
Branched Chondroitin Sulfate Oligosaccharides Derived from the Sea Cucumber Acaudina molpadioides Stimulate Neurite Outgrowth
Abstract
:1. Introduction
2. Results and Discussion
2.1. Preparation of Homogenous Fractions and Oligosaccharides from the Depolymerized FCSAm
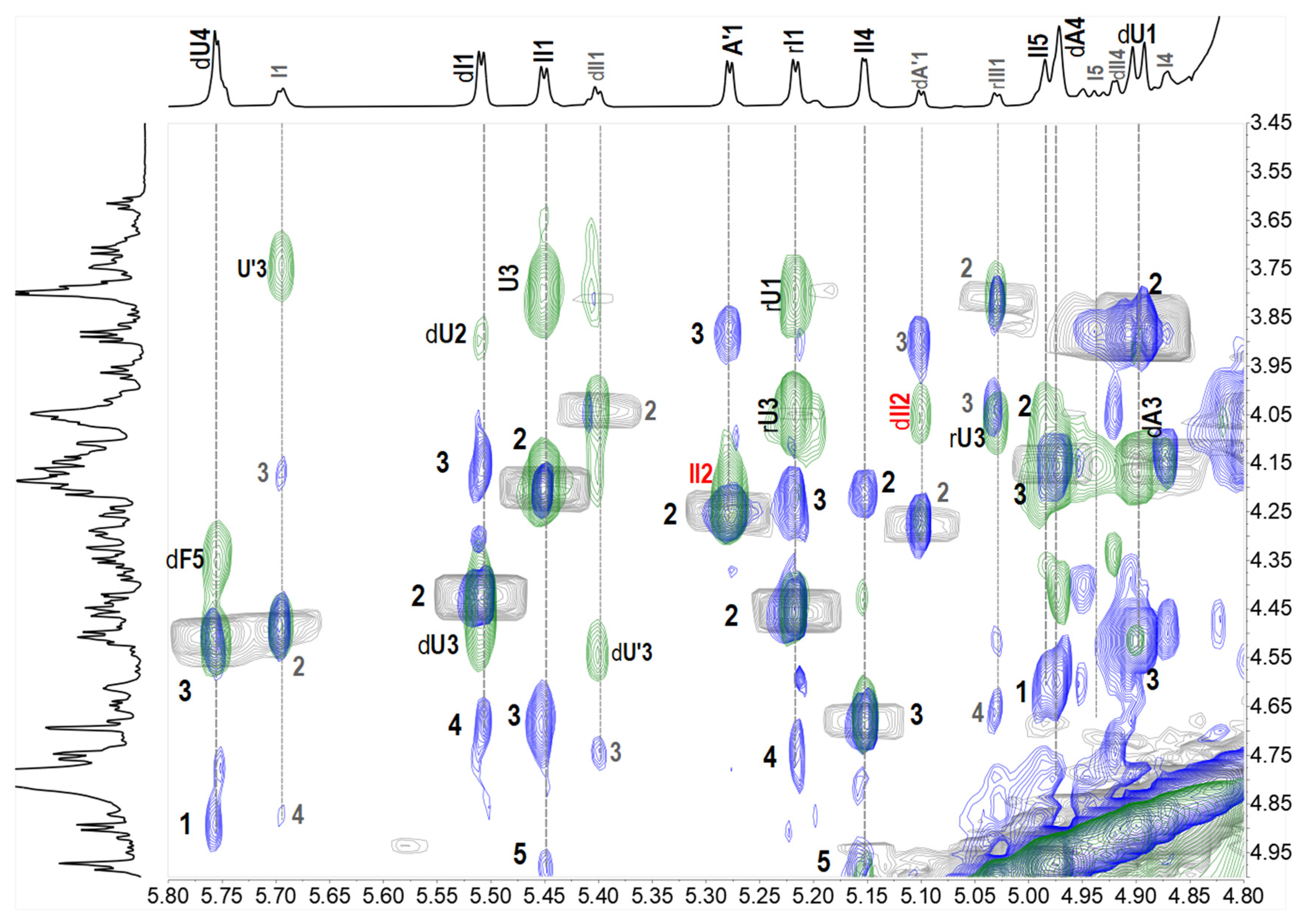
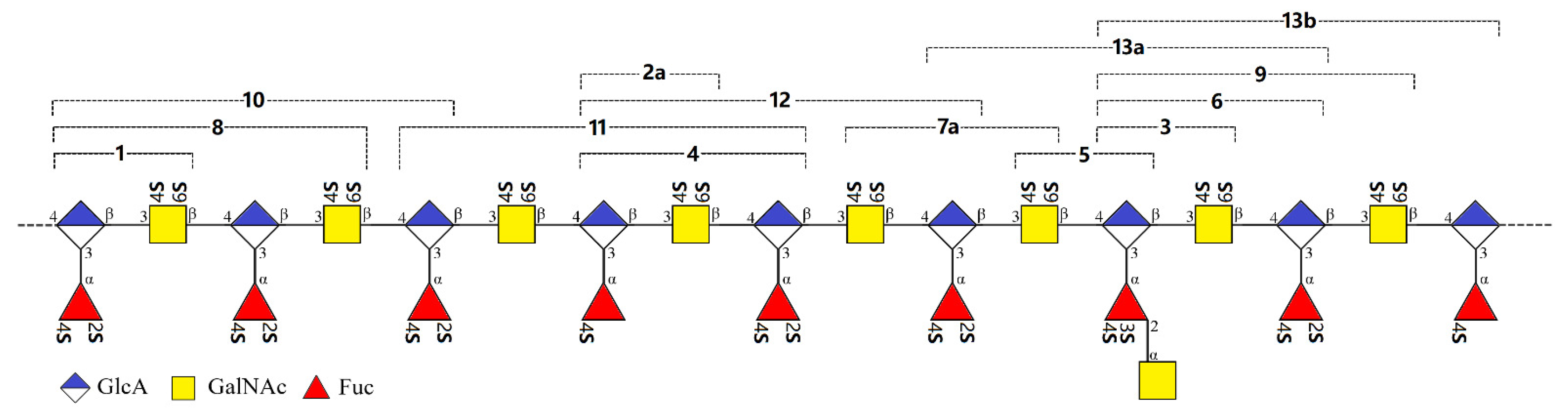
2.2. Structural Elucidation of the Oligosaccharides
2.3. Structural Confirmation of the Native FCSAm
2.4. Neurite Outgrowth-Promoting Activity of FCSAm and Its Derivatives
3. Materials and Methods
3.1. Materials
3.2. Depolymerization of FCSAm
3.3. Preparation of Size-Homogenous Fractions and Oligosaccharides from Depolymerizated FCSAm
3.4. Structural Elucidation of the Oligosaccharides
3.5. Neuronal Cultures and Coating Plates
3.6. Immunofluorescence
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dow, K.E.; Mirski, S.; Roder, J.; Riopelle, R. Neuronal proteoglycans: Biosynthesis and functional interaction with neurons in vitro. J. Neurosci. 1988, 8, 3278–3289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snow, D.M.; Letourneau, P.C. Neurite outgrowth on a step gradient of chondroitin sulfate proteoglycan (CS-PG). J. Neurosci. 1992, 23, 322–336. [Google Scholar] [CrossRef] [PubMed]
- Zuo, J.; Neubauer, D.; Dyess, K.; Ferguson, T.A.; Muir, D. Degradation of chondroitin sulfate proteoglycan enhances the neurite-promoting potential of spinal cord tissue. Exp. Neurol. 1998, 154, 654–662. [Google Scholar] [CrossRef] [PubMed]
- Maeda, N.; Noda, M. 6B4 proteoglycan/phosphacan is a repulsive substratum but promotes morphological differentiation of cortical neurons. Development 1996, 122, 647–658. [Google Scholar] [CrossRef]
- Hikino, M.; Mikami, T.; Faissner, A.; Vilela-Silva, A.-C.E.S.; Pavão, M.S.G.; Sugahara, K. Oversulfated dermatan sulfate exhibits neurite outgrowth-promoting activity toward embryonic mouse hippocampal neurons. J. Biol. Chem. 2003, 278, 43744–43754. [Google Scholar] [CrossRef] [Green Version]
- Sugahara, K.; Mikami, T.; Uyama, T.; Mizuguchi, S.; Nomura, K.; Kitagawa, H. Recent advances in the structural biology of chondroitin sulfate and dermatan sulfate. Curr. Opin. Struct. Biol. 2003, 13, 612–620. [Google Scholar] [CrossRef]
- Tully, S.E.; Mabon, R.; Gama, C.I.; Tsai, S.M.; Liu, X.; Hsieh-Wilson, L.C. A chondroitin sulfate small molecule that stimulates neuronal growth. J. Am. Chem. Soc. 2004, 126, 7736–7737. [Google Scholar] [CrossRef] [Green Version]
