
Evidence, Challenges, and Knowledge Gaps Regarding Latent Tuberculosis in Animals
by
 , , and
, , and
Pamela Ncube
,
Bahareh Bagheri
,
Wynand Johan Goosen
 ,
,
Michele Ann Miller
and
Samantha Leigh Sampson
* DSI/NRF Centre of Excellence for Biomedical Tuberculosis Research, South African Medical Research Council Centre for Tuberculosis Research, Department of Biomedical Sciences, Division of Molecular Biology and Human Genetics, Faculty of Medicine and Health Sciences, Stellenbosch University, Francie Van Zijl Dr, Parow, Cape Town 7505, South Africa
*
Author to whom correspondence should be addressed.
Microorganisms 2022, 10(9), 1845; https://doi.org/10.3390/microorganisms10091845
Submission received: 12 August 2022
/
Revised: 8 September 2022
/
Accepted: 9 September 2022
/
Published: 15 September 2022
(This article belongs to the Special Issue Pathogen Infection in Wildlife)
Abstract
:Mycobacterium bovis and other Mycobacterium tuberculosis complex (MTBC) pathogens that cause domestic animal and wildlife tuberculosis have received considerably less attention than M. tuberculosis, the primary cause of human tuberculosis (TB). Human TB studies have shown that different stages of infection can exist, driven by host–pathogen interactions. This results in the emergence of heterogeneous subpopulations of mycobacteria in different phenotypic states, which range from actively replicating (AR) cells to viable but slowly or non-replicating (VBNR), viable but non-culturable (VBNC), and dormant mycobacteria. The VBNR, VBNC, and dormant subpopulations are believed to underlie latent tuberculosis (LTB) in humans; however, it is unclear if a similar phenomenon could be happening in animals. This review discusses the evidence, challenges, and knowledge gaps regarding LTB in animals, and possible host–pathogen differences in the MTBC strains M. tuberculosis and M. bovis during infection. We further consider models that might be adapted from human TB research to investigate how the different phenotypic states of bacteria could influence TB stages in animals. In addition, we explore potential host biomarkers and mycobacterial changes in the DosR regulon, transcriptional sigma factors, and resuscitation-promoting factors that may influence the development of LTB.
1. Introduction
Tuberculosis is a global threat to animals and humans. In 2020, there were 1.4 million human TB deaths [1]. Although precise numbers are unknown, an estimated 1%–15% of human TB cases are believed to be zoonotic, with transmission originating from animals [2,3,4]. Generally, M. tuberculosis infection causes human TB [5], with M. bovis the most common infectious agent contributing to animal TB (Table 1) [6]. A wide range of domestic animals (e.g., cattle, goats, sheep, pigs) [7,8,9,10] and wildlife (e.g., buffaloes, elephants, lions, wild dogs, warthogs) [6,11,12] can be infected by M. bovis and other M. tuberculosis complex (MTBC) species including Mycobacterium caprae, Mycobacterium canettii, Mycobacterium pinnipedii, Mycobacterium mungi, Mycobacterium africanum, Mycobacterium suricattae, Mycobacterium microti, Mycobacterium orygis, the “chimpanzee bacillus”, and the “dassie bacillus” (Table 1) [13,14,15,16]. Animals with TB can be a source of infection to other animals and humans [17,18]. Given the zoonotic potential and the detrimental effect that TB can have on humans and animals at human–animal–environment interfaces [19], it is important to develop an understanding of TB pathogenesis during different stages of M. bovis infection in animals in order to improve control and management strategies.
While there is increased recognition of the complexity of TB stages of infection in humans [20] and animals [21,22], limited studies define the characteristics and diagnosis of these stages in naturally infected animals [21,23,24,25]. The identification of animals that are TB test-positive based on immunological responses [e.g., tuberculin skin test (TST) or interferon-gamma (IFN-γ) release assay (IGRA)] but lack visible lesions on necropsy or are mycobacterial culture-negative, has led to a debate on whether animals can develop LTB [24,25,26,27]. Host-(e.g., genetic variation, immunocompetence, comorbidities) [1,28] and pathogen-specific (e.g., MTBC strains, virulence, infecting dose) [29,30,31] factors likely impact the outcome of infection. However, it is unclear whether these stages in animals mimic those observed in humans, and this requires additional research [32].
One global challenge is that current animal TB control programs in many countries require the culling of any suspected infected animal to prevent its spread [33,34,35]. The culling of TB test-positive animals or depopulation of a herd is usually performed without clear knowledge of the animal’s infection stage; this can lead to unnecessary loss of latently infected animals that are not shedding [36,37,38]. Culling animals has significant negative economic implications for animal owners. Similarly, the removal of wildlife can have negative conservation impacts, including loss of genetic diversity for some high-value species, such as rhinoceros [39,40]. It is also important to consider the implications of latent infection in livestock destined for consumption [18,41,42,43] and wildlife such as buffaloes or lions (which are maintenance hosts) [44,45,46]. In addition, the diagnosis of TB can lead to movement restrictions [40], resulting in financial losses from game sales, hunting, and ecotourism [47]. Therefore, there is a crucial need to improve the understanding of animal TB pathogenesis, infection stages, and epidemiology to inform disease control and management strategies.
Knowledge gaps still exist regarding infection stages in animal TB, as well as host–pathogen interactions and the pathogen metabolic processes involved [48]. Although research on TB diagnosis in different species has progressed [12], studies on TB pathogenesis are complicated by the lack of clinical signs in animals until the disease is advanced, which can take months to years [6,49]. Additionally, TB in wildlife may only be detected while conducting postmortem examinations [50]. Cases with early or subclinical infection can be easily missed without a thorough examination [51], especially during abattoir inspection or field necropsy [52]. These challenges have delayed progress in research on the pathogenesis and elucidation of different infection stages in animals.
The importance of understanding the stages of TB infection in animals stems from the impact it has on the diagnosis and management of infection in individuals as well as intra- and inter-species transmission risk [53]. In turn, this can inform disease control strategies in domestic and wild animals, which affects livelihoods, food security, and conservation [2,39,54]. This review describes the current state of latent TB research in domestic and wild animals and the challenges that still hinder this area of investigation. Specifically, there is a focus on understanding how different hosts (humans vs. domestic animals vs. wildlife) vary in the progression of infection, mycobacterial metabolic activity/pathways that may underly latent TB, and the influence of host–pathogen interactions. Further, we discuss the challenges that exist with TB diagnostic tools in animals. By identifying the knowledge gaps surrounding the progression of TB infection in animals, future investigations can target translational research on animal TB management and control strategies, especially for endangered wildlife and species with high commercial value. This would protect the livelihoods of domestic farmers, especially in developing countries, so as to mitigate TB disease and potential zoonotic outbreaks at human–domestic animal–wildlife interfaces.
2. The Epidemiology of TB at the Domestic–Wildlife–Human Interfaces
Targeted epidemiologic studies at domestic–wildlife–human interfaces provide a foundation for effective control, ecological management, surveillance, and eradication of TB [95,146,147,148]. Over the years, the advances in strain-typing methods (such as spoligotyping [149,150,151], variable number of tandem repeats [152,153,154], whole genome sequencing [143,155,156,157,158,159], and other tools for characterising MTBC pathogens [12,22,46,160,161,162] have improved the understanding of genotype distributions [154], demographic risk factors for disease [41,163,164], MTBC geographical distributions of infected animal populations [165,166], and transmission patterns [34,80,167,168]. Transmission and TB manifestations in some hosts are associated with age and sex risk factors [169]. For example, in white-tailed deer, the odds of being TB positive is greater in males than females [95,147,169], highlighting sex-based differences (possibly hormonal) that may influence the risk of TB.
Whole genome sequencing data of MTBC isolates have shown 7 lineage classifications that have different TB epidemiological consequences worldwide [170,171]. Human-adapted MTBC lineages such as M. africanum and M. tuberculosis sensu stricto are associated with specific geographical locations, variations in virulence, and transmission capacity [172,173,174]. In New Zealand, 782 M. bovis genomes (across 8261 locations) over 30 years were used to track TB outbreaks in domestic and wildlife herds [159], demonstrating the significance of whole genome sequencing in understanding infection and transmission patterns. Heterogenous transmission rates and infectivity of M. bovis are influenced by community and populations with animal movements playing a key role in TB epidemiology [175,176]. Modifications in animal husbandry practices and changes in ecological and population densities are some factors that can influence transmission events among animals as well as increase zoonotic risks to human populations [177]. For example, it has been shown that modern farming, environmental changes from deforestation, settlement, and expansion of agriculture intensifies TB transmission from animals to humans [177].
The epidemiology of TB in domestic animals such as cattle is well documented in the United States [169], the United Kingdom [178,179,180], Iran [181], Italy [154], France [175], and Spain [182]. However, similar studies in wildlife are limited to some reports from wildlife species such as buffaloes [70], lions [46,88,183], and rhinoceros [93,95,184]. Furthermore, in wildlife settings, the epidemiology and pathogenesis of TB in European badgers have been studied extensively in countries like the United Kingdom and Ireland [46,175]. Badgers are considered maintenance hosts due to their longevity and social structures and are implicated in TB transmission to cattle [147]. Studies have shown that M. bovis shed by badgers into the environment can result in transmission to other badgers and cattle (based on genetically related strains) [185,186]. Thus, it is important to fill gaps in epidemiological knowledge on the wide TB host range to improve the detection of infections and mitigate transmission in captive and free-ranging animals.
It has also been demonstrated that domestic animals and wildlife can be an important source of zoonotic pathogens for human populations, particularly in low-income countries [43,187,188,189,190]. Infection with M. bovis can result in zoonotic transmission affecting a wide variety of mammals [115,177,191,192]. The zoonotic risk factors include herding infected animals, working in meat abattoirs [108,193], consumption of unpasteurised milk [18,194,195,196], blood [18], and undercooked or raw meat [18,191,193,197]. The zoonotic transmission of tuberculosis from animals to humans and vice versa is an under-reported problem, with few reports on its magnitude, particularly in developing countries [188,198]. However, in recent years, veterinarians, researchers, the World Health Organisation, and the World Organisation for Animal Health have emphasised improving the knowledge base on TB zoonosis using multidisciplinary One Health approaches [41,196,199,200,201,202,203]. This approach should increase understanding of the effects of TB zoonosis on a global scale.
3. Response of Different Hosts to MTBC Infection?
3.1. Human TB Infection Stages
Characterising different stages of TB infection in humans is an active area of research [204,205], and is typically better understood compared to disease progression in animals [8,120]. A spectrum of clinical stages, ranging from infection clearance, LTB, incipient TB, subclinical TB, and active TB (ATB) disease, has been proposed (Table 2) [32,204]. However, approaches to diagnose, differentiate, and treat these stages are complicated, controversial, and inadequate [20].
Upon infection with pathogenic mycobacteria, there are several potential outcomes. Host defenses may clear the infection; alternatively, the infection may be contained in the form of LTB [146,206], or progress to ATB [204,207]. LTB is defined as an asymptomatic clinical state where individuals are not infectious but may have a risk of progression to active TB disease over time [58]. Characterising the LTB stage in humans is important to prevent disease progression, but it can be difficult to completely eradicate it with existing chemotherapeutic regimens [208]. Therefore, LTB continues to present a global health burden and a major risk factor for the development of ATB disease, particularly in immunocompromised individuals [20,209,210,211].
People with incipient and subclinical TB lack signs and symptoms of disease, while individuals with ATB disease exhibit coughing, night sweats, loss of appetite, weight loss, or yellowish purulent sputum. Symptomatic individuals are a risk to others since they can transmit infectious bacilli in aerosols through speaking, coughing, or sneezing [146,206]. The complex adaptative mechanisms of M. tuberculosis in the host [212], host genetic variation influencing TB susceptibility [28,213,214], host-pathogen interactions [171,215,216,217,218], pathogen-pathogen co-infections [105,219], and pathogen-non-tuberculous mycobacterial co-infections [220,221] further complicate the investigation of infection stages. The factors that determine the stages of infection in humans are still unclear but are believed to involve host immune responses and pathogen adaptation mechanisms [222,223].
The hypothesis that different TB infection stages in humans are associated with different mycobacterial metabolic states has been proposed [204,205]. For instance, LTB has been linked to subpopulations of bacilli that are (i) viable but non or slowly replicating (VBNR) [224,225], (ii) viable but non-culturable (VBNC) [226,227], and (iii) metabolically inactive dormant bacilli [228]. Broadly, these subpopulations are categorised in the reversibly drug-tolerant, but not genetically drug-resistant population, termed “persisters” [227,229,230,231]; as shown in (Figure 1). Persister bacteria appear to be associated with LTB, incipient and subclinical infection [204,232,233], while actively replicating (AR) mycobacterial populations are present in ATB [204]. Even though human TB research has progressed, additional research is needed to fully understand these hypothesised TB stages in humans.
3.2. Domestic and Wildlife Infection Stages of TB
Domestic animals are considered good models to understand TB pathogenesis and stages due to similar physiology, immunology, and anatomy to humans [30,234,235], and thus may be considered to further understand infection stages in other mammals. Transmission may occur after exposure to infectious secretions including saliva, aerosolised respiratory droplets, urine, faeces, exudate from fistulated lymph nodes, and ingestion of infected tissue by scavengers and predators [49,70,190]. The source of infection may influence the development of clinical signs, which can take months to years to appear, and include nonspecific changes, such as weight loss, reproductive failure, decreased milk production, lymphadenopathy, and coughing [6,8]. However, routes of MTBC infection vary in different host species and this may impact TB pathogenesis and influence the development of clinical signs depending on the number of bacterial loads of the source of infection [236]. Thus, the higher the bacterial load from the source of infection (i.e., ingestion of infected tissue from a TB diseased animal) the higher the likelihood of the infected individual developing clinical signs earlier than a source with a lower bacterial load (i.e., infected grass or water or soil) [18,237,238]. Additionally, signs and sites of infection can vary significantly in different species; for example, elbow hygromas and osteomyelitis in lions [46].
The wide range of animal hosts likely has different immune mechanisms, which would result in variable responses to MTBC infection [31,176,239], adding complications to our understanding of different TB stages. For example, elephants [118,240] and suids [241,242] have an early robust humoral response, whereas carnivores such as lions only develop humoral responses in the presence of disease [88,221,243]. Therefore, when conducting research these immune differences must be considered to improve understanding.
Extrapolation of techniques to diagnose human LTB to the study of natural MTBC infections in different species can be difficult due to the logistics of handling large or dangerous animals, long intervals between infection and development of clinical signs of disease, limited ability to identify the source and route of transmission, as well as a paucity of diagnostic tools [6,8,46,49]. The possibility of latent infections in cattle has been discussed in the literature, however, the evidence is not compelling [24,27,244]. Stages of MTBC infections have been hypothesised based on experimental [21,245] and veterinary field experiences with livestock and wildlife (Table 2). These include cleared infection, LTB, subclinical infection, and ATB disease, based on the presence of viable bacteria, detectable immune responses, clinical signs, and radiographic or gross pathological changes [222]. However, additional concrete evidence for these TB stages in animals is needed, which requires finding solutions to the aforementioned challenges and bridging knowledge gaps through research.
3.3. Diagnostic Challenges in Differential Stages of TB
Accurate diagnostic tests are crucial for controlling the spread of TB in humans and domestic, and wild animals [246]. Since animals can be infected with M. bovis (or some other MTBC members), definitive diagnosis relies on identifying the specific MTBC organism [247]. One key problem with the microbiological detection of mycobacteria antemortem is the assumption that the individual is shedding at the time of testing [204]. However, this approach is suboptimal, especially when using the limited antemortem samples available in animals, and often leads to false-negative results [74]. Even if MTBC infection is detected, there is a paucity of clinical and diagnostic tools to differentiate between putative LTB, incipient infection or subclinical infection, and ATB disease (Table 3) [248]. In addition, few studies have investigated differences in virulence and other characteristics of MTBC strains that may influence infection stages in animals [249,250].
Latent TB in humans is diagnosed by detectable antigen-specific immunological responses in the absence of clinical symptoms and evidence of pathological changes (Table 3) [4]. The QuantiFERON TB Gold In-tube Plus interferon-gamma release assay is used in human patients that are asymptomatic, lack radiographic changes, and are sputum smear, culture, or PCR negative, to identify LTB [204,206,222]. However, validated mycobacterial antigen-specific IGRAs are only available for a handful of animal species [12], and radiography is limited by the size of the animal as well as logistics, especially with wildlife [28]. For example, presumptive M. tuberculosis infection in Asian elephants has been diagnosed using serological responses to specific mycobacterial antigens [240]. However, these individuals may be asymptomatic for years before confirming infection by the culture of trunk wash, tracheobronchial lavage fluids, or post-mortem tissue, and the presence of pathological changes [46].
Since existing TB diagnostic tools can not differentiate between stages of TB infection, any positive test for bovine TB (bTB) usually results in the culling of positive animals, regardless of whether clinical signs or lesions are present [35]. In these cases, a post-mortem inspection may aid in determining if possible latent, subclinical, and active infection is present [241,251]. For example, there are reports of cattle with positive immunological assay results but with no evidence of disease, which could be consistent with LTB [26]. These animals lacked gross and histopathological lesions, yet in vivo TST or in vitro blood-based mycobacterial antigen-specific IGRA was positive, although mycobacterial culture results were negative (Table 3) [26,241,252,253]. Often these animals were considered to have false-positive immunological test results rather than the possibility of having LTB. However, other explanations include suboptimal culture detection in paucibacillary samples or false-positive immunologic test results [26].
In summary, many of the tools available to diagnose LTB in humans are unavailable or have not been thoroughly investigated in animals. There is a lack of validated TB diagnostic tests, which often require species-specific approaches [241]. In animals, respiratory samples are rarely submitted for mycobacterial culture and PCR, due to a paucity of veterinary diagnostic laboratories capable of performing these techniques. Therefore, unless there is a suspicion of TB, the lack of diagnostic workup in animals, especially in wildlife, results in a large knowledge gap to demonstrate the possibility of LTB and other possible infection stages.
4. The Influence of MTBC Characteristics on TB Stages
Eleven MTBC ecotypes evolved from M. canettii including M. tuberculosis and M. bovis which are 99.95% genetically similar, and the primary causative agents of TB in humans and animals, respectively [29,249]. MTBC genotyping in different hosts has been studied across Africa including countries like South Africa, Mozambique, Algeria, and Ethiopia with M. bovis being the key aetiological pathogen for animal TB [258].
Despite primarily infecting humans, M. tuberculosis can occasionally infect domestic animals and wildlife [118,259]. Furthermore, Mycobacterium bovis and M. tuberculosis share some similar patterns of transmission and pathogenesis [21,25], however, there are genetic [260,261,262], metabolic [263,264], and physiological differences [249,250] between M. bovis and M. tuberculosis [265], and among M. bovis [266] and M. tuberculosis [267] strains. These differences could affect granuloma formation [23,268], replicative states during infection, and subsequent infection stages in humans and animals.
M. tuberculosis phenotypic heterogeneity has been observed during different in vitro stress conditions such as macrophage exposure and acid stress [214,225,269]. Mouton et al. [225] demonstrated that during macrophage infection, intracellular M. tuberculosis showed a relatively homogeneous population structure at early time points, with a similar replication rate at 24 h to the in vitro-cultured bacteria. Yet, from 48 h onwards, a more diverse population emerged, with a distinct slower-growing population at 96 h, demonstrating the ability of M. tuberculosis to adapt to stress. However, there are relatively few studies on M. bovis phenotypic heterogeneity under in vitro stressors [268]. Therefore, application of in vitro and in vivo stress models, previously utilised for M. tuberculosis, to M. bovis could reveal new insights into replication dynamics and adaptation to stress.
Similar to M. tuberculosis [264,265,269,270,271], in vitro and in vivo studies using M. bovis BCG have investigated replication dynamics under different host stressors [117,121,122,123,124]. Results demonstrated how human alveolar macrophages effectively used nitric oxide production as a mechanism to suppress M. bovis BCG growth [272], similar to observations with M. tuberculosis [273]. In addition, 23 genes likely involved in energy metabolism were downregulated while only a narX gene (encoding nitrate reductase) was upregulated in aerobic dormant BCG [274], suggesting this gene plays a role in dormancy responses, similar to M. tuberculosis. Boon and Dick [275] revealed that the dormancy survival regulator (dosR) dormancy survival regulator gene that encodes for Rv3133c (α-crystallin small heat shock protein), universal stress protein encoded by Rv2623, and cystathionine β-synthase protein encoded by Rv2626c were upregulated in M. bovis BCG during anaerobic conditions. These findings were similar to the behaviour shown by M. tuberculosis, in which these genes induced non-replicating or dormant bacilli. Furthermore, the differential abilities of MTBC species to enter a non-replicative state was demonstrated using a rabbit model whereby extrapulmonary tuberculosis was exclusively observed in M. bovis Ravenel and M. bovis AF2122/97 compared to M. tuberculosis CDC1551 and M. tuberculosis H37Rv [267].
