
Kocuria flava, a Bacterial Endophyte of the Marine Macroalga Bryopsis plumosa, Emits 8-Nonenoic Acid Which Inhibits the Aquaculture Pathogen Saprolegnia parasitica

Abstract
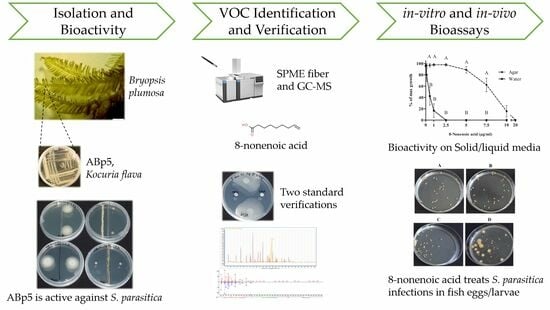
:
1. Introduction
2. Results
2.1. Bioactivity Assays
2.2. VOC Identification
2.2.1. Testing Standards for Bioactivity against S. parasitica
2.2.2. 8-Nonenoic Acid Identification by GC Comparison
2.3. In Vitro 8-Nonenoic Acid Concentration Bioassay
2.4. In Vivo Influence of 8-Nonenoic Acid on Tilapia Eggs Challenged with S. parasitica
3. Discussion
4. Materials and Methods
4.1. Endophyte Isolation and Maintenance
4.2. Bioactivity Assays
4.3. VOC Identification
4.3.1. Testing Standards for Bioactivity against S. parasitica
4.3.2. 8-Nonenoic Acid Identification by GC Comparison
4.4. In Vitro 8-Nonenoic Acid Concentration Bioassay
4.5. In Vivo Influence of 8-Nonenoic Acid on Tilapia Eggs Challenged with S. parasitica
4.6. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Rodriguez, R.; Redman, R. More than 400 million years of evolution and some plants still can’t make it on their own: Plant stress tolerance via fungal symbiosis. J. Exp. Bot. 2008, 59, 1109–1114. [Google Scholar] [CrossRef]
- Liarzi, O.; Bar, E.; Lewinsohn, E.; Ezra, D. Use of the endophytic fungus Daldinia cf. concentrica and Its volatiles as bio-control agents. PLoS ONE 2016, 11, e0168242. [Google Scholar] [CrossRef] [PubMed]
- Bacon, C.W.; White, J.F. Functions, mechanisms and regulation of endophytic and epiphytic microbial communities of plants. Symbiosis 2016, 68, 87–98. [Google Scholar] [CrossRef]
- Gouda, S.; Das, G.; Sen, S.K.; Shin, H.S.; Patra, J.K. Endophytes: A treasure house of bioactive compounds of medicinal importance. Front. Microbiol. 2016, 7, 1538. [Google Scholar] [CrossRef]
- Masand, M.; Jose, P.A.; Menghani, E.; Jebakumar, S.R.D. Continuing hunt for endophytic actinomycetes as a source of novel biologically active metabolites. World J. Microbiol. Biotechnol. 2015, 31, 1863–1875. [Google Scholar] [CrossRef]
- Newman, D.J.; Cragg, G.M. Plant endophytes and epiphytes: Burgeoning sources of known and “unknown” cytotoxic and antibiotic agents? Planta Med. 2020, 86, 891–905. [Google Scholar] [CrossRef] [PubMed]
- Flewelling, A.J.; Ellsworth, K.T.; Sanford, J.; Forward, E.; Johnson, J.A.; Gray, C.A. Macroalgal endophytes from the Atlantic coast of Canada: A potential source of antibiotic natural products? Microorganisms 2013, 1, 175–187. [Google Scholar] [CrossRef]
- Girão, M.; Ribeiro, I.; Ribeiro, T.; Azevedo, I.C.; Pereira, F.; Urbatzka, R.; Leão, P.N.; Carvalho, M.F. Actinobacteria isolated fromI laminaria ochroleuca: A source of new bioactive compounds. Front. Microbiol. 2019, 10, 683. [Google Scholar] [CrossRef]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, physiology, and natural products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef]
- Anandan, R.; Dharumadurai, D.; Manogaran, G.P. An Introduction to Actinobacteria; IntechOpen: London, UK, 2016; ISBN 978-953-51-2248-7. [Google Scholar]
