
Lactiplantibacillus plantarum Induces Apoptosis in Melanoma and Breast Cancer Cells
 , and
, and 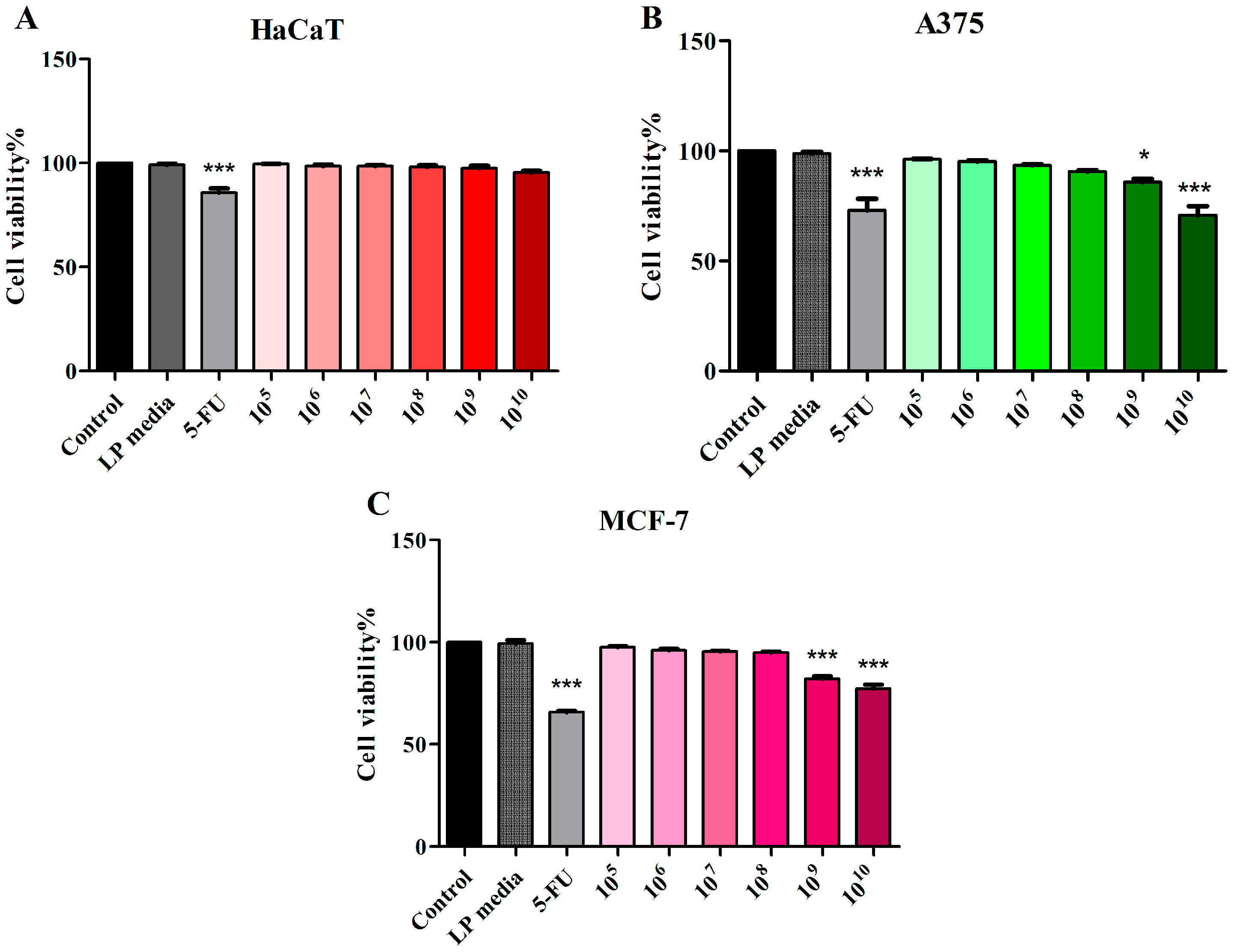{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacteria and Cell Lines
2.2. Cellular Viability Assessment
2.3. Chromatin Condensation
2.4. Annexin-V-FITC/Propidium Iodide (PI) Staining
2.5. BAX and Bcl-2 Detection
2.6. Western Blot
2.7. Statistical Analysis
3. Results
3.1. Assessment of LP Cytotoxic Activity
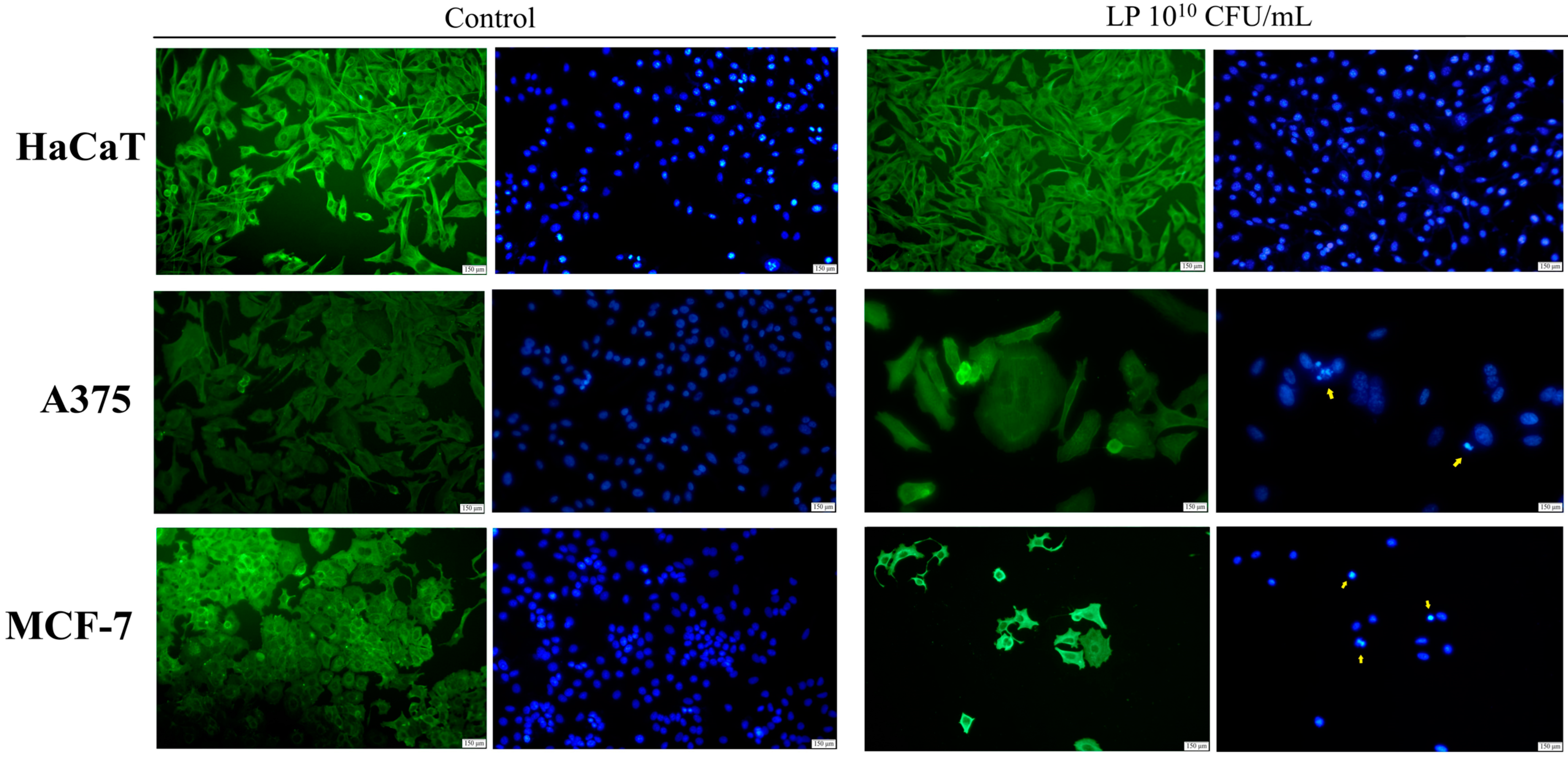
3.2. Evaluation of Apoptotic Features by Immunofluorescent Staining
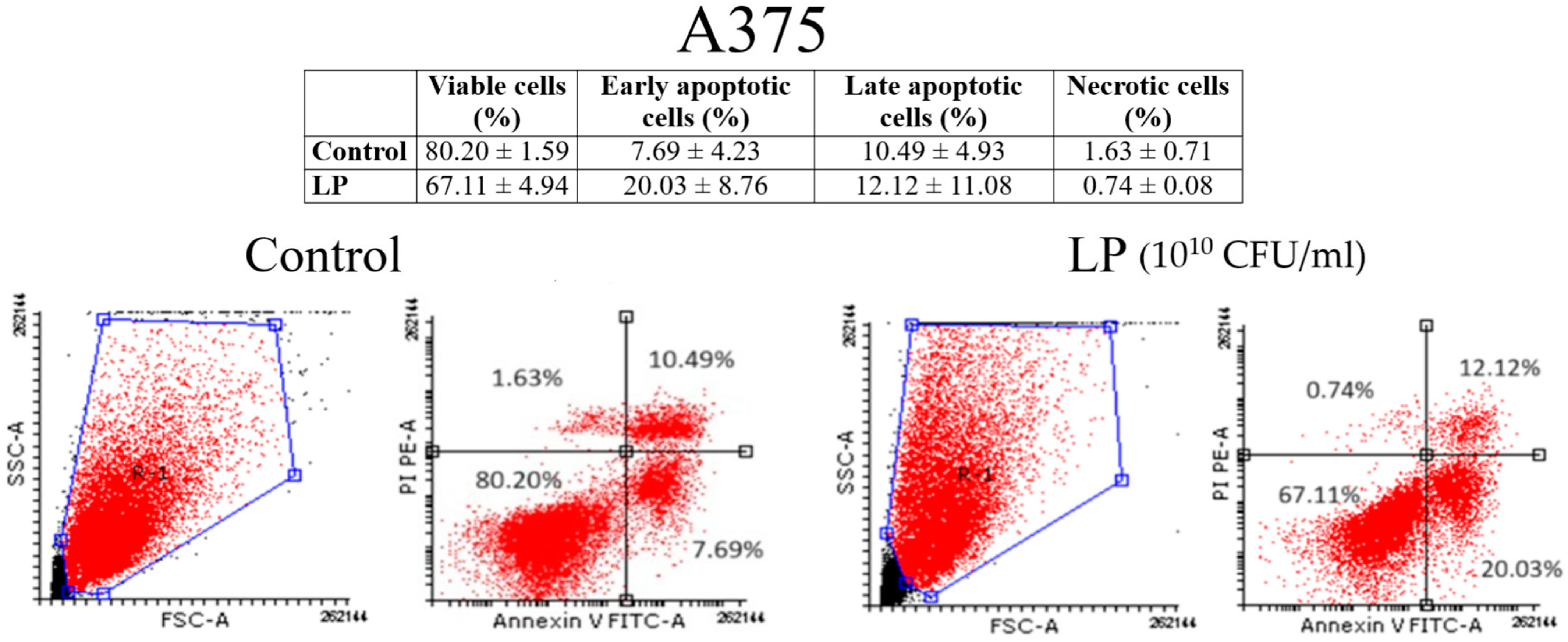
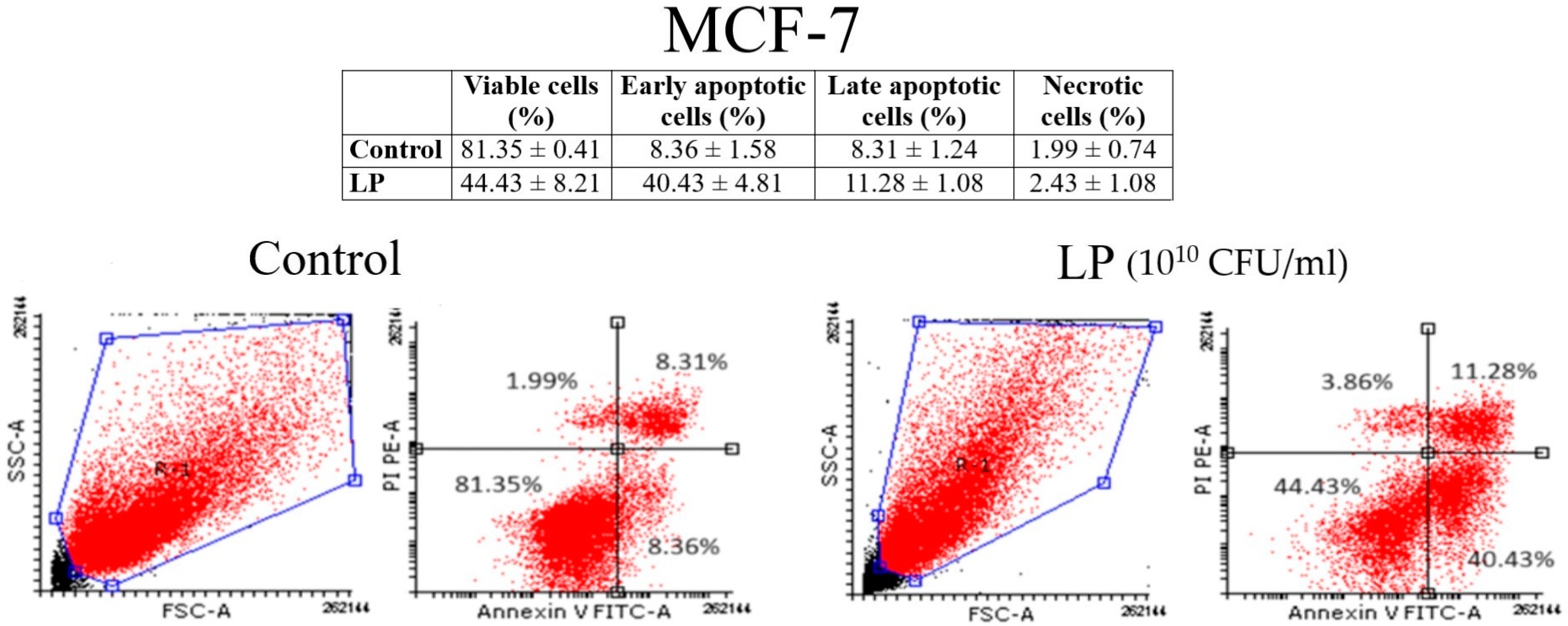
3.3. Annexin V/PI Analysis
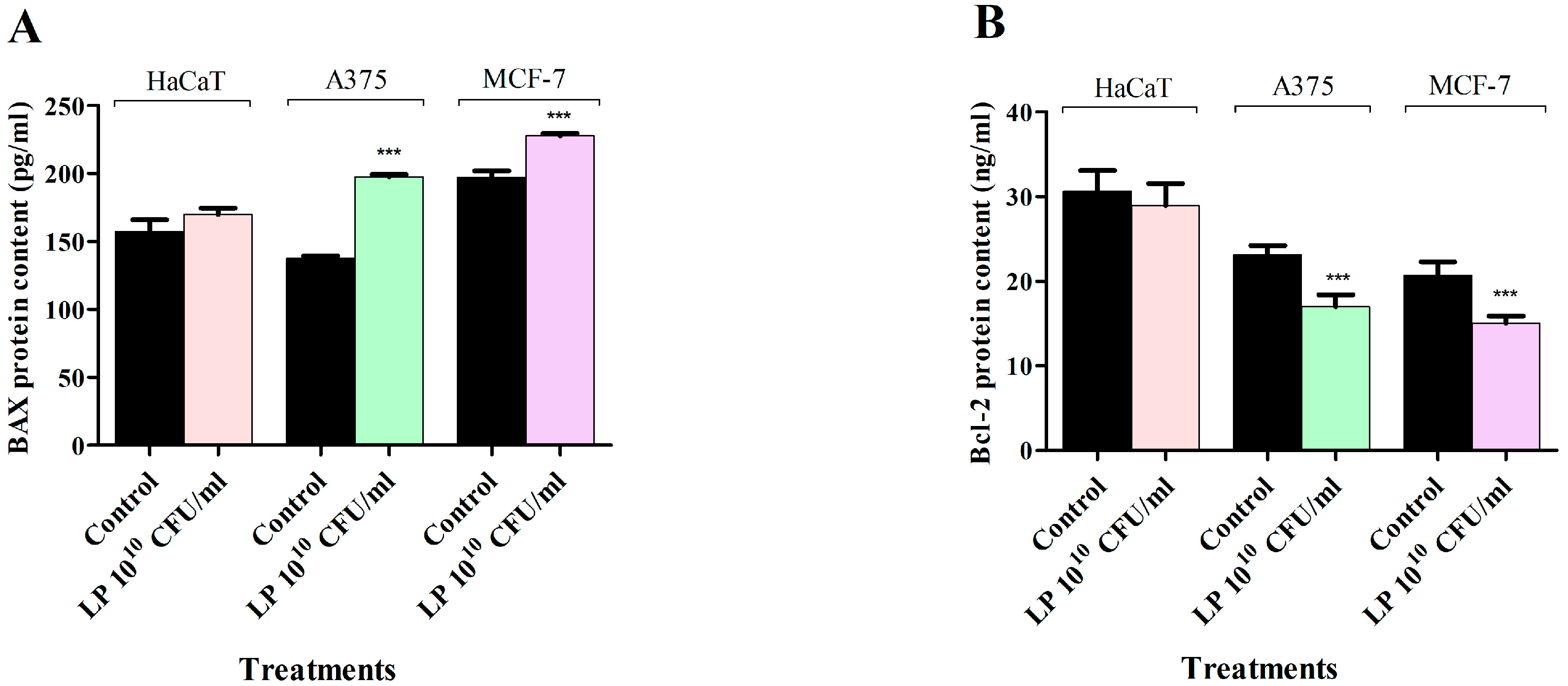
3.4. Apoptosis Assay—LP Effect on BAX and Bcl-2 Protein Levels
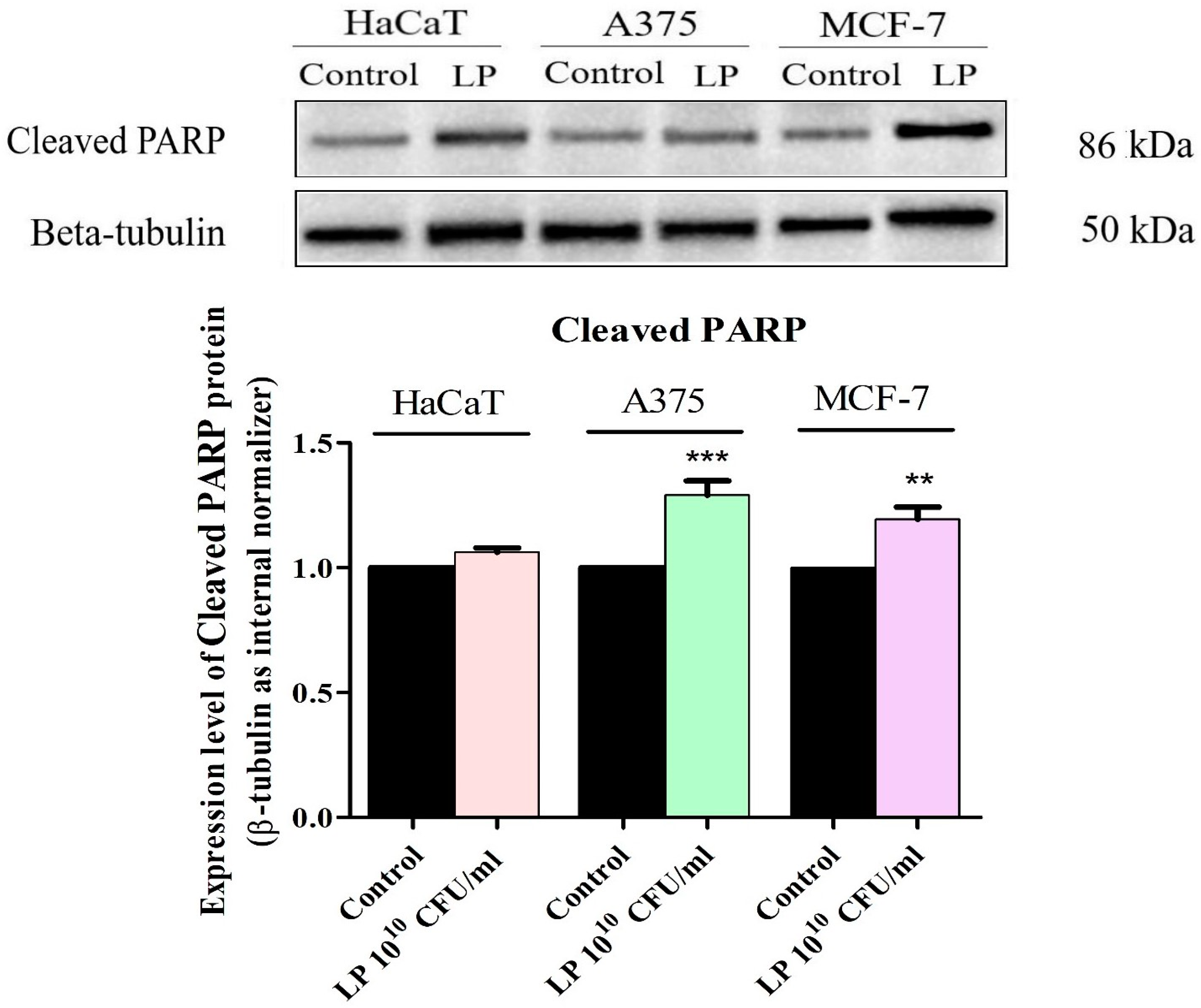
3.5. Western Blot Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Śliżewska, K.; Markowiak-Kopeć, P.; Śliżewska, W. The Role of Probiotics in Cancer Prevention. Cancers 2020, 13, 20. [Google Scholar] [CrossRef]
- Górska, A.; Przystupski, D.; Niemczura, M.J.; Kulbacka, J. Probiotic Bacteria: A Promising Tool in Cancer Prevention and Therapy. Curr. Microbiol. 2019, 76, 939–949. [Google Scholar] [CrossRef]
- Bedada, T.L.; Feto, T.K.; Awoke, K.S.; Garedew, A.D.; Yifat, F.T.; Birri, D.J. Probiotics for cancer alternative prevention and treatment. Biomed. Pharmacother. 2020, 129, 110409. [Google Scholar] [CrossRef]
- Widyastuti, Y.; Febrisiantosa, A.; Tidona, F. Health-Promoting Properties of Lactobacilli in Fermented Dairy Products. Front. Microbiol. 2021, 12, 673890. [Google Scholar] [CrossRef] [PubMed]
- Garbacz, K. Anticancer activity of lactic acid bacteria. Semin. Cancer Biol. 2022, 86, 356–366. [Google Scholar] [CrossRef]
