
Hao1 Is Not a Pathogenic Factor for Ectopic Ossifications but Functions to Regulate the TCA Cycle In Vivo

 ,
,  , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Hao1 Expression Significantly Decreased with Osteoblastic Differentiation
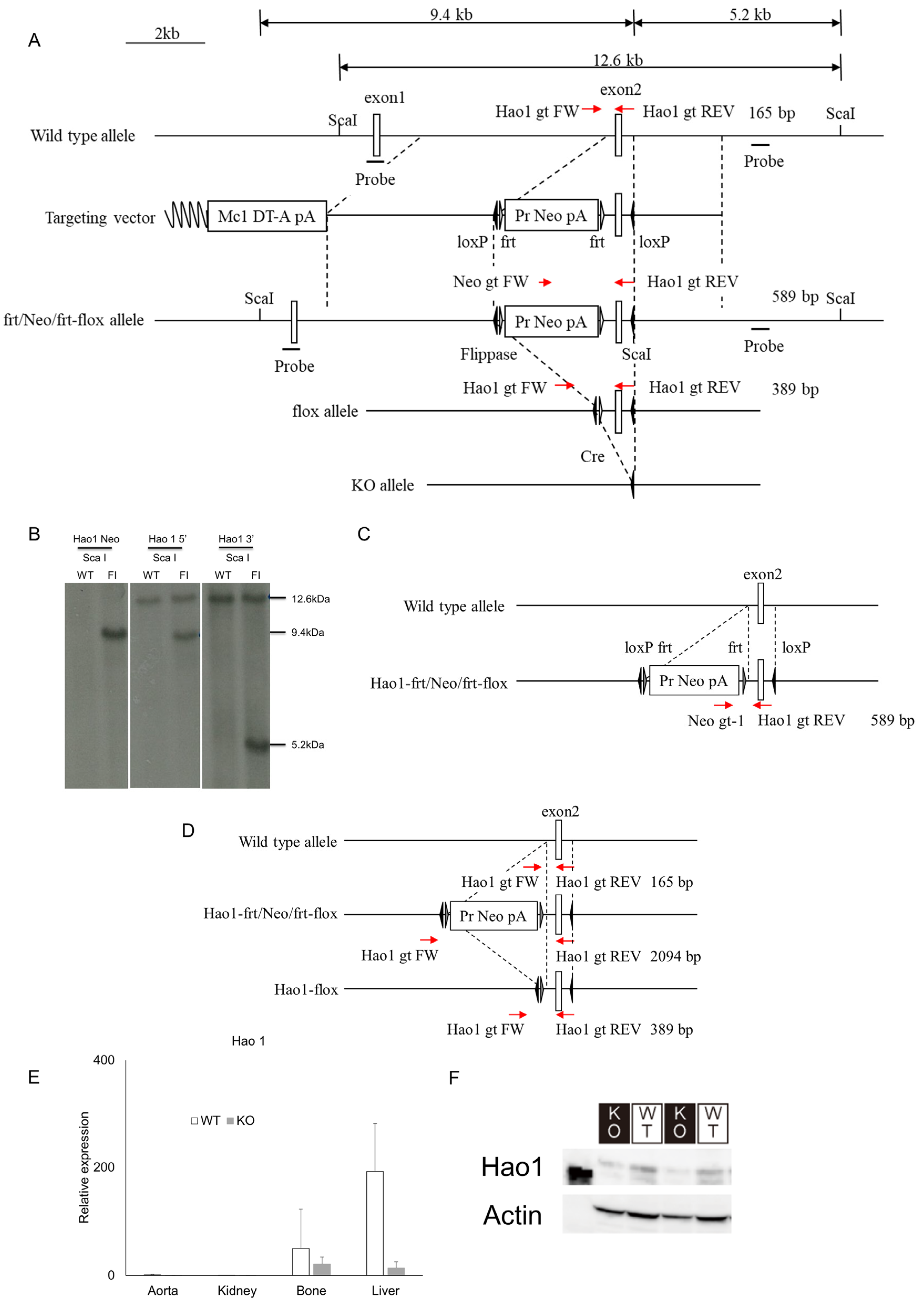
2.2. Generation of Hao1 Knockout Mice
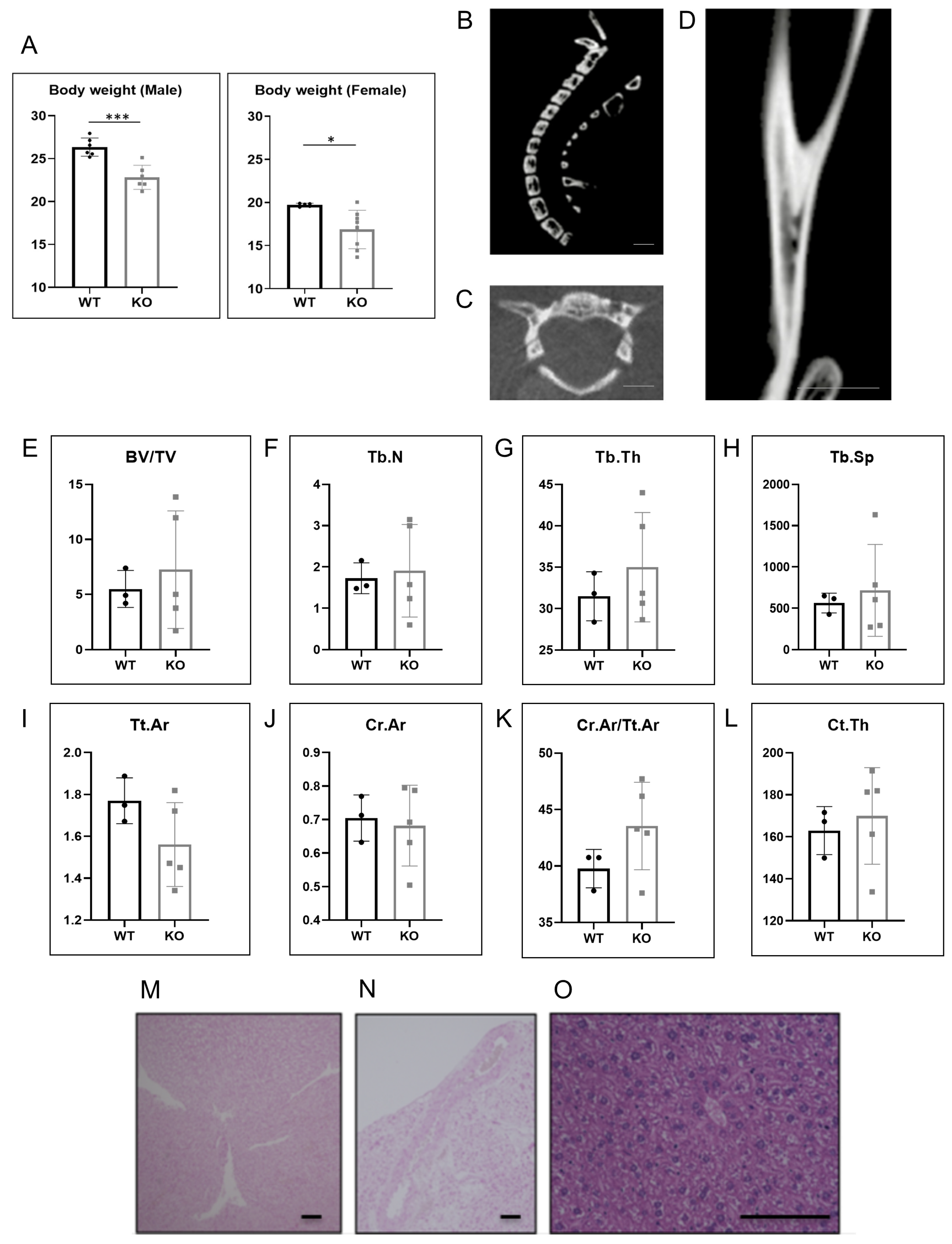
2.3. Hao1 KO Mice Did Not Show Ectopic Ossification or Calcification
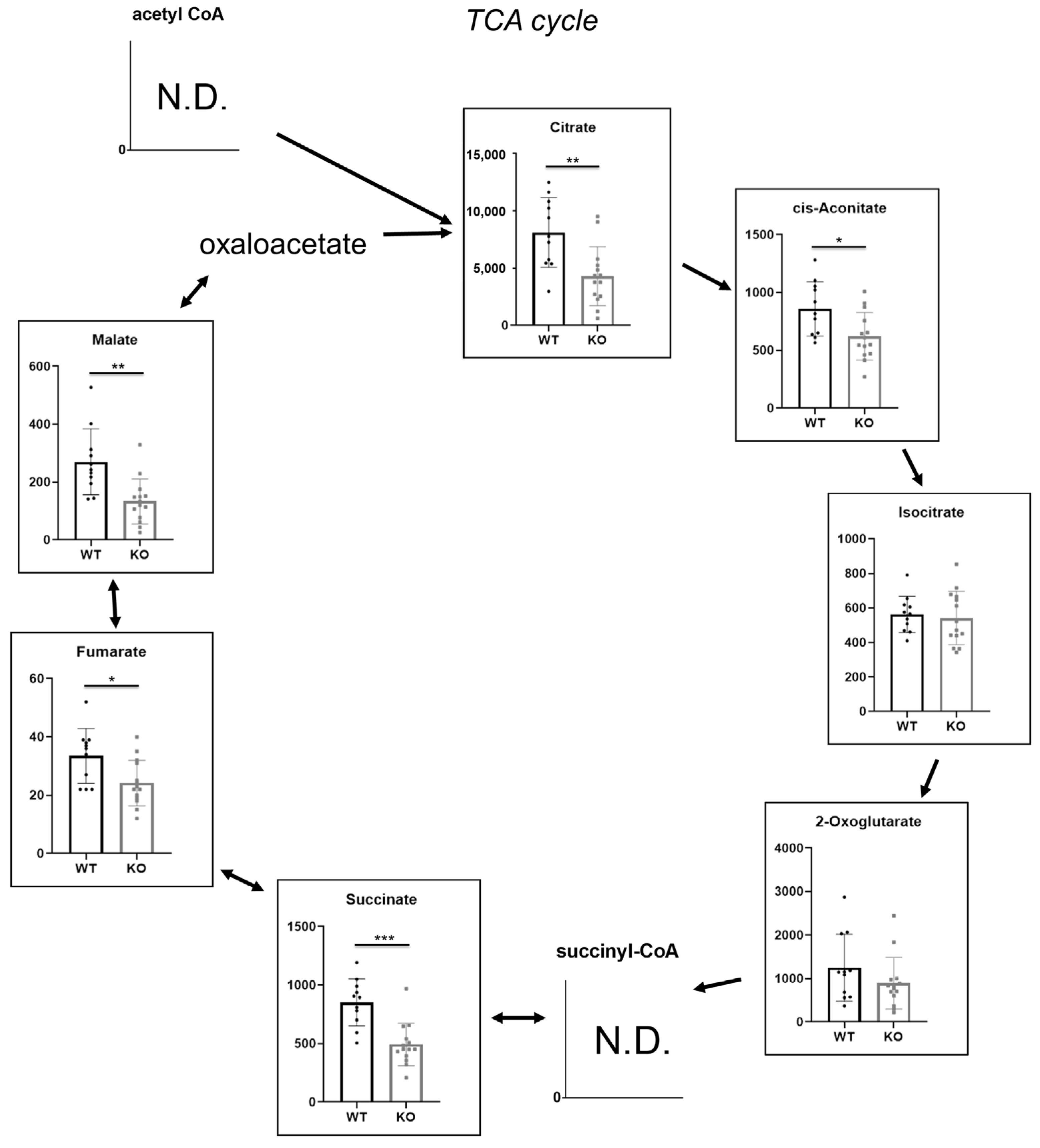
2.4. Hao1 Functions in Regulation of the TCA Cycle
3. Discussion
4. Materials and Methods
4.1. Mice
4.2. Quantitative Real-Time PCR
- Actb—forward: 5′-TGAGAGGGAAATCGTGCGTGAC-3′
- Actb—reverse: 5′-AAGAAGGAAGGCTGGAAAAGAG-3′
- Alp—forward: 5′-CACCATTTTTAGTACTGGCCATCG-3′
- Alp—reverse: 5′-GCTACATTGGTGTTGAGCTTTTGG-3′
- Hao1—forward: 5′-CTTGCTGAATATGTGGCACAAGC-3′
- Hao1—reverse: 5′-TAACAGCTTCCTTGGCATCATCA-3′
- Rspo2—forward: 5′-AGCCAGCAAAAGACACAATACCAT-3′
- Rspo2—reverse: 5′-TTCTCTTTTGCCTTTGGTGTTCTCA-3′
- Eif3e—forward: 5′-CAGAACCAATCGTGAAGATGTTTG-3′
- Eif3e—reverse: 5′-TCTTGCCTAAACCCATGTTTGTCT-3′
- Ccdc91—forward: 5′-CCATCGAGAAGCAATATGTGTCTG-3′
- Ccdc91—reverse: 5′-TGAGTCAAAGCTTCCTGGATTTTC-3′
- Eif3h—forward: 5′-GGATAAGCACGAATTGCTCAGTCT-3′
- Eif3h—reverse: 5′-GCGCATGTACGTGTTGTATTTGAT-3′
- Cdc5l—forward: 5′-TGTGGGAGGAATGCTACAGTCA-3′
- Cdc5l—reverse: 5′-TCTGTTGTCATGTGACCCCTGT-3′
- Rsph9—forward: 5′-GTTGCAGAAGGTTAACGAAGGAGA-3′
- Rsph9—reverse: 5′-AGCTACAGCCTTGTCAATCTGGTC-3′
- Tmem151b—forward: 5′-CTGGTGGAAGGCCATCAGTTATC-3′
- Tmem151b—reverse: 5′-TAGTCATCCAAGCCCTCGTTCTC-3′
4.3. Generation of Hao1 Conditional Knockout Mice
4.4. Metabolome Analysis
4.5. Analysis of Skeletal Morphology
4.6. Statistical Analysis
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Matsunaga, S.; Kukita, M.; Hayashi, K.; Shinkura, R.; Koriyama, C.; Sakou, T.; Komiya, S. Pathogenesis of myelopathy in patients with ossification of the posterior longitudinal ligament. J. Neurosurg. 2002, 96, 168–172. [Google Scholar] [CrossRef] [PubMed]
- Taketomi, E.; Sakou, T.; Matsunaga, S.; Yamaguchi, M. Family study of a twin with ossification of the posterior longitudinal ligament in the cervical spine. Spine 1992, 17, S55–S56. [Google Scholar] [CrossRef] [PubMed]
