

Lipid Metabolism Reprogramming and Trastuzumab Resistance in Breast Cancer Cell Lines Overexpressing the ERBB2 Membrane Receptor
 , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Cell Culture Reagents
2.2. Flow Cytometry [FCM] Analysis and Antibodies
2.3. Cell Imaging
2.4. Sample Preparation, Mass Spectrometer Setup and Protein Identification
2.5. Electron Microscopy
2.6. Statistical Analyses
3. Results
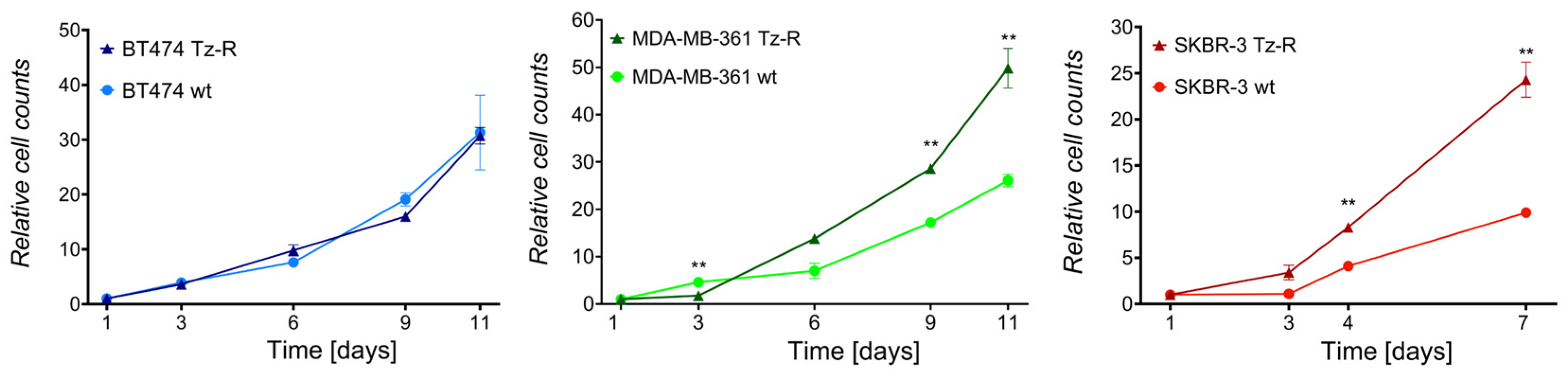
3.1. Generation and Characterization of ERBB2+ Tz-R Cell Lines
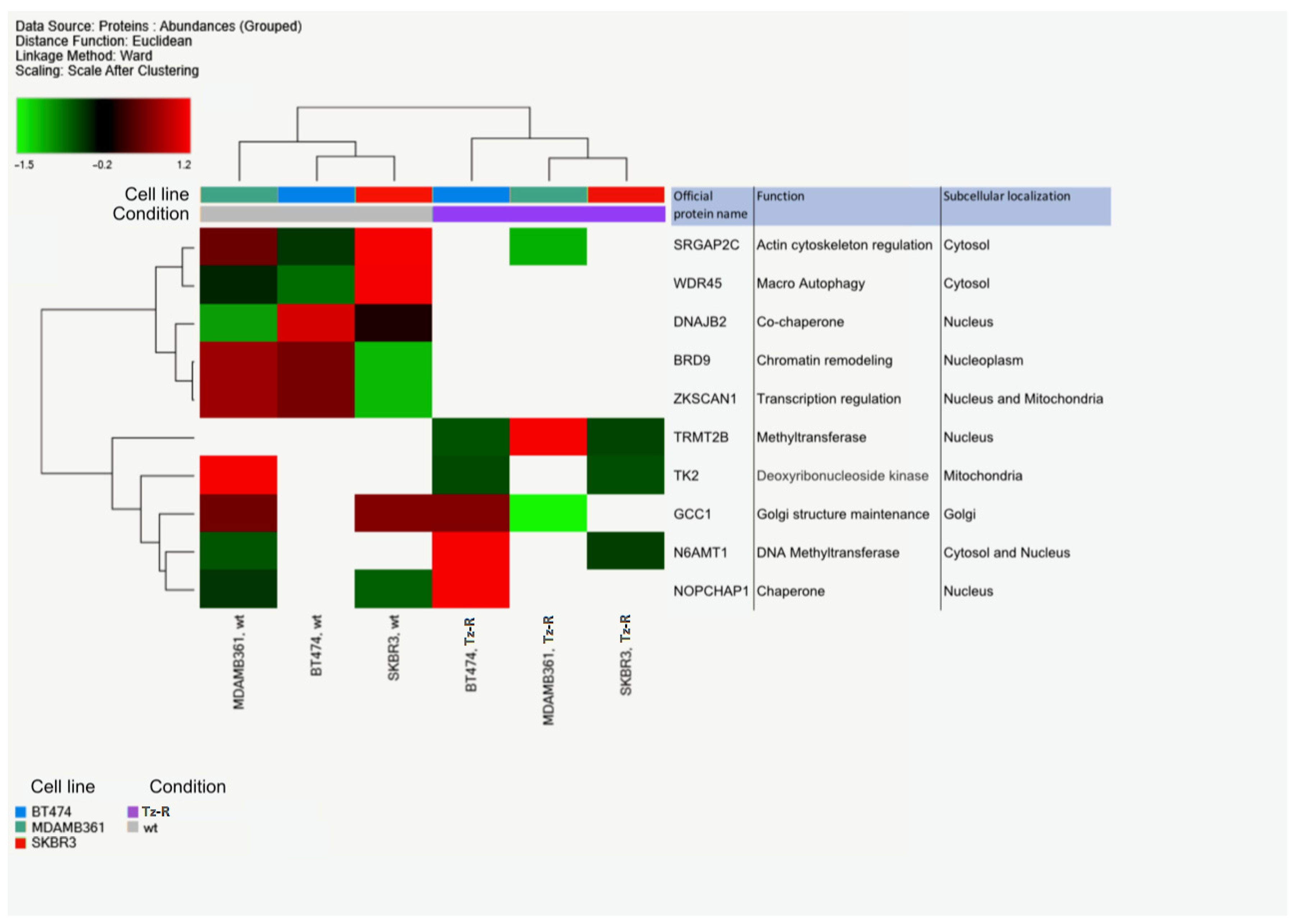
3.2. Proteomic Analysis of ERBB2+ and Tz-R Cell Lines Reveals Deregulation of Lipid Metabolism, Organophosphate Biosynthetic Process, and Macromolecule Methylation
3.3. Electron Microscopy Revealed Changes in Lipid Droplets Content in Tz-R Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human Breast Cancer: Correlation of Relapse and Survival with Amplification of the HER-2/Neu Oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [PubMed]
- Slamon, D.J.; Godolphin, W.; Jones, L.A.; Holt, J.A.; Wong, S.G.; Keith, D.E.; Levin, W.J.; Stuart, S.G.; Udove, J.; Ullrich, A. Studies of the HER-2/Neu Proto-Oncogene in Human Breast and Ovarian Cancer. Science 1989, 244, 707–712. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z. ErbB Receptors and Cancer. Methods Mol. Biol. 2017, 1652, 3–35. [Google Scholar] [CrossRef]
- Braicu, C.; Buse, M.; Busuioc, C.; Drula, R.; Gulei, D.; Raduly, L.; Rusu, A.; Irimie, A.; Atanasov, A.G.; Slaby, O.; et al. A Comprehensive Review on MAPK: A Promising Therapeutic Target in Cancer. Cancers 2019, 11, 1618. [Google Scholar] [CrossRef] [PubMed]
- Revathidevi, S.; Munirajan, A.K. Akt in Cancer: Mediator and More. Semin. Cancer Biol. 2019, 59, 80–91. [Google Scholar] [CrossRef]
- Greenberg, P.A.; Hortobagyi, G.N.; Smith, T.L.; Ziegler, L.D.; Frye, D.K.; Buzdar, A.U. Long-Term Follow-up of Patients with Complete Remission Following Combination Chemotherapy for Metastatic Breast Cancer. J. Clin. Oncol. 1996, 14, 2197–2205. [Google Scholar] [CrossRef]