- Chang, J.-H.; Shim, Y.Y.; Cha, S.-K.; Reaney, M.J.T.; Chee, K.M. Effect of Lactobacillus acidophilus KFRI342 on the development of chemically induced precancerous growths in the rat colon. J. Med. Microbiol. 2012, 61, 361–368. [Google Scholar] [CrossRef]
- Ghorbani, E.; Avan, A.; Ryzhikov, M.; Ferns, G.; Khazaei, M.; Soleimanpour, S. Role of lactobacillus strains in the management of colorectal cancer: An overview of recent advances. Nutrition 2022, 103–104, 111828. [Google Scholar] [CrossRef] [PubMed]
- Hizay, A.; Dag, K.; Oz, N.; Comak-Gocer, E.M.; Ozbey-Unlu, O.; Ucak, M.; Keles-Celik, N. Lactobacillus acidophilus regulates abnormal serotonin availability in experimental ulcerative colitis. Anaerobe 2023, 80, 102710. [Google Scholar] [CrossRef]
- Singh, S.; Singh, M.; Gaur, S. Probiotics as multifaceted oral vaccines against colon cancer: A review. Front. Immunol. 2022, 13, 1002674. [Google Scholar] [CrossRef]
- Mendoza, L. Potential effect of probiotics in the treatment of breast cancer. Oncol. Rev. 2019, 13, 422. [Google Scholar] [CrossRef]
- Kim, H.R.; Kim, H.; Jung, B.J.; You, G.E.; Jang, S.; Chung, D.K. Lipoteichoic acid isolated from Lactobacillus plantarum inhibits melanogenesis in B16F10 mouse melanoma cells. Mol. Cells 2015, 38, 163–170. [Google Scholar] [CrossRef]
- Bender, M.J.; McPherson, A.C.; Phelps, C.M.; Pandey, S.P.; Laughlin, C.R.; Shapira, J.H.; Medina Sanchez, L.; Rana, M.; Richie, T.G.; Mims, T.S.; et al. Dietary tryptophan metabolite released by intratumoral Lactobacillus reuteri facilitates immune checkpoint inhibitor treatment. Cell 2023, 186, 1846.e26–1862.e26. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Hu, M.; Feng, X.; XiaoLi, W.; Dong, D.; Wang, W. Preventive effect of Lactobacillus reuteri on melanoma. Biomed. Pharmacother. 2020, 126, 109929. [Google Scholar] [CrossRef] [PubMed]
