
Aerotolerant Thiosulfate-Reducing Bacterium Fusibacter sp. Strain WBS Isolated from Littoral Bottom Sediments of the White Sea—Biochemical and Genome Analysis
 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Sampling
2.2. Isolation and Cultivation of the Strain
2.3. Determination of Specific Activities of Antioxidant Enzymes
2.4. Genome Analysis
3. Results
3.1. Morphological and Physiological Properties of the Strain WBS
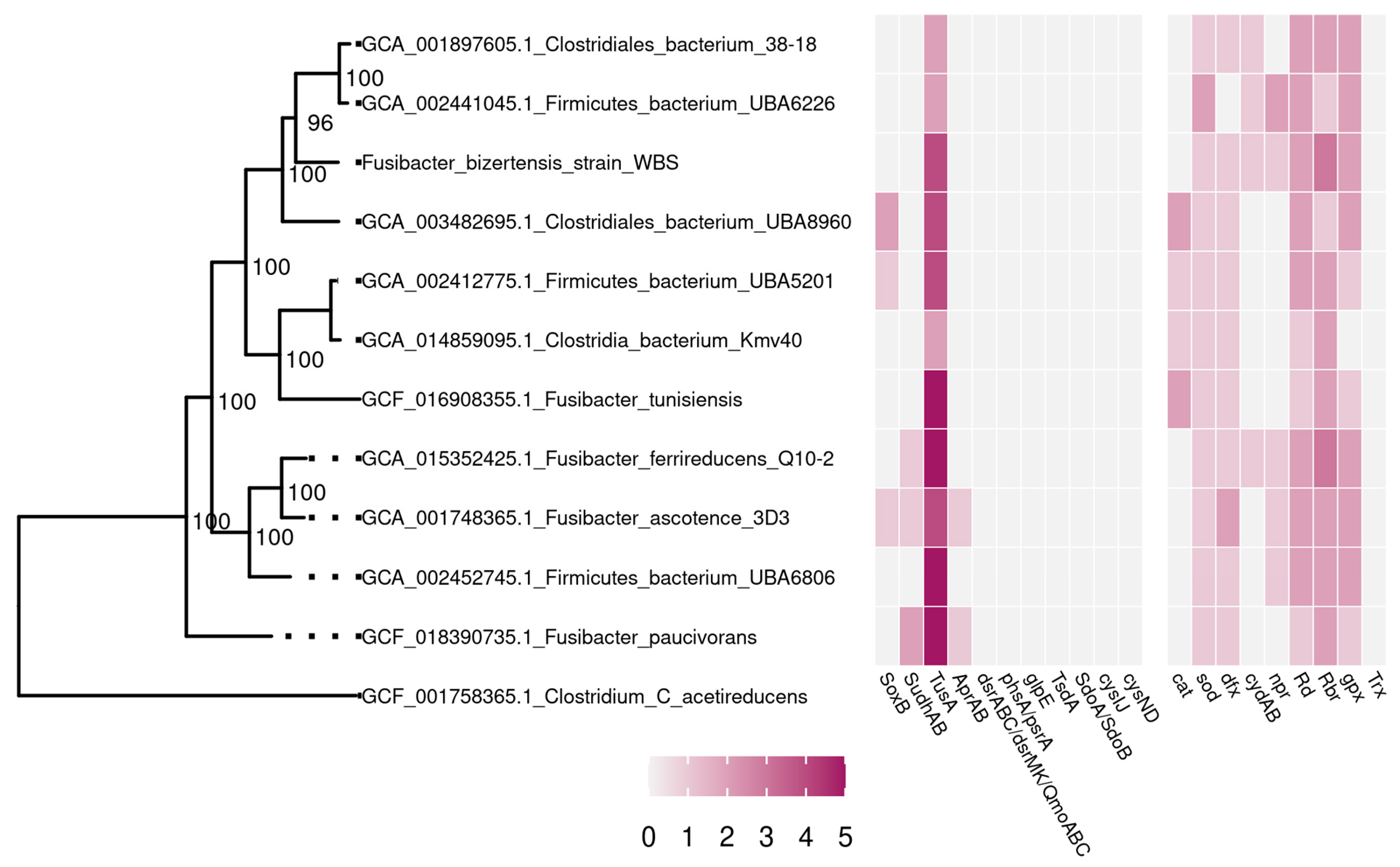
3.2. Phylogenetic and Genome Analysis of the Strain WBS
3.3. Metabolic Capabilities of the Strain WBS According to Genome Analysis
3.4. Systems of Antioxidant Defense of the Strain WBS
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Savvichev, A.S.; Demidenko, N.A.; Rusanov, I.I.; Zakharova, E.E.; Veslopolova, E.F.; Afonina, I.; Ankudinova, I.; Pimenov, N.V.; Ivanov, M.V. Study of the microbial processes in the water column and bottom sediments of the Dolgaya-Vostochnaya Bay (Barents Sea) before construction of the northern tidal power plant. Microbiology 2009, 78, 798–801. [Google Scholar] [CrossRef]
- Ravenschlag, K.; Sahm, K.; Knoblauch, C.; Jørgensen, B.B.; Amann, R. Community structure, cellular rRNA content, and activity of sulfate-reducing bacteria in marine Arctic sediments. Appl. Environ. Microbiol. 2000, 66, 3592–3602. [Google Scholar] [CrossRef] [Green Version]
- Brioukhanov, A.L.; Kadnikov, V.V.; Rusanov, I.I.; Novigatskiy, A.N.; Kanapatskiy, T.A.; Politova, N.V.; Ravin, N.V.; Pimenov, N.V. Phylogenetic diversity in sulphate-reducing bacterial communities from oxidised and reduced bottom sediments of the Barents Sea. Antonie Leeuwenhoek 2022, 115, 801–820. [Google Scholar] [CrossRef]
- Knoblauch, C.; Sahm, K.; Jørgensen, B.B. Psychrophilic sulfate-reducing bacteria isolated from permanently cold Arctic marine sediments: Description of Desulfofrigus oceanense gen. nov., sp. nov., Desulfofrigus fragile sp. nov., Desulfofaba gelida gen. nov., sp. nov., Desulfotalea psychrophila gen. nov., sp. nov. and Desulfotalea arctica sp. nov. Int. J. Syst. Evol. Microbiol. 1999, 49, 1631–1643. [Google Scholar] [CrossRef]
- Könneke, M.; Kuever, J.; Galushko, A.; Jørgensen, B.B. Desulfoconvexum algidum gen. nov., sp. nov., a psychrophilic sulfate-reducing bacterium isolated from a permanently cold marine sediment. Int. J. Syst. Evol. Microbiol. 2013, 63, 959–964. [Google Scholar] [CrossRef]