Adaptation to stressors may provide a greater understanding of differences in the ability of MTBC to become dormant. Exposure of M. bovis CRBIP7.106 (isolated from human bone) to cryogenic treatment and nutrient starvation resulted in stress responses including morphological phenotypic adaptation through the formation of rare extreme morphologic forms (L-forms or cell-wall deficient) bacteria [276], likely to be a long-term survival strategy during unfavourable conditions. Similar to M. tuberculosis, Rv3134c/devR/devS in M. bovis BCG Pasteur strain 1137P2 was responsible for adaptation during nutrient starvation, ex vivo macrophages, and oxygen limitation [277], which would be necessary to create an LTB state. Using a nutrient starvation model, the presence and expression of key genes such as (mpb83, mpb70, mmpL8-papA1-pks2 locus, Mb2651) that synthesise or encode cell wall, lipid metabolism, and transcriptional regulators, respectively, differed between M. tuberculosis H37Rv and M. bovis AF2122/97 [260]. For example, a lipid metabolism mmpL8-papA1-pks2 locus was present only in M. tuberculosis. Mpb83 (encoding cell wall lipoprotein), and mpb70 (encoding immunogenic lipoprotein) genes were upregulated in M. bovis compared to M. tuberculosis. In addition, transcriptional regulators showed different patterns with increased expression of Mb2651, Mb2654c, Mb3109c/virS, Mb3477c in M. bovis, and Rv0196, Rv0275c, and Rv2160A-Rv2160c in M. tuberculosis. These observations emphasise the differential way in which the MTBC strains may respond or adapt to stress, thus impacting the ability to result in LTB. Therefore, research into bacterial characteristics that influence host-pathogen interactions and bacterial virulence is essential to elucidate their role in LTB.
Reactivations of M. tuberculosis infection in humans are well documented [226,278,279]. Similarly, there are reports of M. bovis infection reactivations in elderly people in England [280], immunocompromised elderly from Demark [281] and the Netherlands [163], and in bTB eliminated areas (1% of total cases) [282], suggesting a possible capacity of M. bovis to persist in a latent state in humans. In addition, LTB was diagnosed in Mexican abattoir and dairy farm workers infected with M. bovis; 76.2% were TST positive and 58.5% IGRA positive [4]. Additionally, cases of BCG infection (BCG-osis) have been reported weeks to months after BCG vaccination in children, suggesting that M. bovis BCG can enter a non-replicative persister state as well as reactivate [283,284]. However, more studies are needed to establish M. bovis persistence and investigate the relationship between metabolic states of mycobacteria and different infection stages in animals.
5. Candidate Models to Improve Understanding of TB Stages in Animals
5.1. In Vitro Models
Numerous in vitro experiments have been performed to understand LTB in humans, with a relative paucity of corresponding work using M. bovis. In vitro models are pivotal for simulating the microenvironment that mycobacterial pathogens may encounter in human and animal hosts [285,286]. To date, nine variations of hypoxia models have been exploited for M. tuberculosis studies [287], while very few have been applied to M. bovis strains, although attenuated M. bovis BCG strains have been investigated under hypoxic conditions [274]. In one report, M. bovis BCG was found to survive and adapt to hypoxia through upregulation of DosR (Rv3133c), an essential dormancy response regulator also found in M. tuberculosis [275]. Gideon et al. [288] demonstrated upregulation of genes from the regions of difference (RD) 2 (Rv2659c) and RD11 (Rv2658c) in M. tuberculosis during hypoxia [261]. However, RD11 is absent in all M. bovis strains, so the hypoxic adaptive advantage this RD region may confer on M. tuberculosis would be lost in M. bovis. Furthermore, RD2 is absent in M. bovis BCG, so this strain may not fully represent the adaptation of M. bovis virulent strains [261]. Therefore, it is important to shift research focus from M. bovis BCG and commonly applied lab-adapted virulent M. bovis Ravenel and M. bovis AF2122/97 to pathogenic M. bovis strains isolated from animals with natural infections [266]. This approach will build new knowledge on how pathogenic wild-type strains would respond to hypoxia.
Nutrient starvation, acid stress, and multi-stress models that combine single stressors [hypoxia (5% O2 content), high CO2 (10%), nutrient deprivation (10% Dubos medium), and acidic pH (pH 5.0)] [289] have been explored using M. tuberculosis to mimic multiple host-pathogen interactions upon infection [289]. However, the use of these models with M. bovis has been limited. In vitro nutrient starvation of M. bovis BCG induced upregulation of the Rv3134c/devR/devS transcriptional regulator, which is also present in M. tuberculosis [277]. Further, M. bovis BCG was found to be tolerant of low pH through the adoption of a homeostatic balance between the internal and external pH [290]. Another study found that M. bovis BCG was also tolerant to low pH in the presence of glutamate as a carbon source [291]. These results highlight that in an acidic environment, a subpopulation of M. bovis bacilli, which can utilise glutamate as a carbon source, may have the ability to persist longer than those that cannot utilise glutamate. Thus, these in vitro models appear to be a potential platform for additional investigations using field strains of M. bovis.
In vitro macrophage infection models have been widely applied to understand differences in pathogenesis and host tropism in M. bovis, M. tuberculosis [265], Mycobacterium avium subsp. paratuberculosis, and Mycobacterium avium subsp. avium [292]. Bovine alveolar macrophages [265], blood-derived macrophages [293], human monocyte THP-1 [294], and RAW 264.7 macrophages [295,296] have been used to understand mechanisms that M. bovis employs to adapt to stress, host-pathogen interactions, and vaccine discovery. Bovine alveolar macrophages infected with M. bovis BCG and two virulent M. bovis strains (M. bovis Ravenel and M. bovis AF2122/97) have revealed transcriptional and proteomic differences between M. bovis and M. tuberculosis pathogens under acid shock, hypoxia, and macrophage infection [260,297,298,299]. These studies showed that M. tuberculosis expressed higher levels of nitrate reductase-associated genes narG and narH, while M. bovis expressed more mpb70 and mpb83 (the two most upregulated genes of the overall analysis) [260,298]. A cell wall-associated protein, DipZ (member of sigK regulon), that encodes for MPB70 and MPB83, was among the top ten highly expressed proteins in M. bovis compared to M. tuberculosis under stress conditions. Additionally, Rv0188, an enduring hypoxic response antigen, was found to be solely associated with low pathology scores in cattle infected with M. bovis, suggesting that it may play a role in inducing a non-replicative state [300]. These genes and protein expression differences could potentially influence TB stages in humans and animals [27]; however, additional studies are needed to investigate their effects on the development of LTB using diverse hosts [28,301,302] and MTBC strains. Since bovine alveolar macrophages are isolated from cattle, this model may be considered a more physiologically relevant model for M. bovis and thus suitable for understanding the host-pathogen interactions during M. bovis infections.
5.2. In Vivo Models
Cattle are the most commonly used animals for investigating in vivo M. bovis pathogenesis and host-pathogen interactions [25,300,303]. Although cattle are a natural host of M. bovis, they may not recapitulate infection stages in the wide variety of other animal species infected with M. bovis [268,304]. Experiments to study LTB using cattle have a logistical challenge since there are limited antemortem methods to confirm whether pathological changes are present or absent [244]. Cattle experimentally infected with M. bovis often develop clinical signs and progress to disease [268], whereas infections with M. tuberculosis H37Rv or other M. tuberculosis isolates are reported to delay progression to disease [31,120,244]. Therefore, M. bovis dose-response and strain differences in experimental infection models need further exploration.
Domestic pigs have been recently used as a model to understand domestic animals and human TB [30,305,306,307]. Pigs experimentally infected with M. bovis AF2122/97 exhibited greater morbidity, rapid onset mortality, severe pathology, and weight loss, as compared to less severe disease in those infected with M. tuberculosis Erdman [30]. M. tuberculosis-challenged pigs developed the active disease but had delayed onset of fever, no signs of respiratory distress, and showed no change in health status, compared to M. bovis infection, during the 35-week trial [30]. Therefore, the pig model may be useful for exploring whether different M. bovis field strains develop different infection stages compared to the commonly used M. bovis AF2122/97 and M. bovis Ravenel strains.
The badger is a promising in vivo model for studying LTB in animals [21,308,309]. This model mimics observations in naturally M. bovis-infected animals, with up to 80% of infected badgers containing M. bovis for years, until reactivation occurs when their immune system becomes compromised [308,309]. Chronic progressive pulmonary TB occurs in badgers, therefore this model may be a candidate system for not only LTB but also for subclinical infection studies [308].
The Cornell mouse model is considered a “classic” TB latency model, in which infected mice are treated with antibiotics to reduce bacteria to undetectable levels, before TB reactivation [279,310]. This model results in a steady asymptomatic state that resembles human latency in which bacteria enter a slowly or non-replicating state [205,311,312,313]. A study found that mice vaccinated before being challenged developed different immunopathology, with M. bovis BCG challenged mice having lower histopathological scores than those challenged with M. tuberculosis [314]. Although this is not surprising since BCG is attenuated, it raises the question of whether virulent M. bovis strains would differ in their ability to enter a slowly or non-replicating state in this model.
The guinea pig TB model has also been used to study infection stages [315], M. bovis diagnostics [316], and vaccine development [317]. This has been used to study LTB using M. tuberculosis [318] but hasn’t been evaluated using M. bovis clinical strains. Mice and guinea pig models are relatively cheaper than those using large animals, such as cattle and pigs, although they can be technically challenging and present limitations such as lack of granuloma structure and organisation, no extracellular dissemination, and difficulty establishing LTB without vaccinations [222,317,319] Therefore, badgers, cattle, and pigs may be more physiologically relevant to studying the different TB stages in M. bovis natural hosts.
In vivo research is needed to answer questions about the existence of different TB stages in animals. Although cattle, pig, and badger models appear to be the most appropriate models, they do not produce all features of the different TB stages or reflect the diverse host-pathogen repertoire of responses during infection in other domestic animal and wildlife species [314,315,320].
6. Candidate Host and Pathogen Biomarkers to Improve Understanding of TB Stages
6.1. Candidate Host Markers
Currently, there are no specific host immunologic biomarkers definitively associated with latency in humans and animals because most overlap with those for active disease [48,223,285,321], although in vitro and in vivo models have shed light on possible candidates [205]. A quantitative comparison of cytokine levels in the blood of latently infected versus actively diseased patients showed differences in tumor necrosis factor α, IFN-γ, and interleukin 12 (IL-12) [26,48,205]. Transcriptomic profiling of immune responses in a Gambian study found that an antiapoptotic gene bcl2 was a promising marker that indicated the onset of active disease very early after infection [322]. In addition, the study found that the B-cell lymphoma 2 (BCL2) marker was significantly lower in individuals with ATB disease versus latently infected individuals [322], also making it a potential host candidate biomarker. Another study found that IL-13 and autoimmune regulator (AIRE) were biomarkers that could identify high-risk people with LTB that develop ATB, months before clinical diagnosis [323].
Multiple cytokine assays have been evaluated in human studies for their ability to differentiate active disease from LTB [324,325]. Suzukawa et al. [326] found that combined analysis of IFN-γ, IL-2, IL-5, IL-10, IL-1RA, and MCP-1/CCL2 cytokines found in QuantiFERON supernatant, could distinguish ATB from LTB in humans. In a recent study, in vitro antigen-specific concentrations of IFN-γ, IL-2, IL-5, IL-10, IL-1RA, and monocyte chemoattractant protein-1 (MCP-1/CCL2) showed promise for distinguishing ATB from LTB [321,326]. A recent systematic review that compared human host factors involved in LTB, identified IL-1, IL-6, IL-9, IL-17, and IL-IRA as potential candidates for identifying human latency [48].
In contrast, there are relatively few studies on diagnostic host biomarkers in animal TB, with the majority focusing on developing diagnostic tools and investigating pathogenesis [12,21,31,252]. Therefore, additional investigation is needed to confirm whether these can be used to detect latent infection in animals. The development of an LTB cytokine panel would need to consider the variability of MTBC characteristics, including bacterial heterogeneity, host immunological responses, and pathogenesis, across animal species.
6.2. Candidate Pathogen Biomarkers
Several mycobacterial factors are upregulated during M. tuberculosis infection in response to host defences. In human LTB, these include genes that mediate energy metabolism, lipid production, stress response, growth restriction, hypoxia response, and toxin-antitoxin stress gene regulators of the bacilli [205,227,327]. By extrapolating from human LTB to animals, candidate bacterial biomarkers to investigate would be DosR and other transcriptional factors.
The DosR regulon is one of the best-studied regulatory systems in mycobacterial responses to anaerobic conditions [275,328]. DosR is responsible for the maintenance and upkeep of mycobacterial persister populations believed to underlie latency and is conserved in M. tuberculosis [227,329,330,331] and M. bovis [277]. This regulon is a 48-component system controlled by the DosR regulator [270]. DosR genes include nrdZ, narX, narK2, ctpF, otsB1, fdxA, pdxA, pfkB, acr, acg, and dosR, which respond to environmental stress conditions, leading to mycobacterial dormancy and persistence [270,332,333,334]. The DosR antigen Rv2031c (Acr or HspX) has been associated with adaptation to stressful in vitro conditions such as hypoxia [297,335]. Importantly, studies in cattle infected with M. tuberculosis H37Rv and M. bovis AF2122/97 [244] have demonstrated that this antigen may differentiate between active disease and possible LTB in animals.
A recent study has recognised the potential of DosR and resuscitation-promoting factors (Rpf) antigens [Rv2029c (PfkB), Rv2389c (RpfA), Rv0867c (RpfA), and E6-C1] to discriminate between LTB and ATB in a TB endemic human population [330]. Similar results have been observed in other studies conducted in African, Asian, European, Indian, and Brazilian populations [330,331,336], suggesting that DosR antigens are important for latency, independent of host immune responses, environmental background, human genetics, and circulating M. tuberculosis strains. Furthermore, a study by Jones et al. [300] identifies potential LTB markers in naturally infected cattle such as DosR/S/T antigens (Rv2627c, Rv2628, Rv2029c, and Rv1733c). However, the number of animals with no visible lesions was very low, so LTB may have been missed due to the experimental conditions. Therefore, studies with a larger sample size would provide further clarity and a better understanding of LTB biomarkers.
The main driver for latent M. tuberculosis infection is believed to be hypoxia [288], but more recent studies have discovered additional factors that could play a role. Transcription of mRNA sigma factor genes is implicated in different stress responses and may play a role in the adaptation to stressful host environments [337]. These include sigma factor F gene (sigF) (responsible for slowing growth of bacteria) [338,339], and hspX/ acr/ Rv2031c genes that encode for the α-crystallin homolog, and small heat shock protein (sHSP16.3) [340,341]. SigF is upregulated in M. tuberculosis persisters, suggesting a role in latency [332]. Further, the sigF gene is responsible for regulating oxidative stress, antibiotic stress, cold shock, and nutrient depletion [332,337]. Sigma factor B (sigB) and sigH are additional genes induced by heat and oxidative stress [342,343]. Although sigF has been demonstrated to be induced in hypoxic conditions and the stationary growth phase in M. bovis BCG [337,344], there is a need to further investigate transcriptional factors specifically involved in M. bovis persister bacilli.
The tools and methods to study M. tuberculosis persistence have improved in recent years, hence some mycobacterial candidate genes for persistence may overlap in M. tuberculosis and M. bovis strains due to their genetic similarities. These similar genetic characteristics could make it easier to reproduce models using M. bovis or other MTBC species, thus accelerating latency research in animals. Nevertheless, some of the physiological differences between these strains may reveal unique candidate genes for latency upon infection and this must be taken into consideration during experimental design.
7. How Can Researchers Bridge Existing LTB Knowledge Gaps in Animals?
Although laboratory-based research has highlighted the divergent nature of M. tuberculosis and M. bovis infection stages in mice [314], guinea pigs [290], cattle [31], badgers [21], and domestic pigs [307,345], there is still no compelling evidence on whether LTB occurs in domestic and wild animals infected with M. bovis, M. tuberculosis, or other MTBC pathogens. Therefore, there is a need to identify knowledge gaps and possible approaches to address these.
7.1. Factors Contributing to Existing Knowledge Gaps
- A paucity of studies describing and comparing the pathogenesis of M. bovis infection and other MTBC species in different animal species.
- Limited availability of sensitive and specific diagnostic tools for detection and differentiation of MTBC infection stages in domestic animals and wildlife, especially antemortem tests.
- Incomplete information on the diversity of MTBC virulent clinical strains, primarily M. bovis, and their influence on pathogenesis.
- A limited understanding of the role and variability in immune responses to mycobacterial infection in different animal species.
- A poor understanding of the genetic, metabolic, and physiological characteristics of M. bovis could promote persister bacilli formation.
- A lack of clarity on how TB stages vary in different hosts (humans, domestic and wild animals).
- A lack of well-characterised in vitro and in vivo models of M. bovis infection to simulate different stages of infection, including LTB.
7.2. Recommendations for Future Research
To improve understanding of LTB in animals, data from in vitro and in vivo MTBC archetypal strains (such as M. tuberculosis H37Rv, M. tuberculosis CDC1551, M. bovis Ravenel, and M. bovis AF2122/97) and studies on host responses in M. tuberculosis and M. bovis BCG infections should be used to design future research. This should include:
- Developing a consensus on the definition of latency in domestic and wild animals and identifying a model that could be used to find biomarkers for this state.
- Identification of blood-based host and pathogen biomarkers that can differentiate between ATB and different stages of M. bovis infection in different animal species.
- Utilising available tools to study the phenotypic state of persister bacilli at a single-cell level to understand the physiological, phenotypic, and molecular features of different strains.
- Comparing the pathogenesis of M. bovis and other MTBC in different animal species to characterise the chronic asymptomatic state in infected hosts.
- Exploring host–pathogen similarities and differences of host–pathogen interactions to elucidate factors leading to LTB and susceptibility of different species to latent infections.
8. Conclusions
Animal TB research is still lagging compared to human TB research. Reasons include the complexity introduced by the fact that animals can be infected by a wide range of MTBC species, and a range of animal species can be infected by M. bovis [176]. Further, each MTBC species has diverse strains which could have differential influences on their pathogenesis and virulence. There is a lack of compelling evidence to prove the existence of different TB stages in animals as proposed in humans [204]. Experimental cattle [31] and pig studies [30], that applied laboratory-adapted strains, suggest that M. bovis is less likely to enter a persister state than M. tuberculosis. Comparative genetic and proteomic studies of M. bovis and M. tuberculosis have shown metabolic and replicative differences which could impact how these pathogens adapt to different animal hosts [249,250]. Despite these genetic and proteomic differences, reports of M. bovis reactivation in humans suggest that this species is capable of establishing LTB. However, further investigations are needed to ascertain the likelihood of MTBC pathogens (especially M. bovis) establishing LTB in animals and humans [250,267].
Although there is a lack of compelling evidence for latency in cattle [27,160], some ante-mortem and post-mortem findings suggest a possible LTB stage in animals [21,26,131,308]. Latent TB could occur in some animal species but is under-recognised due to the lack of defined criteria for TB stages, the challenging logistics, and the lack of diagnostic tools that can differentiate between several infection stages. However, there are promising approaches to studying human LTB that could serve as a foundation to elucidate this stage in animals [204]. Blood-based test approaches present a convenient way to distinguish TB stages ante-mortem; however, the current biomarkers of LTB and active disease overlap [326]. While post-mortem tests can also be applied to validate LTB in animals they can only be performed upon culling an individual, therefore presenting feasibility and cost implications. These myriad challenges negatively impact the progress of research to determine if LTB occurs in domestic or wild animals, however, using cross-species studies (M. bovis vs. M. tuberculosis) could advance understanding of LTB at the domestic-wildlife-human interfaces.
Identifying TB stages such as LTB in animals can inform policy, devise targeted management, and improve control strategies based on associated risks. Studying LTB in animals could also differentiate individuals that may be infected but not infectious from those with a greater risk of spreading the disease, provide potential management changes that could prevent infected animals from progressing to active disease (and therefore spread) and minimise the loss of animals due to unnecessary culling (especially for endangered animals such as rhinos). In vitro and in vivo models can be applied to broaden knowledge on whether different field MTBC species, particularly M. bovis, induce persister bacilli believed to underlie LTB. When designing such studies, it is important to be aware of the diversity in host-pathogen interactions and infection outcomes in different animals. Moreover, the knowledge gained from animal TB research can also inform models to study LTB in humans. Future research should also focus on methods to further identify persister bacterial populations, not only in culture but also directly in animal samples to characterize their phenotypes, ultimately bridging some LTB knowledge gaps.
Author Contributions
All authors listed have made a substantial, direct, and intellectual contribution to the work, and approved it for publication. P.N. performed conceptualisation, investigation, writing (original draft, review, and editing), and visualisation; B.B. performed conceptualisation, and writing (original draft, review, and editing), W.J.G. performed writing (original draft, review, and editing) and supervision; M.A.M. and S.L.S. performed conceptualisation, funding acquisition, supervision, and writing (original draft, review, and editing). The opinions expressed in this paper are solely those of the authors. All authors have read and agreed to the published version of the manuscript.
Funding
The authors acknowledge Stellenbosch University Faculty of Medicine and Health Science, the South African Medical Research Council (SAMRC), and the National Research Foundation (NRF) South African Research Chairs Initiative (SARChI grant numbers 86949 and 86539) for financial support. The content is the sole responsibility of the authors and does not necessarily represent the official views of the South African Medical Research Council or the National Research Foundation.
Acknowledgments
We also acknowledge constructive discussions and contributions from Jomien Mouton, and Trisha Parbhoo.
Conflicts of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest. The authors declare that there are no conflict of interest.
References
- WHO. Global Tuberculosis Report. 2021. Available online: https://www.who.int/publications/i/item/9789240037021 (accessed on 9 May 2022).