- Fuerst, M.L. Gut Microbiome Affects Response to Immunotherapy. Oncol. Times 2019, 41, 32. [Google Scholar] [CrossRef]
- Gopalakrishnan, V.; Spencer, C.N.; Nezi, L.; Reuben, A.; Andrews, M.C.; Karpinets, T.V.; Prieto, P.A.; Vicente, D.; Hoffman, K.; Wei, S.C.; et al. Gut microbiome modulates response to anti–PD-1 immunotherapy in melanoma patients. Science 2018, 359, 97–103. [Google Scholar] [CrossRef] [PubMed]
- Spencer, C.N.; McQuade, J.L.; Gopalakrishnan, V.; McCulloch, J.A.; Vetizou, M.; Cogdill, A.P.; Khan, M.A.W.; Zhang, X.; White, M.G.; Peterson, C.B.; et al. Dietary fiber and probiotics influence the gut microbiome and melanoma immunotherapy response. Science 2021, 374, 1632–1640. [Google Scholar] [CrossRef]
- Majtnerova, P.; Capek, J.; Petira, F.; Handl, J.; Rousar, T. Quantitative spectrofluorometric assay detecting nuclear condensation and fragmentation in intact cells. Sci. Rep. 2021, 11, 11921. [Google Scholar] [CrossRef]
- Alfhili, M.A.; Hussein, H.A.M.; Park, Y.; Lee, M.H.; Akula, S.M. Triclosan induces apoptosis in Burkitt lymphoma-derived BJAB cells through caspase and JNK/MAPK pathways. Apoptosis 2021, 26, 96–110. [Google Scholar] [CrossRef]
- Assay Procedure. Available online: https://www.abcam.com/?gclsrc=aw.ds&gclid=Cj0KCQiAutyfBhCMARIsAMgcRJTqXuV8ejoRZWlnUJ1b5th64NWvCAUQZCJ3vXNt-0T9tgWXRutKeaMaAvwCEALw_wcB&gclsrc=aw.ds (accessed on 1 October 2023).
- Pillai-Kastoori, L.; Schutz-Geschwender, A.R.; Harford, J.A. A systematic approach to quantitative Western blot analysis. Anal. Biochem. 2020, 593, 113608. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Lee, H.; Park, K.; Shin, S. Rapid and Efficient Isolation of Exosomes by Clustering and Scattering. J. Clin. Med. 2020, 9, 650. [Google Scholar] [CrossRef]