- Saetia, K.; Cho, D.; Lee, S.; Kim, D.H.; Kim, S.D. Ossification of the posterior longitudinal ligament: A review. Neurosurg. Focus 2011, 30, E1. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, Y.; Nakano, M.; Yasuda, T.; Seki, S.; Hori, T.; Suzuki, K.; Makino, H.; Kimura, T. Characteristics of ossification of the spinal ligament; incidence of ossification of the ligamentum flavum in patients with cervical ossification of the posterior longitudinal ligament—Analysis of the whole spine using multidetector CT. J. Orthop. Sci. 2016, 21, 439–445. [Google Scholar] [CrossRef]
- Koyanagi, I.; Iwasaki, Y.; Hida, K.; Imamura, H.; Fujimoto, S.; Akino, M. Acute cervical cord injury associated with ossification of the posterior longitudinal ligament. Neurosurgery 2003, 53, 887–891; discussion 891–882. [Google Scholar] [CrossRef]
- Chikuda, H.; Seichi, A.; Takeshita, K.; Matsunaga, S.; Watanabe, M.; Nakagawa, Y.; Oshima, K.; Sasao, Y.; Tokuhashi, Y.; Nakahara, S.; et al. Acute cervical spinal cord injury complicated by preexisting ossification of the posterior longitudinal ligament: A multicenter study. Spine 2011, 36, 1453–1458. [Google Scholar] [CrossRef]
- Kim, Y.H.; Ha, K.Y.; Kim, S.I. Spinal Cord Injury and Related Clinical Trials. Clin. Orthop. Surg. 2017, 9, 1–9. [Google Scholar] [CrossRef]
- Komura, S.; Miyamoto, K.; Hosoe, H.; Fushimi, K.; Iwai, C.; Nishimoto, H.; Shimizu, K. Anterior cervical multilevel decompression and fusion using fibular strut as revision surgery for failed cervical laminoplasty. Arch. Orthop. Trauma Surg. 2011, 131, 1177–1185. [Google Scholar] [CrossRef]
- Okawa, A.; Nakamura, I.; Goto, S.; Moriya, H.; Nakamura, Y.; Ikegawa, S. Mutation in Npps in a mouse model of ossification of the posterior longitudinal ligament of the spine. Nat. Genet. 1998, 19, 271–273. [Google Scholar] [CrossRef]
- Lomashvili, K.A.; Garg, P.; Narisawa, S.; Millan, J.L.; O’Neill, W.C. Upregulation of alkaline phosphatase and pyrophosphate hydrolysis: Potential mechanism for uremic vascular calcification. Kidney Int. 2008, 73, 1024–1030. [Google Scholar] [CrossRef] [Green Version]
- Bollen, M.; Gijsbers, R.; Ceulemans, H.; Stalmans, W.; Stefan, C. Nucleotide pyrophosphatases/phosphodiesterases on the move. Crit. Rev. Biochem. Mol. Biol. 2000, 35, 393–432. [Google Scholar] [CrossRef]