- Brioukhanov, A.L.; Pieulle, L.; Dolla, A. Antioxidative defense systems of anaerobic sulfate-reducing microorganisms. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; Volume 1, pp. 148–159. [Google Scholar]
- Widdel, F.; Bak, F. Gram-negative mesophilic sulfate-reducing bacteria. In The Prokaryotes, 2nd ed.; Balows, A., Trüper, H.G., Dworkin, M., Harder, W., Schleifer, K.-H., Eds.; Springer: New York, NY, USA, 1992; Volume 4, pp. 3352–3378. [Google Scholar]
- Jørgensen, B.B. Mineralization of organic matter in the sea bed—the role of sulphate reduction. Nature 1982, 296, 643–645. [Google Scholar] [CrossRef]
- Müller, A.L.; Pelikan, C.; de Rezende, J.R.; Wasmund, K.; Putz, M.; Glombitza, C.; Kjeldsen, K.U.; Jørgensen, B.B.; Loy, A. Bacterial interactions during sequential degradation of cyanobacterial necromass in a sulfidic arctic marine sediment. Environ. Microbiol. 2018, 20, 2927–2940. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ravot, G.; Magot, M.; Fardeau, M.-L.; Patel, B.; Thomas, P.; Garcia, J.-L.; Ollivier, B. Fusibacter paucivorans gen. nov., sp. nov., an anaerobic, thiosulfate-reducing bacterium from an oil-producing well. Int. J. Syst. Evol. Microbiol. 1999, 49, 1141–1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ben Hania, W.; Fraj, B.; Postec, A.; Fadhlaoui, K.; Hamdi, M.; Ollivier, B.; Fardeau, M.-L. Fusibacter tunisiensis sp. nov., isolated from an anaerobic reactor used to treat olive-mill wastewater. Int. J. Syst. Evol. Microbiol. 2012, 62, 1365–1368. [Google Scholar] [CrossRef] [PubMed]
- Smii, L.; Ben Hania, W.; Cayol, J.-L.; Joseph, M.; Hamdi, M.; Ollivier, B.; Fardeau, M.-L. Fusibacter bizertensis sp. nov., isolated from a corroded kerosene storage tank. Int. J. Syst. Evol. Microbiol. 2015, 65, 117–121. [Google Scholar] [CrossRef] [Green Version]
- Fadhlaoui, K.; Ben Hania, W.; Postec, A.; Fauque, G.; Hamdi, M.; Ollivier, B.; Fardeau, M.-L. Fusibacter fontis sp. nov., a sulfur-reducing, anaerobic bacterium isolated from a mesothermic Tunisian spring. Int. J. Syst. Evol. Microbiol. 2015, 65, 3501–3506. [Google Scholar] [CrossRef]
- Qiu, D.; Zeng, X.; Zeng, L.; Li, G.; Shao, Z. Fusibacter ferrireducens sp. nov., an anaerobic, Fe(III)- and sulphur-reducing bacterium isolated from mangrove sediment. Int. J. Syst. Evol. Microbiol. 2021, 71. [Google Scholar] [CrossRef] [PubMed]
- Savvichev, A.S.; Rusanov, I.I.; Zakharova, E.E.; Veslopolova, E.F.; Mitskevich, I.N.; Kravchishina, M.D.; Lein, A.I.; Ivanov, M.V. Microbial processes of the carbon and sulfur cycles in the White Sea. Microbiology 2008, 77, 823–838. [Google Scholar] [CrossRef] [PubMed]