- World Health Organization. Cancer—Fact Sheets. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 1 October 2023).
- Saginala, K.; Barsouk, A.; Aluru, J.S.; Rawla, P.; Barsouk, A. Epidemiology of Melanoma. Med. Sci. 2021, 9, 63. [Google Scholar] [CrossRef]
- Sankarapandian, V.; Venmathi Maran, B.A.; Rajendran, R.L.; Jogalekar, M.P.; Gurunagarajan, S.; Krishnamoorthy, R.; Gangadaran, P.; Ahn, B.-C. An Update on the Effectiveness of Probiotics in the Prevention and Treatment of Cancer. Life 2022, 12, 59. [Google Scholar] [CrossRef]
- Van Holm, W.; Lauwens, K.; De Wever, P.; Schuermans, A.; Zayed, N.; Pamuk, F.; Saghi, M.; Fardim, P.; Bernaerts, K.; Boon, N.; et al. Probiotics for oral health: Do they deliver what they promise? Front. Microbiol. 2023, 14, 1219692. [Google Scholar] [CrossRef]
- Molska, M.; Reguła, J. Potential Mechanisms of Probiotics Action in the Prevention and Treatment of Colorectal Cancer. Nutrients 2019, 11, 2453. [Google Scholar] [CrossRef]
- Salemi, R.; Vivarelli, S.; Ricci, D.; Scillato, M.; Santagati, M.; Gattuso, G.; Falzone, L.; Libra, M. Lactobacillus rhamnosus GG cell-free supernatant as a novel anti-cancer adjuvant. J. Transl. Med. 2023, 21, 195. [Google Scholar] [CrossRef]
- Park, J.; Kwon, M.; Lee, J.; Park, S.; Seo, J.; Roh, S. Anti-Cancer Effects of Lactobacillus plantarum L-14 Cell-Free Extract on Human Malignant Melanoma A375 Cells. Molecules 2020, 25, 3895. [Google Scholar] [CrossRef]
- Aintablian, A.; Jaber, D.F.; Jallad, M.A.; Abdelnoor, A.M. The Effect of Lactobacillus Plantarum and Bacterial Peptidoglycan on the Growth of Mouse Tumors in vivo and in vitro. Am. J. Immunol. 2017, 13, 201–208. [Google Scholar] [CrossRef]
