of the Model Soft Rot Phytopathogen Pectobacterium wasabiae SCC3193. PLoS Pathog. 2012, 8, e1003013. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Fan, Q.; Loria, R. A re-evaluation of the taxonomy of phytopathogenic genera Dickeya and Pectobacterium using whole-genome sequencing data. Syst. Appl. Microbiol. 2016, 39, 252–259. [Google Scholar] [CrossRef]
- Brady, C.L.; Cleenwerck, I.; Denman, S.; Venter, S.N.; Rodriguez-Palenzuela, P.; Coutinho, T.A.; De Vos, P. Proposal to reclassify Brenneria quercina (Hildebrand and Schroth 1967) Hauben et al. 1999 into a new genus, Lonsdalea gen. nov., as Lonsdalea quercina comb. nov., descriptions of Lonsdalea quercina subsp. quercina comb. nov., Lonsdalea quercina subsp. ib. Int. J. Syst. Evol. Microbiol. 2012, 62, 1592–1602. [Google Scholar] [CrossRef]
- Tian, Y.; Zhao, Y.; Yuan, X.; Yi, J.; Fan, J.; Xu, Z.; Hu, B.; De Boer, S.H.; Li, X. Dickeya fangzhongdai sp. nov., a plantpathogenic bacterium isolated from pear trees (Pyrus pyrifolia). Int. J. Syst. Evol. Microbiol. 2016, 66, 2831–2835. [Google Scholar] [CrossRef] [PubMed]
- Hugouvieux-Cotte-Pattat, N.; Jacot-Des-Combes, C.; Briolay, J. Dickeya lacustris sp. nov., a water-living pectinolytic bacterium isolated from lakes in France. Int. J. Syst. Evol. Microbiol. 2019, 69, 721–726. [Google Scholar] [CrossRef] [PubMed]
- Oulghazi, S.; Pédron, J.; Cigna, J.; Lau, Y.Y.; Moumni, M.; Van Gijsegem, F.; Chan, K.-G.; Faure, D. Dickeya undicola sp. nov., a novel species for pectinolytic isolates from surface waters in Europe and Asia. Int. J. Syst. Evol. Microbiol. 2019, 69, 2440–2444. [Google Scholar] [CrossRef] [PubMed]
- Parkinson, N.; DeVos, P.; Pirhonen, M.; Elphinstone, J. Dickeya aquatica sp. nov., isolated from waterways. Int. J. Syst. Evol. Microbiol. 2014, 64, 2264–2266. [Google Scholar] [CrossRef] [PubMed]
- Samson, R.; Legendre, J.; Richard, C.; Fischer-Le Saux, M.; Achouak, W.; Gardan, L. Transfer of Pectobacterium chrysanthemi (Burkholder, 1953) Brenner et al. 1973 and Brenneria paradisiaca to the genus Dickeya gen. nov. as Dickeya chrysanthemi comb. nov. and Dickeya paradisiaca comb. nov. and delineation of four novel species, Dickeya da. Int. J. Syst. Evol. Microbiol. 2005, 55, 1415–1427. [Google Scholar] [CrossRef]
- Golanowska, M.; Galardini, M.; Bazzicalupo, M.; Hugouvieux-Cotte-Pattat, N.; Mengoni, A.; Potrykus, M.; Slawiak, M.; Lojkowska, E. Draft Genome Sequence of a Highly Virulent Strain of the Plant Pathogen Dickeya solani, IFB0099. Genome Announc. 2015, 3, e00109-15. [Google Scholar] [CrossRef] [Green Version]
- Hanage, W.P.; Spratt, B.G.; Turner, K.M.E.; Fraser, C. Modelling bacterial speciation. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 2039–2044. [Google Scholar] [CrossRef] [Green Version]
- Cigna, J.; Dewaegeneire, P.; Beury, A.; Gobert, V.; Faure, D. A gapA PCR-sequencing Assay for Identifying the Dickeya and Pectobacterium Potato Pathogens. Plant Dis. 2017, 101, 1278–1282. [Google Scholar] [CrossRef] [Green Version]