- Vogel, C.L.; Cobleigh, M.A.; Tripathy, D.; Gutheil, J.C.; Harris, L.N.; Fehrenbacher, L.; Slamon, D.J.; Murphy, M.; Novotny, W.F.; Burchmore, M.; et al. Efficacy and Safety of Trastuzumab as a Single Agent in First-Line Treatment of HER2-Overexpressing Metastatic Breast Cancer. J. Clin. Oncol. 2002, 20, 719–726. [Google Scholar] [CrossRef]
- Cortés, J.; Kim, S.-B.; Chung, W.-P.; Im, S.-A.; Park, Y.H.; Hegg, R.; Kim, M.H.; Tseng, L.-M.; Petry, V.; Chung, C.-F.; et al. Trastuzumab Deruxtecan versus Trastuzumab Emtansine for Breast Cancer. N. Engl. J. Med. 2022, 386, 1143–1154. [Google Scholar] [CrossRef]
- Lane, H.A.; Motoyama, A.B.; Beuvink, I.; Hynes, N.E. Modulation of P27/Cdk2 Complex Formation through 4D5-Mediated Inhibition of HER2 Receptor Signaling. Ann. Oncol. 2001, 12 (Suppl. 1), S21–S22. [Google Scholar] [CrossRef]
- Denny, E.C.; Kane, S.E. T-Darpp Promotes Enhanced EGFR Activation and New Drug Synergies in Her2-Positive Breast Cancer Cells. PLoS ONE 2015, 10, e0132267. [Google Scholar] [CrossRef]
- Maadi, H.; Soheilifar, M.H.; Choi, W.-S.; Moshtaghian, A.; Wang, Z. Trastuzumab Mechanism of Action; 20 Years of Research to Unravel a Dilemma. Cancers 2021, 13, 3540. [Google Scholar] [CrossRef] [PubMed]
- Moasser, M.M. Inactivating Amplified HER2: Challenges, Dilemmas, and Future Directions. Cancer Res. 2022, 82, 2811–2820. [Google Scholar] [CrossRef] [PubMed]
- D’Alesio, C.; Bellese, G.; Gagliani, M.C.; Lechiara, A.; Dameri, M.; Grasselli, E.; Lanfrancone, L.; Cortese, K.; Castagnola, P. The Chromodomain Helicase CHD4 Regulates ERBB2 Signaling Pathway and Autophagy in ERBB2+ Breast Cancer Cells. Biol. Open 2019, 8, bio.038323. [Google Scholar] [CrossRef]
- Nagata, Y.; Lan, K.-H.; Zhou, X.; Tan, M.; Esteva, F.J.; Sahin, A.A.; Klos, K.S.; Li, P.; Monia, B.P.; Nguyen, N.T.; et al. PTEN Activation Contributes to Tumor Inhibition by Trastuzumab, and Loss of PTEN Predicts Trastuzumab Resistance in Patients. Cancer Cell 2004, 6, 117–127. [Google Scholar] [CrossRef]
- Kreutzfeldt, J.; Rozeboom, B.; Dey, N.; De, P. The Trastuzumab Era: Current and Upcoming Targeted HER2+ Breast Cancer Therapies. Am. J. Cancer Res. 2020, 10, 1045–1067. [Google Scholar] [PubMed]
- Vivekanandhan, S.; Knutson, K.L. Resistance to Trastuzumab. Cancers 2022, 14, 5115. [Google Scholar] [CrossRef]