- Gilbert, R.J.; McKeon, R.J.; Darr, A.; Calabro, A.; Hascall, V.C.; Bellamkonda, R.V. CS-4,6 is differentially upregulated in glial scar and is a potent inhibitor of neurite extension. Mol. Cell. Neurosci. 2005, 29, 545–558. [Google Scholar] [CrossRef]
- Kim, M.H.; Park, S.R.; Choi, B.H. Comparative analysis of the expression of chondroitin sulfate subtypes and their inhibitory effect on axonal growth in the embryonic, adult, and injured rat brains. Tissue Eng. Regener. Med. 2020, 18, 165–178. [Google Scholar] [CrossRef]
- Clement, A.M.; Sugahara, K.; Faissner, A. Chondroitin sulfate E promotes neurite outgrowth of rat embryonic day 18 hippocampal neurons. Neurosci. Lett. 1999, 269, 125–128. [Google Scholar] [CrossRef]
- Higashi, K.; Takeda, K.; Mukuno, A.; Okamoto, Y.; Masuko, S.; Linhardt, R.J.; Toida, T. Identification of keratan sulfate disaccharide at C-3 position of glucuronate of chondroitin sulfate from Mactra chinensis. Biochem. J. 2016, 473, 4145–4158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shida, M.; Mikami, T.; Tamura, J.-I.; Kitagawa, H. A characteristic chondroitin sulfate trisaccharide unit with a sulfated fucose branch exhibits neurite outgrowth-promoting activity: Novel biological roles of fucosylated chondroitin sulfates isolated from the sea cucumber Apostichopus japonicus. Biochem. Biophys. Res. Commun. 2017, 487, 678–683. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Zhou, Q.; Liu, B.; Chen, F.; Wang, M. Holothurian fucosylated chondroitin sulfates and their potential benefits for human health: Structures and biological activities. Carbohydr. Polym. 2022, 275, 118691. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Yuan, Q.; Lv, K.; Ma, H.; Gao, C.; Liu, Y.; Zhang, S.; Zhao, L. Low-molecular-weight fucosylated glycosaminoglycan and its oligosaccharides from sea cucumber as novel anticoagulants: A review. Carbohydr. Polym. 2021, 251, 117034. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Wu, M.; Xiao, C.; Yang, L.; Zhou, L.; Gao, N.; Li, Z.; Chen, J.; Chen, J.; Liu, J.; et al. Discovery of an intrinsic tenase complex inhibitor: Pure nonasaccharide from fucosylated glycosaminoglycan. Proc. Natl. Acad. Sci. USA 2015, 112, 8284–8289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, R.; Zhou, L.; Gao, N.; Li, Z.; Zhao, L.; Shang, F.; Wu, M.; Zhao, J. Oligosaccharides from depolymerized fucosylated glycosaminoglycan: Structures and minimum size for intrinsic factor Xase complex inhibition. J. Biol. Chem. 2018, 293, 14089–14099. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Zhong, W.; Pan, Y.; Lin, L.; Cai, Y.; Mao, H.; Zhang, T.; Li, S.; Chen, R.; Zhou, L.; et al. Structural characterization and anticoagulant analysis of the novel branched fucosylated glycosaminoglycan from sea cucumber Holothuria nobilis. Carbohydr. Polym. 2021, 269, 118290. [Google Scholar] [CrossRef]
- Yin, R.; Zhou, L.; Gao, N.; Lin, L.; Sun, H.; Chen, D.; Cai, Y.; Zuo, Z.; Hu, K.; Huang, S.; et al. Unveiling the disaccharide-branched glycosaminoglycan and anticoagulant potential of its derivatives. Biomacromolecules 2021, 22, 1244–1255. [Google Scholar] [CrossRef]
- Yin, R.; Pan, Y.; Cai, Y.; Yang, F.; Gao, N.; Ruzemaimaiti, D.; Zhao, J. Re-understanding of structure and anticoagulation: Fucosylated chondroitin sulfate from sea cucumber Ludwigothurea grisea. Carbohydr. Polym. 2022, 294, 119826. [Google Scholar] [CrossRef]
- Huang, N.; Wu, M.; Zheng, C.; Zhu, L.; Zhao, J.; Zheng, Y. The depolymerized fucosylated chondroitin sulfate from sea cucumber potently inhibits HIV replication via interfering with virus entry. Carbohydr. Res. 2013, 380, 64–69. [Google Scholar] [CrossRef]
- Li, X.; Luo, L.; Cai, Y.; Yang, W.; Lin, L.; Li, Z.; Gao, N.; Purcell, S.W.; Wu, M.; Zhao, J. Structural elucidation and biological activity of a highly regular fucosylated glycosaminoglycan from the edible sea cucumber Stichopus herrmanni. J. Agric. Food Chem. 2017, 65, 9315–9323. [Google Scholar] [CrossRef] [PubMed]
- Pomin, V.H.; Vignovich, W.P.; Gonzales, A.V.; Vasconcelos, A.A.; Mulloy, B. Galactosaminoglycans: Medical applications and drawbacks. Molecules 2019, 24, 2803. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, H.; Cai, Y.; Li, S.; Sun, H.; Lin, L.; Pan, Y.; Yang, W.; He, Z.; Chen, R.; Zhou, L.; et al. A new fucosylated glycosaminoglycan containing disaccharide branches from Acaudina molpadioides: Unusual structure and anti-intrinsic tenase activity. Carbohydr. Polym. 2020, 245, 116503. [Google Scholar] [CrossRef]