- Goris, J.; Konstantinidis, K.T.; Klappenbach, J.A.; Coenye, T.; Vandamme, P.; Tiedje, J.M. DNA–DNA hybridization values and their relationship to whole-genome sequence similarities. Int. J. Syst. Evol. Microbiol. 2007, 57, 81–91. [Google Scholar] [CrossRef] [Green Version]
- Raoul des Essarts, Y.; Pédron, J.; Blin, P.; Van Dijk, E.; Faure, D.; Van Gijsegem, F. Common and distinctive adaptive traits expressed in Dickeya dianthicola and Dickeya solani pathogens when exploiting potato plant host. Environ. Microbiol. 2019, 21, 1004–1018. [Google Scholar] [CrossRef]
- Anajjar, B.; Aitmhand, R.; Timinouni, M.; Ennaji, M.M. Caractérisation par PCR de deux souches d’Erwinia carotovora isolées de la rhizosphère de la pomme de terre dans la région du grand Casablanca au Maroc. EPPO Bull. 2007, 37, 175–180. [Google Scholar] [CrossRef]
- Faquihi, H.; Terta, M.; Amdan, M.; Achbani, E.H.; Ennaji, M.M.; Ait Mhand, R. Phenotypic and genotypic diversity of Pectobacterium carotovorum subsp carotovorum causing soft rot disease of potatoes in Morocco. Eur. J. Plant Pathol. 2015, 143, 801–811. [Google Scholar] [CrossRef]
- Terta, M.; El Karkouri, A.; Ait M’Hand, R.; Achbani, E.; Barakate, M.; Amdan, M.; Annajar, B.; El Hassouni, M.; Val, F.; Bouteau, F.; et al. Occurrence of Pectobacterium carotovorum strains isolated from potato soft rot in Morocco. Cell. Mol. Biol. 2010, 56, OL1324–OL1333. [Google Scholar] [PubMed]
- Terta, M.; Azelmat, S.; M’Hand, R.A.; Achbani, E.H.; Barakate, M.; Bouteau, F.; Ennaji, M.M. Molecular typing of Pectobacterium carotovorum isolated from potato tuber soft rot in Morocco. Ann. Microbiol. 2012, 62, 1411–1417. [Google Scholar] [CrossRef]
- Oulghazi, S.; Khayi, S.; Lafkih, N.; Massaoudi, Y.; El Karkouri, A.; El Hassouni, M.; Faure, D.; Moumni, M. First report of Dickeya dianthicola causing blackleg on potato in Morocco. Plant Dis. 2017, 101, 1671–1672. [Google Scholar] [CrossRef]
- Hélias, V.; Hamon, P.; Huchet, E.; Wolf, J.V.D.; Andrivon, D. Two new effective semiselective crystal violet pectate media for isolation of Pectobacterium and Dickeya. Plant Pathol. 2012, 61, 339–345. [Google Scholar] [CrossRef]
- Darrasse, A.; Priou, S.; Kotoujansky, A.; Bertheau, Y. PCR and restriction fragment length polymorphism of a pel gene as a tool to identify Erwinia carotovora in relation to potato diseases. Appl. Environ. Microbiol. 1994, 60, 1437–1443. [Google Scholar] [CrossRef] [Green Version]
- Nassar, A.; Darrasse, A.; Lemattre, M.; Kotoujansky, A.; Dervin, C.; Vedel, R.; Bertheau, Y. Characterization of Erwinia chrysanthemi by pectinolytic isozyme polymorphism and restriction fragment length polymorphism analysis of PCR- amplified fragments of pel genes. Appl. Environ. Microbiol. 1996, 62, 2228–2235. [Google Scholar] [CrossRef] [Green Version]
- Grant, J.R.; Stothard, P. The CGView Server: A comparative genomics tool for circular genomes. Nucleic Acids Res. 2008, 36, W181–W184. [Google Scholar] [CrossRef]
