
Paracoccidioides lutzii Formamidase Contributes to Fungal Survival in Macrophages

 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Fungal Strains and Growth Conditions
2.3. Construction of fmd P. lutzii Silenced Strains with Antisense RNA Technology
2.4. Validation of P. lutzii AsFmd Transformants and Expression Analysis by RT-qPCR
2.5. Growth Curve and Viability of P. lutzii Silenced Cells
2.6. Total Protein Extraction and Fmd Immunoblotting Assay
2.7. Measurement of Formamidase and Urease Activity
2.8. Urease Heterologous Expression
2.9. Ure Polyclonal Antibodies Production and Immunoblotting
2.10. Ex-Vivo Infection of AsFmd in J774 Macrophages
2.11. Statistical Analysis
3. Results
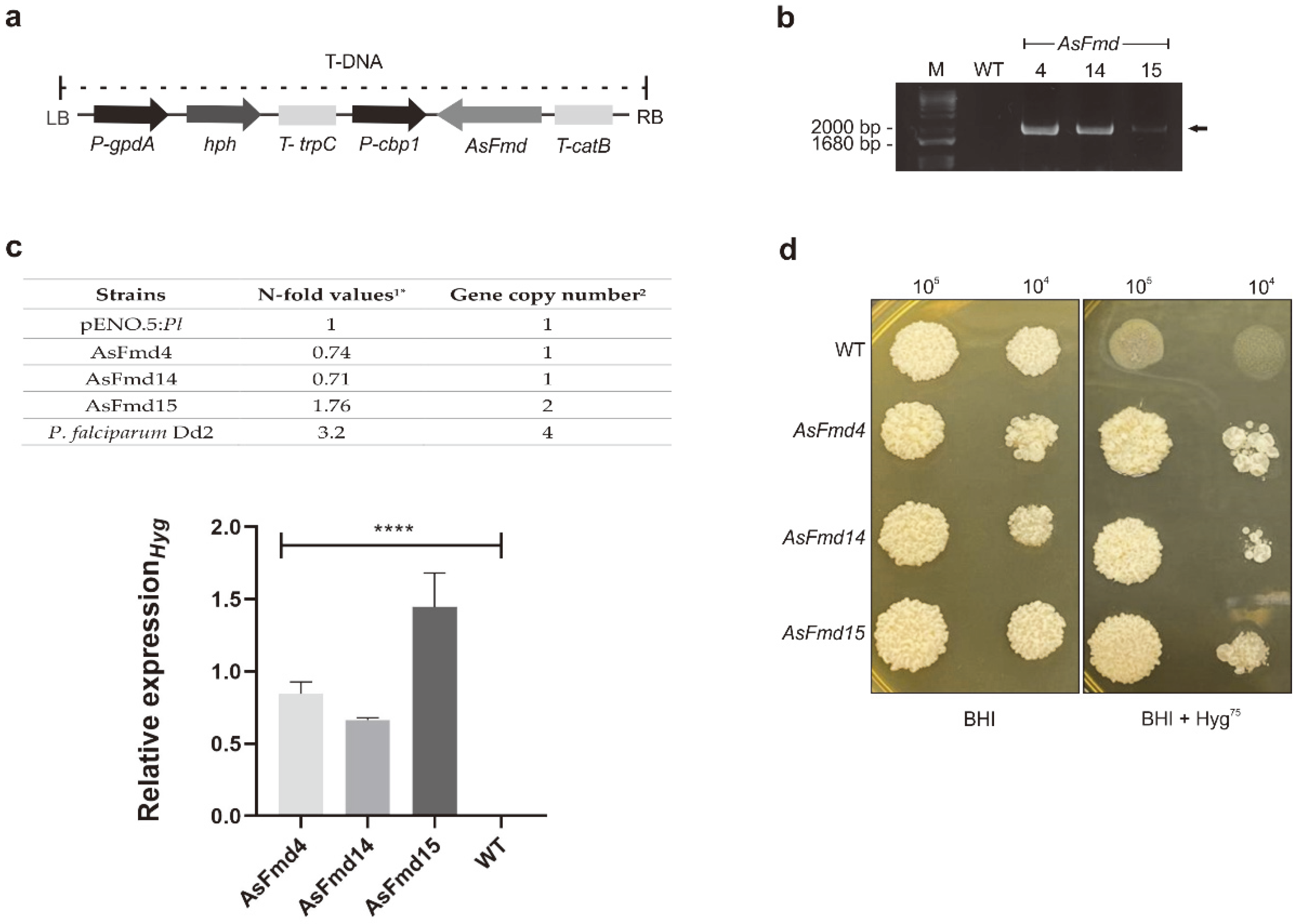
3.1. Production of Fmd P. lutzii Silenced Strains
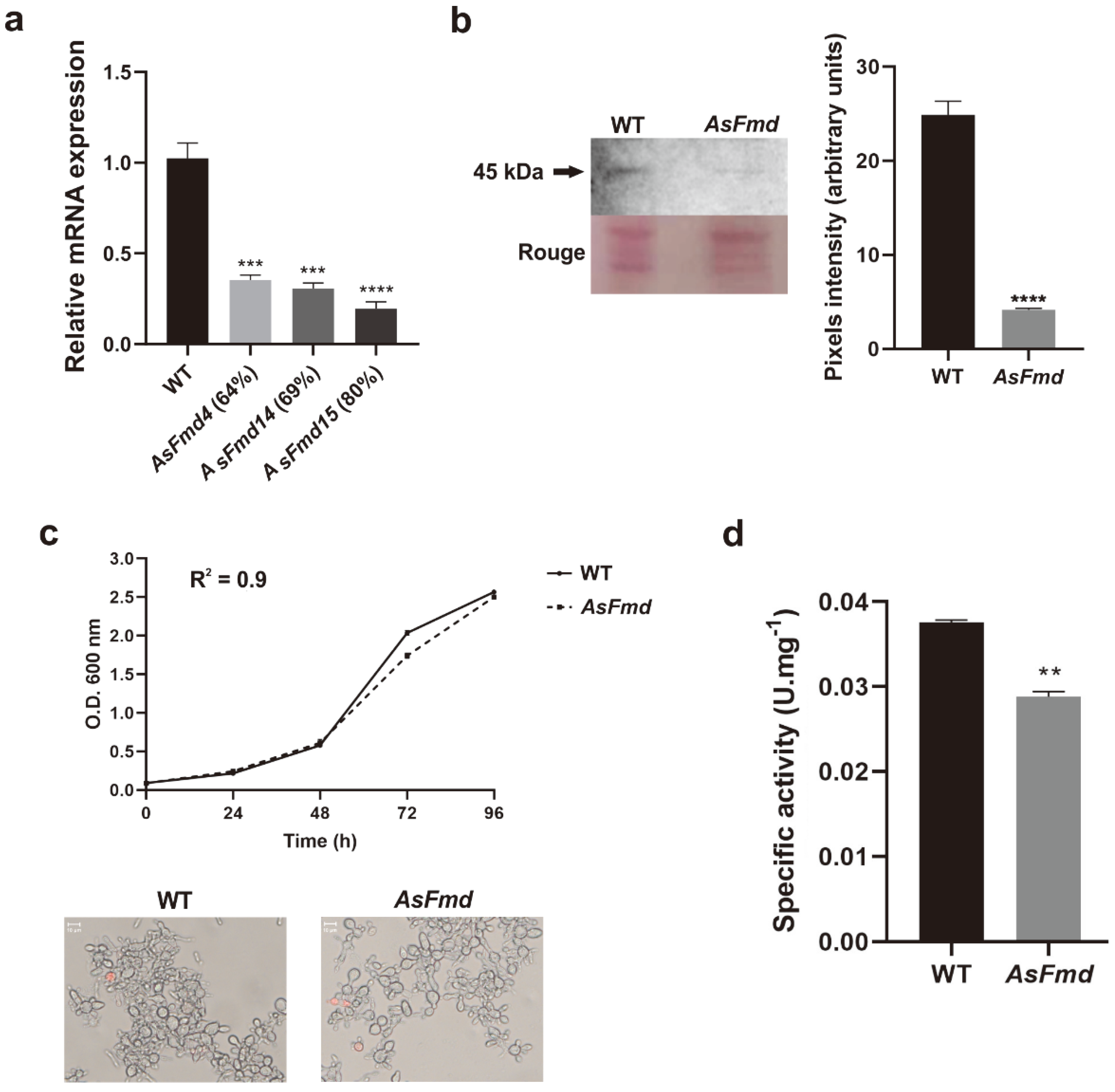
3.2. Reduced Plfmd Gene Expression Does Not Impair Fungal Growth Rate and Cell Viability
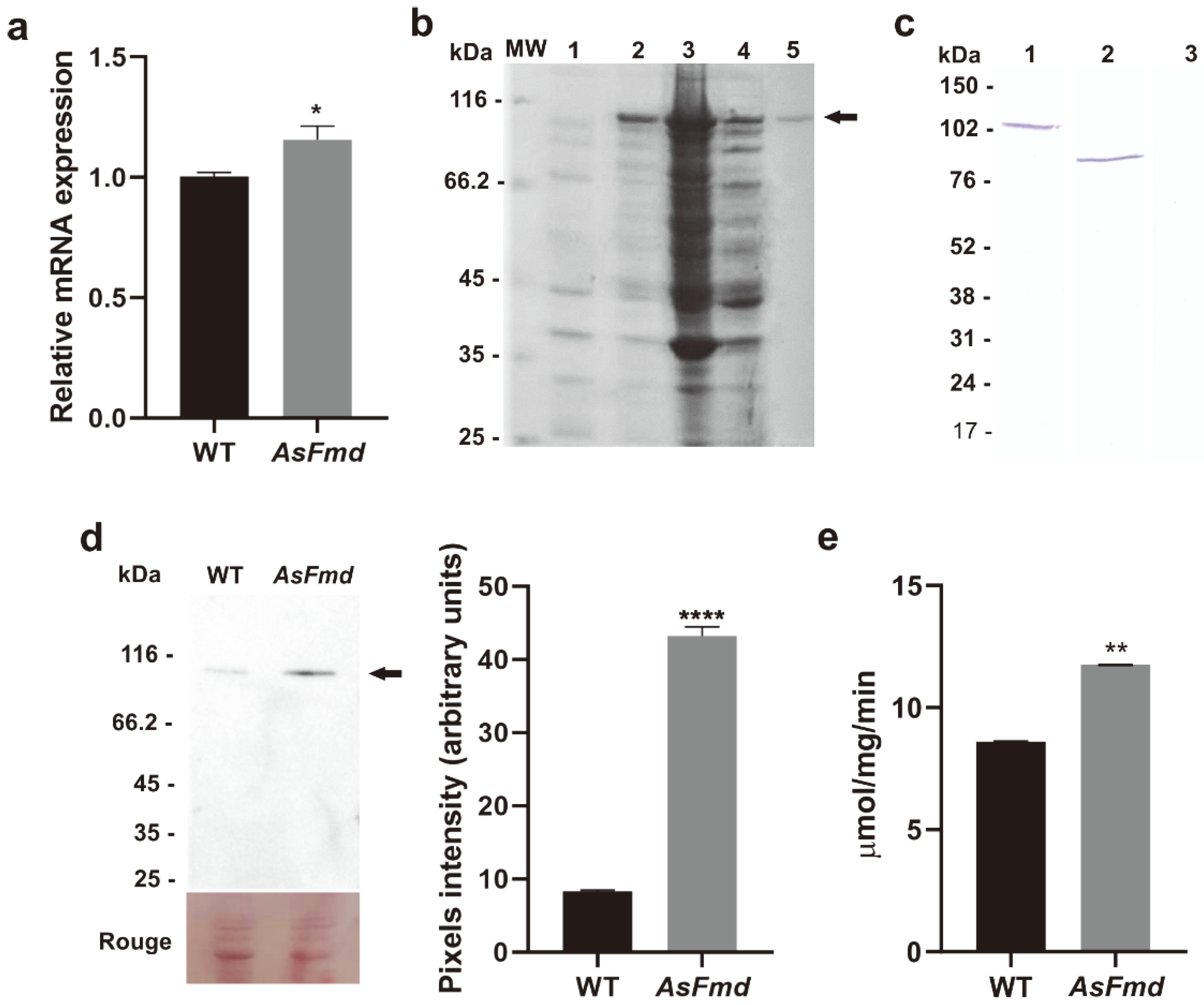
3.3. Decreased PlFmd Expression Levels Influence PlUre Expression and Activity
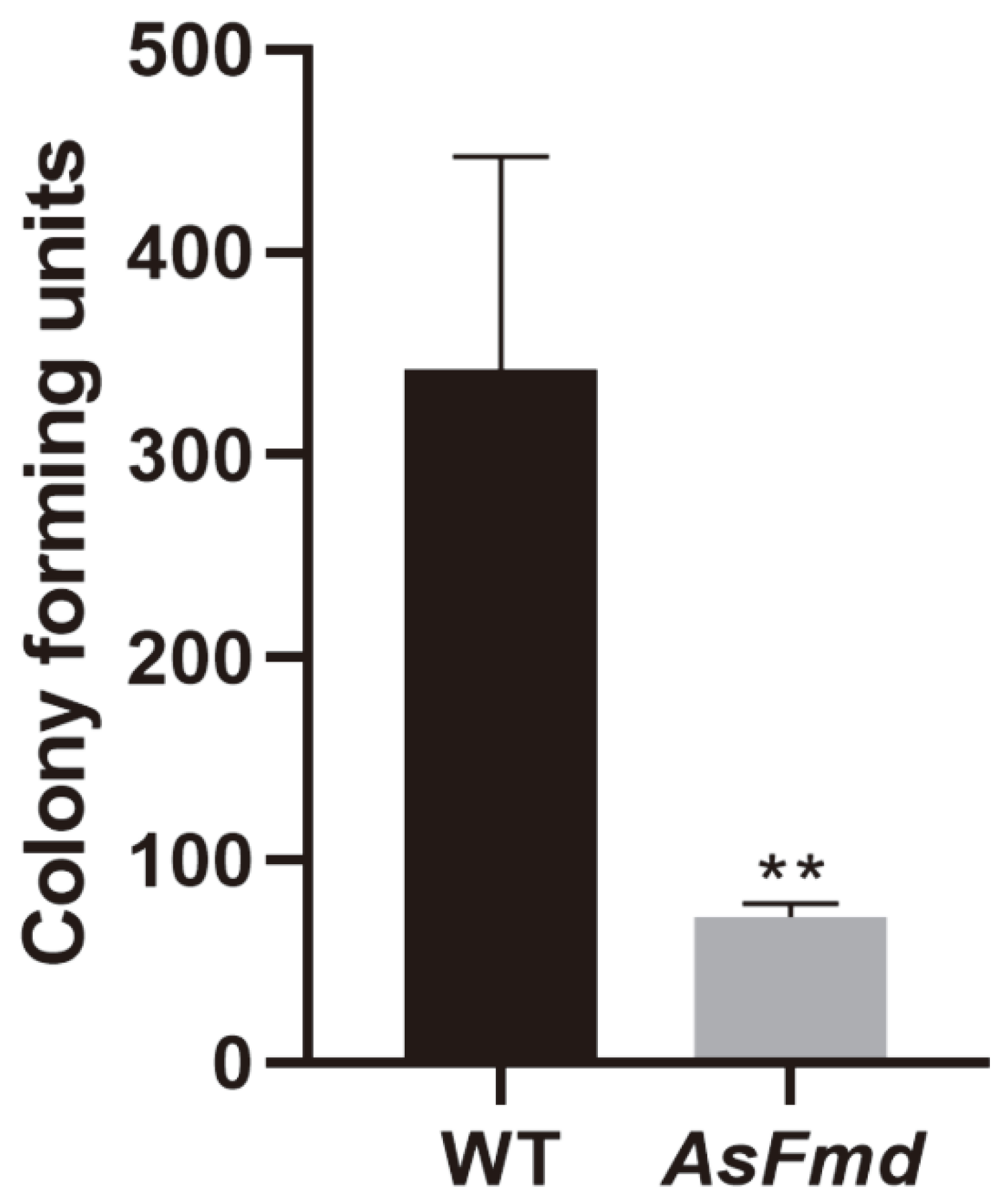
3.4. PlFmd Is Important for P. lutzii Survival during Macrophage Infection
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Turissini, D.A.; Gomez, O.M.; Teixeira, M.M.; McEwen, J.G.; Matute, D.R. Species Boundaries in the Human Pathogen Paracoccidioides. Fungal Genet. Biol. 2017, 106, 9–25. [Google Scholar] [CrossRef]
- Vilela, R.; Huebner, M.; Vilela, C.; Vilela, G.; Pettersen, B.; Oliveira, C.; Mendoza, L. The Taxonomy of Two Uncultivated Fungal Mammalian Pathogens Is Revealed through Phylogeny and Population Genetic Analyses. Sci. Rep. 2021, 11, 18119. [Google Scholar] [CrossRef]
- Martinez, R. New Trends in Paracoccidioidomycosis Epidemiology. J. Fungi 2017, 3, 1. [Google Scholar] [CrossRef] [Green Version]
- Colombo, A.L.; Tobn, A.; Restrepo, A.; Queiroz-Telles, F.; Nucci, M. Epidemiology of Endemic Systemic Fungal Infections in Latin America. Med. Mycol. 2011, 49, 785–798. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shikanai-Yasuda, M.A.; Mendes, R.P.; Colombo, A.L.; de Queiroz-Telles, F.; Kono, A.S.G.; Paniago, A.M.M.; Nathan, A.; do Valle, A.C.F.; Bagagli, E.; Benard, G.; et al. Brazilian Guidelines for the Clinical Management of Paracoccidioidomycosis. Rev. Soc. Bras. Med. Trop. 2017, 50, 715–740. [Google Scholar] [CrossRef]