- Boonlert, B.; Fongcom, A.; Virarach, N. The Safety of Lactobacillus Plantarum Extract Used as a Possible Adjuvant Treatment of Breast Cancer. Thai J. Surg. 2022, 43, 105–112. [Google Scholar]
- Chuah, L.-O.; Foo, H.L.; Loh, T.C.; Alitheen, N.B.M.; Yeap, S.K.; Abdul Mutalib, N.E.; Abdul Rahim, R.; Yusoff, K. Postbiotic metabolites produced by Lactobacillus plantarum strains exert selective cytotoxicity effects on cancer cells. BMC Complement. Altern. Med. 2019, 19, 114. [Google Scholar] [CrossRef]
- Kassayová, M.; Bobrov, N.; Strojný, L.; Kisková, T.; Mikeš, J.; Demečková, V.; Orendáš, P.; Bojková, B.; Péč, M.; Kubatka, P.; et al. Preventive effects of probiotic bacteria Lactobacillus plantarum and dietary fiber in chemically-induced mammary carcinogenesis. Anticancer Res. 2014, 34, 4969–4975. [Google Scholar]
- Malik, S.S.; Saeed, A.; Baig, M.; Asif, N.; Masood, N.; Yasmin, A. Anticarcinogenecity of microbiota and probiotics in breast cancer. Int. J. Food Prop. 2018, 21, 655–666. [Google Scholar] [CrossRef]
- Urbaniak, C.; Cummins, J.; Brackstone, M.; Macklaim, J.M.; Gloor, G.B.; Baban, C.K.; Scott, L.; O’Hanlon, D.M.; Burton, J.P.; Francis, K.P.; et al. Microbiota of Human Breast Tissue. Appl. Environ. Microbiol. 2014, 80, 3007–3014. [Google Scholar] [CrossRef] [PubMed]
- Samkari, A.A.; Alsulami, M.; Bataweel, L.; Altaifi, R.; Altaifi, A.; Saleem, A.M.; Farsi, A.H.; Iskanderani, O.; Akeel, N.Y.; Malibary, N.H.; et al. Body Microbiota and Its Relationship with Benign and Malignant Breast Tumors: A Systematic Review. Cureus 2022, 14. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.; Kim, K.; Noh, J.-Y.; Jung, Y.; Bae, O.-N.; Lim, K.-M.; Chung, J.