- Darling, A.C.E.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef] [Green Version]
- Grissa, I.; Vergnaud, G.; Pourcel, C. CRISPRFinder: A web tool to identify clustered regularly interspaced short palindromic repeats. Nucleic Acids Res. 2007, 35, W52–W57. [Google Scholar] [CrossRef] [Green Version]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khayi, S.; Blin, P.; Pédron, J.; Chong, T.M.; Chan, K.G.; Moumni, M.; Hélias, V.; Van Gijsegem, F.; Faure, D. Population genomics reveals additive and replacing horizontal gene transfers in the emerging pathogen Dickeya solani. BMC Genom. 2015, 16, 788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horvath, P.; Barrangou, R. CRISPR/Cas, the immune system of bacteria and archaea. Science 2010, 327, 167–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reverchon, S.; Nasser, W. Dickeya ecology, environment sensing and regulation of virulence programme. Environ. Microbiol. Rep. 2013, 5, 622–636. [Google Scholar]
- Choi, O.; Kim, J. Pectobacterium carotovorum subsp. brasiliense Causing Soft Rot on Paprika in Korea. J. Phytopathol. 2013, 161, 125–127. [Google Scholar] [CrossRef]
- De Boer, S.H.; Li, X.; Ward, L.J. Pectobacterium spp. Associated with Bacterial Stem Rot Syndrome of Potato in Canada. Phytopathology 2012, 102, 937–947. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Chai, A.; Shi, Y.; Xie, X.; Ma, Z.; Li, B. Emergence of Bacterial Soft Rot in Cucumber Caused by Pectobacterium carotovorum subsp. brasiliense in China. Plant Dis. 2017, 101, 279–287. [Google Scholar] [CrossRef] [Green Version]
- Nunes Leite, L.; de Haan, E.G.; Krijger, M.; Kastelein, P.; van der Zouwen, P.S.; van den Bovenkamp, G.W.; Tebaldi, N.D.; van der Wolf, J.M. First report of potato blackleg caused by Pectobacterium carotovorum subsp. brasiliensis in the Netherlands. New Dis. Rep. 2014, 29, 24. [Google Scholar] [CrossRef] [Green Version]
- Panda, P.; Fiers, M.A.W.J.; Armstrong, K.; Pitman, A.R. First report of blackleg and soft rot of potato caused by Pectobacterium carotovorum subsp. brasiliensis in New Zealand. New Dis. Rep. 2012, 26, 15. [Google Scholar] [CrossRef] [Green Version]
- Van der Merwe, J.J.; Coutinho, T.A.; Korsten, L.; van der Waals, J.E. Pectobacterium carotovorum subsp. brasiliensis causing blackleg on potatoes in South Africa. Eur. J. Plant Pathol. 2010, 126, 175–185. [Google Scholar] [CrossRef] [Green Version]
- Li, X.S.; Yuan, K.X.; Cullis, J.; Lévesque, C.A.; Chen, W.; Lewis, C.T.; De Boer, S.H. Draft Genome Sequences for Canadian Isolates of Pectobacterium carotovorum subsp. brasiliense with Weak Virulence on Potato. Genome Announc. 2015, 3, e00240-15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niemi, O.; Laine, P.; Koskinen, P.; Pasanen, M.; Pennanen, V.; Harjunpää, H.; Nykyri, J.; Holm, L.; Paulin, L.; Auvinen, P.; et al. Genome sequence of the model plant pathogen Pectobacterium carotovorum SCC1. Stand. Genom. Sci. 2017, 12, 87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Naas, H.; Sebaihia, M.; Orfei, B.; Rezzonico, F.; Buonaurio, R.; Moretti, C. Pectobacterium carotovorum subsp. brasiliense and Pectobacterium carotovorum subsp. carotovorum as causal agents of potato soft rot in Algeria. Eur. J. Plant Pathol. 2018, 151, 1027–1034. [Google Scholar] [CrossRef]