- Dai, X.; Cheng, H.; Bai, Z.; Li, J. Breast Cancer Cell Line Classification and Its Relevance with Breast Tumor Subtyping. J. Cancer 2017, 8, 3131–3141. [Google Scholar] [CrossRef] [PubMed]
- Narayan, M.; Wilken, J.A.; Harris, L.N.; Baron, A.T.; Kimbler, K.D.; Maihle, N.J. Trastuzumab-Induced HER Reprogramming in “Resistant” Breast Carcinoma Cells. Cancer Res. 2009, 69, 2191–2194. [Google Scholar] [CrossRef]
- Cardinali, B.; Lunardi, G.; Millo, E.; Armirotti, A.; Damonte, G.; Profumo, A.; Gori, S.; Iacono, G.; Levaggi, A.; Del Mastro, L. Trastuzumab Quantification in Serum: A New, Rapid, Robust ELISA Assay Based on a Mimetic Peptide That Specifically Recognizes Trastuzumab. Anal. Bioanal. Chem. 2014, 406, 4557–4561. [Google Scholar] [CrossRef]
- Perez-Riverol, Y.; Bai, J.; Bandla, C.; García-Seisdedos, D.; Hewapathirana, S.; Kamatchinathan, S.; Kundu, D.J.; Prakash, A.; Frericks-Zipper, A.; Eisenacher, M.; et al. The PRIDE Database Resources in 2022: A Hub for Mass Spectrometry-Based Proteomics Evidences. Nucleic Acids Res. 2022, 50, D543–D552. [Google Scholar] [CrossRef]
- D’Alesio, C.; Bellese, G.; Gagliani, M.C.; Aiello, C.; Grasselli, E.; Marcocci, G.; Bisio, A.; Tavella, S.; Daniele, T.; Cortese, K.; et al. Cooperative Antitumor Activities of Carnosic Acid and Trastuzumab in ERBB2+ Breast Cancer Cells. J. Exp. Clin. Cancer Res. 2017, 36, 154. [Google Scholar] [CrossRef] [PubMed]
- Zuo, Q.; Liu, J.; Zhang, J.; Wu, M.; Guo, L.; Liao, W. Development of Trastuzumab-Resistant Human Gastric Carcinoma Cell Lines and Mechanisms of Drug Resistance. Sci. Rep. 2015, 5, 11634. [Google Scholar] [CrossRef] [PubMed]
- Ignatov, T.; Gorbunow, F.; Eggemann, H.; Ortmann, O.; Ignatov, A. Loss of HER2 after HER2-Targeted Treatment. Breast Cancer Res. Treat. 2019, 175, 401–408. [Google Scholar] [CrossRef] [PubMed]
- Swain, S.M.; Shastry, M.; Hamilton, E. Targeting HER2-Positive Breast Cancer: Advances and Future Directions. Nat. Rev. Drug Discov. 2023, 22, 101–126. [Google Scholar] [CrossRef] [PubMed]
- Menyhárt, O.; Santarpia, L.; Győrffy, B. A Comprehensive Outline of Trastuzumab Resistance Biomarkers in HER2 Overexpressing Breast Cancer. Curr. Cancer Drug Targets 2015, 15, 665–683. [Google Scholar] [CrossRef]