- Seider, K.; Heyken, A.; Lüttich, A.; Miramón, P.; Hube, B. Interaction of Pathogenic Yeasts with Phagocytes: Survival, Persistence and Escape. Curr. Opin. Microbiol. 2010, 13, 392–400. [Google Scholar] [CrossRef]
- Gonzalez, A.; Hernandez, O. New Insights into a Complex Fungal Pathogen: The Case of Paracoccidioides spp. Yeast 2016, 33, 113–128. [Google Scholar] [CrossRef] [Green Version]
- Brummer, E.; Hanson, L.H.; Restrepo, A.; Stevens, D.A. Intracellular Multiplication of Paracoccidioides brasiliensis in Macrophages: Killing and Restriction of Multiplication by Activated Macrophages. Infect. Immun. 1989, 57, 2289–2294. [Google Scholar] [CrossRef] [Green Version]
- Moscardi-Bacchi, M.; Brummer, E.; Stevens, D.A. Support of Paracoccidioides brasiliensis Multiplication by Human Monocytes or Macrophages: Inhibition by Activated Phagocytes. J. Med. Microbiol. 1994, 40, 159–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cezar-dos-Santos, F.; Assolini, J.P.; Okuyama, N.C.M.; Viana, K.F.; de Oliveira, K.B.; Itano, E.N. Unraveling the Susceptibility of Paracoccidioidomycosis: Insights towards the Pathogen-Immune Interplay and Immunogenetics. Infect. Genet. Evol. 2020, 86, 104586. [Google Scholar] [CrossRef] [PubMed]
- Santos, L.A.; Grisolia, J.C.; Burger, E.; de Araujo Paula, F.B.; Dias, A.L.T.; Malaquias, L.C.C. Virulence Factors of Paracoccidioides brasiliensis as Therapeutic Targets: A Review. Antonie Leeuwenhoek Int. J. Gen. Mol. Microbiol. 2020, 113, 593–604. [Google Scholar] [CrossRef]
- Broach, J.R. Nutritional Control of Growth and Development in Yeast. Genetics 2012, 192, 73–105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fleck, C.B.; Schöbel, F.; Brock, M. Nutrient Acquisition by Pathogenic Fungi: Nutrient Availability, Pathway Regulation, and Differences in Substrate Utilization. Int. J. Med. Microbiol. 2011, 301, 400–407. [Google Scholar] [CrossRef] [PubMed]
- Ries, L.N.A.; Beattie, S.; Cramer, R.A.; Goldman, G.H. Overview of Carbon and Nitrogen Catabolite Metabolism in the Virulence of Human Pathogenic Fungi. Mol. Microbiol. 2018, 107, 277–297. [Google Scholar] [CrossRef] [Green Version]