- Ozturk, M.; Aksoy, H.M.; Potrykus, M.; Lojkowska, E. Correction to: Genotypic and phenotypic variability of Pectobacterium strains causing blackleg and soft rot on potato in Turkey. Eur. J. Plant Pathol. 2018, 152, 1. [Google Scholar] [CrossRef]
- Van der Wolf, J.; de Haan, E.G.; Kastelein, P.; Krijger, M.; de Haas, B.H.; Velvis, H.; Mendes, O.; Kooman-Gersmann, M.; van der Zouwen, P.S. Virulence of Pectobacterium carotovorum subsp. brasiliense on potato compared with that of other Pectobacterium and Dickeya species under climatic conditions prevailing in the Netherlands. Plant Pathol. 2017, 66, 571–583. [Google Scholar] [CrossRef]
- Charkowski, A.O. The Changing Face of Bacterial Soft-Rot Diseases. Annu. Rev. Phytopathol. 2018, 56, 16éà. [Google Scholar] [CrossRef]
- Schöner, T.A.; Gassel, S.; Osawa, A.; Tobias, N.J.; Okuno, Y.; Sakakibara, Y.; Shindo, K.; Sandmann, G.; Bode, H.B. Aryl Polyenes, a Highly Abundant Class of Bacterial Natural Products, Are Functionally Related to Antioxidative Carotenoids. ChemBioChem 2016, 17, 247–253. [Google Scholar] [CrossRef]
- Alič, Š.; Pédron, J.; Dreo, T.; Van Gijsegem, F. Genomic characterisation of the new Dickeya fangzhongdai species regrouping plant pathogens and environmental isolates. BMC Genom. 2019, 20, 34. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism | Accession Number | Genome Size | N50 (pb) | Number of Contigs | Coverage | Number of CDS | Number of tRNAs |
|---|---|---|---|---|---|---|---|
| Pectobacterium polaris S4.16.03.2B | QZDF00000000 | 4,862,009 | 155,865 | 65 | 41 | 4355 | 54 |
| Pectobacterium brasiliense S1.16.01.3k | QZDG00000000 | 4,946,598 | 146,844 | 74 | 410 | 4337 | 36 |
| Pectobacterium brasiliense S1.15.11.2D | QZDH00000000 | 4,818,836 | 99,392 | 91 | 420 | 4206 | 35 |
| Pectobacterium brasiliense S4.16.03.1C | QZDI00000000 | 4,944,722 | 139,665 | 74 | 467 | 4336 | 37 |
| Pectobacterium carotovorum S1-A16 | QZDJ00000000 | 4,835,633 | 255,206 | 37 | 55 | 4261 | 63 |
| Pectobacterium versatile S4.16.03.3I | QZDK00000000 | 4,854,084 | 8262 | 108 | 246 | 4247 | 34 |
| Pectobacterium versatile S4.16.03.3K | QZDL00000000 | 4,870,940 | 90,195 | 106 | 237 | 4262 | 34 |
| Pectobacterium versatile S4.16.03.3F | QZDM00000000 | 4,852,595 | 89,731 | 114 | 143 | 4244 | 40 |
| Dickeya dianthicola S4.16.03.P2.4 | QZDN00000000 | 4,865,147 | 92,028 | 101 | 415 | 4238 | 37 |
| Dickeya dianthicola LAR.16.03.LID | QZDO00000000 | 4,863,939 | 71,707 | 108 | 344 | 4238 | 37 |
| ANI Values | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Strains | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 |