- Marko, T.A.; Shamsan, G.A.; Edwards, E.N.; Hazelton, P.E.; Rathe, S.K.; Cornax, I.; Overn, P.R.; Varshney, J.; Diessner, B.J.; Moriarity, B.S.; et al. Slit-Robo GTPase-Activating Protein 2 as a Metastasis Suppressor in Osteosarcoma. Sci. Rep. 2016, 6, 39059. [Google Scholar] [CrossRef]
- Fei, H.; Chen, X. A Novel Autophagy-Related Prognostic Risk Model and a Nomogram for Survival Prediction of Oral Cancer Patients. Biomed Res. Int. 2022, 2022, 2067540. [Google Scholar] [CrossRef]
- Feng, W.W.; Kurokawa, M. Lipid metabolic reprogramming as an emerging mechanism of resistance to kinase inhibitors in breast cancer. Cancer Drug Resist. 2019, 3, 1–17. [Google Scholar] [CrossRef]
- Menendez, J.A.; Vellon, L.; Mehmi, I.; Oza, B.P.; Ropero, S.; Colomer, R.; Lupu, R. Inhibition of Fatty Acid Synthase (FAS) Suppresses HER2/Neu ( Erb B-2) Oncogene Overexpression in Cancer Cells. Proc. Natl. Acad. Sci. USA 2004, 101, 10715–10720. [Google Scholar] [CrossRef]
- Feng, W.W.; Wilkins, O.; Bang, S.; Ung, M.; Li, J.; An, J.; Del Genio, C.; Canfield, K.; DiRenzo, J.; Wells, W.; et al. CD36-Mediated Metabolic Rewiring of Breast Cancer Cells Promotes Resistance to HER2-Targeted Therapies. Cell Rep. 2019, 29, 3405–3420.e5. [Google Scholar] [CrossRef]
- Yamashita, S.; Hattori, N.; Fujii, S.; Yamaguchi, T.; Takahashi, M.; Hozumi, Y.; Kogawa, T.; El-Omar, O.; Liu, Y.-Y.; Arai, N.; et al. Multi-Omics Analyses Identify HSD17B4 Methylation-Silencing as a Predictive and Response Marker of HER2-Positive Breast Cancer to HER2-Directed Therapy. Sci. Rep. 2020, 10, 15530. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Mukai, H.; Yamashita, S.; Fujii, S.; Ushijima, T. Comprehensive DNA Methylation and Extensive Mutation Analyses of HER2-Positive Breast Cancer. Oncology 2015, 88, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Palomeras, S.; Diaz-Lagares, Á.; Viñas, G.; Setien, F.; Ferreira, H.J.; Oliveras, G.; Crujeiras, A.B.; Hernández, A.; Lum, D.H.; Welm, A.L.; et al. Epigenetic Silencing of TGFBI Confers Resistance to Trastuzumab in Human Breast Cancer. Breast Cancer Res. 2019, 21, 79. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| BT474 wt vs. Tz-R | MDA-MB-361 wt vs. Tz-R | SKBR-3 wt vs. Tz-R | ||||||