-H. Simvastatin induces the apoptosis of normal vascular smooth muscle through the disruption of actin integrity via the impairment of RhoA/Rac-1 activity. Thromb. Haemost. 2016, 116, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Płochocka, A.Z.; Ramirez Moreno, M.; Davie, A.M.; Bulgakova, N.A.; Chumakova, L. Robustness of the microtubule network self-organization in epithelia. eLife 2021, 10, e59529. [Google Scholar] [CrossRef]
- Fu, A.; Yao, B.; Dong, T.; Chen, Y.; Yao, J.; Liu, Y.; Li, H.; Bai, H.; Liu, X.; Zhang, Y.; et al. Tumor-resident intracellular microbiota promotes metastatic colonization in breast cancer. Cell 2022, 185, 1356.e26–1372.e26. [Google Scholar] [CrossRef]
- Zamberi, N.R.; Abu, N.; Mohamed, N.E.; Nordin, N.; Keong, Y.S.; Beh, B.K.; Zakaria, Z.A.B.; Nik Abdul Rahman, N.M.A.; Alitheen, N.B. The Antimetastatic and Antiangiogenesis Effects of Kefir Water on Murine Breast Cancer Cells. Integr. Cancer Ther. 2016, 15, NP53–NP66. [Google Scholar] [CrossRef] [PubMed]
- Brentnall, M.; Rodriguez-Menocal, L.; De Guevara, R.L.; Cepero, E.; Boise, L.H. Caspase-9, caspase-3 and caspase-7 have distinct roles during intrinsic apoptosis. BMC Cell Biol. 2013, 14, 32. [Google Scholar] [CrossRef]
- Karimi Ardestani, S.; Tafvizi, F.; Tajabadi Ebrahimi, M. Heat-killed probiotic bacteria induce apoptosis of HT-29 human colon adenocarcinoma cell line via the regulation of Bax/Bcl2 and caspases pathway. Hum. Exp. Toxicol. 2019, 38, 1069–1081. [Google Scholar] [CrossRef]
- Sentürk, M.; Ercan, F.; Yalcin, S. The secondary metabolites produced by Lactobacillus plantarum downregulate BCL-2 and BUFFY genes on breast cancer cell line and model organism Drosophila melanogaster: Molecular docking approach. Cancer Chemother. Pharmacol. 2020, 85, 33–45. [Google Scholar] [CrossRef]