- Terkeltaub, R. Physiologic and pathologic functions of the NPP nucleotide pyrophosphatase/phosphodiesterase family focusing on NPP1 in calcification. Purinergic Signal. 2006, 2, 371–377. [Google Scholar] [CrossRef] [Green Version]
- Uchida, K.; Yayama, T.; Sugita, D.; Nakajima, H.; Rodriguez Guerrero, A.; Watanabe, S.; Roberts, S.; Johnson, W.E.; Baba, H. Initiation and progression of ossification of the posterior longitudinal ligament of the cervical spine in the hereditary spinal hyperostotic mouse (twy/twy). Eur. Spine J. 2012, 21, 149–155. [Google Scholar] [CrossRef] [Green Version]
- Watanabe, R.; Fujita, N.; Sato, Y.; Kobayashi, T.; Morita, M.; Oike, T.; Miyamoto, K.; Kuro, O.M.; Michigami, T.; Fukumoto, S.; et al. Enpp1 is an anti-aging factor that regulates Klotho under phosphate overload conditions. Sci. Rep. 2017, 7, 7786. [Google Scholar] [CrossRef] [Green Version]
- Rutsch, F.; Ruf, N.; Vaingankar, S.; Toliat, M.R.; Suk, A.; Höhne, W.; Schauer, G.; Lehmann, M.; Roscioli, T.; Schnabel, D.; et al. Mutations in ENPP1 are associated with ‘idiopathic’ infantile arterial calcification. Nat. Genet. 2003, 34, 379–381. [Google Scholar] [CrossRef]
- Rutsch, F.; Vaingankar, S.; Johnson, K.; Goldfine, I.; Maddux, B.; Schauerte, P.; Kalhoff, H.; Sano, K.; Boisvert, W.A.; Superti-Furga, A.; et al. PC-1 nucleoside triphosphate pyrophosphohydrolase deficiency in idiopathic infantile arterial calcification. Am. J. Pathol. 2001, 158, 543–554. [Google Scholar] [CrossRef] [Green Version]
- Levy-Litan, V.; Hershkovitz, E.; Avizov, L.; Leventhal, N.; Bercovich, D.; Chalifa-Caspi, V.; Manor, E.; Buriakovsky, S.; Hadad, Y.; Goding, J.; et al. Autosomal-recessive hypophosphatemic rickets is associated with an inactivation mutation in the ENPP1 gene. Am. J. Hum. Genet. 2010, 86, 273–278. [Google Scholar] [CrossRef] [Green Version]
- Lorenz-Depiereux, B.; Schnabel, D.; Tiosano, D.; Häusler, G.; Strom, T.M. Loss-of-function ENPP1 mutations cause both generalized arterial calcification of infancy and autosomal-recessive hypophosphatemic rickets. Am. J. Hum. Genet. 2010, 86, 267–272. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, Y.; Nakano, M.; Yasuda, T.; Seki, S.; Suzuki, K.; Yahara, Y.; Makino, H.; Kitajima, I.; Kimura, T. Serum biomarkers in patients with ossification of the posterior longitudinal ligament (OPLL): Inflammation in OPLL. PLoS ONE 2017, 12, e0174881. [Google Scholar] [CrossRef] [Green Version]