- Tadesse, T.; Lelo, U.; Markos, T.; Tadesse, T.; Birhan, F.; Tona, T. Review on the Epidemiology, Public Health and Economic Importance of Bovine Tuberculosis. J. Biol. Agric. Healthc. 2017, 7, 16–23. [Google Scholar]
- Lesslie, I.W.; Magnus, K.; Stewart, C.J. The Prevalence of Bovine Type Tuberculous Infection in Man in the English Rural Population. Tubercle 1972, 53, 198–204. [Google Scholar] [CrossRef]
- Torres-Gonzalez, P.; Soberanis-Ramos, O.; Martinez-Gamboa, A.; Chavez-Mazari, B.; Barrios-Herrera, M.T.; Torres-Rojas, M.; Cruz-Hervert, L.P.; Garcia-Garcia, L.; Singh, M.; Gonzalez-Aguirre, A.; et al. Prevalence of Latent and Active Tuberculosis among Dairy Farm Workers Exposed to Cattle Infected by Mycobacterium bovis. PLoS Negl. Trop. Dis. 2013, 7, e2177. [Google Scholar] [CrossRef]
- Kanabalan, R.D.; Lee, L.J.; Lee, T.Y.; Chong, P.P.; Hassan, L.; Ismail, R.; Chin, V.K. Human Tuberculosis and Mycobacterium tuberculosis Complex: A Review on Genetic Diversity, Pathogenesis and Omics Approaches in Host Biomarkers Discovery. Microbiol. Res. 2021, 246, 126674. [Google Scholar] [CrossRef]
- Miller, M.A.; Lyashchenko, K.P. 15 Mycobacterial Infections in Other Zoo Animals. In Tuberculosis, Leprosy and Mycobacterial Diseases of Man and Animals: The Many Hosts of Mycobacteria; CABI: London, UK, 2015. [Google Scholar] [CrossRef]
- Zanardi, G.; Boniotti, M.B.; Gaffuri, A.; Casto, B.; Zanoni, M.; Pacciarini, M.L. Tuberculosis Transmission by Mycobacterium bovis in a Mixed Cattle and Goat Herd. Res. Vet. Sci. 2013, 95, 430–433. [Google Scholar] [CrossRef]
- Pesciaroli, M.; Alvarez, J.; Boniotti, M.B.; Cagiola, M.; Di Marco, V.; Marianelli, C.; Pacciarini, M.; Pasquali, P. Tuberculosis in Domestic Animal Species. Res. Vet. Sci. 2014, 97, S78–S85. [Google Scholar] [CrossRef] [PubMed]
- Muñoz Mendoza, M.; de Juan, L.; Menéndez, S.; Ocampo, A.; Mourelo, J.; Sáez, J.L.; Domínguez, L.; Gortázar, C.; García Marín, J.F.; Balseiro, A. Tuberculosis Due to Mycobacterium bovis and Mycobacterium caprae in Sheep. Vet. J. 2012, 191, 267–269. [Google Scholar] [CrossRef]
- Hang’ombe, M.B.; Munyeme, M.; Nakajima, C.; Fukushima, Y.; Suzuki, H.; Matandiko, W.; Ishii, A.; Mweene, A.S.; Suzuki, Y. Mycobacterium bovis Infection at the Interface between Domestic and Wild Animals in Zambia. BMC Vet. Res. 2012, 8, 221. [Google Scholar] [CrossRef]
- Olivier, T.T.; Viljoen, I.M.; Hofmeyr, J.; Hausler, G.A.; Goosen, W.J.; Tordiffe, A.S.W.; Buss, P.; Loxton, A.G.; Warren, R.M.; Miller, M.A.; et al. Development of a Gene Expression Assay for the Diagnosis of Mycobacterium bovis Infection in African Lions (Panthera leo). Transbound. Emerg. Dis. 2017, 64, 774–781. [Google Scholar] [CrossRef]
- Bernitz, N.; Kerr, T.J.; Goosen, W.J.; Chileshe, J.; Higgitt, R.L.; Roos, E.O.; Meiring, C.; Gumbo, R.; de Waal, C.; Clarke, C.; et al. Review of Diagnostic Tests for Detection of Mycobacterium bovis Infection in South African Wildlife. Front. Vet. Sci. 2021, 8, 588697. [Google Scholar] [CrossRef]
- Malone, K.M.; Gordon, S.V. Mycobacterium tuberculosis Complex Members Adapted to Wild and Domestic Animals. Adv. Exp. Med. Biol. 2017, 1019, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Mostowy, S.; Cousins, D.; Behr, M.A. Genomic Interrogation of the Dassie Bacillus Reveals It as a Unique RD1 Mutant within the Mycobacterium tuberculosis Complex. J. Bacteriol. 2004, 186, 104–109. [Google Scholar] [CrossRef] [PubMed]
- Brites, D.; Loiseau, C.; Menardo, F.; Borrell, S.; Boniotti, M.B.; Warren, R.; Dippenaar, A.; Parsons, S.D.C.; Beisel, C.; Behr, M.A.; et al. A New Phylogenetic Framework for the Animal-Adapted Mycobacterium tuberculosis Complex. Front. Microbiol. 2018, 9, 2820. [Google Scholar] [CrossRef] [PubMed]
- Coscolla, M.; Lewin, A.; Metzger, S.; Maetz-Rennsing, K.; Calvignac-Spencer, S.; Nitsche, A.; Dabrowski, P.W.; Radonic, A.; Niemann, S.; Parkhill, J.; et al. Novel Mycobacterium tuberculosis Complex Isolate from a Wild Chimpanzee. Emerg. Infect. Dis. 2013, 19, 969. [Google Scholar] [CrossRef]
- Gummow, B. A Survey of Zoonotic Diseases Contracted by South African Veterinarians. J. S. Afr. Vet. Assoc. 2003, 74, 72–76. [Google Scholar] [CrossRef]
- Sichewo, P.R.; Vander Kelen, C.; Thys, S.; Michel, A.L. Risk Practices for Bovine Tuberculosis Transmission to Cattle and Livestock Farming Communities Living at Wildlife-Livestock-Human Interface in Northern Kwazulu Natal, South Africa. PLoS Negl. Trop. Dis. 2020, 14, 7681. [Google Scholar] [CrossRef]
- Moyo, M.; Lebina, L.; Milovanovic, M.; MacPherson, P.; Michel, A.; Martinson, N. Tuberculosis Patients at the Human-Animal Interface: Potential Zooanthroponotic and Zoonotic Transmission. One Health 2021, 13, 100319. [Google Scholar] [CrossRef]
- Behr, M.A.; Edelstein, P.H.; Ramakrishnan, L. Revisiting the Timetable of Tuberculosis. BMJ 2018, 362, k2738. [Google Scholar] [CrossRef]
- Gormley, E.; Corner, L.A.L. Pathogenesis of Mycobacterium bovis Infection: The Badger Model as a Paradigm for Understanding Tuberculosis in Animals. Front. Vet. Sci. 2018, 4, 247. [Google Scholar] [CrossRef]
- Michel, A.L.; Lane, E.P.; de Klerk-Lorist, L.M.; Hofmeyr, M.; van der Heijden, E.M.D.L.; Botha, L.; van Helden, P.; Miller, M.; Buss, P. Experimental Mycobacterium bovis Infection in Three White Rhinoceroses (Ceratotherium simum): Susceptibility, Clinical and Anatomical Pathology. PLoS ONE 2017, 12, e0179943. [Google Scholar] [CrossRef]
- Palmer, M.V.; Kanipe, C.; Boggiatto, P.M. The Bovine Tuberculoid Granuloma. Pathogens 2022, 11, 61. [Google Scholar] [CrossRef]
- Álvarez, A.H.; Estrada-Chávez, C.; Flores-Valdez, M.A. Molecular Findings and Approaches Spotlighting Mycobacterium bovis Persistence in Cattle. Vet. Res. 2009, 40, 22. [Google Scholar] [CrossRef] [PubMed]
- Pollock, J.M.; Neill, S.D. Mycobacterium bovis Infection and Tuberculosis in Cattle. Vet. J. 2002, 163, 115–127. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, A.H.; Gutiérrez-Ortega, A.; Gómez-Entzin, V.; Pérez-Mayorga, G.; Naranjo-Bastién, J.; González-Martínez, V.; Milián-Suazo, F.; Martínez-Velázquez, M.; Herrera-Rodríguez, S.; Hinojoza-Loza, E. Assessment of Antigenic Supplementation of Bovine Purified Protein Derivative for Diagnosis of Subclinical Infection with Mycobacterium bovis in Cattle. Microb. Pathog. 2017, 108, 114–121. [Google Scholar] [CrossRef] [PubMed]
- García, J.S.Y.; Bigi, M.M.; Klepp, L.I.; García, E.A.; Blanco, F.C.; Bigi, F. Does Mycobacterium bovis Persist in Cattle in a Non-Replicative Latent State as Mycobacterium tuberculosis in Human Beings? Vet. Microbiol. 2020, 247, 108758. [Google Scholar] [CrossRef] [PubMed]
- Mukundan, H.; Chambers, M.; Waters, R.; Larsen, M. Tuberculosis, Leprosy and Mycobacterial Diseases of Man and Animals: The Many Hosts of Mycobacteria; CABI: London, UK, 2015. [Google Scholar] [CrossRef]
- Brosch, R.; Gordon, S.V.; Marmiesse, M.; Brodin, P.; Buchrieser, C.; Eiglmeier, K.; Garnier, T.; Gutierrez, C.; Hewinson, G.; Kremer, K.; et al. A New Evolutionary Scenario for the Mycobacterium tuberculosis Complex. Proc. Natl. Acad. Sci. USA 2002, 99, 3684–3689. [Google Scholar] [CrossRef]
- Niroula, N.; Lim, Z.L.; Walker, S.; Huang, Y.; Gerdts, V.; Zriba, S.; Drever, K.; Chen, J.M. Domestic Pigs Experimentally Infected with Mycobacterium bovis and Mycobacterium tuberculosis Exhibit Different Disease Outcomes. Tuberculosis 2022, 133, 102167. [Google Scholar] [CrossRef]
- Villarreal-Ramos, B.; Berg, S.; Whelan, A.; Holbert, S.; Carreras, F.; Salguero, F.J.; Khatri, B.L.; Malone, K.; Rue-Albrecht, K.; Shaughnessy, R.; et al. Experimental Infection of Cattle with Mycobacterium tuberculosis Isolates Shows the Attenuation of the Human Tubercle Bacillus for Cattle. Sci. Rep. 2018, 8, 894. [Google Scholar] [CrossRef]
- Barry, C.E.; Boshoff, H.; Dartois, V.; Dick, T.; Ehrt, S.; Flynn, J.; Schnappinger, D.; Wilkinson, R.J.; Young, D. The Spectrum of Latent Tuberculosis: Rethinking the Goals of Prophylaxis. Nat. Rev. Microbiol. 2009, 7, 845–855. [Google Scholar] [CrossRef]
- Jenkins, H.E.; Woodroffe, R.; Donnelly, C.A.; Cox, D.R.; Johnston, W.T.; Bourne, F.J.; Cheeseman, C.L.; Clifton-Hadley, R.S.; Gettinby, G.; Gilks, P.; et al. Effects of Culling on Spatial Associations of Mycobacterium bovis Infections in Badgers and Cattle. J. Appl. Ecol. 2007, 44, 897–908. [Google Scholar] [CrossRef]
- Van Tonder, A.J.; Thornton, M.J.; Conlan, A.J.K.; Jolley, K.A.; Goolding, L.; Mitchell, A.P.; Dale, J.; Palkopoulou, E.; Hogarth, P.J.; Hewinson, R.G.; et al. Inferring Mycobacterium bovis Transmission between Cattle and Badgers Using Isolates from the Randomised Badger Culling Trial. PLoS Pathog. 2021, 17, e1010075. [Google Scholar] [CrossRef] [PubMed]
- Gormley, E.; Corner, L.A.L. Wild Animal Tuberculosis: Stakeholder Value Systems and Management of Disease. Front. Vet. Sci. 2018, 5, 327. [Google Scholar] [CrossRef] [PubMed]
- Smith, R.L.; Tauer, L.W.; Schukken, Y.H.; Lu, Z.; Grohn, Y.T. Minimization of Bovine Tuberculosis Control Costs in US Dairy Herds. Prev. Vet. Med. 2013, 112, 266–275. [Google Scholar] [CrossRef]
- Karolemeas, K.; de la Rua-Domenech, R.; Cooper, R.; Goodchild, A.V.; Clifton-Hadley, R.S.; Conlan, A.J.K.; Mitchell, A.P.; Hewinson, R.G.; Donnelly, C.A.; Wood, J.L.N.; et al. Estimation of the Relative Sensitivity of the Comparative Tuberculin Skin Test in Tuberculous Cattle Herds Subjected to Depopulation. PLoS ONE 2012, 7, e43217. [Google Scholar] [CrossRef]
- Smith, R.L.; YH, S.; Lu, Z.; RM, M.; YT, G. Development of a Model to Simulate Infection Dynamics of Mycobacterium bovis in Cattle Herds in the United States. J. Am. Vet. Med. Assoc. 2013, 243, 411–423. [Google Scholar] [CrossRef]
- Miller, M.A.; Buss, P.; Parsons, S.D.C.; Roos, E.; Chileshe, J.; Goosen, W.J.; van Schalkwyk, L.; de Klerk-Lorist, L.M.; Hofmeyr, M.; Hausler, G.; et al. Conservation of White Rhinoceroses Threatened by Bovine Tuberculosis, South Africa, 2016–2017. Emerg. Infect. Dis. 2018, 24, 2373–2375. [Google Scholar] [CrossRef] [PubMed]
- Michel, A.L.; Bengis, R.G.; Keet, D.F.; Hofmeyr, M.; De Klerk, L.M.; Cross, P.C.; Jolles, A.E.; Cooper, D.; Whyte, I.J.; Buss, P.; et al. Wildlife Tuberculosis in South African Conservation Areas: Implications and Challenges. Vet. Microbiol. 2006, 112, 91–100. [Google Scholar] [CrossRef]
- Gompo, T.R.; Shrestha, A.; Ranjit, E.; Gautam, B.; Ale, K.; Shrestha, S.; Bhatta, D.D. Risk Factors of Tuberculosis in Human and Its Association with Cattle TB in Nepal: A One Health Approach. One Health 2020, 10, 100156. [Google Scholar] [CrossRef]
- Silva, M.R.; Rocha, A.D.S.; Araújo, F.R.; Fonseca-Júnior, A.A.; de Alencar, A.P.; Suffys, P.N.; da Costa, R.R.; Moreira, M.A.S.; Guimarães, M.D.C. Risk Factors for Human Mycobacterium bovis Infections in an Urban Area of Brazil. Mem. Inst. Oswaldo Cruz 2018, 113, e170445. [Google Scholar] [CrossRef] [Green Version]
- Sichewo, P.R.; Michel, A.L.; Musoke, J.; Etter, E.M.C. Risk Factors for Zoonotic Tuberculosis at the Wildlife–Livestock–Human Interface in South Africa. Pathogens 2019, 8, 101. [Google Scholar] [CrossRef]
- Bernitz, N.; Clarke, C.; Roos, E.O.; Goosen, W.J.; Cooper, D.; van Helden, P.D.; Parsons, S.D.C.; Miller, M.A. Detection of Mycobacterium bovis Infection in African Buffaloes (Syncerus caffer) Using QuantiFERON®-TB Gold (QFT) Tubes and the Qiagen Cattletype® IFN-Gamma ELISA. Vet. Immunol. Immunopathol. 2018, 196, 48–52. [Google Scholar] [CrossRef] [PubMed]
- Clarke, C.; Cooper, D.; Goosen, W.J.; McFadyen, R.; Warren, R.M.; van Helden, P.D.; Parsons, S.D.C.; Miller, M.A. Antigen-Specific Interferon-Gamma Release Is Decreased Following the Single Intradermal Comparative Cervical Skin Test in African Buffaloes (Syncerus caffer). Vet. Immunol. Immunopathol. 2018, 201, 12–15. [Google Scholar] [CrossRef]
- Miller, M.; Buss, P.; Hofmeyr, J.; Olea-Popelka, F.; Parsons, S.; van Helden, P. Antemortem Diagnosis of Mycobacterium bovis Infection in Free-Ranging African Lions (Panthera Leo) and Implications for Transmission. J. Wildl. Dis. 2015, 51, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Van Der Merwe, P.; Saayman, M. Determining the Economic Value of Game Farm Tourism. Koedoe 2003, 46, 103–112. [Google Scholar] [CrossRef]
- Herrera, M.; Vera, C.; Keynan, Y.; Rueda, Z.V. Gaps in Study Design for Immune Parameter Research for Latent Tuberculosis Infection: A Systematic Review. J. Immunol. Res. 2020, 2020, 8074183. [Google Scholar] [CrossRef]
- Miller, M. Tuberculosis in South African Wildlife: Why Is It Important? 2015. Available online: http://www.sun.ac.za/english/Inaugurallectures/Inaugurallectures/InauguralLectureProfMiller.pdf (accessed on 17 January 2019).