- Kim, H.; Kim, H.S.; Park, W.J.; Chung, D.K. Inhibitory Effect of Lactobacillus plantarum Extracts on HT-29 Colon Cancer Cell Apoptosis Induced by Staphylococcus aureus and Its Alpha-Toxin. J. Microbiol. Biotechnol. 2015, 25, 1849–1855. [Google Scholar] [CrossRef] [PubMed]
- Changizi, V.; Azadbakht, O.; Ghanavati, R.; Behrouj, H.; Motevaseli, E.; Khanzadeh, P. Effect of Lactobacillus species on apoptosis-related genes BCL2, BAX, and caspase 3 in the testes of gamma-irradiated rats. Rev. Assoc. Med. Bras. 2021, 67, 1581–1585. [Google Scholar] [CrossRef] [PubMed]
- Hiraishi, N.; Kanmura, S.; Oda, K.; Arima, S.; Kumagai, K.; Mawatari, S.; Tanoue, S.; Sasaki, F.; Hashimoto, S.; Ido, A. Extract of Lactobacillus plantarum strain 06CC2 induces JNK/p38 MAPK pathway-mediated apoptosis through endoplasmic reticulum stress in Caco2 colorectal cancer cells. Biochem. Biophys. Rep. 2019, 20, 100691. [Google Scholar] [CrossRef] [PubMed]
- Jeong, S.; Kim, Y.; Park, S.; Lee, D.; Lee, J.; Hlaing, S.P.; Yoo, J.-W.; Rhee, S.H.; Im, E. Lactobacillus plantarum Metabolites Elicit Anticancer Effects by Inhibiting Autophagy-Related Responses. Molecules 2023, 28, 1890. [Google Scholar] [CrossRef]
- Wu, H.; Schiff, D.S.; Lin, Y.; Neboori, H.J.R.; Goyal, S.; Feng, Z.; Haffty, B.G. Ionizing radiation sensitizes breast cancer cells to Bcl-2 inhibitor, ABT-737, through regulating Mcl-1. Radiat. Res. 2014, 182, 618–625. [Google Scholar] [CrossRef]
- Boulares, A.H.; Yakovlev, A.G.; Ivanova, V.; Stoica, B.A.; Wang, G.; Iyer, S.; Smulson, M. Role of poly(ADP-ribose) polymerase (PARP) cleavage in apoptosis. Caspase 3-resistant PARP mutant increases rates of apoptosis in transfected cells. J. Biol. Chem. 1999, 274, 22932–22940. [Google Scholar] [CrossRef]