- Kawaguchi, Y.; Kitajima, I.; Nakano, M.; Yasuda, T.; Seki, S.; Suzuki, K.; Yahara, Y.; Makino, H.; Ujihara, Y.; Ueno, T.; et al. Increase of the Serum FGF-23 in Ossification of the Posterior Longitudinal Ligament. Global Spine J. 2019, 9, 492–498. [Google Scholar] [CrossRef] [Green Version]
- Cai, G.D.; Zhu, Z.C.; Wang, J.Q.; Chen, G.; Wang, Z.; Yang, C.S.; Sun, G.X. Multiplex analysis of serum hormone and cytokine in patients with cervical cOPLL: Towards understanding the potential pathogenic mechanisms. Growth Factors 2017, 35, 171–178. [Google Scholar] [CrossRef]
- Ikegawa, S. Genomic study of ossification of the posterior longitudinal ligament of the spine. Proc. Jpn. Acad. Ser. B Phys. Biol. Sci. 2014, 90, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Saito, T.; Shimizu, Y.; Hori, M.; Taguchi, M.; Igarashi, T.; Fukumoto, S.; Fujitab, T. A patient with hypophosphatemic rickets and ossification of posterior longitudinal ligament caused by a novel homozygous mutation in ENPP1 gene. Bone 2011, 49, 913–916. [Google Scholar] [CrossRef]
- Delatycki, M.; Rogers, J.G. The genetics of fibrodysplasia ossificans progressiva. Clin. Orthop. Relat. Res. 1998, 346, 15–18. [Google Scholar] [CrossRef]
- Kaplan, F.S.; Groppe, J.; Pignolo, R.J.; Shore, E.M. Morphogen receptor genes and metamorphogenes: Skeleton keys to metamorphosis. Ann. N. Y. Acad. Sci. 2007, 1116, 113–133. [Google Scholar] [CrossRef]
- Hino, K.; Horigome, K.; Nishio, M.; Komura, S.; Nagata, S.; Zhao, C.; Jin, Y.; Kawakami, K.; Yamada, Y.; Ohta, A.; et al. Activin-A enhances mTOR signaling to promote aberrant chondrogenesis in fibrodysplasia ossificans progressiva. J. Clin. Invest. 2017, 127, 3339–3352. [Google Scholar] [CrossRef] [Green Version]
- Sakou, T.; Matsunaga, S.; Koga, H. Recent progress in the study of pathogenesis of ossification of the posterior longitudinal ligament. J. Orthop. Sci. 2000, 5, 310–315. [Google Scholar] [CrossRef]
- Terayama, K. Genetic studies on ossification of the posterior longitudinal ligament of the spine. Spine 1989, 14, 1184–1191. [Google Scholar] [CrossRef]