- Siddique, A.B.; Hussain, R.; Jamal, A.; Hossain, M.B.; Ahmad, Z.; Mansoor, M.K.; Khan, I.; Zahra, K.; Khan, A. Histopathological Investigations and Molecular Confirmation Reveal Mycobacterium bovis in One-Horned Rhinoceros (Rhinoceros Unicorns). BioMed Res. Int. 2022, 2022, 5816986. [Google Scholar] [CrossRef]
- Alvarez, A.H. Revisiting Tuberculosis Screening: An Insight to Complementary Diagnosis and Prospective Molecular Approaches for the Recognition of the Dormant TB Infection in Human and Cattle Hosts. Microbiol. Res. 2021, 252, 126853. [Google Scholar] [CrossRef] [PubMed]
- Lopes, B.C.; dos Reis, E.M.; de Bitencourt, F.B.R.; Loiko, M.R.; Bezerra, A.V.A.; Bueno, T.S.; Lape, I.T.; Cerva, C.; Coppola, M.D.M.; Rodrigues, R.O.; et al. A Molecular Strategy to Optimize Bovine Tuberculosis Post-Mortem Diagnosis and the Exposure to Mycobacterium tuberculosis Variant Bovis. Mol. Biol. Rep. 2020, 47, 7291–7296. [Google Scholar] [CrossRef]
- Meurens, F.; Dunoyer, C.; Fourichon, C.; Gerdts, V.; Haddad, N.; Kortekaas, J.; Lewandowska, M.; Monchatre-Leroy, E.; Summerfield, A.; Wichgers Schreur, P.J.; et al. Animal Board Invited Review: Risks of Zoonotic Disease Emergence at the Interface of Wildlife and Livestock Systems. Animal 2021, 15, 100241. [Google Scholar] [CrossRef]
- Odoi, A.; Chavez-Lindell, T.L.; Moncayo, A.L.; Fernanda, M.; Veloz, V. An Exploratory Assessment of Human and Animal Health Concerns of Smallholder Farmers in Rural Communities of Chimborazo, Ecuador. PeerJ 2022, 9, e12208. [Google Scholar] [CrossRef]
- Hlokwe, T.M.; Jenkins, A.O.; Streicher, E.M.; Venter, E.H.; Cooper, D.; Godfroid, J.; Michel, A.L. Molecular Characterisation of Mycobacterium bovis Isolated from African Buffaloes (Syncerus Caffer) in Hluhluwe-IMfolozi Park in KwaZulu-Natal, South Africa. Onderstepoort J. Vet. Res. 2011, 78, 6. [Google Scholar] [CrossRef] [PubMed]
- Roug, A.; Muse, E.A.; Clifford, D.L.; Paul, G.; Mpanduji, D.; Makingi, G.; Magesa, W.; Josephat, E.; Mazet, J.; Bird, B.; et al. Health of African Buffalos (Syncerus caffer) in Ruaha National Park, Tanzania. J. Wildl. Dis. 2020, 56, 495–498. [Google Scholar] [CrossRef]
- Shah, Y.; Paudel, S. Protect Elephants from Tuberculosis. Science 2021, 374, 832–833. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Finnegan, M.; Storms, T.; Garner, M.; Lyashchenko, K.P. Outbreak of Mycobacterium tuberculosis in a Herd of Captive Asian Elephants (Elephas maximus): Antemortem Diagnosis, Treatment, and Lessons Learned. J. Zoo Wildl. Med. 2018, 49, 748–754. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.A.; Hogan, J.N.; Meehan, C.L. Housing and Demographic Risk Factors Impacting Foot and Musculoskeletal Health in African Elephants [Loxodonta africana] and Asian Elephants [Elephas Maximus] in North American Zoos. PLoS ONE 2016, 11, e0155223. [Google Scholar] [CrossRef]
- Glaeser, S.S.; Edwards, K.L.; Paris, S.; Scarlata, C.; Lee, B.; Wielebnowski, N.; Finnell, S.; Somgird, C.; Brown, J.L. Characterization of Longitudinal Testosterone, Cortisol, and Musth in Male Asian Elephants (Elephas maximus), Effects of Aging, and Adrenal Responses to Social Changes and Health Events. Animals 2022, 12, 1332. [Google Scholar] [CrossRef] [PubMed]
- Ruetten, M.; Steinmetz, H.W.; Thiersch, M.; Kik, M.; Vaughan, L.; Altamura, S.; Muckenthaler, M.U.; Gassmann, M. Iron Regulation in Elderly Asian Elephants (Elephas maximus) Chronically Infected With Mycobacterium tuberculosis. Front. Vet. Sci. 2020, 7, 596379. [Google Scholar] [CrossRef]
- Mikota, S.K.; Gairhe, K.; Giri, K.; Hamilton, K.; Miller, M.; Paudel, S.; Lyashchenko, K.; Larsen, R.S.; Payeur, J.B.; Waters, W.R.; et al. Tuberculosis Surveillance of Elephants (Elephas maximus) in Nepal at the Captive-Wild Interface. Eur. J. Wildl. Res. 2015, 61, 221–229. [Google Scholar] [CrossRef]
- Katale, B.Z.; Mbugi, E.V.; Kendal, S.; Fyumagwa, R.D.; Kibiki, G.S.; Godfrey-Faussett, P.; Keyyu, J.D.; van Helden, P.; Matee, M.I. Bovine Tuberculosis at the Human-Livestock-Wildlife Interface: Is It a Public Health Problem in Tanzania?: A Review. Onderstepoort J. Vet. Res. 2012, 79, 8. [Google Scholar] [CrossRef] [PubMed]
- Ayele, W.; Neill, S.; Zinsstag, J.; Weiss, M.; Pavlik, I. Bovine Tuberculosis: An Old Disease but a New Threat to Africa. Int. J. Tuberc. Lung Dis. 2004, 8, 924–937. [Google Scholar] [CrossRef]
- Kaneene, J.B.; Miller, R.; de Kantor, I.N.; Thoen, C.O. Tuberculosis in Wild Animals. Int. J. Tuberc. Lung Dis. 2010, 14, 1508–1512. [Google Scholar] [PubMed]
- Fitzgerald, S.D.; Kaneene, J.B. Wildlife Reservoirs of Bovine Tuberculosis Worldwide: Hosts, Pathology, Surveillance, and Control. Vet. Pathol. 2013, 50, 488–499. [Google Scholar] [CrossRef]
- Goodchild, A.V.; Clifton-Hadley, R.S. Cattle-to-Cattle Transmission of Mycobacterium bovis. Tuberculosis 2001, 81, 23–41. [Google Scholar] [CrossRef] [PubMed]
- Sanou, A.; Tarnagda, Z.; Kanyala, E.; Zingué, D.; Nouctara, M.; Ganamé, Z.; Combary, A.; Hien, H.; Dembele, M.; Kabore, A.; et al. Mycobacterium bovis in Burkina Faso: Epidemiologic and Genetic Links between Human and Cattle Isolates. PLoS Negl. Trop. Dis. 2014, 8, 3142. [Google Scholar] [CrossRef] [PubMed]
- Neill, S.D.; Cassidy, J.; Hanna, J.; Mackie, D.P.; Pollock, J.M.; Clements, A.; Walton, E.; Bryson, D.G. Detection of Mycobacterium bovis Infection in Skin Test-Negative Cattle with an Assay for Bovine Interferon-Gamma. Vet. Rec. 1994, 135, 134–135. [Google Scholar] [CrossRef]
- De Vos, V.; Bengis, R.G.; Kriek, N.P.; Michel, A.; Keet, D.F.; Raath, J.P.; Huchzermeyer, H.F. The Epidemiology of Tuberculosis in Free-Ranging African Buffalo (Syncerus caffer) in the Kruger National Park, South Africa. Onderstepoort J. Vet. Res. 2001, 68, 119–130. [Google Scholar]
- Keet, D.F.; Kriek, N.P.J.; Bengis, R.G.; Grobler, D.G.; Michel, A. The Rise and Fall of Tuberculosis in a Free-Ranging Chacma Baboon Troop in the Kruger National Park. Onderstepoort J. Vet. Res. 2000, 67, 115–122. [Google Scholar]
- Keet, D.F.; Kriek, N.P.J.; Bengis, R.G.; Michel, A.L. Tuberculosis in Kudus (Tragelaphus strepsiceros) in the Kruger National Park. Onderstepoort J. Vet. Res. 2001, 68, 225–230. [Google Scholar]
- Paine, R.; Martinaglia, G. Tuberculosis in Wild Buck Living Under Natural Conditions. J. Comp. Pathol. Ther. 1929, 42, 1–8. [Google Scholar] [CrossRef]
- Goosen, W.J.; Kleynhans, L.; Kerr, T.J.; van Helden, P.D.; Buss, P.; Warren, R.M.; Miller, M.A. Improved Detection of Mycobacterium tuberculosis and M. Bovis in African Wildlife Samples Using Cationic Peptide Decontamination and Mycobacterial Culture Supplementation. J. Vet. Diagnostic Investig. 2022, 34, 61–67. [Google Scholar] [CrossRef]
- Hlokwe, T.M.; Id, O.; Article, O. First Detection of Mycobacterium bovis Infection in Giraffe (Giraffa camelopardalis) in the Greater Kruger National Park Complex: Role and Implications. Transbound Emerg. Dis. 2019, 66, 2264–2270. [Google Scholar] [CrossRef]
- Gavier-Widen, D.; Chambers, M.A.; Palmer, N.; Newell, D.G.; Hewinson, R.G. Pathology of Natural Mycobacterium bovis Infection in European Badgers (Meles meles) and Its Relationship with Bacterial Excretion. Vet. Rec. 2001, 148, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Martin, L.E.R.; Byrne, A.W.; O’Keeffe, J.; Miller, M.A.; Olea-Popelka, F.J. Weather Influences Trapping Success for Tuberculosis Management in European Badgers (Meles meles). Eur. J. Wildl. Res. 2017, 63, 30. [Google Scholar] [CrossRef]
- Jolma, E.R.; Delahay, R.J.; Smith, F.; Drewe, J.A. Serologic Responses Correlate with Current but Not Future Bacterial Shedding in Badgers Naturally Infected with Mycobacterium bovis. Transbound. Emerg. Dis. 2021, 69, 1922–1932. [Google Scholar] [CrossRef] [PubMed]
- Infantes-Lorenzo, J.A.; Dave, D.; Moreno, I.; Anderson, P.; Lesellier, S.; Gormley, E.; Dominguez, L.; Balseiro, A.; Gortázar, C.; Dominguez, M.; et al. New Serological Platform for Detecting Antibodies against Mycobacterium tuberculosis Complex in European Badgers. Vet. Med. Sci. 2019, 5, 61–69. [Google Scholar] [CrossRef]
- Katale, B.Z.; Mbugi, E.V.; Siame, K.K.; Keyyu, J.D.; Kendall, S.; Kazwala, R.R.; Dockrell, H.M.; Fyumagwa, R.D.; Michel, A.L.; Rweyemamu, M.; et al. Isolation and Potential for Transmission of Mycobacterium bovis at Human–Livestock–Wildlife Interface of the Serengeti Ecosystem, Northern Tanzania. Transbound. Emerg. Dis. 2017, 64, 815–825. [Google Scholar] [CrossRef]
- Michel, A.L.; Coetzee, M.L.; Keet, D.F.; Maré, L.; Warren, R.; Cooper, D.; Bengis, R.G.; Kremer, K.; van Helden, P. Molecular Epidemiology of Mycobacterium bovis Isolates from Free-Ranging Wildlife in South African Game Reserves. Vet. Microbiol. 2009, 133, 335–343. [Google Scholar] [CrossRef]
- Bouts, T.; Vordermeier, M.; Flach, E.; Routh, A. Positive Skin and Serologic Test Results of Diagnostic Assays for Bovine Tuberculosis and Subsequent Isolation of Mycobacterium interjectum in a Pygmy Hippopotamus (Hexaprotodon liberiensis). J. Zoo Wildl. Med. 2009, 40, 536–542. [Google Scholar] [CrossRef]
- Kerr, T.J.; Goosen, W.J.; Gumbo, R.; de Klerk-Lorist, L.M.; Pretorius, O.; Buss, P.E.; Kleynhans, L.; Lyashchenko, K.P.; Warren, R.M.; van Helden, P.D.; et al. Diagnosis of Mycobacterium bovis Infection in Free-Ranging Common Hippopotamus (Hippopotamus amphibius). Transbound. Emerg. Dis. 2022, 69, 378–384. [Google Scholar] [CrossRef]
- De Vos, V.; McCully, R.M.; van Niekerk, C.A.W.J. Mycobacteriosis in the Kruger National Park. Koedoe 1977, 20, a928. [Google Scholar] [CrossRef]
- Tanner, M.; Michel, A.L. Investigation of the Viability of Mycobacterium bovis under Different Environmental Conditions in the Kruger National Park. Onderstepoort J. Vet. Res. 1999, 66, 185–190. [Google Scholar]
- Hlokwe, T.M.; Mogano, R.M. Utility of Xpert® MTB/RIF Ultra Assay in the Rapid Diagnosis of Bovine Tuberculosis in Wildlife and Livestock Animals from South Africa. Prev. Vet. Med. 2020, 177, 104980. [Google Scholar] [CrossRef]
- Palmer, M.V. Mycobacterium bovis: Characteristics of Wildlife Reservoir Hosts. Transbound. Emerg. Dis. 2013, 60 (Suppl. S1), 1–13. [Google Scholar] [CrossRef]
- Miller, M.A.; Buss, P.; Sylvester, T.T.; Lyashchenko, K.P.; Deklerk-Lorist, L.-M.; Bengis, R.; Hofmeyr, M.; Hofmeyr, J.; Mathebula, N.; Hausler, G.; et al. Mycobacterium bovis in Free-Ranging Lions (Panthera leo)—Evaluation of Serological and Tuberculin Skin Tests for Detection of Infection and Disease. J. Zoo Wildl. Med. 2019, 50, 7. [Google Scholar] [CrossRef] [PubMed]
- Gumbo, R.; Sylvester, T.T.; Goosen, W.J.; Buss, P.E.; de Klerk-Lorist, L.-M.; van Schalkwyk, O.L.; McCall, A.; Warren, R.M.; van Helden, P.D.; Miller, M.A.; et al. Adaptation and Diagnostic Potential of a Commercial Cat Interferon Gamma Release Assay for the Detection of Mycobacterium bovis Infection in African Lions (Panthera leo). Pathogens 2022, 11, 765. [Google Scholar] [CrossRef] [PubMed]
- Brüns, A.C.; Brüns, B.; Tanner, M.; Williams, M.C.; Botha, L.; O’brien, A.; Fosgate, G.T.; Van Helden, P.D.; Clarke, J.; Michel, A.L. Diagnosis and Implications of Mycobacterium bovis Infection in Banded Mongooses (Mungos mungo) in the Kruger National Park, South Africa. J. Wildl. Dis. 2017, 53, 19–29. [Google Scholar] [CrossRef]
- Palmer, M.V.; Thacker, T.C.; Rabideau, M.M.; Jones, G.J.; Kanipe, C.; Vordermeier, H.M.; Waters, W.R. Veterinary Immunology and Immunopathology Biomarkers of Cell-Mediated Immunity to Bovine Tuberculosis. Vet. Immunol. Immunopathol. 2020, 220, 109988. [Google Scholar] [CrossRef]
- Thomas, J.; Balseiro, A.; Gortázar, C.; Risalde, M.A. Diagnosis of Tuberculosis in Wildlife: A Systematic Review. Vet. Res. 2021, 52, 31. [Google Scholar] [CrossRef]
- Miller, M.; Chavey, P.S.; Hofmeyr, J.; Mathebula, N.; Doering, A.; Buss, P.; Olea-Popelka, F. Evaluation of Serum Ferritin and Serum Iron in Free-Ranging Black Rhinoceros (Diceros bicornis) as a Tool to Understand Factors Affecting Iron-Overload Disorder. J. Zoo Wildl. Med. 2016, 47, 820–826. [Google Scholar] [CrossRef]
- Miller, M.A.; Greenwald, R.; Lyashchenko, K.P. Potential for Serodiagnosis of Tuberculosis in Black Rhinoceros (Diceros bicornis). J. Zoo Wildl. Med. 2015, 46, 100–104. [Google Scholar] [CrossRef]
- Dwyer, R.A.; Witte, C.; Buss, P.; Goosen, W.J.; Miller, M.; Caron, A. Epidemiology of Tuberculosis in Multi-Host Wildlife Systems: Implications for Black (Diceros bicornis) and White (Ceratotherium simum) Rhinoceros. Front. Vet. Sci. 2020, 7, 580476. [Google Scholar] [CrossRef] [PubMed]
- Richomme, C.; Réveillaud, E.; Moyen, J.-L.; Sabatier, P.; De Cruz, K.; Michelet, L.; Boschiroli, M.L. Mycobacterium bovis Infection in Red Foxes in Four Animal Tuberculosis Endemic Areas in France. Microorganisms 2020, 8, 1070. [Google Scholar] [CrossRef]
- Roos, E.O.; Buss, P.; de Klerk-Lorist, L.M.; Hewlett, J.; Hausler, G.A.; Rossouw, L.; McCall, A.J.; Cooper, D.; van Helden, P.D.; Parsons, S.D.C.; et al. Test Performance of Three Serological Assays for the Detection of Mycobacterium bovis Infection in Common Warthogs (Phacochoerus africanus). Vet. Immunol. Immunopathol. 2016, 182, 79–84. [Google Scholar] [CrossRef] [PubMed]
- Roos, E.O.; Olea-Popelka, F.; Buss, P.; Hausler, G.A.; Warren, R.; Van Helden, P.D.; Parsons, S.D.C.; De Klerk-Lorist, L.M.; Miller, M.A. Measuring Antigen-Specific Responses in Mycobacterium bovis-Infected Warthogs (Phacochoerus africanus) Using the Intradermal Tuberculin Test. BMC Vet. Res. 2018, 14, 360. [Google Scholar] [CrossRef] [PubMed]
- Roos, E.O.; Olea-Popelka, F.; Buss, P.; de Klerk-Lorist, L.M.; Cooper, D.; van Helden, P.D.; Parsons, S.D.C.; Miller, M.A. Seroprevalence of Mycobacterium bovis Infection in Warthogs (Phacochoerus africanus) in Bovine Tuberculosis-Endemic Regions of South Africa. Transbound. Emerg. Dis. 2018, 65, 1182–1189. [Google Scholar] [CrossRef]
- Campana, M.G.; Parker, L.D.; Hawkins, M.T.R.; Young, H.S.; Helgen, K.M.; Szykman Gunther, M.; Woodroffe, R.; Maldonado, J.E.; Fleischer, R.C. Genome Sequence, Population History, and Pelage Genetics of the Endangered African Wild Dog (Lycaon pictus). BMC Genom. 2016, 17, 1. [Google Scholar] [CrossRef] [PubMed]
- Meiring, C.; Schurz, H.; van Helden, P.; Hoal, E.; Tromp, G.; Kinnear, C.; Kleynhans, L.; Glanzmann, B.; van Schalkwyk, L.; Miller, M.; et al. African Wild Dogs (Lycaon pictus) from the Kruger National Park, South Africa Are Currently Not Inbred but Have Low Genomic Diversity. Sci. Rep. 2022, 12, 14979. [Google Scholar] [CrossRef]
- Overton, J.M.C.; Elizalde Castells, D.; Figueira Fernandes Elizalde, S.R.; Valério, H.M.; Alexandre Zumbo, M.N.; Groom, R.J.; Durant, S.M. Endangered African Wild Dogs (Lycaon pictus Temm.) in Angola: Filling a 50-Year Gap of Knowledge with Findings from Two National Parks. Afr. J. Ecol. 2020, 58, 582–587. [Google Scholar] [CrossRef]
- Higgitt, R.L.; Van Schalkwyk, O.L.; De Klerk-Lorist, L.M.; Buss, P.E.; Caldwell, P.; Rossouw, L.; Manamela, T.; Hausler, G.A.; Hewlett, J.; Mitchell, E.P.; et al. Mycobacterium bovis Infection in African Wild Dogs, Kruger National Park, South Africa. Emerg. Infect. Dis. 2019, 25, 1425–1427. [Google Scholar] [CrossRef]
- Kock, R.A.; Woodford, M.H.; Rossiter, P.B. Disease Risks Associated with the Translocation of Wildlife. OIE Rev. Sci. Tech. 2010, 29, 329–350. [Google Scholar] [CrossRef]
- Hlokwe, T.M.; van Helden, P.; Michel, A.L. Evidence of Increasing Intra and Inter-Species Transmission of Mycobacterium bovis in South Africa: Are We Losing the Battle? Prev. Vet. Med. 2014, 115, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Miller, M.; Michel, A.; van Helden, P.; Buss, P. Tuberculosis in Rhinoceros: An Underrecognized Threat? Transbound. Emerg. Dis. 2017, 64, 1071–1078. [Google Scholar] [CrossRef] [PubMed]
- Monies, B.; de la Rua, R.; Jahans, K. Bovine Tuberculosis in Cats. Vet. Rec. 2006, 158, 490–491. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, K.; Chauhan, D.S.; Gupta, P.; Singh, H.B.; Sharma, V.D.; Yadav, V.S.; Sreekumaran; Thakral, S.S.; Dharamdheeran, J.S.; Nigam, P.; et al. Isolation of Mycobacterium bovis & Mycobacterium tuberculosis from Cattle of Some Farms in North India-Possible Relevance in Human Health. Indian J. Med. Res. 2008, 128, 26–31. [Google Scholar]
- Sweetline Anne, N.; Ronald, B.S.M.; Kumar, T.M.A.S.; Kannan, P.; Thangavelu, A. Molecular Identification of Mycobacterium tuberculosis in Cattle. Vet. Microbiol. 2017, 198, 81–87. [Google Scholar] [CrossRef]
- Ghodbane, R.; Drancourt, M. Non-Human Sources of Mycobacterium tuberculosis. Tuberculosis 2013, 93, 589–595. [Google Scholar] [CrossRef]
- Ravindran, R.; Krishnan, V.V.; Dhawan, R.; Wunderlich, M.L.; Lerche, N.W.; Flynn, J.L.; Luciw, P.A.; Khan, I.H. Plasma Antibody Profiles in Non-Human Primate Tuberculosis. J. Med. Primatol. 2014, 43, 59–71. [Google Scholar] [CrossRef]
- Parsons, S.D.C.; Warren, R.M.; Ottenhoff, T.H.M.; Gey van Pittius, N.C.; van Helden, P.D. Detection of Mycobacterium tuberculosis Infection in Dogs in a High-Risk Setting. Res. Vet. Sci. 2012, 92, 414–419. [Google Scholar] [CrossRef]
- Haydock, L.A.J.; Abrams-Ogg, A.C.G.; Weese, J.S.; Goldstein, M.R.; Clifford, A.B.; Sebastian, A.; Rea, E.H.; Jamieson, F.B.; Duncan, C.; Andrievskaia, O.; et al. Diagnostic and Public Health Investigation of Mycobacterium tuberculosis Infection in a Dog in Ontario, Canada. J. Vet. Diagn. Investig. 2022, 34, 292–297. [Google Scholar] [CrossRef]
- Marfil, M.J.; Barandiaran, S.; Zumárraga, M.J.; Germani, L.; Faccini, T.; Linares, M.; Capra, S.; Gramajo, L.; Martínez Vivot, M.; Falzoni, E. Mycobacterium tuberculosis Infection in a Free-Ranging Urban Dog from Argentina. Vet. Res. Commun. 2022, 46, 781–788. [Google Scholar] [CrossRef]
- Une, Y.; Mori, T. Tuberculosis as a Zoonosis from a Veterinary Perspective. Comp. Immunol. Microbiol. Infect. Dis. 2007, 30, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Montali, R.J.; Mikota, S.K.; Cheng, L.I. Mycobaterium tuberculosis in Zoo and Wildlife Species. Sci. Tech. Rev. Int. Off. Epizoot. 2001, 20, 291–303. [Google Scholar] [CrossRef] [PubMed]
- Rosen, L.E.; Hanyire, T.G.; Dawson, J.; Foggin, C.M.; Michel, A.L.; Huyvaert, K.P.; Miller, M.A.; Olea-Popelka, F.J. Tuberculosis Serosurveillance and Management Practices of Captive African Elephants (Loxodonta africana) in the Kavango-Zambezi Transfrontier Conservation Area. Transbound. Emerg. Dis. 2018, 65, e344–e354. [Google Scholar] [CrossRef] [PubMed]
- Landolfi, J.A.; Terio, K.A.; Miller, M.; Junecko, B.F.; Reinhart, T. Pulmonary Tuberculosis in Asian Elephants (Elephas maximus): Histologic Lesions with Correlation to Local Immune Responses. Vet. Pathol. 2015, 52, 535–542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarad, P.; Tsubota, T. Tuberculosis in Elephants: A Zoonotic Disease at the Human-Elephant Interface. Japanese J. Zoo Wildl. Med. 2016, 21, 65–69. [Google Scholar] [CrossRef]
- Bezos, J.; Casal, C.; Díez-Delgado, I.; Romero, B.; Liandris, E.; Álvarez, J.; Sevilla, I.A.; de Juan, L.; Domínguez, L.; Gortázar, C. Goats Challenged with Different Members of the Mycobacterium tuberculosis Complex Display Different Clinical Pictures. Vet. Immunol. Immunopathol. 2015, 167, 185–189. [Google Scholar] [CrossRef]
- Cadmus, S.I.B.; Jenkins, A.O.; Godfroid, J.; Osinusi, K.; Adewole, I.F.; Murphy, R.L.; Taiwo, B.O. Mycobacterium tuberculosis and Mycobacterium africanum in Stools from Children Attending an Immunization Clinic in Ibadan, Nigeria. Int. J. Infect. Dis. 2009, 13, 740–744. [Google Scholar] [CrossRef]
- Silva, M.L.; Cá, B.; Osório, N.S.; Rodrigues, P.N.S.; Maceiras, A.R.; Saraiva, M. Tuberculosis Caused by Mycobacterium africanum: Knowns and Unknowns. PLoS Pathog. 2022, 18, e1010490. [Google Scholar] [CrossRef]
- Gonzalo-Asensio, J.; Astarie-Dequeker, C.; Malaga, W.; Brosch, R.; Moreau, F.; Passemar, C.; Martin, C.; Laval, F.; Daffe, M.; Guilhot, C.; et al. Evolutionary History of Tuberculosis Shaped by Conserved Mutations in the PhoPR Virulence Regulator. Proc. Natl. Acad. Sci. USA 2014, 111, 11491–11496. [Google Scholar] [CrossRef]
- Rahim, Z.; Möllers, M.; Te Koppele-Vije, A.; De Beer, J.; Zaman, K.; Matin, M.A.; Kamal, M.; Raquib, R.; Van Soolingen, D.; Baqi, M.A.; et al. Characterization of Mycobacterium africanum Subtype 1 among Cows in a Dairy Farm in Bangladesh Using Spoligotyping. Southeast Asian J. Trop. Med. Public Health 2007, 38, 706–713. [Google Scholar]
- Gudan, A.; Artuković, B.; Cvetnić, Ž.; Špičić, S.; Beck, A.; Hohšteter, M.; Naglić, T.; Bata, I.; Grabarević, Ž. Disseminated Tuberculosis in Hyrax (Procavia capensis) Caused by Mycobacterium africanum. J. Zoo Wildl. Med. 2008, 39, 386–391. [Google Scholar] [CrossRef] [PubMed]
- Cousins, D.V.; Bastida, R.; Cataldi, A.; Quse, V.; Redrobe, S.; Dow, S.; Duignan, P.; Murray, A.; Dupont, C.; Ahmed, N.; et al. Tuberculosis in Seals Caused by a Novel Member of the Mycobacterium tuberculosis Complex: Mycobacterium pinnipedii Sp. Nov. Int. J. Syst. Evol. Microbiol. 2003, 53, 1305–1314. [Google Scholar] [CrossRef]
- Kiers, A.; Klarenbeek, A.; Mendelts, B.; Van Soolingen, D.; Koëter, G. Transmission of Mycobacterium pinnipedii to Humans in a Zoo with Marine Mammals. Int. J. Tuberc. Lung Dis. 2008, 12, 1469–1473. [Google Scholar] [PubMed]
- Macedo, R.; Isidro, J.; Gomes, M.C.; Botelho, A.; Albuquerque, T.; Sogorb, A.; Bernardino, R.; Fernandes, T.L.; Mourato, T.; Durval, M.; et al. Animal-to-Human Transmission of Mycobacterium pinnipedii. Eur. Respir. J. 2020, 56, 2000371. [Google Scholar] [CrossRef]
- Loeffler, S.H.; de Lisle, G.W.; Neill, M.A.; Collins, D.M.; Price-Carter, M.; Paterson, B.; Crews, K.B. The Seal Tuberculosis Agent, Mycobacterium pinnipedii, Infects Domestic Cattle in New Zealand: Epidemiologic Factors and DNA Strain Typing. J. Wildl. Dis. 2014, 50, 180–187. [Google Scholar] [CrossRef] [PubMed]
- Aranaz, A.; Liebana, E.; Gomez-Mampaso, E.; Galán, J.C.; Cousins, D.; Ortega, A.; Blázquez, J.; Baquero, F.; Mateos, A.; Súarez, G.; et al. Mycobacterium tuberculosis Subsp. caprae subsp. nov.: A Taxonomic Study of a New Member of the Mycobacterium tuberculosis Complex Isolated from Goats in Spain. Int. J. Syst. Bacteriol. 1999, 49, 1263–1273. [Google Scholar] [CrossRef] [PubMed]
- Gormley, E.; Corner, L.A.L.; Costello, E.; Rodriguez-Campos, S. Bacteriological Diagnosis and Molecular Strain Typing of Mycobacterium bovis and Mycobacterium caprae. Res. Vet. Sci. 2014, 97, S30–S43. [Google Scholar] [CrossRef]
- Shrestha, A.; Picoy, J.; Torres, A.; Moore, D.A.; Gilman, R.H.; Coronel, J.; Grandjean, L. A Case Report of Transmission and Disease Caused by Mycobacterium caprae and Mycobacterium bovis in Lima, Peru. BMC Infect. Dis. 2021, 21, 4–9. [Google Scholar] [CrossRef]
- Huitema, H.; Jaartsveld, F.H.J. Mycobacterium microti Infection in a Cat and Some Pigs. Antonie Van Leeuwenhoek 1967, 33, 209–212. [Google Scholar] [CrossRef]
- Wells, A.Q.; Robb-Smith, A.H.T. Murine Type of Tubercle Bacillus (the Vole Acid-Fast Bacillus). 1946. Available online: https://agris.fao.org/agris-search/search.do?recordID=US201300343701 (accessed on 11 August 2022).