- Ljungdahl, P.O.; Daignan-Fornier, B. Regulation of Amino Acid, Nucleotide, and Phosphate Metabolism in Saccharomyces cerevisiae. Genetics 2012, 190, 885–929. [Google Scholar] [CrossRef] [Green Version]
- Fraser, J.A.; Davis, M.A.; Hynes, M.J. The Formamidase Gene of Aspergillus nidulans: Regulation by Nitrogen Metabolite Repression and Transcriptional Interference by an Overlapping Upstream Gene. Genetics 2001, 157, 119–131. [Google Scholar] [CrossRef]
- Cruz-Leite, V.R.M.; Salem-Izacc, S.M.; Novaes, E.; Neves, B.J.; de Almeida Brito, W.; Silva, L.O.S.; Paccez, J.D.; Parente-Rocha, J.A.; Pereira, M.; de Almeida Soares, C.M.; et al. Nitrogen Catabolite Repression in Members of Paracoccidioides Complex. Microb. Pathog. 2020, 149, 104281. [Google Scholar] [CrossRef]
- Felipe, M.S.S.; Andrade, R.V.; Petrofeza, S.S.; Maranhão, A.Q.; Torres, F.A.G.; Albuquerque, P.; Arraes, F.B.M.; Arruda, M.; Azevedo, M.O.; Baptista, A.J.; et al. Transcriptome Characterization of the Dimorphic and Pathogenic Fungus Paracoccidioides brasiliensis by EST Analysis. Yeast 2003, 20, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Bailão, A.M.; Borges, C.L.; Maria de Almeida Soares, C.; Pereira, M.; Soares Mendes-Giannini, M.J.; Schrank, A.; Dutra, V.; Walquíria Inês Molinari-Madlum, E.E.; Martins, W.S.; Soares Felipe, M.S. Differential Gene Expression by Paracoccidioides brasiliensis in Host Interaction Conditions: Representational Difference Analysis Identifies Candidate Genes Associated with Fungal Pathogenesis. Microbes Infect. 2006, 8, 2686–2697. [Google Scholar] [CrossRef] [PubMed]
- Borges, C.L.; Pereira, M.; Felipe, M.S.S.; De Faria, F.P.; Gomez, F.J.; Deepe, G.S.; de Almeida Soares, C.M. The Antigenic and Catalytically Active Formamidase of Paracoccidioides brasiliensis: Protein Characterization, CDNA and Gene Cloning, Heterologous Expression and Functional Analysis of the Recombinant Protein. Microbes Infect. 2005, 7, 66–77. [Google Scholar] [CrossRef]
- Araújo, D.S.; de Sousa Lima, P.; Baeza, L.C.; Parente, A.F.A.; Melo Bailão, A.; Borges, C.L.; de Almeida Soares, C.M. Employing Proteomic Analysis to Compare Paracoccidioides lutzii Yeast and Mycelium Cell Wall Proteins. Biochim. Biophys. Acta Proteins Proteom. 2017, 1865, 1304–1314. [Google Scholar] [CrossRef] [PubMed]