- Nakajima, M.; Takahashi, A.; Tsuji, T.; Karasugi, T.; Baba, H.; Uchida, K.; Kawabata, S.; Okawa, A.; Shindo, S.; Takeuchi, K.; et al. A genome-wide association study identifies susceptibility loci for ossification of the posterior longitudinal ligament of the spine. Nat. Genet. 2014, 46, 1012–1016. [Google Scholar] [CrossRef]
- Knight, M.N.; Hankenson, K.D. R-spondins: Novel matricellular regulators of the skeleton. Matrix Biol. 2014, 37, 157–161. [Google Scholar] [CrossRef]
- Abed, É.; Chan, T.F.; Delalandre, A.; Martel-Pelletier, J.; Pelletier, J.P.; Lajeunesse, D. R-spondins are newly recognized players in osteoarthritis that regulate Wnt signaling in osteoblasts. Arthritis Rheum. 2011, 63, 3865–3875. [Google Scholar] [CrossRef] [PubMed]
- Recalcati, S.; Tacchini, L.; Alberghini, A.; Conte, D.; Cairo, G. Oxidative stress-mediated down-regulation of rat hydroxyacid oxidase 1, a liver-specific peroxisomal enzyme. Hepatology 2003, 38, 1159–1166. [Google Scholar] [CrossRef] [PubMed]
- Frishberg, Y.; Zeharia, A.; Lyakhovetsky, R.; Bargal, R.; Belostotsky, R. Mutations in HAO1 encoding glycolate oxidase cause isolated glycolic aciduria. J. Med. Genet. 2014, 51, 526–529. [Google Scholar] [CrossRef] [PubMed]
- Dutta, C.; Avitahl-Curtis, N.; Pursell, N.; Larsson Cohen, M.; Holmes, B.; Diwanji, R.; Zhou, W.; Apponi, L.; Koser, M.; Ying, B.; et al. Inhibition of Glycolate Oxidase With Dicer-substrate siRNA Reduces Calcium Oxalate Deposition in a Mouse Model of Primary Hyperoxaluria Type 1. Mol. Ther. 2016, 24, 770–778. [Google Scholar] [CrossRef] [Green Version]
- Matsunaga, S.; Sakou, T.; Taketomi, E.; Nakanisi, K. Effects of strain distribution in the intervertebral discs on the progression of ossification of the posterior longitudinal ligaments. Spine 1996, 21, 184–189. [Google Scholar] [CrossRef]
- Ohishi, H.; Furukawa, K.; Iwasaki, K.; Ueyama, K.; Okada, A.; Motomura, S.; Harata, S.; Toh, S. Role of prostaglandin I2 in the gene expression induced by mechanical stress in spinal ligament cells derived from patients with ossification of the posterior longitudinal ligament. J. Pharmacol. Exp. Ther. 2003, 305, 818–824. [Google Scholar] [CrossRef]
- Albright, R.A.; Stabach, P.; Cao, W.; Kavanagh, D.; Mullen, I.; Braddock, A.A.; Covo, M.S.; Tehan, M.; Yang, G.; Cheng, Z.; et al. ENPP1-Fc prevents mortality and vascular calcifications in rodent model of generalized arterial calcification of infancy. Nat. Commun. 2015, 6, 10006. [Google Scholar] [CrossRef]