| 1-Pp NIBIO 1006T | 96.8 | 92.9 | 92.9 | 93.5 | 93.7 | 93.5 | 93.7 | 93.4 | 93.4 | 93.5 | 93.5 | 92.2 | 79.3 | 79 | 79.1 | |
| 2-Pp S4.16.03.2B | 73.30 | 93 | 92.9 | 93.5 | 93.7 | 93.5 | 93.7 | 93.4 | 93.5 | 93.5 | 93.4 | 92.2 | 78.9 | 79 | 79 | |
| 3-Pc ICMP5702T | 52.3 | 52.10 | 97.2 | 92.6 | 92.9 | 92.8 | 92.8 | 95.1 | 95.2 | 95.2 | 95.2 | 94.8 | 78.9 | 78.9 | 78.9 | |
| 4-Pc S1-A16 | 52.2 | 51.90 | 76.30 | 92.6 | 92.9 | 92.7 | 92.9 | 95.3 | 95.3 | 95.3 | 95.3 | 94.8 | 78.8 | 79.6 | 79.6 | |
| 5-Pb LMG 21371T | 54.4 | 54.5 | 50.9 | 50.9 | 96.1 | 95.9 | 96.1 | 92.2 | 92.2 | 92.2 | 92.3 | 93.00 | 78.6 | 79.5 | 79.5 | |
| 6-Pb S4.16.03.1C | 56 | 55.7 | 52 | 51.7 | 68.5 | 96.3 | 100 | 92.3 | 91.2 | 91.1 | 91.2 | 91.9 | 78.7 | 79.6 | 79.6 | |
| 7-Pb S1.15.11.2D | 54.7 | 54.2 | 51.3 | 51 | 67.1 | 69.4 | 96.3 | 92.3 | 92.2 | 92.3 | 92.3 | 91.8 | 79.1 | 79.1 | 79.1 | |
| 8-Pb S1.16.01.3K | 56 | 55.7 | 76 | 51.7 | 68.5 | 100 | 71.4 | 92.4 | 92.4 | 92.4 | 92.3 | 91.9 | 79.6 | 79.6 | 79.7 | |
| 9-Pv SCC1 | 54.3 | 54.3 | 63.5 | 63.9 | 48.9 | 49.8 | 49.5 | 49.3 | 99.5 | 99.5 | 99.5 | 94.7 | 79.2 | 79.1 | 79.3 | |
| 10-Pv S4.16.03.3F | 54.4 | 54.3 | 63.9 | 64.3 | 49.1 | 49.9 | 49.5 | 49.9 | 96.6 | 100 | 100 | 94.7 | 79.2 | 79.2 | 79.2 | |
| 11-Pv S4.16.03.3k | 54.4 | 54.3 | 63.8 | 64.2 | 49.2 | 49.9 | 49.5 | 49.9 | 96.6 | 99.3 | 100 | 94.7 | 79.3 | 79.3 | 79.4 | |
| 12-Pv S4.16.03.3I | 54.4 | 54.3 | 63.8 | 64.2 | 49.1 | 49.9 | 49.5 | 49.8 | 96.5 | 100 | 99.3 | 94.7 | 79.3 | 79.3 | 79.3 | |
| 13-Po BCS7 | 49.2 | 49 | 61.3 | 60.6 | 47.4 | 47.8 | 47.6 | 47.8 | 60.4 | 60.7 | 60.6 | 60.6 | 79.1 | 79.1 | 79.1 | |
| 14-Ddi NCPPB 453T | 21.1 | 20.7 | 20.5 | 20.7 | 20.9 | 21 | 20.6 | 21 | 21.2 | 21 | 21 | 21 | 21.1 | 99.5 | 99.5 | |
| 15-Ddi S4.16.03.P2.4 | 20.8 | 20.6 | 20.4 | 20.9 | 21.1 | 20.9 | 20.7 | 20.9 | 21 | 21 | 21 | 21 | 20.8 | 95.6 | 100 | |
| 16-Ddi LAR.16.03.LID | 20.8 | 20.6 | 20.4 | 21 | 21.1 | 20.9 | 20.6 | 20.9 | 21 | 21 | 21 | 21 | 20.8 | 95.6 | 100 | |
| is-DDH | ||||||||||||||||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oulghazi, S.; Moumni, M.; Khayi, S.; Robic, K.; Sarfraz, S.; Lopez-Roques, C.; Vandecasteele, C.; Faure, D. Diversity of Pectobacteriaceae Species in Potato Growing Regions in Northern Morocco. Microorganisms 2020, 8, 895. https://doi.org/10.3390/microorganisms8060895
Oulghazi S, Moumni M, Khayi S, Robic K, Sarfraz S, Lopez-Roques C, Vandecasteele C, Faure D. Diversity of Pectobacteriaceae Species in Potato Growing Regions in Northern Morocco. Microorganisms. 2020; 8(6):895. https://doi.org/10.3390/microorganisms8060895
Chicago/Turabian StyleOulghazi, Saïd, Mohieddine Moumni, Slimane Khayi, Kévin Robic, Sohaib Sarfraz, Céline Lopez-Roques, Céline Vandecasteele, and Denis Faure. 2020. "Diversity of Pectobacteriaceae Species in Potato Growing Regions in Northern Morocco" Microorganisms 8, no. 6: 895. https://doi.org/10.3390/microorganisms8060895