- Borges, C.L.; Parente, J.A.; Barbosa, M.S.; Santana, J.M.; Báo, S.N.; de Sousa, M.V.; de Almeida Soares, C.M. Detection of a Homotetrameric Structure and Protein-Protein Interactions of Paracoccidioides brasiliensis Formamidase Lead to New Functional Insights. FEMS Yeast Res. 2010, 10, 104–113. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.L.E.; Oliveira, M.A.P.; Silva, L.O.S.; Inácio, M.M.; Bailão, A.M.; Parente-Rocha, J.A.; Cruz-Leite, V.R.M.; Paccez, J.D.; de Almeida Soares, C.M.; Weber, S.S.; et al. Immunoproteomic Approach of Extracellular Antigens From Paracoccidioides Species Reveals Exclusive B-Cell Epitopes. Front. Microbiol. 2020, 10, 2968. [Google Scholar] [CrossRef]
- Parente-Rocha, J.A.; Parente, A.F.A.; Baeza, L.C.; Bonfim, S.M.R.C.; Hernandez, O.; McEwen, J.G.; Bailão, A.M.; Taborda, C.P.; Borges, C.L.; De Almeida Soares, C.M. Macrophage Interaction with Paracoccidioides brasiliensis Yeast Cells Modulates Fungal Metabolism and Generates a Response to Oxidative Stress. PLoS ONE 2015, 10, e137619. [Google Scholar] [CrossRef] [Green Version]
- Chaves, E.G.A.; Parente-Rocha, J.A.; Baeza, L.C.; Araújo, D.S.; Borges, C.L.; de Oliveira, M.A.P.; de Almeida Soares, C.M. Proteomic Analysis of Paracoccidioides brasiliensis during Infection of Alveolar Macrophages Primed or Not by Interferon-Gamma. Front. Microbiol. 2019, 10, 96. [Google Scholar] [CrossRef]
- Pigosso, L.L.; Baeza, L.C.; Tomazett, M.V.; Faleiro, M.B.R.; Brianezi Dignani de Moura, V.M.; Bailão, A.M.; Borges, C.L.; Parente-Rocha, J.A.; Fernandes, G.R.; Gauthier, G.M.; et al. Paracoccidioides brasiliensis Presents Metabolic Reprogramming and Secretes a Serine Proteinase during Murine Infection. Virulence 2017, 8, 1417–1434. [Google Scholar] [CrossRef] [Green Version]
- Moreira, A.L.E.; Cruz-Leite, V.R.M.; Silva, L.O.S.; Parente, A.F.A.; Bailão, A.M.; de Almeida Soares, C.M.; Parente-Rocha, J.A.; Ruiz, O.H.; Borges, C.L. Proteome Characterization of Paracoccidioides lutzii Conidia by Using NanoUPLC-MSE. Fungal Biol. 2020, 124, 766–780. [Google Scholar] [CrossRef] [PubMed]