- Tokuhashi, Y.; Ajiro, Y.; Umezawa, N. A patient with two re-surgeries for delayed myelopathy due to progression of ossification of the posterior longitudinal ligaments after cervical laminoplasty. Spine 2009, 34, E101–E105. [Google Scholar] [CrossRef]
- Takeda, K.; Kou, I.; Hosogane, N.; Otomo, N.; Yagi, M.; Kaneko, S.; Kono, H.; Ishikawa, M.; Takahashi, Y.; Ikegami, T.; et al. Association of Susceptibility Genes for Adolescent Idiopathic Scoliosis and Intervertebral Disc Degeneration With Adult Spinal Deformity. Spine 2019, 44, 1623–1629. [Google Scholar] [CrossRef]
- Laufer, V.A.; Tiwari, H.K.; Reynolds, R.J.; Danila, M.I.; Wang, J.; Edberg, J.C.; Kimberly, R.P.; Kottyan, L.C.; Harley, J.B.; Mikuls, T.R.; et al. Genetic influences on susceptibility to rheumatoid arthritis in African-Americans. Hum. Mol. Genet. 2019, 28, 858–874. [Google Scholar] [CrossRef]
- Okada, Y.; Wu, D.; Trynka, G.; Raj, T.; Terao, C.; Ikari, K.; Kochi, Y.; Ohmura, K.; Suzuki, A.; Yoshida, S.; et al. Genetics of rheumatoid arthritis contributes to biology and drug discovery. Nature 2014, 506, 376–381. [Google Scholar] [CrossRef]
- McAllister, K.; Yarwood, A.; Bowes, J.; Orozco, G.; Viatte, S.; Diogo, D.; Hocking, L.J.; Steer, S.; Wordsworth, P.; Wilson, A.G.; et al. Identification of BACH2 and RAD51B as rheumatoid arthritis susceptibility loci in a meta-analysis of genome-wide data. Arthritis Rheum. 2013, 65, 3058–3062. [Google Scholar] [CrossRef] [Green Version]
- Barawkar, D.A.; Meru, A.; Bandyopadhyay, A.; Banerjee, A.; Deshpande, A.M.; Athare, C.; Koduru, C.; Khose, G.; Gundu, J.; Mahajan, K.; et al. Potent and Selective Inhibitors of Long Chain l-2-Hydroxy Acid Oxidase Reduced Blood Pressure in DOCA Salt-Treated Rats. ACS Med. Chem. Lett. 2011, 2, 919–923. [Google Scholar] [CrossRef] [Green Version]
- Martin-Higueras, C.; Luis-Lima, S.; Salido, E. Glycolate Oxidase Is a Safe and Efficient Target for Substrate Reduction Therapy in a Mouse Model of Primary Hyperoxaluria Type I. Mol. Ther. 2016, 24, 719–725. [Google Scholar] [CrossRef] [Green Version]