- Hungate, R.E.; Macy, J. The roll-tube method for cultivation of strict anaerobes. Bull. Ecol. Res. Comm. 1973, 17, 123–125. [Google Scholar]
- Widdel, F.; Pfennig, N. Studies on dissimilatory sulphate reducing bacteria that decompose fatty acids II. Incomplete oxidation of propionate by Desulfobulbus propionicus gen. nov., sp. nov. Arch. Microbiol. 1982, 131, 360–365. [Google Scholar] [CrossRef]
- Trüper, H.G.; Schlegel, H.G. Sulphur metabolism in Thiorhodaceae. 1. Quantitative measurements on growing cells of Chromatium okenii. Antonie Leeuwenhoek 1964, 30, 225–238. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. The utility of superoxide dismutase in studying free radical reactions. I. Radicals generated by the interaction of sulfite, dimethyl sulfoxide, and oxygen. J. Biol. Chem. 1969, 244, 6056–6063. [Google Scholar] [CrossRef]
- Gallati, H. Horseradish peroxidase: A study of the kinetics and the determination of optimal reaction conditions, using hydrogen peroxide and 2,2′-azinobis 3-ethylbenzthiazoline-6-sulfonic acid (ABTS) as substrates. J. Clin. Chem. Clin. Biochem. 1979, 17, 1–7. [Google Scholar]
- Coulter, E.; Shenvi, N.; Kurtz, D. NADH peroxidase activity of rubrerythrin. Biochem. Biophys. Res. Commun. 1999, 255, 317–323. [Google Scholar] [CrossRef]
- Nurk, S.; Bankevich, A.; Antipov, D.; Gurevich, A.A.; Korobeynikov, A.; Lapidus, A.; Prjibelski, A.D.; Pyshkin, A.; Sirotkin, A.; Sirotkin, Y.; et al. Assembling single-cell genomes and mini-metagenomes from chimeric MDA products. J. Comput. Biol. 2013, 20, 714–737. [Google Scholar] [CrossRef] [Green Version]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef] [PubMed]
- Brettin, T.; Davis, J.J.; Disz, T.; Edwards, R.A.; Gerdes, S.; Olsen, G.J.; Olson, R.; Overbeek, R.; Parrello, B.; Pusch, G.D.; et al. RASTtk: A modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 2015, 5, 8365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez-R, L.M.; Konstantinidis, K.T. The enveomics collection: A toolbox for specialized analyses of microbial genomes and metagenomes. PeerJ Prepr. 2016, 4, e1900v1. [Google Scholar] [CrossRef] [Green Version]
- Chun, J.; Oren, A.; Ventosa, A.; Christensen, H.; Arahal, D.R.; da Costa, M.S.; Rooney, A.P.; Yi, H.; Xu, X.-W.; De Meyer, S.; et al. Proposed minimal standards for the use of genome data for the taxonomy of prokaryotes. Int. J. Syst. Evol. Microbiol. 2018, 68, 461–466. [Google Scholar] [CrossRef] [PubMed]