- Van Soolingen, D.; Van Der Zanden, A.G.M.; De Haas, P.E.W.; Noordhoek, G.T.; Kiers, A.; Foudraine, N.A.; Portaels, F.; Kolk, A.H.J.; Kremer, K.; Van Embden, J.D.A. Diagnosis of Mycobacterium microti Infections among Humans by Using Novel Genetic Markers. J. Clin. Microbiol. 1998, 36, 1840–1845. [Google Scholar] [CrossRef]
- van Ingen, J.; Brosch, R.; van Soolingen, D. Characterization of Mycobacterium orygis. Emerg. Infect. Dis. 2013, 19, 521–522. [Google Scholar] [CrossRef] [PubMed]
- Van Soolingen, D.; De Haas, P.E.W.; Haagsma, J.; Eger, T.; Hermans, P.W.M.; Ritacco, V.; Alito, A.; Va Embdenl, J.D.A. Use of Various Genetic Markers in Differentiation of Mycobacterium bovis Strains from Animals and Humans and for Studying Epidemiology of Bovine Tuberculosis. J. Clin. Microbiol. 1994, 32, 2425–2433. [Google Scholar] [CrossRef] [PubMed]
- Parsons, S.D.C. Mycobacterium orygis: A Zoonosis, Zooanthroponosis, or Both? Lancet Microbe 2020, 1, e240. [Google Scholar] [CrossRef]
- Pfyffer, G.E.; Auckenthaler, R.; Van Embden, J.D.A.; Van Soolingen, D. Mycobacterium canettii, the Smooth Variant of Mycobacterium tuberculosis, Isolated from a Swiss Patient Exposed in Africa. Emerg. Infect. Dis. 1998, 4, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Van Soolingen, D.; Hoogenboezem, T.; De Haas, P.E.W.; Hermans, P.W.M.; Koedam, M.A.; Teppema, K.S.; Brennan, P.J.; Besra, G.S.; Portaels, F.; Top, J.; et al. A Novel Pathogenic Taxon of the Mycobacterium tuberculosis Complex, Canetti: Characterization of an Exceptional Isolate from Africa. Int. J. Syst. Bacteriol. 1997, 47, 1236–1245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supply, P.; Brosch, R. The Biology and Epidemiology of Mycobacterium canettii. Adv. Exp. Med. Biol. 2017, 1019, 27–41. [Google Scholar] [CrossRef]
- Alexander, K.A.; Laver, P.N.; Michel, A.L.; Williams, M.; van Helden, P.D.; Warren, R.M.; van Pittius, N.C.G. Novel Mycobacterium tuberculosis Complex Pathogen, M. Mungi. Emerg. Infect. Dis. 2010, 16, 1296–1299. [Google Scholar] [CrossRef]
- Dippenaar, A.; Parsons, S.D.C.; Sampson, S.L.; Van Der Merwe, R.G.; Drewe, J.A.; Abdallah, A.M.; Siame, K.K.; Gey Van Pittius, N.C.; Van Helden, P.D.; Pain, A.; et al. Whole Genome Sequence Analysis of Mycobacterium suricattae. Tuberculosis 2015, 95, 682–688. [Google Scholar] [CrossRef]
- Clarke, C.; Van Helden, P.; Miller, M.; Parsons, S. Animal-Adapted Members of the Mycobacterium tuberculosis Complex Endemic to the Southern African Subregion. J. S. Afr. Vet. Assoc. 2016, 87, 1322. [Google Scholar] [CrossRef]
- Parsons, S.D.C.; Drewe, J.A.; van Pittius, N.C.G.; Warren, R.M.; van Helden, P.D. Novel Cause of Tuberculosis in Meerkats, South Africa. Emerg. Infect. Dis. 2013, 19, 2004–2007. [Google Scholar] [CrossRef]
- Glaziou, P.; Floyd, K.; Raviglione, M.C. Global Epidemiology of Tuberculosis. Semin. Respir. Crit. Care Med. 2015, 39, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Morris, R.S.; Pfeiffer, D.U.; Jackson, R. The Epidemiology of Mycobacterium bovis Infections. Vet. Microbiol. 1994, 40, 153–177. [Google Scholar] [CrossRef]
- Phipps, E.; McPhedran, K.; Edwards, D.; Russell, K.; O’Connor, C.M.; Gunn-Moore, D.A.; O’Halloran, C.; Roberts, T.; Morris, J. Bovine Tuberculosis in Working Foxhounds: Lessons Learned from a Complex Public Health Investigation. Epidemiol. Infect. 2019, 147, E24. [Google Scholar] [CrossRef] [PubMed]
- Haddad, N.; Ostyn, A.; Karoui, C.; Masselot, M.; Thorel, M.F.; Hughes, S.L.; Inwald, J.; Hewinson, R.G.; Durand, B. Spoligotype Diversity of Mycobacterium bovis Strains Isolated in France from 1979 to 2000. J. Clin. Microbiol. 2001, 39, 3623–3632. [Google Scholar] [CrossRef] [PubMed]
- Almaw, G.; Mihret, A.; Abebe, T.; Ameni, G.; Gumi, B.; Olani, A.; Tamiru, M.; Koran, T.; Aliy, A.; Sombo, M.; et al. Spoligotype Analysis of Mycobacterium bovis Isolates from Cattle and Assessment of Zoonotic TB Transmission among Individuals Working in Bovine TB- Infected Dairy Farms in Ethiopia. Zoonoses Public Health 2022, 69, 663–672. [Google Scholar] [CrossRef]
- Branger, M.; Loux, V.; Cochard, T.; Boschiroli, M.L.; Biet, F.; Michelet, L. The Complete Genome Sequence of Mycobacterium bovis Mb3601, a SB0120 Spoligotype Strain Representative of a New Clonal Group. Infect. Genet. Evol. 2020, 82, 104309. [Google Scholar] [CrossRef]
- Boniotti, M.B.; Goria, M.; Loda, D.; Garrone, A.; Benedetto, A.; Mondo, A.; Tisato, E.; Zanoni, M.; Zoppi, S.; Dondo, A.; et al. Molecular Typing of Mycobacterium bovis Strains Isolated in Italy from 2000 to 2006 and Evaluation of Variable-Number Tandem Repeats for Geographically Optimized Genotyping. J. Clin. Microbiol. 2009, 47, 636–644. [Google Scholar] [CrossRef]
- Hlokwe, T.M.; Van Helden, P.; Michel, A. Evaluation of the Discriminatory Power of Variable Number of Tandem Repeat Typing of Mycobacterium bovis Isolates from Southern Africa. Transbound. Emerg. Dis. 2013, 60 (Suppl. S1), 111–120. [Google Scholar] [CrossRef]
- Amato, B.; Di Marco Lo Presti, V.; Gerace, E.; Capucchio, M.T.; Vitale, M.; Zanghì, P.; Pacciarini, M.L.; Marianelli, C.; Boniotti, M.B. Molecular Epidemiology of Mycobacterium tuberculosis Complex Strains Isolated from Livestock and Wild Animals in Italy Suggests the Need for a Different Eradication Strategy for Bovine Tuberculosis. Transbound. Emerg. Dis. 2018, 65, e416–e424. [Google Scholar] [CrossRef]
- Broeckl, S.; Krebs, S.; Varadharajan, A.; Straubinger, R.K.; Blum, H.; Buettner, M. Investigation of Intra-Herd Spread of Mycobacterium caprae in Cattle by Generation and Use of a Whole-Genome Sequence. Vet. Res. Commun. 2017, 41, 113–128. [Google Scholar] [CrossRef]
- Orloski, K.; Robbe-Austerman, S.; Tod Stuber, B.H.; Schoenbaum, M. Whole Genome Sequencing of Mycobacterium bovis Isolated from Livestock in the United States, 1989–2018. Front. Vet. Sci. 2018, 5, 253. [Google Scholar] [CrossRef]
- Valcheva, V.; Perea, C.; Savova-Lalkovska, T.; Dimitrova, A.; Radulski, L.; Mokrousov, I.; Marinov, K.; Najdenski, H.; Bonovska, M. Mycobacterium bovis and Mycobacterium caprae in Bulgaria: Insight into Transmission and Phylogeography Gained through Whole-Genome Sequencing. BMC Vet. Res. 2022, 18, 148. [Google Scholar] [CrossRef] [PubMed]
- Sandoval-Azuara, S.E.; Muñiz-Salazar, R.; Perea-Jacobo, R.; Robbe-Austerman, S.; Perera-Ortiz, A.; López-Valencia, G.; Bravo, D.M.; Sanchez-Flores, A.; Miranda-Guzmán, D.; Flores-López, C.A.; et al. Whole Genome Sequencing of Mycobacterium bovis to Obtain Molecular Fingerprints in Human and Cattle Isolates from Baja California, Mexico. Int. J. Infect. Dis. 2017, 63, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Price-Carter, M.; Brauning, R.; de Lisle, G.W.; Livingstone, P.; Neill, M.; Sinclair, J.; Paterson, B.; Atkinson, G.; Knowles, G.; Crews, K.; et al. Whole Genome Sequencing for Determining the Source of Mycobacterium bovis Infections in Livestock Herds and Wildlife in New Zealand. Front. Vet. Sci. 2018, 5, 272. [Google Scholar] [CrossRef] [PubMed]
- de la Rua-Domenech, R.; Goodchild, A.T.; Vordermeier, H.M.; Hewinson, R.G.; Christiansen, K.H.; Clifton-Hadley, R.S. Ante Mortem Diagnosis of Tuberculosis in Cattle: A Review of the Tuberculin Tests, γ-Interferon Assay and Other Ancillary Diagnostic Techniques. Res. Vet. Sci. 2006, 81, 190–210. [Google Scholar] [CrossRef]
- Roos, E.O.; Olea-Popelka, F.; Buss, P.; de Klerk-Lorist, L.M.; Cooper, D.; Warren, R.M.; van Helden, P.D.; Parsons, S.D.C.; Miller, M.A. IP-10: A Potential Biomarker for Detection of Mycobacterium bovis Infection in Warthogs (Phacochoerus africanus). Vet. Immunol. Immunopathol. 2018, 201, 43–48. [Google Scholar] [CrossRef]
- Roos, E.O. Detection and Characterization of Mycobacterial Infections Occurring in Phacochoerus Africanus (Gmelin, 1788) (Common Warthog); Stellenbosch University: Stellenbosch, South Africa, 2018. [Google Scholar]
- Majoor, C.J.; Magis-Escurra, C.; van Ingen, J.; Boeree, M.J.; van Soolingen, D. Epidemiology of Mycobacterium bovis Disease in Humans, The Netherlands, 1993–2007. Emerg. Infect. Dis. 2011, 17, 457–463. [Google Scholar] [CrossRef]
- Rabaa, M.A.; Tue, N.T.; Phuc, T.M.; Carrique-Mas, J.; Saylors, K.; Cotten, M.; Bryant, J.E.; Nghia, H.D.T.; Van Cuong, N.; Pham, H.A.; et al. The Vietnam Initiative on Zoonotic Infections (VIZIONS): A Strategic Approach to Studying Emerging Zoonotic Infectious Diseases. Ecohealth 2015, 12, 726–735. [Google Scholar] [CrossRef]
- Durr, P.A.; Hewinson, R.G.; Clifton-Hadley, R.S. Molecular Epidemiology of Bovine Tuberculosis. I. Mycobacterium bovis Genotyping. Rev. Sci. Tech. 2000, 19, 675–688. [Google Scholar] [CrossRef]
- Mostowy, S.; Behr, M.A. Comparative Genomics in the Fight against Tuberculosis: Diagnostics, Epidemiology, and BCG Vaccination. Am. J. Pharm. 2002, 2, 189–196. [Google Scholar] [CrossRef]
- Campbell, E.L.; Byrne, A.W.; Menzies, F.D.; Milne, G.; McBride, K.R.; McCormick, C.M.; Scantlebury, D.M.; Reid, N. Quantifying Intraherd Cattle Movement Metrics: Implications for Disease Transmission Risk. Prev. Vet. Med. 2020, 185, 106192. [Google Scholar] [CrossRef] [PubMed]
- Ghebremariam, M.K.; Michel, A.L.; Vernooij, J.C.M.; Nielen, M.; Rutten, V.P.M.G. Prevalence of Bovine Tuberculosis in Cattle, Goats, and Camels of Traditional Livestock Raising Communities in Eritrea. BMC Vet. Res. 2018, 14, 73. [Google Scholar] [CrossRef]
- O’Brien, D.J.; Schmitt, S.M.; Fierke, J.S.; Hogle, S.A.; Winterstein, S.R.; Cooley, T.M.; Moritz, W.E.; Diegel, K.L.; Fitzgerald, S.D.; Berry, D.E.; et al. Epidemiology of Mycobacterium bovis in Free-Ranging White-Tailed Deer, Michigan, USA, 1995–2000. Prev. Vet. Med. 2002, 54, 47–63. [Google Scholar] [CrossRef]
- Zimpel, C.K.; Patané, J.S.L.; Guedes, A.C.P.; de Souza, R.F.; Silva-Pereira, T.T.; Camargo, N.C.S.; de Souza Filho, A.F.; Ikuta, C.Y.; Neto, J.S.F.; Setubal, J.C.; et al. Global Distribution and Evolution of Mycobacterium bovis Lineages. Front. Microbiol. 2020, 11, 843. [Google Scholar] [CrossRef] [PubMed]
- Galagan, J.E. Genomic Insights into Tuberculosis. Nat. Rev. Genet. 2014, 15, 307–320. [Google Scholar] [CrossRef]
- Coscolla, M.; Gagneux, S. Consequences of Genomic Diversity in Mycobacterium tuberculosis. Semin. Immunol. 2014, 26, 431–444. [Google Scholar] [CrossRef]
- Otchere, I.D.; van Tonder, A.J.; Asante-Poku, A.; Sánchez-Busó, L.; Coscollá, M.; Osei-Wusu, S.; Asare, P.; Aboagye, S.Y.; Ekuban, S.A.; Yahayah, A.I.; et al. Molecular Epidemiology and Whole Genome Sequencing Analysis of Clinical Mycobacterium bovis from Ghana. PLoS ONE 2019, 14, e0209395. [Google Scholar] [CrossRef]
- De Jong, B.C.; Adetifa, I.; Walther, B.; Hill, P.C.; Antonio, M.; Ota, M.; Adegbola, R.A. Differences between Tuberculosis Cases Infected with Mycobacterium africanum, West African Type 2, Relative to Euro-American Mycobacterium tuberculosis: An Update. FEMS Immunol. Med. Microbiol. 2010, 58, 102–105. [Google Scholar] [CrossRef]
- Palisson, A.; Courcoul, A.; Durand, B. Role of Cattle Movements in Bovine Tuberculosis Spread in France between 2005 and 2014. PLoS ONE 2016, 11, e0152578. [Google Scholar] [CrossRef]
- Pereira, A.C.; Reis, A.C.; Ramos, B.; Cunha, M.V. Animal Tuberculosis: Impact of Disease Heterogeneity in Transmission, Diagnosis and Control. Transbound. Emerg. Dis. 2020, 2019, 13539. [Google Scholar] [CrossRef]
- Jones, B.A.; Grace, D.; Kock, R.; Alonso, S.; Rushton, J.; Said, M.Y.; McKeever, D.; Mutua, F.; Young, J.; McDermott, J.; et al. Zoonosis Emergence Linked to Agricultural Intensification and Environmental Change. Proc. Natl. Acad. Sci. USA 2013, 110, 8399–8404. [Google Scholar] [CrossRef] [PubMed]
- Gopal, R.; Goodchild, A.; Hewinson, G.; de la Rua Domenech, R.; Clifton-Hadley, R. Introduction of Bovine Tuberculosis to North-East England by Bought-in Cattle. Vet. Rec. 2006, 159, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Brooks-Pollock, E.; Roberts, G.O.; Keeling, M.J. A Dynamic Model of Bovine Tuberculosis Spread and Control in Great Britain. Nature 2014, 511, 228–231. [Google Scholar] [CrossRef]
- Reynolds, D. A Review of Tuberculosis Science and Policy in Great Britain. Vet. Microbiol. 2006, 112, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Mosavari, N.; Feizabadi, M.M.; Jamshidian, M.; Shahpouri, M.R.S.; Forbes, K.J.; Pajoohi, R.A.; Keshavarz, R.; Taheri, M.M.; Tadayon, K. Molecular Genotyping and Epidemiology of Mycobacterium bovis Strains Obtained from Cattle in Iran. Vet. Microbiol. 2011, 151, 148–152. [Google Scholar] [CrossRef]
- Pozo, P.; VanderWaal, K.; Grau, A.; de la Cruz, M.L.; Nacar, J.; Bezos, J.; Perez, A.; Minguez, O.; Alvarez, J. Analysis of the Cattle Movement Network and Its Association with the Risk of Bovine Tuberculosis at the Farm Level in Castilla y Leon, Spain. Transbound. Emerg. Dis. 2019, 66, 327–340. [Google Scholar] [CrossRef]
- Sylvester, T.T.; Martin, L.E.R.; Buss, P.; Loxton, A.G.; Hausler, G.A.; Rossouw, L.; van Helden, P.; Parsons, S.D.C.; Olea-Popelka, F.; Miller, M.A. Prevalence and Risk Factors for Mycobacterium bovis Infection in African Lions (Panthera leo) in the Kruger National Park. J. Wildl. Dis. 2017, 53, 372–376. [Google Scholar] [CrossRef]
- Botha, L.; Gey van Pittius, N.C.; van Helden, P.D. Mycobacteria and Disease in Southern Africa. Transbound. Emerg. Dis. 2013, 60 (Suppl. S1), 147–156. [Google Scholar] [CrossRef]
- Corner, L.A.L.; Murphy, D.; Gormley, E. Mycobacterium bovis Infection in the Eurasian Badger (Meles meles): The Disease, Pathogenesis, Epidemiology and Control. J. Comp. Pathol. 2011, 144, 1–24. [Google Scholar] [CrossRef]
- Corner, L.A.L.; O’Meara, D.; Costello, E.; Lesellier, S.; Gormley, E. The Distribution of Mycobacterium bovis Infection in Naturally Infected Badgers. Vet. J. 2012, 194, 166–172. [Google Scholar] [CrossRef]
- Turgenbayev, K.A.; Borsynbayeva, A.M.; Plazun, A.A.; Turgenbayev, R.K. Tuberculosis Prevalence in Animals and Humans in the Republic of Kazakhstan. Vet. World 2021, 14, 2362–2370. [Google Scholar] [CrossRef] [PubMed]
- Luciano, S.A.; Roess, A. Human Zoonotic Tuberculosis and Livestock Exposure in Low- and Middle-Income Countries: A Systematic Review Identifying Challenges in Laboratory Diagnosis. Zoonoses Public Health 2020, 67, 97–111. [Google Scholar] [CrossRef]
- Sichewo, P.R.; Etter, E.M.C.C.; Michel, A.L. Wildlife-Cattle Interactions Emerge as Drivers of Bovine Tuberculosis in Traditionally Farmed Cattle. Prev. Vet. Med. 2020, 174, 104847. [Google Scholar] [CrossRef]
- Sichewo, P.R.; Hlokwe, T.M.; Etter, E.M.C.C.; Michel, A.L. Tracing Cross Species Transmission of Mycobacterium bovis at the Wildlife / Livestock Interface in South Africa. BMC Microbiol. 2020, 20, 49. [Google Scholar] [CrossRef]