- Kim, B.-K.; Yoon, Y.-S.; Ryu, Y.; Chung, M.-J. Probiotic-derived p8 protein induce apoptosis via regulation of RNF152 in colorectal cancer cells. Am. J. Cancer Res. 2021, 11, 746–759. [Google Scholar]
- Madempudi, R.S.; Kalle, A.M. Antiproliferative Effects of Bacillus coagulans Unique IS2 in Colon Cancer Cells. Nutr. Cancer 2017, 69, 1062–1068. [Google Scholar] [CrossRef] [PubMed]
- Eslami-S, Z.; Majidzadeh-A, K.; Halvaei, S.; Babapirali, F.; Esmaeili, R. Microbiome and Breast Cancer: New Role for an Ancient Population. Front. Oncol. 2020, 10, 120. [Google Scholar] [CrossRef]
- Thu, M.S.; Ondee, T.; Nopsopon, T.; Farzana, I.A.K.; Fothergill, J.L.; Hirankarn, N.; Campbell, B.J.; Pongpirul, K. Effect of Probiotics in Breast Cancer: A Systematic Review and Meta-Analysis. Biology 2023, 12, 280. [Google Scholar] [CrossRef]






Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budu, O.; Mioc, A.; Soica, C.; Caruntu, F.; Milan, A.; Oprean, C.; Lighezan, D.; Rotunjanu, S.; Ivan, V.; Banciu, C. Lactiplantibacillus plantarum Induces Apoptosis in Melanoma and Breast Cancer Cells. Microorganisms 2024, 12, 182. https://doi.org/10.3390/microorganisms12010182
Budu O, Mioc A, Soica C, Caruntu F, Milan A, Oprean C, Lighezan D, Rotunjanu S, Ivan V, Banciu C. Lactiplantibacillus plantarum Induces Apoptosis in Melanoma and Breast Cancer Cells. Microorganisms. 2024; 12(1):182. https://doi.org/10.3390/microorganisms12010182
Chicago/Turabian StyleBudu, Oana, Alexandra Mioc, Codruta Soica, Florina Caruntu, Andreea Milan, Camelia Oprean, Daniel Lighezan, Slavita Rotunjanu, Viviana Ivan, and Christian Banciu. 2024. "Lactiplantibacillus plantarum Induces Apoptosis in Melanoma and Breast Cancer Cells" Microorganisms 12, no. 1: 182. https://doi.org/10.3390/microorganisms12010182