|---|---|---|---|---|---|---|---|---|
| KEGG Pathway | % of DEPs | p-Value | KEGG Pathway | % of DEPs | p-Value | KEGG Pathway | % of DEPs | p-Value |
| Metabolic pathways | 18.6 | 5.00 × 10−5 | Chronic myeloid leukemia | 2.9 | 1.40 × 10−2 | Metabolic pathways | 28 | 7.00 × 10−3 |
| Biosynthesis of cofactors | 4.3 | 5.00 × 10−3 | TGF-beta signaling pathway | 2.9 | 2.50 × 10−2 | Thyroid hormone signaling pathway | 6 | 1.10 × 10−2 |
| Metabolic pathways | 17 | 4.30 × 10−2 | Glycerophospholipid metabolism | 5 | 2.40 × 10−2 | |||
| Proteoglycans in cancer | 5 | 4.90 × 10−2 | ||||||
| PATTERN | Identifier/Term | Category | Descrlptlon | Count | % | Log10(P) | Log10(q) |
|---|---|---|---|---|---|---|---|
| ■■■ | R-HSA-556833 | Reactome Gene Sets | metabolism of lipids | 36 | 8.98 | −10.5 | −6.15 |
| ■■■ | GO:0090407 | GO Biological Processes | organophosphate biosynthetic process | 28 | 6.98 | −8.89 | −4.84 |
| ■■■ | GO:0043414 | GO Biological Processes | macromolecule methylation | 11 | 7.97 | −7.18 | −3.13 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortese, K.; Ponassi, M.; Profumo, A.; Coronel Vargas, G.; Iervasi, E.; Gagliani, M.C.; Bellese, G.; Tavella, S.; Castagnola, P. Lipid Metabolism Reprogramming and Trastuzumab Resistance in Breast Cancer Cell Lines Overexpressing the ERBB2 Membrane Receptor. Membranes 2023, 13, 540. https://doi.org/10.3390/membranes13060540
Cortese K, Ponassi M, Profumo A, Coronel Vargas G, Iervasi E, Gagliani MC, Bellese G, Tavella S, Castagnola P. Lipid Metabolism Reprogramming and Trastuzumab Resistance in Breast Cancer Cell Lines Overexpressing the ERBB2 Membrane Receptor. Membranes. 2023; 13(6):540. https://doi.org/10.3390/membranes13060540
Chicago/Turabian StyleCortese, Katia, Marco Ponassi, Aldo Profumo, Gabriela Coronel Vargas, Erika Iervasi, Maria Cristina Gagliani, Grazia Bellese, Sara Tavella, and Patrizio Castagnola. 2023. "Lipid Metabolism Reprogramming and Trastuzumab Resistance in Breast Cancer Cell Lines Overexpressing the ERBB2 Membrane Receptor" Membranes 13, no. 6: 540. https://doi.org/10.3390/membranes13060540