- Pillai, I.C.L.; Li, S.; Romay, M.; Lam, L.; Lu, Y.; Huang, J.; Dillard, N.; Zemanova, M.; Rubbi, L.; Wang, Y.; et al. Cardiac Fibroblasts Adopt Osteogenic Fates and Can Be Targeted to Attenuate Pathological Heart Calcification. Cell Stem. Cell 2017, 20, 218–232.e215. [Google Scholar] [CrossRef] [Green Version]
- Kuro-o, M.; Matsumura, Y.; Aizawa, H.; Kawaguchi, H.; Suga, T.; Utsugi, T.; Ohyama, Y.; Kurabayashi, M.; Kaname, T.; Kume, E.; et al. Mutation of the mouse klotho gene leads to a syndrome resembling ageing. Nature 1997, 390, 45–51. [Google Scholar] [CrossRef]
- Kurosu, H.; Yamamoto, M.; Clark, J.D.; Pastor, J.V.; Nandi, A.; Gurnani, P.; McGuinness, O.P.; Chikuda, H.; Yamaguchi, M.; Kawaguchi, H.; et al. Suppression of aging in mice by the hormone Klotho. Science 2005, 309, 1829–1833. [Google Scholar] [CrossRef] [Green Version]
- Hasegawa, T.; Sasaki, M.; Yamada, T.; Ookido, I.; Yamamoto, T.; Hongo, H.; Yamamoto, T.; Oda, K.; Yokoyama, K.; Amizuka, N. Histochemical examination of vascular medial calcification of aorta in klotho-deficient mice. J. Oral. Biosci. 2013, 55, 10–15. [Google Scholar] [CrossRef]
- Akram, M. Citric acid cycle and role of its intermediates in metabolism. Cell Biochem. Biophys. 2014, 68, 475–478. [Google Scholar] [CrossRef]
- Kiyonari, H.; Kaneko, M.; Abe, S.; Aizawa, S. Three inhibitors of FGF receptor, ERK, and GSK3 establishes germline-competent embryonic stem cells of C57BL/6N mouse strain with high efficiency and stability. Genesis 2010, 48, 317–327. [Google Scholar] [CrossRef]
- Sakai, K.; Miyazaki, J. A transgenic mouse line that retains cre recombinase activity in mature oocytes irrespective of the cre transgene transmission. Biochem. Biophys. Res. Commun. 1997, 18, 318–324. [Google Scholar] [CrossRef]
- Kobayashi, H.; Nakamura, S.; Sato, Y.; Kobayashi, T.; Miyamoto, K.; Oya, A.; Matsumoto, M.; Nakamura, M.; Kanaji, A.; Miyamoto, T. ALDH2 mutation promotes skeletal muscle atrophy in mice via accumulation of oxidative stress. Bone 2021, 142, 115739. [Google Scholar] [CrossRef]
- Soma, T.; Iwasaki, R.; Sato, Y.; Kobayashi, T.; Nakamura, S.; Kaneko, Y.; Ito, E.; Okada, H.; Watanabe, H.; Miyamoto, K.; et al. Tooth extraction in mice administered zoledronate increases inflammatory cytokine levels and promotes osteonecrosis of the jaw. J. Bone Miner. Metab. 2021, 39, 372–384. [Google Scholar] [CrossRef]