- Rezende, T.C.V.; Borges, C.L.; Magalhães, A.D.; de Sousa, M.V.; Ricart, C.A.O.; Bailão, A.M.; Soares, C.M.A. A Quantitative View of the Morphological Phases of Paracoccidioides brasiliensis Using Proteomics. J. Proteom. 2011, 75, 572–587. [Google Scholar] [CrossRef]
- Parente-Rocha, J.A.; Tomazett, M.V.; Pigosso, L.L.; Bailão, A.M.; Ferreira de Souza, A.; Paccez, J.D.; Baeza, L.C.; Pereira, M.; Silva Bailão, M.G.; Borges, C.L.; et al. In Vitro, Ex Vivo and in Vivo Models: A Comparative Analysis of Paracoccidioides spp. Proteomic Studies. Fungal Biol. 2018, 122, 505–513. [Google Scholar] [CrossRef]
- Almeida, A.J.; Carmona, J.A.; Cunha, C.; Carvalho, A.; Rappleye, C.A.; Goldman, W.E.; Hooykaas, P.J.; Leão, C.; Ludovico, P.; Rodrigues, F. Towards a Molecular Genetic System for the Pathogenic Fungus Paracoccidioides brasiliensis. Fungal Genet. Biol. 2007, 44, 1387–1398. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.F.; Tomazett, M.V.; E Silva, K.S.F.; de Curcio, J.S.; Pereira, C.A.; Baeza, L.C.; Paccez, J.D.; Gonçales, R.A.; Rodrigues, F.; Pereira, M.; et al. Interacting with Hemoglobin: Paracoccidioides spp. Recruits Hsp30 on Its Cell Surface for Enhanced Ability to Use This Iron Source. J. Fungi 2021, 7, 21. [Google Scholar] [CrossRef] [PubMed]
- Rappleye, C.A.; Engle, J.T.; Goldman, W.E. RNA Interference in Histoplasma capsulatum Demonstrates a Role for α-(1,3)-Glucan in Virulence. Mol. Microbiol. 2004, 53, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Bookout, A.L.; Cummins, C.L.; Mangelsdorf, D.J.; Pesola, J.M.; Kramer, M.F. High-Throughput Real-Time Quantitative Reverse Transcription PCR. Curr. Protoc. Mol. Biol. 2006, 73, 1–28. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Leite, V.R.M.; Tomazett, M.V.; Santana de Curcio, J.; Sbaraini, N.; Bailão, A.M.; Gonçales, R.A.; Moraes, D.; Pereira, M.; Vainstein, M.H.; Schrank, A.; et al. Bioluminescence Imaging in Paracoccidioides spp.: A Tool to Monitor the Infectious Processes. Microbes Infect. 2022, 24, 104975. [Google Scholar] [CrossRef]