- Greening, C.; Biswas, A.; Carere, C.; Jackson, C.; Taylor, M.; Stott, M.; Cook, G.M.; Morales, S.E. Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilised energy source for microbial growth and survival. ISME J. 2016, 10, 761–777. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biegel, E.; Schmidt, S.; González, J.M.; Müller, V. Biochemistry, evolution and physiological function of the Rnf complex, a novel ion-motive electron transport complex in prokaryotes. Cell. Mol. Life Sci. 2011, 68, 613–634. [Google Scholar] [CrossRef]
- Malinen, A.M.; Belogurov, G.A.; Baykov, A.A.; Lahti, R. Na+-pyrophosphatase: A novel primary sodium pump. Biochemistry 2007, 46, 8872–8878. [Google Scholar] [CrossRef]
- Marschall, C.; Frenzel, P.; Cypionka, H. Influence of oxygen on sulfate reduction and growth of sulfate-reducing bacteria. Arch. Microbiol. 1993, 159, 168–173. [Google Scholar] [CrossRef]
- Serrano, A.E.; Escudero, L.V.; Tebes-Cayo, C.; Acosta, M.; Encalada, O.; Fernández-Moroso, S.; Demergasso, C. First draft genome sequence of a strain from the genus Fusibacter isolated from Salar de Ascotán in Northern Chile. Stand. Genom. Sci. 2017, 12, 43–51. [Google Scholar] [CrossRef] [Green Version]
- Ravot, G.; Ollivier, B.; Magot, M.; Patel, B.; Crolet, J.; Fardeau, M.; Garcia, J. Thiosulfate reduction, an important physiological feature shared by members of the order Thermotogales. Appl. Environ. Microbiol. 1995, 61, 2053–2055. [Google Scholar] [CrossRef] [Green Version]
- Fardeau, M.-L.; Faudon, C.; Cayol, J.-L.; Magot, M.; Patel, B.; Ollivier, B. Effect of thiosulphate as electron acceptor on glucose and xylose oxidation by Thermoanaerobacter finnii and a Thermoanaerobacter sp. isolated from oil field water. Res. Microbiol. 1996, 147, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Crolet, J.-L.; Magot, M. Non-SRB sulfidogenic bacteria from oil field production facilities. Mater. Perform. 1996, 37, 60–64. [Google Scholar]
- Bak, F.; Pfennig, N. Chemolithotrophic growth of Desulfovibrio sulfodismutans sp. nov. by disproportionation of inorganic sulfur compounds. Arch. Microbiol. 1987, 147, 184–189. [Google Scholar] [CrossRef]
- Janssen, P.H.; Schuhmann, A.; Bak, F.; Liesack, W. Disproportionation of inorganic sulfur compounds by the sulfate-reducing bacterium Desulfocapsa thiozymogenes gen. nov., sp. nov. Arch. Microbiol. 1996, 166, 184–192. [Google Scholar] [CrossRef]