- Etter, E.; Donado, P.; Jori, F.; Caron, A.; Goutard, F.; Roger, F. Risk Analysis and Bovine Tuberculosis, a Re-Emerging Zoonosis. Ann. N. Y. Acad. Sci. 2006, 1081, 61–73. [Google Scholar] [CrossRef]
- Thoen, C.; LoBue, P.; De Kantor, I. The Importance of Mycobacterium bovis as a Zoonosis. Vet. Microbiol. 2006, 112, 339–345. [Google Scholar] [CrossRef]
- Awah Ndukum, J.; Caleb Kudi, A.; Bradley, G.; Ane-Anyangwe, I.N.; Fon-Tebug, S.; Tchoumboue, J. Prevalence of Bovine Tuberculosis in Abattoirs of the Littoral and Western Highland Regions of Cameroon: A Cause for Public Health Concern. Vet. Med. Int. 2010, 2010, 495015. [Google Scholar] [CrossRef]
- Ali, Z.I.; Hanafy, M.; Hansen, C.; Saudi, A.M.; Talaat, A.M. Genotypic Analysis of Nontuberculous Mycobacteria Isolated from Raw Milk and Human Cases in Wisconsin. J. Dairy Sci. 2020, 104, 211–220. [Google Scholar] [CrossRef]
- Collins, Á.B.; Floyd, S.; Gordon, S.V.; More, S.J. Prevalence of Mycobacterium bovis in Milk on Dairy Cattle Farms: An International Systematic Literature Review and Meta-Analysis. Tuberculosis 2022, 132, 102166. [Google Scholar] [CrossRef] [PubMed]
- Bapat, P.R.; Dodkey, R.S.; Shekhawat, S.D.; Husain, A.A.; Nayak, A.R.; Kawle, A.P.; Daginawala, H.F.; Singh, L.K.; Kashyap, R.S. Prevalence of Zoonotic Tuberculosis and Associated Risk Factors in Central Indian Populations. J. Epidemiol. Glob. Health 2017, 7, 277–283. [Google Scholar] [CrossRef] [PubMed]
- Desta, G.B.; Jena, P.K.; Hassen, H.M.; Behera, M.R. Factors Associated with Zoonosis and Reverse Zoonosis of Mycobacterium tuberculosis and Mycobacterium bovis in Ethiopia: A Systematic Review and Meta-Analysis. Int. J. Health Sci. 2022, 6, 1630–1653. [Google Scholar] [CrossRef]
- Tober, A.V.; Govender, D.; Russo, I.-R.M.; Cable, J. The Microscopic Five of the Big Five: Managing Zoonotic Diseases within and beyond African Wildlife Protected Areas. Adv. Parasitol. 2022, 117, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Dürr, S.; Müller, B.; Alonso, S.; Hattendorf, J.; Laisse, C.J.M.; van Helden, P.D.; Zinsstag, J. Differences in Primary Sites of Infection between Zoonotic and Human Tuberculosis: Results from a Worldwide Systematic Review. PLoS Negl. Trop. Dis. 2013, 7, e2399. [Google Scholar] [CrossRef] [PubMed]
- Thoen, C.O.; LoBue, P.A.; Enarson, D.A. Tuberculosis in Animals and Humans: An Overview. Int. J. Tuberc. Lung Dis. 2010, 14, 1075–1078. [Google Scholar] [CrossRef]
- Carruth, L.; Roess, A.A.; Mekonnen, Y.T.; Melaku, S.K.; Nichter, M.; Salman, M. Zoonotic Tuberculosis in Africa: Challenges and Ways Forward. Lancet 2016, 388, 2460–2461. [Google Scholar] [CrossRef]
- Kock, R.; Michel, A.L.; Yeboah-Manu, D.; Azhar, E.I.; Torrelles, J.B.; Cadmus, S.I.; Brunton, L.; Chakaya, J.M.; Marais, B.; Mboera, L.; et al. Zoonotic Tuberculosis–The Changing Landscape. Int. J. Infect. Dis. 2021, 113, S68–S72. [Google Scholar] [CrossRef]
- van den Brom, R.; de Jong, A.; van Engelen, E.; Heuvelink, A.; Vellema, P. Zoonotic Risks of Pathogens from Sheep and Their Milk Borne Transmission. Small Rumin. Res. 2020, 189, 106123. [Google Scholar] [CrossRef]
- Drain, P.K.; Bajema, K.L.; Dowdy, D.; Dheda, K.; Naidoo, K.; Schumacher, S.G.; Ma, S.; Meermeier, E.; Lewinsohn, D.M.; Sherman, D.R.; et al. Incipient and Subclinical Tuberculosis: A Clinical Review of Early Stages and Progression of Infection. Clin. Microbiol. Rev. 2018, 31, e00021-18. [Google Scholar] [CrossRef]
- Dutta, N.K.; Karakousis, P.C. Latent Tuberculosis Infection: Myths, Models, and Molecular Mechanisms. Microbiol. Mol. Biol. Rev. 2014, 78, 343–371. [Google Scholar] [CrossRef]
- Furin, J.; Cox, H.; Pai, M. Tuberculosis. Lancet 2019, 393, 1642–1656. [Google Scholar] [CrossRef]
- Esmail, H.; Barry, C.E.; Wilkinson, R.J. Understanding Latent Tuberculosis: The Key to Improved Diagnostic and Novel Treatment Strategies. Drug Discov. Today 2012, 17, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Suárez, I.; Fünger, S.M.; Kröger, S.; Rademacher, J.; Gerd Fätkenheuer, J.; Rybniker, A. The Diagnosis and Treatment of Tuberculosis. Dtsch. Arztebl. Int. 2019, 116, 729–735. [Google Scholar] [CrossRef] [PubMed]
- Zumla, A.I.; Lawn, S.D. Tuberculosis. Lancet 2011, 378, 57–72. [Google Scholar] [CrossRef] [PubMed]
- Faryar, K.A.; Braun, R.; Ancona, R.M.; Ajayi, E.; Bryant, W.; Rehman, S.; Sall, H.; Lyons, M.S.; Huaman, M.A. Emergency Department Screening for Latent Tuberculosis Infection. Am. J. Emerg. Med. 2021, 54, 323.e5–323.e8. [Google Scholar] [CrossRef]
- Houben, R.M.G.J.; Dodd, P.J. The Global Burden of Latent Tuberculosis Infection: A Re-Estimation Using Mathematical Modelling. PLoS Med. 2016, 13, e1002152. [Google Scholar] [CrossRef] [Green Version]
- Bañuls, A.L.; Sanou, A.; Van Anh, N.T.; Godreuil, S. Mycobacterium tuberculosis: Ecology and Evolution of a Human Bacterium. J. Med. Microbiol. 2015, 64, 1261–1269. [Google Scholar] [CrossRef]
- Hershberg, R.; Lipatov, M.; Small, P.M.; Sheffer, H.; Niemann, S.; Homolka, S.; Roach, J.C.; Kremer, K.; Petrov, D.A.; Feldman, M.W.; et al. High Functional Diversity in Mycobacterium tuberculosis Driven by Genetic Drift and Human Demography. PLoS Biol. 2008, 6, 2658–2671. [Google Scholar] [CrossRef]
- Parbhoo, T.; Sampson, S.L.; Mouton, J.M. Recent Developments in the Application of Flow Cytometry to Advance Our Understanding of Mycobacterium tuberculosis Physiology and Pathogenesis. Cytom. Part A 2020, 97, 683–693. [Google Scholar] [CrossRef]
- Gengenbacher, M.; Kaufmann, S.H.E. Mycobacterium tuberculosis: Success through Dormancy. FEMS Microbiol. Rev. 2012, 36, 514–532. [Google Scholar] [CrossRef]
- Borah, K.; Xu, Y.; McFadden, J. Dissecting Host-Pathogen Interactions in TB Using Systems-Based Omic Approaches. Front. Immunol. 2021, 12, 762315. [Google Scholar] [CrossRef]
- Gagneux, S.; DeRiemer, K.; Van, T.; Kato-Maeda, M.; De Jong, B.C.; Narayanan, S.; Nicol, M.; Niemann, S.; Kremeri, K.; Gutierrez, M.C.; et al. Variable Host-Pathogen Compatibility in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2006, 103, 2869–2873. [Google Scholar] [CrossRef]
- Manina, G.; Dhar, N.; McKinney, J.D. Stress and Host Immunity Amplify Mycobacterium tuberculosis Phenotypic Heterogeneity and Induce Nongrowing Metabolically Active Forms. Cell Host Microbe 2015, 17, 32–46. [Google Scholar] [CrossRef] [PubMed]
- Laureillard, D.; Marcy, O.; Madec, Y.; Chea, S.; Chan, S.; Borand, L.; Fernandez, M.; Prak, N.; Kim, C.; Dim, B.; et al. Paradoxical Tuberculosis-Associated Immune Reconstitution Inflammatory Syndrome after Early Initiation of Antiretroviral Therapy in a Randomized Clinical Trial. Aids 2013, 27, 2577–2586. [Google Scholar] [CrossRef] [PubMed]
- Varela-Castro, L.; Barral, M.; Arnal, M.C.; Luco, D.F.D.; Gortázar, C.; Garrido, J.M.; Sevilla, I.A. Beyond Tuberculosis: Diversity and Implications of Non-Tuberculous Mycobacteria at the Wildlife-Livestock Interface. Transbound. Emerg. Dis. 2022. [Google Scholar] [CrossRef]
- Keet, D.F.; Michel, A.L.; Bengis, R.G.; Becker, P.; van Dyk, D.S.; van Vuuren, M.; Rutten, V.P.M.G.; Penzhorn, B.L. Intradermal Tuberculin Testing of Wild African Lions (Panthera leo) Naturally Exposed to Infection with Mycobacterium bovis. Vet. Microbiol. 2010, 144, 384–391. [Google Scholar] [CrossRef] [PubMed]
- Riska, P.F.; Carleton, S. Latent Tuberculosis: Models, Mechanisms, and Novel Prospects for Eradication. Semin. Pediatr. Infect. Dis. 2002, 13, 263–272. [Google Scholar] [CrossRef]
- Meier, N.R.; Jacobsen, M.; Ottenhoff, T.H.M.; Ritz, N. A Systematic Review on Novel Mycobacterium tuberculosis Antigens and Their Discriminatory Potential for the Diagnosis of Latent and Active Tuberculosis. Front. Immunol. 2018, 9, 2476. [Google Scholar] [CrossRef]
- Helaine, S.; AM, C.; KG, W.; LM, F.; SA, M.; DW, H.; Helaine, S.; Cheverton, A.M.; Watson, K.G.; Faure, L.M.; et al. Internalization of Salmonella by Macrophages Induces Formation of Nonreplicating Persisters. Science 2014, 343, 204–208. [Google Scholar] [CrossRef] [PubMed]
- Mouton, J.M.; Helaine, S.; Holden, D.W.; Sampson, S.L. Elucidating Population-Wide Mycobacterial Replication Dynamics at the Single-Cell Level. Microbiology 2016, 162, 966–978. [Google Scholar] [CrossRef]
- Chengalroyen, M.D.; Beukes, G.M.; Gordhan, B.G.; Streicher, E.M.; Churchyard, G.; Hafner, R.; Warren, R.; Otwombe, K.; Martinson, N.; Kana, B.D. Detection and Quantification of Differentially Culturable Tubercle Bacteria in Sputum from Patients with Tuberculosis. Am. J. Respir. Crit. Care Med. 2016, 194, 1532–1540. [Google Scholar] [CrossRef]
- Chao, M.C.; Rubin, E.J. Letting Sleeping Dos Lie: Does Dormancy Play a Role in Tuberculosis? Annu. Rev. Microbiol. 2010, 64, 293–311. [Google Scholar] [CrossRef]
- Hice, S.A.; Santoscoy, M.C.; Soupir, M.L.; Cademartiri, R. Distinguishing between Metabolically Active and Dormant Bacteria on Paper. Appl. Microbiol. Biotechnol. 2018, 102, 367–375. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y. Persisters, Persistent Infections and the Yin-Yang Model. Emerg. Microbes Infect. 2014, 3, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Brauner, A.; Fridman, O.; Gefen, O.; Balaban, N.Q. Distinguishing between Resistance, Tolerance and Persistence to Antibiotic Treatment. Nat. Rev. Microbiol. 2016, 14, 320–330. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Gerdes, K.; Lewis, K.; McKinney, J.D. A Problem of Persistence: Still More Questions than Answers? Nat. Rev. Microbiol. 2013, 11, 587–591. [Google Scholar] [CrossRef] [PubMed]
- Balaban, N.Q.; Merrin, J.; Chait, R.; Kowalik, L.; Leibler, S. Bacterial Persistence as a Phenotypic Switch. Science 2004, 305, 1622–1625. [Google Scholar] [CrossRef]
- Li, L.; Mendis, N.; Trigui, H.; Oliver, J.D.; Sebastien, P. The Importance of the Viable but Non-Culturable State in Human Bacterial Pathogens. Front. Microbiol. 2014, 5, 258. [Google Scholar] [CrossRef]
- Buddle, B.M.; Vordermeier, H.M.; Hewinson, R.G. Experimental Infection Models of Tuberculosis in Domestic Livestock. Microbiol. Spectr. 2016, 4, 4. [Google Scholar] [CrossRef]
- Domingo, M.; Vidal, E.; Marco, A. Pathology of Bovine Tuberculosis. Res. Vet. Sci. 2014, 97, S20–S29. [Google Scholar] [CrossRef]
- Thoen, C.O. Tuberculosis in Wild and Domestic Mammals. Tuberculosis 1994, 157–162. [Google Scholar] [CrossRef]
- Courtenay, O.; Reilly, L.A.; Sweeney, F.P.; Hibberd, V.; Bryan, S.; Ul-Hassan, A.; Newman, C.; Macdonald, D.W.; Delahay, R.J.; Wilson, G.J.; et al. Is Mycobacterium bovis in the Environment Important for the Persistence of Bovine Tuberculosis? Biol. Lett. 2006, 2, 460–462. [Google Scholar] [CrossRef] [PubMed]
- Ghodbane, R.; Medie, F.M.; Lepidi, H.; Nappez, C.; Drancourt, M. Long-Term Survival of Tuberculosis Complex Mycobacteria in Soil. Microbiology 2014, 160 Pt 3, 496–501. [Google Scholar] [CrossRef] [PubMed]
- Lyashchenko, K.P.; Sikar-Gang, A.; Sridhara, A.A.; Johnathan-Lee, A.; Elahi, R.; Lambotte, P.; Esfandiari, J.; Duthi, M.; Reed, S.G.; Jones, G.; et al. Novel Polyprotein Antigens Designed for Improved Serodiagnosis of Bovine Tuberculosis. Vet. Immunol. Immunopathol. 2021, 240, 110320. [Google Scholar] [CrossRef]
- Lyashchenko, K.P.; Gortázar, C.; Miller, M.A.; Waters, W.R. Spectrum of Antibody Profiles in Tuberculous Elephants, Cervids, and Cattle. Vet. Microbiol. 2018, 214, 89–92. [Google Scholar] [CrossRef] [PubMed]
- Thomas, R.; Chambers, M. Review of Methods Used for Diagnosing Tuberculosis in Captive and Free-Ranging Non-Bovid Species (2012–2020). Pathogens 2021, 10, 584. [Google Scholar] [CrossRef] [PubMed]
- Boadella, M.; Lyashchenko, K.; Greenwald, R.; Esfandiari, J.; Jaroso, R.; Carta, T.; Garrido, J.M.; Vicente, J.; de la Fuente, J.; Gortázar, C. Serologic Tests for Detecting Antibodies against Mycobacterium bovis and Mycobacterium avium Subspecies Paratuberculosis in Eurasian Wild Boar (Sus scrofa scrofa). J. Vet. Diagn. Investig. 2011, 23, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Higgitt, R.L.; Buss, P.E.; Van Helden, P.D.; Miller, M.A.; Parsons, S.D.C. Development of Gene Expression Assays Measuring Immune Responses in the Spotted Hyena (Crocuta crocuta). Afr. Zool. 2017, 52, 99–104. [Google Scholar] [CrossRef]
- Whelan, A.O.; Coad, M.; Cockle, P.J.; Hewinson, G.; Vordermeier, M.; Gordon, S.V. Revisiting Host Preference in the Mycobacterium tuberculosis Complex: Experimental Infection Shows Mycobacterium tuberculosis H37Rv to Be Avirulent in Cattle. PLoS ONE 2010, 5, e8527. [Google Scholar] [CrossRef]
- Parikka, M.; Hammarén, M.M.; Harjula, S.-K.E.; Halfpenny, N.J.A.; Oksanen, K.E.; Lahtinen, M.J.; Pajula, E.T.; Iivanainen, A.; Pesu, M.; Rämet, M. Mycobacterium marinum Causes a Latent Infection That Can Be Reactivated by Gamma Irradiation in Adult Zebrafish. PLoS Pathog. 2012, 8, e1002944. [Google Scholar] [CrossRef]
- Ramos, D.F.; Silva, P.E.A.; Dellagostin, O.A. Diagnosis of Bovine Tuberculosis: Review of Main Techniques. Braz. J. Biol. 2015, 75, 830–837. [Google Scholar] [CrossRef]
- Warren, R.M.; Gey Van Pittius, N.C.; Barnard, M.; Hesseling, A.; Engelke, E.; De Kock, M.; Gutierrez, M.C.; Chege, G.K.; Victor, T.C.; Hoal, E.G.; et al. Differentiation of Mycobacterium tuberculosis Complex by PCR Amplification of Genomic Regions of Difference. Int. J. Tuberc. Lung Dis. 2006, 10, 818–822. [Google Scholar]
- Gong, W.; Wu, X. Differential Diagnosis of Latent Tuberculosis Infection and Active Tuberculosis: A Key to a Successful Tuberculosis Control Strategy. Front. Microbiol. 2021, 12, 745592. [Google Scholar] [CrossRef]
- Bigi, F.; Zumárraga, J.; Cataldi, A.A.; Soria, M.A. Polymorphisms of Twenty Regulatory Proteins between Mycobacterium tuberculosis and Mycobacterium bovis. Microbiol. Immunol. 2016, 60, 552–560. [Google Scholar] [CrossRef]
- Li, P.; Wang, R.; Dong, W.; Hu, L.; Zong, B.; Zhang, Y.; Wang, X.; Guo, A.; Zhang, A.; Xiang, Y.; et al. Comparative Proteomics Analysis of Human Macrophages Infected with Virulent Mycobacterium bovis. Front. Cell. Infect. Microbiol. 2017, 7, 65. [Google Scholar] [CrossRef] [PubMed]
- Meiring, C.; Van Helden, P.; Goosen, W.J. TB Control in Humans and Animals in South Africa: A Perspective on Problems and Successes. Front. Vet. Sci. 2018, 5, 298. [Google Scholar] [CrossRef] [PubMed]
- Lim, A.; Steibel, J.P.; Coussens, P.M.; Grooms, D.L.; Bolin, S.R. Differential Gene Expression Segregates Cattle Confirmed Positive for Bovine Tuberculosis from Antemortem Tuberculosis Test-False Positive Cattle Originating from Herds Free of Bovine Tuberculosis. Vet. Med. Int. 2012, 2012, 192926. [Google Scholar] [CrossRef] [PubMed]
- Patrick, B. Michigan Bovine Tuberculosis Eradication Project: Activities Report. 2003. Available online: https://digitalcommons.unl.edu/michbovinetb/12/ (accessed on 8 March 2022).