- Ferreira, I.D.; do Rosário, V.E.; Cravo, P.V. Real-Time Quantitative PCR with SYBR Green I Detection for Estimating Copy Numbers of Nine Drug Resistance Candidate Genes in Plasmodium falciparum. Malar. J. 2006, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Silva, M.G.; de Curcio, J.S.; Silva-Bailão, M.G.; Lima, R.M.; Tomazett, M.V.; de Souza, A.F.; Cruz-Leite, V.R.M.; Sbaraini, N.; Bailão, A.M.; Rodrigues, F.; et al. Molecular Characterization of Siderophore Biosynthesis in Paracoccidioides brasiliensis. IMA Fungus 2020, 11, 11. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Follmer, C. Insights into the Role and Structure of Plant Ureases. Phytochemistry 2008, 69, 18–28. [Google Scholar] [CrossRef]
- Carlini, C.R.; Polacco, J.C. Toxic Properties of Urease. Crop Sci. 2008, 48, 1665–1672. [Google Scholar] [CrossRef]
- Carlini, C.R.; Ligabue-Braun, R. Ureases as Multifunctional Toxic Proteins: A Review. Toxicon 2016, 110, 90–109. [Google Scholar] [CrossRef]
- Lin, W.; Mathys, V.; Ang, E.L.Y.; Koh, V.H.Q.; Martínez Gómez, J.M.; Ang, M.L.T.; Zainul Rahim, S.Z.; Tan, M.P.; Pethe, K.; Alonso, S. Urease Activity Represents an Alternative Pathway for Mycobacterium tuberculosis Nitrogen Metabolism. Infect. Immun. 2012, 80, 2771–2779. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fu, M.S.; Coelho, C.; De Leon-Rodriguez, C.M.; Rossi, D.C.P.; Camacho, E.; Jung, E.H.; Kulkarni, M.; Casadevall, A. Cryptococcus neoformans Urease Affects the Outcome of Intracellular Pathogenesis by Modulating Phagolysosomal PH. PLoS Pathog. 2018, 14, e1007144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rappleye, C.A.; Goldman, W.E. Defining Virulence Genes in the Dimorphic Fungi. Annu. Rev. Microbiol. 2006, 60, 281–303. [Google Scholar] [CrossRef] [PubMed]
- Brummer, E.; Castaneda, E.; Restrepo, A.; De Microbiologia, G.; De Bogota, S. Paracoccidioidomycosis: An Update. Clin. Microbiol. Rev. 1993, 6, 89–117. [Google Scholar] [CrossRef] [PubMed]
- Almeida, A.J.; Cunha, C.; Carmona, J.A.; Sampaio-Marques, B.; Carvalho, A.; Malavazi, I.; Steensma, H.Y.; Johnson, D.I.; Leão, C.; Logarinho, E.; et al. Cdc42p Controls Yeast-Cell Shape and Virulence of Paracoccidioides brasiliensis. Fungal Genet. Biol. 2009, 46, 919–926. [Google Scholar] [CrossRef]