- Umezawa, K.; Kojima, H.; Kato, Y.; Fukui, M. Disproportionation of inorganic sulfur compounds by a novel autotrophic bacterium belonging to Nitrospirota. Syst. Appl. Microbiol. 2020, 43, 126110. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Huang, H.; Wang, Y.; Li, L.; Yi, J.; Wang, S. A Cytoplasmic NAD(P)H-dependent polysulfide reductase with thiosulfate reductase activity from the hyperthermophilic bacterium Thermotoga maritima. Microbiol. Spectr. 2022, 10, e0043622. [Google Scholar] [CrossRef]


{kind=link}
| Characteristic (Optimum) | Strain WBS | Strain LTF Kr01T [12] |
|---|---|---|
| Morphology | Long rods | Short rods |
| DNA G + C content, mol% | 35.1 | 37.4 |
| Temperature growth range | 8–32 °C (18 °C) | 15–40 °C (30 °C) |
| pH growth range | 5.20–8.30 (7.20) | 5.50–8.16 (7.20) |
| NaCl growth range | 0–70 g L−1 (32) | 0–50 g L−1 (5) |
| Electron acceptors | ||
| Sulfate | + | − |
| Thiosulfate | + | + |
| Sulfur | + | + |
| Substrates utilized | ||
| Lactate | + | − |
| d-Glucose | + | + |
| d-Fructose | + | + |
| d-Ribose | + | + |
| Cellobiose | ± | + |
| d-Mannitol | ± | + |
| d-Xylose | ± | + |
| d-Galactose | + | + |
| d-Arabinose | − | − |
| Maltose | ± | + |
| d-Sucrose | − | + |
| d-Trehalose | − | + |
| d-Mannose | ± | − |
| d-Sorbose | − | − |
| Propionate | − | − |
| SOD activity | 17.4 ± 1.5 U mg−1 prot. | |
| Catalase activity | − | |
| Peroxidase activity | 3.2 ± 0.2 U mg−1 prot. | |
| NADH peroxidase activity | + | |
| O2-consumption rate | 11 ± 2 nmol O2 min−1 mg−1 prot. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brioukhanov, A.L.; Kadnikov, V.V.; Beletsky, A.V.; Savvichev, A.S. Aerotolerant Thiosulfate-Reducing Bacterium Fusibacter sp. Strain WBS Isolated from Littoral Bottom Sediments of the White Sea—Biochemical and Genome Analysis. Microorganisms 2023, 11, 1642. https://doi.org/10.3390/microorganisms11071642
Brioukhanov AL, Kadnikov VV, Beletsky AV, Savvichev AS. Aerotolerant Thiosulfate-Reducing Bacterium Fusibacter sp. Strain WBS Isolated from Littoral Bottom Sediments of the White Sea—Biochemical and Genome Analysis. Microorganisms. 2023; 11(7):1642. https://doi.org/10.3390/microorganisms11071642
Chicago/Turabian StyleBrioukhanov, Andrei L., Vitaly V. Kadnikov, Alexey V. Beletsky, and Alexander S. Savvichev. 2023. "Aerotolerant Thiosulfate-Reducing Bacterium Fusibacter sp. Strain WBS Isolated from Littoral Bottom Sediments of the White Sea—Biochemical and Genome Analysis" Microorganisms 11, no. 7: 1642. https://doi.org/10.3390/microorganisms11071642