- Parsons, S.D.C.; Cooper, D.; Mccall, A.J.; Mccall, W.A.; Streicher, E.M.; Le Maitre, N.C.; Müller, A.; Gey Van Pittius, N.C.; Warren, R.M.; Van Helden, P.D. Modification of the QuantiFERON-TB Gold (In-Tube) Assay for the Diagnosis of Mycobacterium bovis Infection in African buffaloes (Syncerus caffer). Vet. Immunol. Immunopathol. 2011, 142, 113–118. [Google Scholar] [CrossRef] [PubMed]
- Pai, M.; Denkinger, C.M.; Kik, S.V.; Rangaka, M.X.; Zwerling, A.; Oxlade, O.; Metcalfe, J.Z.; Cattamanchi, A.; Dowdy, D.W.; Dheda, K.; et al. Gamma Interferon Release Assays for Detection of Mycobacterium tuberculosis Infection. Clin. Microbiol. Rev. 2014, 27, 3–20. [Google Scholar] [CrossRef]
- Denis, M.; Wedlock, D.N.; McCarthy, A.R.; Parlane, N.A.; Cockle, P.J.; Vordermeier, H.M.; Hewinson, R.G.; Buddle, B.M. Enhancement of the Sensitivity of the Whole-Blood Gamma Interferon Assay for Diagnosis of Mycobacterium bovis Infections in Cattle. Clin. Vaccine Immunol. 2007, 14, 1483–1489. [Google Scholar] [CrossRef]
- Clegg, T.A.; Good, M.; Doyle, M.; Duignan, A.; More, S.J.; Gormley, E. The Performance of the Interferon Gamma Assay When Used as a Diagnostic or Quality Assurance Test in Mycobacterium bovis Infected Herds. Prev. Vet. Med. 2017, 140, 116–121. [Google Scholar] [CrossRef]
- Frederico, O.; Id, I.; Soares, P.; Yumi, C.; Id, I.; Machado, A.; Ferreira, S.; Id, N.; Id, B.H.; Acha, S.J.; et al. Evolutionary Analysis of Mycobacterium bovis Genotypes across Africa Suggests Co-Evolution with Livestock and Humans. PLoS Negl. Trop. Dis. 2020, 14, e0008081. [Google Scholar] [CrossRef]
- Sukumar, R.; Joshi, B.; Lagrange, P.H.; Ophe Longuet, C.; Verma-Kumar, S.; Abraham, D.; Dendukuri, N.; Cheeran, J.V.; Narayanaswamy Balaji, K. Serodiagnosis of Tuberculosis in Asian Elephants (Elephas maximus) in Southern India: A Latent Class Analysis. PLoS ONE 2012, 7, e49548. [Google Scholar] [CrossRef]
- Golby, P.; Hatch, K.A.; Bacon, J.; Cooney, R.; Riley, P.; Allnutt, J.; Hinds, J.; Nunez, J.; Marsh, P.D.; Hewinson, R.G.; et al. Comparative Transcriptomics Reveals Key Gene Expression Differences between the Human and Bovine Pathogens of the Mycobacterium tuberculosis Complex. Microbiology 2007, 153, 3323–3336. [Google Scholar] [CrossRef] [PubMed]
- Garnier, T.; Eiglmeier, K.; Camus, J.C.; Medina, N.; Mansoor, H.; Pryor, M.; Duthoy, S.; Grondin, S.; Lacroix, C.; Monsempe, C.; et al. The Complete Genome Sequence of Mycobacterium bovis. Proc. Natl. Acad. Sci. USA 2003, 100, 7877–7882. [Google Scholar] [CrossRef]
- Marimani, M.; Ahmad, A.; Duse, A. The Role of Epigenetics, Bacterial and Host Factors in Progression of Mycobacterium tuberculosis Infection. Tuberculosis 2018, 113, 200–214. [Google Scholar] [CrossRef]
- Keating, L.A.; Wheeler, P.R.; Mansoor, H.; Inwald, J.K.; Dale, J.; Hewinson, R.G.; Gordon, S.V. The Pyruvate Requirement of Some Members of the Mycobacterium tuberculosis Complex Is Due to an Inactive Pyruvate Kinase: Implications for in vivo Growth. Mol. Microbiol. 2005, 56, 163–174. [Google Scholar] [CrossRef]
- Sohaskey, C.D.; Modesti, L. Differences in Nitrate Reduction between Mycobacterium tuberculosis and Mycobacterium bovis Are Due to Differential Expression of Both NarGHJI and NarK2. FEMS Microbiol. Lett. 2009, 290, 129–134. [Google Scholar] [CrossRef]
- Queval, C.J.; Fearns, A.; Botella, L.; Smyth, A.; Schnettger, L.; Mitermite, M.; Wooff, E.; Villarreal-Ramos, B.; Garcia-Jimenezi, W.; Heunis, T.; et al. Macrophage-Specific Responses to Humanand Animal-Adapted Tubercle Bacilli Reveal Pathogen and Host Factors Driving Multinucleated Cell Formation. PLoS Pathog. 2021, 17, e1009410. [Google Scholar] [CrossRef]
- Cheng, G.; Hussain, T.; Sabir, N.; Ni, J.; Li, M.; Zhao, D.; Zhou, X. Comparative Study of the Molecular Basis of Pathogenicity of Mycobacterium bovis Strains in a Mouse Model. Int. J. Mol. Sci. 2019, 20, 5. [Google Scholar] [CrossRef]
- Nedeltchev, G.G.; Raghunand, T.R.; Jassal, M.S.; Lun, S.; Cheng, Q.J.; Bishai, W.R. Extrapulmonary Dissemination of Mycobacterium bovis but Not Mycobacterium tuberculosis in a Bronchoscopic Rabbit Model of Cavitary Tuberculosis. Infect. Immun. 2009, 77, 598–603. [Google Scholar] [CrossRef]
- Palmer, M.V.; Thacker, T.C.; Kanipe, C.; Boggiatto, P.M.; Palmer, M.V. Heterogeneity of Pulmonary Granulomas in Cattle Experimentally Infected with Mycobacterium bovis. Front. Vet. Sci. 2021, 8, 671460. [Google Scholar] [CrossRef] [PubMed]
- Mouton, J.M.; Heunis, T.; Dippenaar, A.; Gallant, J.L.; Kleynhans, L.; Sampson, S.L. Comprehensive Characterization of the Attenuated Double Auxotroph Mycobacterium tuberculosisΔleudΔpanCD as an Alternative to H37Rv. Front. Microbiol. 2019, 10, 1922. [Google Scholar] [CrossRef] [PubMed]
- Voskuil, M.I.; Schnappinger, D.; Visconti, K.C.; Harrell, M.I.; Dolganov, G.M.; Sherman, D.R.; Schoolnik, G.K. Inhibition of Respiration by Nitric Oxide Induces a Mycobacterium tuberculosis Dormancy Program. J. Exp. Med. 2003, 198, 705–713. [Google Scholar] [CrossRef]
- Tullius, M.V.; Harth, G.; Horwitz, M.A. Glutamine Synthetase GlnA1 Is Essential for Growth of Mycobacterium tuberculosis in Human THP-1 Macrophages and Guinea Pigs. Infect. Immun. 2003, 71, 3927–3936. [Google Scholar] [CrossRef]
- Nozaki, Y.; Hasegawa, Y.; Ichiyama, S.; Nakashima, I.; Shimokata, K. Mechanism of Nitric Oxide-Dependent Killing of Mycobacterium bovis BCG in Human Alveolar Macrophages. Infect. Immun. 1997, 65, 3644–3647. [Google Scholar] [CrossRef] [PubMed]
- Denis, M. Interferon-Gamma-Treated Murine Macrophages Inhibit Growth of Tubercle Bacilli via the Generation of Reactive Nitrogen Intermediates. Cell. Immunol. 1991, 132, 150–157. [Google Scholar] [CrossRef]
- Hutter, B.; Dick, T. Up-Regulation of NarX, Encoding a Putative “fused Nitrate Reductase” in Anaerobic Dormant Mycobacterium bovis BCG. FEMS Microbiol. Lett. 1999, 178, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Boon, C.; Dick, T. Mycobacterium bovis BCG Response Regulator Essential for Hypoxic Dormancy. J. Bacteriol. 2002, 184, 6760–6767. [Google Scholar] [CrossRef]
- Slavchev, G.; Michailova, L.; Markova, N. Stress-Induced L-Forms of Mycobacterium bovis: A Challenge to Survivability. New Microbiol. 2013, 36, 157–166. [Google Scholar]
- Rodriguez, J.G.; Burbano, C.S.; Nuñez, C.; González, C.E.; Zambrano, M.M.; García, M.J.; Del Portillo, P. Rv3134c/DevR/DevS Operon of Mycobacterium bovis BCG Is Differentially Transcribed under “in vitro” Stress Conditions. Tuberculosis 2008, 88, 273–282. [Google Scholar] [CrossRef]
- Malherbe, S.T.; Shenai, S.; Ronacher, K.; Loxton, A.G.; Dolganov, G.; Kriel, M.; Van, T.; Chen, R.Y.; Warwick, J.; Via, E.; et al. Persisting PET-CT Lesion Activity and Mycobacterium tuberculosis mRNA after Pulmonary Tuberculosis Cure. Nat. Med. 2016, 22, 1094–1100. [Google Scholar] [CrossRef] [PubMed]
- Scanga, C.A.; Mohan, V.P.; Joseph, H.; Yu, K.; Chan, J.; Flynn, J.L. Reactivation of Latent Tuberculosis: Variations on the Cornell Murine Model. Infect. Immun. 1999, 67, 4531–4538. [Google Scholar] [CrossRef] [PubMed]
- Palacios, J.J.; Navarro, Y.; Romero, B.; Penedo, A.; Menéndez González, Á.; Pérez Hernández, M.D.; Fernández-Verdugo, A.; Copano, F.; Torreblanca, A.; Bouza, E.; et al. Molecular and Epidemiological Population-Based Integrative Analysis of Human and Animal Mycobacterium bovis Infections in a Low-Prevalence Setting. Vet. Microbiol. 2016, 195, 30–36. [Google Scholar] [CrossRef]
- Larsen, M.V.; Sørensen, I.J.; Thomsen, V.; Ravn, P. Re-Activation of Bovine Tuberculosis in a Patient Treated with Infliximab. Eur. Respir. J. 2008, 32, 229–231. [Google Scholar] [CrossRef]
- Cosivi, O.; Grange, J.M.; Daborn, C.J.; Raviglione, M.C.; Fujikura, T.; Cousins, D.; Robinson, R.A.; Huchzermeyer, H.F.A.K.; De Kantor, I.; Meslin, F.X. Zoonotic Tuberculosis Due to Mycobacterium bovis in Developing Countries. Emerg. Infect. Dis. 1998, 4, 59–70. [Google Scholar] [CrossRef]
- Shahmohammadi, S.; Saffar, M.J.; Rezai, M.S. BCG-Osis after BCG Vaccination in Immunocompromised Children: Case Series and Review. J. Pediatr. Rev. 2014, 2, 62–74. [Google Scholar] [CrossRef]
- Bhola, R.K.; Sarangi, R.; Dey, P.; Samal, P. Disseminated BCG-Osis with Haemophagocytosis, Tubercular Bacteraemia, and Unusual Haematological Findings with Its Haematology Analyser-Based Expression. J. Hematop. 2018, 11, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Gibson, S.E.R.; Harrison, J.; Cox, J.A.G. Modelling a Silent Epidemic: A Review of the in vitro Models of Latent Tuberculosis. Pathogens 2018, 7, 88. [Google Scholar] [CrossRef]
- Alnimr, A.M. Dormancy Models for Mycobacterium tuberculosis: A Minireview. Brazilian J. Microbiol. 2015, 46, 641–647. [Google Scholar] [CrossRef]
- Kundu, M.; Basu, J. Applications of Transcriptomics and Proteomics for Understanding Dormancy and Resuscitation in Mycobacterium tuberculosis. Front. Microbiol. 2021, 12, 642487. [Google Scholar] [CrossRef]
- Gideon, H.P.; Wilkinson, K.A.; Rustad, T.R.; Oni, T.; Guio, H.; Kozak, R.A.; Sherman, D.R.; Meintjes, G.; Behr, M.A.; Vordermeier, H.M.; et al. Hypoxia Induces an Immunodominant Target of Tuberculosis Specific T Cells Absent from Common BCG Vaccines. PLoS Pathog. 2010, 6, e1001237. [Google Scholar] [CrossRef]
- Deb, C.; Lee, C.M.; Dubey, V.S.; Daniel, J.; Abomoelak, B.; Sirakova, T.D.; Pawar, S.; Rogers, L.; Kolattukudy, P.E. A Novel in vitro Multiple-Stress Dormancy Model for Mycobacterium tuberculosis Generates a Lipid-Loaded, Drug-Tolerant, Dormant Pathogen. PLoS ONE 2009, 4, e6077. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.; Streur, T.L.; Aldwell, F.E.; Cook, G.M. Intracellular PH Regulation by Mycobacterium smegmatis and Mycobacterium bovis BCG. Microbiology. 2001, 147, 1017–1024. [Google Scholar] [CrossRef] [PubMed]
- Gallant, J.L.; Viljoen, A.J.; Van Helden, P.D.; Wiid, I.J.F. Glutamate Dehydrogenase Is Required by Mycobacterium bovis BCG for Resistance to Cellular Stress. PLoS ONE 2016, 11, 147706. [Google Scholar] [CrossRef] [PubMed]
- Weiss, D.J.; Evanson, O.A.; Moritz, A.; Deng, M.Q.; Abrahamsen, M.S. Differential Responses of Bovine Macrophages to Mycobacterium avium Subsp. paratuberculosis and Mycobacterium avium Subsp. avium. Infect. Immun. 2002, 70, 5556–5561. [Google Scholar] [CrossRef] [PubMed]
- Piercy, J.; Werling, D.; Coffey, T.J. Differential Responses of Bovine Macrophages to Infection with Bovine-Specific and Non-Bovine Specific Mycobacteria. Tuberculosis 2007, 87, 415–420. [Google Scholar] [CrossRef]
- Medley, J.; Goff, A.; Bettencourt, P.J.G.; Dare, M.; Cole, L.; Cantillon, D.; Waddell, S.J. Dissecting the Mycobacterium bovis BCG Response to Macrophage Infection to Help Prioritize Targets for Anti-Tuberculosis Drug and Vaccine Discovery. Vaccines 2022, 10, 113. [Google Scholar] [CrossRef]
- Zhan, X.; Yuan, W.; Zhou, Y.; Ma, R.; Ge, Z. Small RNA Sequencing and Bioinformatics Analysis of RAW264.7-Derived Exosomes after Mycobacterium bovis Bacillus Calmette-Guérin Infection. BMC Genom. 2022, 23, 355. [Google Scholar] [CrossRef]
- Lin, W.; Xin, T. The Influence of Gene Rv3671c in Mycobacterium bovis to Its Replication and Acid Resistance. bioRxiv 2020. [Google Scholar] [CrossRef]
- Ma, R.; Farrell, D.; Gonzalez, G.; Browne, J.A.; Nakajima, C.; Suzuki, Y.; Gordon, S.V. The TbD1 Locus Mediates a Hypoxia-Induced Copper Response in Mycobacterium bovis. Front. Microbiol. 2022, 13, 817952. [Google Scholar] [CrossRef]
- Malone, K.M.; Rue-Albrecht, K.; Magee, D.A.; Conlon, K.; Schubert, O.T.; Nalpas, N.C.; Browne, J.A.; Smyth, A.; Gormley, E.; Aebersold, R.; et al. Comparative ’omics Analyses Differentiate Mycobacterium tuberculosis and Mycobacterium bovis and Reveal Distinct Macrophage Responses to Infection with the Human and Bovine Tubercle Bacilli. Microb. Genom. 2018, 4, 163. [Google Scholar] [CrossRef] [PubMed]
- Gibson, A.J.; Passmore, I.J.; Faulkner, V.; Xia, D.; Nobeli, I.; Stiens, J.; Willcocks, S.; Clark, T.G.; Sobkowiak, B.; Werling, D.; et al. Probing Differences in Gene Essentiality Between the Human and Animal Adapted Lineages of the Mycobacterium tuberculosis Complex Using TnSeq. Front. Vet. Sci. 2021, 8, 717. [Google Scholar] [CrossRef] [PubMed]
- Jones, G.J.; Pirson, C.; Gideon, H.P.; Wilkinson, K.A.; Sherman, D.R.; Wilkinson, R.J.; Hewinson, R.G.; Vordermeier, H.M. Immune Responses to the Enduring Hypoxic Response Antigen Rv0188 Are Preferentially Detected in Mycobacterium bovis Infected Cattle with Low Pathology. PLoS ONE 2011, 6, e21371. [Google Scholar] [CrossRef]
- Larsen, M.H.; Lacourciere, K.; Parker, T.M.; Kraigsley, A.; Achkar, M.; Adams, L.B.; Dupnik, K.M.; Hall-stoodley, L.; Kanipe, C.; Kurtz, S.L.; et al. The Many Hosts of Mycobacteria 8 (MHM8): A Conference Report. Tuberculosis 2020, 8, 101914. [Google Scholar] [CrossRef] [PubMed]
- León, A.D.; Zumárraga, M.J.; Jiménez Oropeza, R.; Gioffré, A.K.; Bernardelli, A.; Orozco Estévez, H.; Cataldi, A.A.; Hernández Pando, R. Mycobacterium bovis with Different Genotypes and from Different Hosts Induce Dissimilar Immunopathological Lesions in a Mouse Model of Tuberculosis. Clin. Exp. Immunol. 2009, 157, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Phillips, C.J.C.; Foster, C.R.W.; Morris, P.A.; Teverson, R. The Transmission of Mycobacterium bovis Infection to Cattle. Res. Vet. Sci. 2003, 74, 1–15. [Google Scholar] [CrossRef]
- Alvarez Herrera, A. Are Cattle a Surrogate Model for Pathogenic Mycobacterial Latent Infection? Mycobact. Dis. 2014, 4, 4–5. [Google Scholar] [CrossRef] [Green Version]
- Ippolito, D.; Boniotti, M.B.; Fiasconaro, M.; Fontana, S.; Boifava, M.; Ciarello, F.P.; Amato, B.; Pacciarini, M.; Presti, V.D.M.L. Development and Evaluation of a Multi-Antigen Serological Assay for the Intra-Vitam Diagnosis of Tuberculosis Caused by Mycobacterium bovis in Pigs. J. Immunol. Methods 2022, 503, 113234. [Google Scholar] [CrossRef]
- Nol, P.; Wehtje, M.E.; Bowen, R.A.; Robbe-austerman, S.; Thacker, T.C.; Lantz, K.; Rhyan, J.C.; Baeten, L.A.; Juste, A.; Sevilla, I.A.; et al. Pathogens E Ff Ects of Inactivated Mycobacterium bovis Vaccination on Molokai-Origin Wild Pigs Experimentally Infected with Virulent M. Bovis. Pathogens 2020, 9, 199. [Google Scholar] [CrossRef]
- Bolin, C.A.; Whipple, D.L.; Khanna, K.V.; Risdahl, J.M.; Peterson, P.K.; Molitor, T.W. Infection of Swine with Mycobacterium bovis as a Model of Human Tuberculosis. J. Infect. Dis. 1997, 176, 1559–1566. [Google Scholar] [CrossRef]
- Gallagher, J.; Clifton-Hadley, R.S. Tuberculosis in Badgers; a Review of the Disease and Its Significance for Other Animals. Res. Vet. Sci. 2000, 69, 203–217. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, J.; Monies, R.; Gavier-Widen, M.; Rule, B. Role of Infected, Non-Diseased Badgers in the Pathogenesis of Tuberculosis in the Badger. Vet. Rec. 1998, 142, 710–714. [Google Scholar] [CrossRef] [PubMed]
- Kupz, A.; Zedler, U.; Stäber, M.; Kaufmann, S.H.E. A Mouse Model of Latent Tuberculosis Infection to Study Intervention Strategies to Prevent Reactivation. PLoS ONE 2016, 11, e0158849. [Google Scholar] [CrossRef] [PubMed]