- Tamayo, D.; Muñoz, J.F.; Torres, I.; Almeida, A.J.; Restrepo, A.; McEwen, J.G.; Hernández, O. Involvement of the 90kDa Heat Shock Protein during Adaptation of Paracoccidioides brasiliensis to Different Environmental Conditions. Fungal Genet. Biol. 2013, 51, 34–41. [Google Scholar] [CrossRef]
- Bailão, E.F.L.C.; Parente, J.A.; Pigosso, L.L.; de Castro, K.P.; Fonseca, F.L.; Silva-Bailão, M.G.; Báo, S.N.; Bailão, A.M.; Rodrigues, M.L.; Hernandez, O.; et al. Hemoglobin Uptake by Paracoccidioides spp. Is Receptor-Mediated. PLoS Negl. Trop. Dis. 2014, 8, e2856. [Google Scholar] [CrossRef] [Green Version]
- Fernandes, F.F.; Oliveira, A.F.; Landgraf, T.N.; Cunha, C.; Carvalho, A.; Vendruscolo, P.E.; Gonçales, R.A.; Almeida, F.; da Silva, T.A.; Rodrigues, F.; et al. Impact of Paracoccin Gene Silencing on Paracoccidioides brasiliensis Virulence. mBio 2017, 8, e00537-17. [Google Scholar] [CrossRef] [Green Version]
- Oliveira, L.N.; Gonçales, R.A.; Silva, M.G.; Lima, R.M.; Tomazett, M.V.; de Curcio, J.S.; Paccez, J.D.; Cruz-Leite, V.R.M.; Rodrigues, F.; Lima, P.d.S.; et al. Characterization of a Heme-Protein Responsive to Hypoxia in Paracoccidioides brasiliensis. Fungal Genet. Biol. 2020, 144, 103446. [Google Scholar] [CrossRef]
- Mirbod-Donovan, F.; Schaller, R.; Hung, C.Y.; Xue, J.; Reichard, U.; Cole, G.T. Urease Produced by Coccidioides posadasii Contributes to the Virulence of This Respiratory Pathogen. Infect. Immun. 2006, 74, 504–515. [Google Scholar] [CrossRef]
- Dixon, N.E.; Riddles, P.W.; Gazzola, C.; Blakeley, R.L.; Zerner, B. Jack Bean Urease (EC 3.5.1.5). V. On the Mechanism of Action of Urease on Urea, Formamide, Acetamide, N -Methylurea, and Related Compounds. Can. J. Biochem. 1980, 58, 1335–1344. [Google Scholar] [CrossRef] [PubMed]
- Skouloubris, S.; Labigne, A.; De Reuse, H. The AmiE Aliphatic Amidase and AmiF Formamidase of Helicobacter Pylori: Natural Evolution of Two Enzyme Paralogues. Mol. Microbiol. 2001, 40, 596–609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenwood, J. Physiological Regulation, Purification and Properties of Urease from Methylophilus methylotrophus. FEMS Microbiol. Lett. 1998, 160, 131–135. [Google Scholar] [CrossRef]