- Umezu, T.; Nakamura, S.; Sato, Y.; Kobayashi, T.; Ito, E.; Abe, T.; Kaneko, M.; Nomura, M.; Yoshimura, A.; Oya, A.; et al. Smad2 and Smad3 expressed in skeletal muscle promote immobilization-induced bone atrophy in mice. Biochem. Biophys. Res. Commun. 2021, 582, 111–117. [Google Scholar] [CrossRef]
- Sugimoto, M.; Wong, D.T.; Hirayama, A.; Soga, T.; Tomita, M. Capillary electrophoresis mass spectrometry-based saliva metabolomics identified oral, breast and pancreatic cancer-specific profiles. Metabolomics 2010, 6, 78–95. [Google Scholar] [CrossRef] [Green Version]
- Soga, T.; Igarashi, K.; Ito, C.; Mizobuchi, K.; Zimmermann, H.P.; Tomita, M. Metabolomic profiling of anionic metabolites by capillary electrophoresis mass spectrometry. Anal. Chem. 2009, 81, 6165–6174. [Google Scholar] [CrossRef] [Green Version]
- Hirayama, A.; Sugimoto, M.; Suzuki, A.; Hatakeyama, Y.; Enomoto, A.; Harada, S.; Soga, T.; Tomita, M.; Takebayashi, T. Effects of processing and storage conditions on charged metabolomic profiles in blood. Electrophoresis 2015, 36, 2148–2155. [Google Scholar] [CrossRef]
- Bouxsein, M.L.; Boyd, S.K.; Christiansen, B.A.; Guldberg, R.E.; Jepsen, K.J.; Müller, R. Guidelines for Assessment of Bone Microstructure in Rodents Using Micro–Computed Tomography. J. Bone Miner. Res. 2010, 7, 1468–1486. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kimura, A.; Hirayama, A.; Matsumoto, T.; Sato, Y.; Kobayashi, T.; Ikeda, S.; Maruyama, M.; Kaneko, M.; Shigeta, M.; Ito, E.; et al. Hao1 Is Not a Pathogenic Factor for Ectopic Ossifications but Functions to Regulate the TCA Cycle In Vivo. Metabolites 2022, 12, 82. https://doi.org/10.3390/metabo12010082
Kimura A, Hirayama A, Matsumoto T, Sato Y, Kobayashi T, Ikeda S, Maruyama M, Kaneko M, Shigeta M, Ito E, et al. Hao1 Is Not a Pathogenic Factor for Ectopic Ossifications but Functions to Regulate the TCA Cycle In Vivo. Metabolites. 2022; 12(1):82. https://doi.org/10.3390/metabo12010082
Chicago/Turabian StyleKimura, Atsushi, Akiyoshi Hirayama, Tatsuaki Matsumoto, Yuiko Sato, Tami Kobayashi, Satsuki Ikeda, Midori Maruyama, Mari Kaneko, Mayo Shigeta, Eri Ito, and et al. 2022. "Hao1 Is Not a Pathogenic Factor for Ectopic Ossifications but Functions to Regulate the TCA Cycle In Vivo" Metabolites 12, no. 1: 82. https://doi.org/10.3390/metabo12010082