- Manivasagan, P.; Venkatesan, J.; Sivakumar, K.; Kim, S.K. Pharmaceutically active secondary metabolites of marine Actinobacteria. Microbiol. Res. 2014, 169, 262–278. [Google Scholar] [CrossRef]
- Zhou, G.; Luo, X.; Tang, Y.; Zhang, L.; Yang, Q.; Qiu, Y.; Fang, C.X. Kocuria flava sp. nov. and Kocuria turfanensis sp. nov., airborne Actinobacteria isolated from Xinjiang, China. Int. J. Syst. Evol. Microbiol. 2008, 58, 1304–1307. [Google Scholar] [CrossRef] [PubMed]
- Rakshit, A.; Sukul, S. Studies on two indole acetic acid (IAA) producing endophytic bacteria from Thelypteris interrupta (Wild.) K. Iwats and their effect on seed germination. Res. J. Agric. Sci. 2021, 12, 2037–2043. [Google Scholar]
- Pachaiyappan, A.; Sadhasivam, G.; Kumar, M.; Muthuvel, A. Biomedical potential of astaxanthin from novel endophytic pigment producing bacteria Pontibacter korlensis AG6. Waste Biomass Valori. 2021, 12, 2119–2129. [Google Scholar] [CrossRef]
- Singh, N.K.; Rai, U.N.; Verma, D.K.; Rathore, G. Kocuria flava induced growth and chromium accumulation in Cicer arietinum L. Int. J. Phytoremediation 2013, 16, 14–28. [Google Scholar] [CrossRef] [PubMed]
- Kaur, H.; Dolma, K.; Kaur, N.; Malhotra, A.; Kumar, N.; Dixit, P.; Sharma, D.; Mayilraj, S.; Choudhury, A.R. Marine microbe as nano-factories for copper biomineralization. Biotechnol. Bioprocess Eng. 2015, 20, 51–57. [Google Scholar] [CrossRef]
- Sun, W.; Liu, C.; Zhang, F.; Zhao, M.; Li, Z. Comparative genomics provides insights into the marine adaptation in sponge-derived Kocuria flava S43. Front. Microbiol. 2018, 9, 1257. [Google Scholar] [CrossRef]
- Deutsch, Y.; Gur, L.; Berman Frank, I.; Ezra, D. Endophytes from algae, a potential source for new biologically active metabolites for disease management in aquaculture. Front. Mar. Sci. 2021, 8, 636636. [Google Scholar] [CrossRef]
- Ba-akdah, M.A.; Satheesh, S. Characterization and antifouling activity analysis of extracellular polymeric substances produced by an epibiotic bacterial strain Kocuria flava associated with the green macroalga Ulva lactuca. Acta Oceanol. Sin. 2021, 40, 107–115. [Google Scholar] [CrossRef]
- Najjar, A.; Hassan, E.A.; Zabermawi, N.; Saber, S.H.; Bajrai, L.H.; Almuhayawi, M.S.; Abujamel, T.S.; Almasaudi, S.B.; Azhar, L.E.; Moulay, M.; et al. Optimizing the catalytic activities of methanol and thermo tolerant Kocuria flava lipases for biodiesel production from cooking oil wastes. Sci. Rep. 2021, 11, 13659. [Google Scholar] [CrossRef]
- Hollants, J.; Decleyre, H.; Leliaert, F.; De Clerck, O.; Willems, A. Life without a cell membrane: Challenging the specificity of bacterial endophytes within Bryopsis (Bryopsidales, chlorophyta). BMC Microbiol. 2011, 11, 255. [Google Scholar] [CrossRef]
- Van West, P. Saprolegnia parasitica, an oomycete pathogen with a fishy appetite: New challenges for an old problem. Mycologist 2006, 20, 99–104. [Google Scholar] [CrossRef]
- Earle, G.; Hintz, W. New approaches for controlling Saprolegnia parasitica, the causal agent of a devastating fish disease. Trop. Life Sci. Res. 2014, 25, 101–109. [Google Scholar] [PubMed]