- Karakousis, P.C.; Yoshimatsu, T.; Lamichhane, G.; Woolwine, S.C.; Nuermberger, E.L.; Grosset, J.; Bishai, W.R. Dormancy Phenotype Displayed by Extracellular Mycobacterium tuberculosis within Artificial Granulomas in Mice. J. Exp. Med. 2004, 200, 647–657. [Google Scholar] [CrossRef]
- Zhan, L.; Tang, J.; Lin, S.; Xu, Y.; Xu, Y.; Qin, C. Prophylactic Use of Ganoderma Lucidum Extract May Inhibit Mycobacterium tuberculosis Replication in a New Mouse Model of Spontaneous Latent Tuberculosis Infection. Front. Microbiol. 2016, 6, 1490. [Google Scholar] [CrossRef]
- Rhoades, E.R. Progression of Chronic Pulmonary Tuberculosis in Mice Aerogenically Infected with Virulent Mycobacterium tuberculosis. Tuber. Lung Dis. 1997, 78, 57–66. [Google Scholar] [CrossRef]
- Lu, J.B.; Chen, B.W.; Wang, G.Z.; Fu, L.L.; Shen, X.B.; Su, C.; Du, W.X.; Yang, L.; Xu, M. Recombinant Tuberculosis Vaccine AEC/BC02 Induces Antigen-Specific Cellular Responses in Mice and Protects Guinea Pigs in a Model of Latent Infection. J. Microbiol. Immunol. Infect. 2015, 48, 597–603. [Google Scholar] [CrossRef] [Green Version]
- Clark, S.; Hall, Y.; Williams, A. Animal Models of Tuberculosis: Guinea Pigs. Cold Spring Harb. Perspect. Med. 2015, 5, a018572. [Google Scholar] [CrossRef]
- Chambers, M.A.; Williams, A.; Gavier-Widén, D.; Whelan, A.; Hughes, C.; Hall, G.; Lever, M.S.; Marsh, P.D.; Hewinson, R.G. A Guinea Pig Model of Low-Dose Mycobacterium bovis Aerogenic Infection. Vet. Microbiol. 2001, 80, 213–226. [Google Scholar] [CrossRef]
- Chandran, A.; Williams, K.; Mendum, T.; Stewart, G.; Clark, S.; Zadi, S.; McLeod, N.; Williams, A.; Villarreal-Ramos, B.; Vordermeier, M.; et al. Development of a Diagnostic Compatible BCG Vaccine against Bovine Tuberculosis. Sci. Rep. 2019, 9, 17791. [Google Scholar] [CrossRef]
- Kashino, S.S.; Napolitano, D.R.; Skobe, Z.; Campos-Neto, A. Guinea Pig Model of Mycobacterium tuberculosis Dormant Infection. Microbes Infect. 2008, 10, 1469–1476. [Google Scholar] [CrossRef]
- Lenaerts, A.J.; Chapman, P.L.; Orme, I.M. Statistical Limitations to the Cornell Model of Latent Tuberculosis Infection for the Study of Relapse Rates. Tuberculosis 2004, 84, 361–364. [Google Scholar] [CrossRef] [PubMed]
- Zhan, L.; Tang, J.; Sun, M.; Qin, C. Animal Models for Tuberculosis in Translational and Precision Medicine. Front. Microbiol. 2017, 8, 717. [Google Scholar] [CrossRef] [PubMed]
- Bapat, P.R.; Husain, A.A.; Daginawala, H.F.; Agrawal, N.P.; Panchbhai, M.S.; Satav, A.R.; Taori, G.M.; Kashyap, R.S. The Assessment of Cytokines in Quantiferon Supernatants for the Diagnosis of Latent TB Infection in a Tribal Population of Melghat, India. J. Infect. Public Health 2015, 8, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Sutherland, J.S.; Hill, P.C.; Adetifa, I.M.; de Jong, B.C.; Donkor, S.; Joosten, S.A.; Opmeer, L.; Haks, M.C.; Ottenhoff, T.H.M.; Adegbola, R.A.; et al. Identification of Probable Early-Onset Biomarkers for Tuberculosis Disease Progression. PLoS ONE 2011, 6, e25230. [Google Scholar] [CrossRef]
- Sloot, R.; Schim van der Loeff, M.F.; van Zwet, E.W.; Haks, M.C.; Keizer, S.T.; Scholing, M.; Ottenhoff, T.H.M.; Borgdorff, M.W.; Joosten, S.A. Biomarkers Can Identify Pulmonary Tuberculosis in HIV-Infected Drug Users Months Prior to Clinical Diagnosis. EbioMedicine 2015, 2, 172–179. [Google Scholar] [CrossRef]
- Chegou, N.N.; Black, G.F.; Kidd, M.; van Helden, P.D.; Walzl, G. Host Markers in Quantiferon Supernatants Differentiate Active TB from Latent TB Infection: Preliminary Report. BMC Pulm. Med. 2009, 9, 21. [Google Scholar] [CrossRef]
- Kim, S.Y.; Park, M.S.; Kim, Y.S.; Kim, S.K.; Chang, J.; Lee, H.J.; Cho, S.N.; Kang, Y.A. The Responses of Multiple Cytokines Following Incubation of Whole Blood from TB Patients, Latently Infected Individuals and Controls with the TB Antigens ESAT-6, CFP-10 and TB7.7. Scand. J. Immunol. 2012, 76, 580–586. [Google Scholar] [CrossRef]
- Suzukawa, M.; Akashi, S.; Nagai, H.; Nagase, H.; Nakamura, H.; Matsui, H.; Hebisawa, A.; Ohta, K. Combined Analysis of IFN-γ, IL-2, IL-5, IL-10, IL-1RA and MCP-1 in QFT Supernatant Is Useful for Distinguishing Active Tuberculosis from Latent Infection. PLoS ONE 2016, 11, e0152483. [Google Scholar] [CrossRef]
- Singh, G.; Yadav, M.; Ghosh, C.; Rathore, J.S. Bacterial Toxin-Antitoxin Modules: Classification, Functions, and Association with Persistence. Curr. Res. Microb. Sci. 2021, 2, 100047. [Google Scholar] [CrossRef]
- Boon, C.; Li, R.; Qi, R.; Dick, T. Proteins of Mycobacterium bovis BCG Induced in the Wayne Dormancy Model. J. Bacteriol. 2001, 183, 2672–2676. [Google Scholar] [CrossRef] [PubMed]
- Leistikow, R.L.; Morton, R.A.; Bartek, I.L.; Frimpong, I.; Wagner, K.; Voskuil, M.I. The Mycobacterium tuberculosis DosR Regulon Assists in Metabolic Homeostasis and Enables Rapid Recovery from Nonrespiring Dormancy. J. Bacteriol. 2010, 192, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, L.; Marín, D.; Franken, K.L.M.C.; Ottenhoff, T.H.M.; Barrera, L.F. Potential of DosR and Rpf Antigens from Mycobacterium tuberculosis to Discriminate between Latent and Active Tuberculosis in a Tuberculosis Endemic Population of Medellin Colombia. BMC Infect. Dis. 2018, 18, 26. [Google Scholar] [CrossRef]
- Black, G.F.; Thiel, B.A.; Ota, M.O.; Parida, S.K.; Adegbola, R.; Boom, W.H.; Dockrell, H.M.; Franken, K.L.M.C.; Friggen, A.H.; Hill, P.C.; et al. Immunogenicity of Novel DosR Regulon-Encoded Candidate Antigens of Mycobacterium tuberculosis in Three High-Burden Populations in Africa. Clin. Vaccine Immunol. 2009, 16, 1203–1212. [Google Scholar] [CrossRef]
- Keren, I.; Minami, S.; Rubin, E.; Lewis, K. Characterization and Transcriptome Analysis of Mycobacterium tuberculosis Persisters. mBio 2011, 2, 11. [Google Scholar] [CrossRef] [PubMed]
- O’ Toole, R.; Smeulders, M.J.; Blokpoel, M.C.; Kay, E.J.; Lougheed, K.; Williams, H.D. A Two-Component Regulator of Universal Stress Protein Expression and Adaptation to Oxygen Starvation in Mycobacterium smegmatis. J. Bacteriol. 2003, 185, 1543–1554. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.R.; Robertson, B.D.; Young, D.B. Tuberculosis: A Problem with Persistence. Nat. Rev. Microbiol. 2003, 1, 97–105. [Google Scholar] [CrossRef]
- Flores-Valdez, M.A.; Aceves-Sánchez, M.D.J.; Peterson, E.J.R.; Baliga, N.; Bravo-Madrigal, J.; De la Cruz-Villegas, M.Á.; Ares, M.A.; Born, S.; Voskuil, M.; Pérez-Padilla, N.A.; et al. Transcriptional Portrait of M. Bovis BCG during Biofilm Production Shows Genes Differentially Expressed during Intercellular Aggregation and Substrate Attachment. Sci. Rep. 2020, 10, 12578. [Google Scholar] [CrossRef]
- Hozumi, H.; Tsujimura, K.; Yamamura, Y.; Seto, S.; Uchijima, M.; Nagata, T.; Miwa, S.; Hayakawa, H.; Fujisawa, T.; Hashimoto, D.; et al. Immunogenicity of Dormancy-Related Antigens in Individuals Infected with Mycobacterium tuberculosis in Japan. Int. J. Tuberc. Lung Dis. 2013, 17, 818–824. [Google Scholar] [CrossRef]
- Demaio, J.; Zhang, Y.; Ko, C.; Youngt, D.B.; Bishai, W.R. A Stationary-Phase Stress-Response Sigma Factor from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 1996, 93, 2790–2794. [Google Scholar] [CrossRef]
- Michele, T.M.; Ko, C. Exposure to Antibiotics Induces Expression of the Mycobacterium tuberculosis SigF Gene: Implications for Chemotherapy against Mycobacterial Persistors. Society 2010, 43, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.P.; Lee, J.-H.; Bishai, W.R.; Colantuoni, C.; Karakousis, P.C. Mycobacterium tuberculosis SigF Regulates Genes Encoding Cell Wall-Associated Proteins and Directly Regulates the Transcriptional Regulatory Gene PhoY1. J. Bacteriol. 2007, 189, 4234–4242. [Google Scholar] [CrossRef] [PubMed]
- Sherman, R.A.; Shimoda, K.J. Tuberculosis Tracking: Determining the Frequency of the Booster Effect in Patients and Staff. Am. J. Infect. Control 2001, 29, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Jee, B.; Singh, Y.; Yadav, R.; Lang, F. Small Heat Shock Protein16.3 of Mycobacterium tuberculosis: After Two Decades of Functional Characterization. Cell. Physiol. Biochem. 2018, 49, 368–380. [Google Scholar] [CrossRef]
- Rodrigue, S.; Provvedi, R.; Jacques, P.É.; Gaudreau, L.; Manganelli, R. The σ Factors of Mycobacterium tuberculosis. FEMS Microbiol. Rev. 2006, 30, 926–941. [Google Scholar] [CrossRef] [PubMed]
- Sachdeva, P.; Misra, R.; Tyagi, A.K.; Singh, Y. The Sigma Factors of Mycobacterium tuberculosis: Regulation of the Regulators. FEBS J. 2010, 277, 605–626. [Google Scholar] [CrossRef]
- Manganelli, R.; Dubnau, E.; Tyagi, S.; Kramer, F.R.; Smith, I. Differential Expression of 10 Sigma Factor Genes in Mycobacterium tuberculosis. Mol. Microbiol. 1999, 30, 715–724. [Google Scholar] [CrossRef]
- Cuerda, M.X.; Colombatti, M.A.; Gravisaco, M.J.; Marfil, M.J.; Barandiaran, S.; Sevilla, I.A.; Garrido, J.M.; Moyano, R.D.; Zumárraga, M.J.; Romano, M.I.; et al. Pathogenesis of Domestic Pigs Submitted to Mycobacterial Sensitizations Previous to Experimental Infection with Mycobacterium bovis. Span. J. Agric. Res. 2022, 20, e0502. [Google Scholar] [CrossRef]
Figure 1.
Latent TB stage and underlying bacterial phenotypes. Latent TB is speculated to encompass several bacterial phenotypes, under the broad heading of persister bacteria. The subpopulations of persister bacilli include viable but non-replicating (VBNR) bacilli, viable but non-culturable VBNC bacilli, and dormant bacilli. These have varying phenotypic characteristics and may vary with each TB stage. (Original figure).
Figure 1.
Latent TB stage and underlying bacterial phenotypes. Latent TB is speculated to encompass several bacterial phenotypes, under the broad heading of persister bacteria. The subpopulations of persister bacilli include viable but non-replicating (VBNR) bacilli, viable but non-culturable VBNC bacilli, and dormant bacilli. These have varying phenotypic characteristics and may vary with each TB stage. (Original figure).


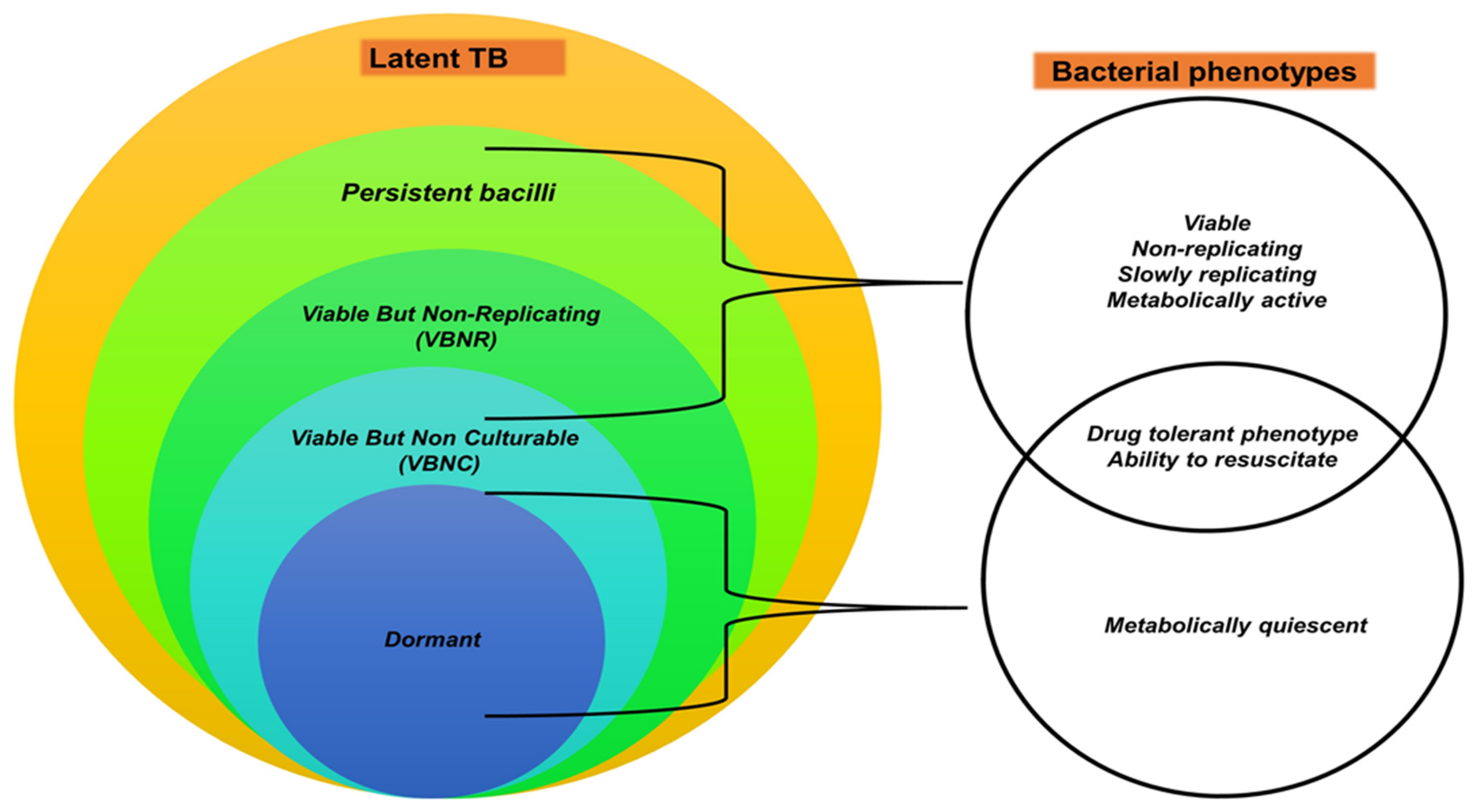{kind=link}
Table 1.
Mycobacterium tuberculosis complex pathogens and their hosts.
| MTBC Pathogens | Primary Hosts | Secondary/Sporadic Hosts |
|---|---|---|
| M. bovis | African buffaloes (Syncerus caffer) [44,55,56], Asian elephants (Elephas maximus) [57,58,59,60,61,62], bushbucks (Tragelaphus scriptus) [28,63,64], bush pigs [40,65,66] (Potamochoerus porcus), cattle [18,67,68,69], chacma baboons (Papio ursinus) [28,70,71], cheetahs (Acinonyx jubatus) [70,72], common duiker (Syvicapra grimmia) [12,73], African elephants (Loxodonta africana) [28,59,74], eland (Taurotragus oryx) [28,40,66], giraffe (Giraffa camelopardalis) [75], european badger (Meles meles) [76,77,78,79], honey badger (Mellivora capensis) [12,28,80], greater kudu (Tragelaphus strepsiceros) [72,81], hippopotamus (Hippopotamus amphibius) [82,83], impala (Aepyceros melampus) [28,40,84], African leopards (Panthera pardus) [28,35,85], African lions (Panthera leo) [6,11,28,86,87,88,89], banded mongoose (Mungos mungo) [90,91], nyala (Tragelaphus angasii) [28,92] black rhinoceros (Diceros bicornis) [6,93,94,95], red fox (Vulpes vulpes) [96], spotted genet (Genetta tigrine) [28,65,81], springbok (Antidorcas marsupialis) [12,28], common warthog (Phacochoerus africanus) [28,97,98,99], wild boars (Sus scrofa) [96], African wild dog (Lycaon pictus) [100,101,102,103], blue wildebeest (Connochaetes taurinus) [104,105], wild deer (Capreolus capreolus, Cervus elaphus) [96], white rhinoceros (Ceratotherium simum) [35,95,104,106] | Humans [17,19,43], cats [107] |
| M. tuberculosis | Humans [1,19,28] | Cattle [108,109], non-human primates [110,111], dogs [112,113,114], cats [115], zoo animals e.g., black and white rhinos [116], African elephants, and Asian elephants [117,118,119], goats [120] |
| Mycobacterium africanum | Humans [121,122,123] | Cattle [124], rock hyraxes (Procavia capensis) [29,125] |
| Mycobacterium pinnipedii | Seals (pinnipedts) [126] | Humans [127,128], cattle [129] |
| Mycobacterium caprae | Goats, sheep, and pigs [130,131] | Humans [132] |
| Mycobacterium microti | Cats, pigs [133], bank vole, wood mouse, shrew [134] | Humans [135] |
| Mycobacterium orygis | Antelope, deer, waterbuck [136], oryxes, gazelles [137], | Humans [138] |
| Mycobacterium canettii | Humans [29,139,140,141] | - |
| Mycobacterium mungi | Banded mongooses [142] | - |
| Mycobacterium suricattae | Meerkats (Suricata suricatta) [143] | - |
| Chimpanzee bacillus | Chimpanzee (Pan troglodytes) [16] | - |
| Dassie bacillus | Rock hyraxes [14,144], meerkats [143,145] | - |
Table 2.
Hypothetical stages of mycobacterial infection and host–pathogen interactions in domestic and wild animals. Infection stages may vary in different species depending on the host’s immune response and risk factors, as well as strain and infecting dose of Mycobacterium bovis. (Adapted and adjusted from [25,204]).
Table 2.
Hypothetical stages of mycobacterial infection and host–pathogen interactions in domestic and wild animals. Infection stages may vary in different species depending on the host’s immune response and risk factors, as well as strain and infecting dose of Mycobacterium bovis. (Adapted and adjusted from [25,204]).
| bTB Stage | Clinical Signs | Host–Pathogen Interactions | |
|---|---|---|---|
| Uninfected | Absent |
| |
| Infected | Cleared infection | Absent |
|
| Latent infection | Absent |
| |
| Subclinical infection | Absent |
| |
| Active TB disease | Present |
| |
Table 3.
Hypothetical infection stages and associated antemortem and postmortem test results for categorising TB stages.
Table 3.
Hypothetical infection stages and associated antemortem and postmortem test results for categorising TB stages.
| Animal TB Stage (a) | Antemortem Tests (b,c) | Postmortem Tests (d) | |||
|---|---|---|---|---|---|
| TST | IGRA | Bacterial Culture | Tissue Histopathology Consistent with bTB (Yes/No) | ||
| Uninfected | Negative | Negative | Negative | No | |
| Infected | Cleared infection | Negative (innate immune system clearance) Positive (adaptive system immune clearance) | Positive for a short period before waning (adaptive immune clearance) | Negative | No |
| Latent infection | Positive | Positive | Negative | No | |
| Subclinical infection | Positive | Positive | Positive | Yes (early-stage granulomas) | |
| Active disease | Positive | Positive | Positive | Yes (multifocal and/or confluent late-stage granulomas | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ncube, P.; Bagheri, B.; Goosen, W.J.; Miller, M.A.; Sampson, S.L. Evidence, Challenges, and Knowledge Gaps Regarding Latent Tuberculosis in Animals. Microorganisms 2022, 10, 1845. https://doi.org/10.3390/microorganisms10091845
AMA Style
Ncube P, Bagheri B, Goosen WJ, Miller MA, Sampson SL. Evidence, Challenges, and Knowledge Gaps Regarding Latent Tuberculosis in Animals. Microorganisms. 2022; 10(9):1845. https://doi.org/10.3390/microorganisms10091845
Chicago/Turabian StyleNcube, Pamela, Bahareh Bagheri, Wynand Johan Goosen, Michele Ann Miller, and Samantha Leigh Sampson. 2022. "Evidence, Challenges, and Knowledge Gaps Regarding Latent Tuberculosis in Animals" Microorganisms 10, no. 9: 1845. https://doi.org/10.3390/microorganisms10091845
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.