- Puccia, R.; Carmona, A.K.; Gesztesi, J.L.; Juliano, L.; Travassos, L.R. Exocellular Proteolytic Activity of Paracoccidioides brasiliensis: Cleavage of Components Associated with the Basement Membrane. Med. Mycol. 1998, 36, 345–348. [Google Scholar] [CrossRef] [Green Version]
- Lee, M.H.; Mulrooney, S.B.; Renner, M.J.; Markowicz, Y.; Hausinger, R.P. Klebsiella aerogenes Urease Gene Cluster: Sequence of UreD and Demonstration That Four Accessory Genes (UreD, UreE, UreF, and UreG) Are Involved in Nickel Metallocenter Biosynthesis. J. Bacteriol. 1992, 174, 4324–4330. [Google Scholar] [CrossRef] [Green Version]
- Fong, Y.H.; Wong, H.C.; Yuen, M.H.; Lau, P.H.; Chen, Y.W.; Wong, K.-B. Structure of UreG/UreF/UreH Complex Reveals How Urease Accessory Proteins Facilitate Maturation of Helicobacter pylori Urease. PLoS Biol. 2013, 11, e1001678. [Google Scholar] [CrossRef] [Green Version]
- Castilho, D.G.; Chaves, A.F.A.; Xander, P.; Zelanis, A.; Kitano, E.S.; Serrano, S.M.T.; Tashima, A.K.; Batista, W.L.; Mirassol, R. Exploring Potential Virulence Regulators in Paracoccidioides brasiliensis Isolates of Varying Virulence through Quantitative Proteomics. J. Proteome Res. 2014, 13, 4259–4271. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silva, L.O.S.; Moreira, T.R.; Gonçales, R.A.; Tomazett, M.V.; Parente-Rocha, J.A.; Mattos, K.; Paccez, J.D.; Ruiz, O.H.; Pereira, M.; Soares, C.M.d.A.; et al. Paracoccidioides lutzii Formamidase Contributes to Fungal Survival in Macrophages. Microorganisms 2022, 10, 2011. https://doi.org/10.3390/microorganisms10102011
Silva LOS, Moreira TR, Gonçales RA, Tomazett MV, Parente-Rocha JA, Mattos K, Paccez JD, Ruiz OH, Pereira M, Soares CMdA, et al. Paracoccidioides lutzii Formamidase Contributes to Fungal Survival in Macrophages. Microorganisms. 2022; 10(10):2011. https://doi.org/10.3390/microorganisms10102011
Chicago/Turabian StyleSilva, Lana O’Hara Souza, Thalison Rodrigues Moreira, Relber Aguiar Gonçales, Mariana Vieira Tomazett, Juliana Alves Parente-Rocha, Karine Mattos, Juliano Domiraci Paccez, Orville Hernandez Ruiz, Maristela Pereira, Célia Maria de Almeida Soares, and et al. 2022. "Paracoccidioides lutzii Formamidase Contributes to Fungal Survival in Macrophages" Microorganisms 10, no. 10: 2011. https://doi.org/10.3390/microorganisms10102011