- Schar, D.; Klein, E.Y.; Laxminarayan, R.; Gilbert, M.; Boeckel, T.P. Van Global trends in antimicrobial use in aquaculture. Sci. Rep. 2020, 10, 21878. [Google Scholar] [CrossRef] [PubMed]
- Luthman, O.; Jonell, M.; Troell, M. Governing the salmon farming industry: Comparison between national regulations and the ASC salmon standard. Mar. Policy 2019, 106, 103534. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nation. The State of World Fisheries and Aquaculture 2020; Sustainability in Action; FAO: Rome, Italy, 2020; pp. 1–244. [Google Scholar]
- Siri-udom, S.; Suwannarach, N.; Lumyong, S. Existence of Muscodor vitigenus, M. equiseti and M. heveae Sp. Nov. in leaves of the rubber tree (Hevea brasiliensis Müll. Arg.), and their biocontrol potential. Ann. Microbiol. 2016, 66, 437–448. [Google Scholar] [CrossRef]
- Burranboina, K.K.; Kumar, K.M.; Manjunatha Reddy, G.B.; Yogisharadhya, R.; Prashantha, C.N.; Dhulappa, A. GC-MS analysis, molecular docking and pharmacokinetic studies of various bioactive compounds from methanolic leaf extracts of Leucas aspera (L) against anti-capripox viral activity. Chem. Data Collect. 2022, 39, 100873. [Google Scholar] [CrossRef]
- Sisido, K.; Kawanisi, M.; Kondö, K.; Morimoto, T.; Saitö, A.; Hukue, N. Syntheses of 9-keto- and 10-hydroxy-trans-2-decenoic acids and related compounds. J. Org. Chem. 1962, 27, 4073–4076. [Google Scholar] [CrossRef]
- Chan, T.H.; Stossel, D. Chemistry of l,3,5-tris(trimethylsiloxy)-l-methoxyhexa-l,3,5-triene, a β-tricarbonyl trianion equivalent. J. Org. Chem. 1986, 51, 2423–2428. [Google Scholar] [CrossRef]
- Kula, J.; Smigielski, K.; Quang, T.B.; Grzelak, I.; Sikora, M. Preparation of ω-hydroxynonanoic acid and its ester derivatives. JAOCS J. Am. Oil Chem. Soc. 1999, 76, 811–817. [Google Scholar] [CrossRef]
- Unanyan, M.P.; Vinogradova, L.P.; Filippov, V.V.; Zav’yalov, S.I. New method of synthesis of biologically important derivatives of pelargonic acid. Bull. Acad. Sci. USSR Div. Chem. Sci. 1967, 16, 1722–1724. [Google Scholar] [CrossRef]
- De Facci, M.; Wang, H.L.; Yuvaraj, J.K.; Dublon, I.A.N.; Svensson, G.P.; Chapman, T.W.; Anderbrant, O. Chemical composition of anal droplets of the eusocial gall-inducing thrips Kladothrips intermedius. Chemoecology 2014, 24, 85–94. [Google Scholar] [CrossRef]
- Khazanehdari, C.; Buglass, A.J.; Waterhouse, J.S. Anal Gland secretion of european mole: Volatile constituents and significance in territorial maintenance. J. Chem. Ecol. 1996, 22, 383–392. [Google Scholar] [CrossRef]
- Yakubu, O.E.; Abah, M. Comparative determination of antioxidant activities and phytochemicals from fractions of ethanol extract of Senna occidentalis using GC-MS. J. Emerg. Technol. Innov. Res. 2021, 8, 195–211. [Google Scholar]
- Ravichandran, Y.D.; Yetayih, M.M. The GC–MS analysis of the diethylether and ethylacetate fraction of the peel of Solanum incanum and the study of their antibacterial activity. Adv. Tradit. Med. 2022, 22, 809–821. [Google Scholar] [CrossRef]
- Liarzi, O.; Benichis, M.; Gamliel, A.; Ezra, D. trans-2-octenal, a single compound of a fungal origin, controls Sclerotium rolfsii, both in vitro and in soil. Pest Manag. Sci. 2020, 76, 2068–2071. [Google Scholar] [CrossRef]
- Willoughby, L.G.; Roberts, R.J. Towards strategic use of fungicides against Saprolegnia parasitica in salmonid fish hatcheries. J. Fish Dis. 1992, 15, 1–13. [Google Scholar] [CrossRef]
- Tedesco, P.; Fioravanti, M.L.; Galuppi, R. In vitro activity of chemicals and commercial products against Saprolegnia parasitica and Saprolegnia delica strains. J. Fish Dis. 2019, 42, 237–248. [Google Scholar]
- Oono, H.; Hatai, K.; Miura, M.; Tuchida, N.; Kiryu, T. The use of bronopol to control fungal infection in rainbow trout eggs. Biocontrol Sci. 2007, 12, 55–57. [Google Scholar] [CrossRef] [PubMed]
- Eissa, A.E.; Abdelsalam, M.; Tharwat, N.; Zaki, M. Detection of Saprolegnia parasitica in eggs of angelfish Pterophyllum scalare (cuvier–valenciennes) with a history of decreased hatchability. Int. J. Vet. Sci. 2013, 1, 7–14. [Google Scholar] [CrossRef]
- Cosentino, S.; Voldby Larsen, M.; Møller Aarestrup, F.; Lund, O. Pathogenfinder–distinguishing friend from foe using bacterial whole genome sequence data. PLoS ONE 2013, 8, e77302. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Retention Time (Min) | Compound * | Family | Molecular Formula | Mass | Score | Height | Area |
|---|---|---|---|---|---|---|---|
| 6.882 | Oxime-, methoxy-phenyl-_ | Phenol | C8H9NO2 | 151.1 | 79.12 | 1,321,307 | 53,113,398 |
| 9.488 | 2-Propyl-1-pentanol | Alcohol | C8H18O | 130.1 | 85.71 | 1,055,720 | 42,679,237 |
| 10.773 | 2-Nonanone | Ketone | C9H18O | 142.1 | 82.44 | 2,906,369 | 85,262,293 |
| 12.137 | 2-Decanone | Ketone | C10H20O | 156.2 | 75.06 | 1,167,692 | 33,491,708 |
| 12.383 | 8-Nonenoic acid | Fatty acid | C9H16O2 | 156.1 | 82.06 | 755,023 | 28,399,821 |
| 14.513 | 2-Undecanone | Ketone | C11H22O | 170.2 | 91.03 | 3,424,025 | 79,041,922 |
| 14.623 | 2-Undecanol | Alcohol | C11H24O | 172.2 | 83.82 | 1,391,479 | 44,357,455 |
| 15.716 | 2-Dodecanone | Ketone | C12H24O | 184.2 | 73.23 | 1,460,936 | 39,991,523 |
| 17.23 | 2-Tridecanone | Ketone | C13H26O | 198.2 | 88.75 | 1,502,874 | 36,978,277 |
| 19.379 | 2,2,4-Trimethyl-1,3-pentanediol diisobutyrate | Ester | C16H30O4 | 286.2 | 83.7 | 2,495,851 | 73,829,637 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Deutsch, Y.; Samara, M.; Nasser, A.; Berman-Frank, I.; Ezra, D. Kocuria flava, a Bacterial Endophyte of the Marine Macroalga Bryopsis plumosa, Emits 8-Nonenoic Acid Which Inhibits the Aquaculture Pathogen Saprolegnia parasitica. Mar. Drugs 2023, 21, 476. https://doi.org/10.3390/md21090476
Deutsch Y, Samara M, Nasser A, Berman-Frank I, Ezra D. Kocuria flava, a Bacterial Endophyte of the Marine Macroalga Bryopsis plumosa, Emits 8-Nonenoic Acid Which Inhibits the Aquaculture Pathogen Saprolegnia parasitica. Marine Drugs. 2023; 21(9):476. https://doi.org/10.3390/md21090476
Chicago/Turabian StyleDeutsch, Ynon, Mohamed Samara, Ahmed Nasser, Ilana Berman-Frank, and David Ezra. 2023. "Kocuria flava, a Bacterial Endophyte of the Marine Macroalga Bryopsis plumosa, Emits 8-Nonenoic Acid Which Inhibits the Aquaculture Pathogen Saprolegnia parasitica" Marine Drugs 21, no. 9: 476. https://doi.org/10.3390/md21090476