- Brewer, G.J.; Torricelli, J.; Evege, E.; Price, P. Optimized survival of hippocampal neurons in B27-supplemented neurobasal™, a new serum-free medium combination. J. Neurosci. Res. 1993, 35, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Brewer, G.J.; Torricelli, J.R. Isolation and culture of adult neurons and neurospheres. Nat. Protoc. 2007, 2, 1490–1498. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.; Cong, R.; Na, L.; Ju, G.; You, S.-W. Long-term primary culture of highly-pure rat embryonic hippocampal neurons of low-density. Neurochem. Res. 2010, 35, 1333–1342. [Google Scholar] [CrossRef]
- Gama, C.I.; Tully, S.E.; Sotogaku, N.; Clark, P.M.; Rawat, M.; Vaidehi, N.; Goddard, W.A.; Nishi, A.; Hsieh-Wilson, L.C. Sulfation patterns of glycosaminoglycans encode molecular recognition and activity. Nat. Chem. Biol. 2006, 2, 467–473. [Google Scholar] [CrossRef] [Green Version]
- Sotogaku, N.; Tully, S.E.; Gama, C.I.; Higashi, H.; Tanaka, M.; Hsieh-Wilson, L.C.; Nishi, A. Activation of phospholipase C pathways by a synthetic chondroitin sulfate-E tetrasaccharide promotes neurite outgrowth of dopaminergic neurons. J. Neurochem. 2007, 103, 749–760. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comp. | Residue | δH, ppm | δC, ppm | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| H1 | H2 | H3 | H4 | H5 | H6 | H8 | C1 | C2 | C3 | C4 | C5 | C6 | C7 | C8 | ||
| 9 | A’ | 5.09 | 4.27 | 3.89 | 4.00 | 4.17 | 3.78/3.74 | 2.09 | 102.4 | 52.1 | 71.0 | 71.5 | 74.6 | 64.3 | 178.0 | 25.1 |
| II | 5.40 | 4.04 | 4.74 | 4.91 | 4.34 | 1.31 | 101.1 | 75.1 | 76.7 | 81.9 | 69.2 | 18.6 | ||||
| dU | 4.90 | 3.84 | 4.53 | 5.74 | 105.8 | 73.3 | 78.9 | 109.4 | 149.6 | 171.6 | ||||||
| A | 4.62 | 4.12 | 4.18 | 4.98 | 4.06 | 4.30/4.25 | 2.06 | 102.4 | 54.2 | 78.7 | 79.2 | 74.5 | 70.2 | 177.8 | 25.3 | |
| I | 5.68 | 4.50 | 4.15 | 4.86 | 4.96 | 1.38 | 99.6 | 77.9 | 69.3 | 83.9 | 69.1 | 18.5 | ||||
| U | 4.60 | 3.73 | 3.79 | 3.94 | 3.79 | 105.8 | 77.1 | 80.3 | 78.1 | 79.0 | 178.0 | |||||
| rA | 3.70 | 4.27 | 4.28 | 4.50 | 4.41 | 4.15 | 2.01 | 62.5 | 54.4 | 77.8 | 80.6 | 70.3 | 72.4 | 177.0 | 24.8 | |
| 10 | dI | 5.51 | 4.44 | 4.15 | 4.70 | 4.36 | 1.31 | 99.0 | 77.7 | 69.2 | 83.5 | 69.1 | 18.4 | |||
| dU | 4.91 | 3.90 | 4.51 | 5.76 | 105.9 | 73.1 | 79.1 | 109.1 | 149.8 | 171.6 | ||||||
| dA | 4.61 | 4.12 | 4.15 | 4.98 | 4.04 | 4.31/4.20 | 2.06 | 102.6 | 54.1 | 78.7 | 79.1 | 75.0 | 70.1 | 177.7 | 25.3 | |
| I | 5.70 | 4.49 | 4.16 | 4.88 | 4.94 | 1.39 | 99.3 | 77.8 | 69.3 | 83.9 | 69.1 | 18.6 | ||||
| U | 4.48 | 3.67 | 3.76 | 3.96 | 3.70 | 106.6 | 76.6 | 79.9 | 78.1 | 79.7 | 177.7 | |||||
| rA | 4.78 | 4.07 | 4.01 | 4.83 | 4.06 | 4.30/4.20 | 2.06 | 103.5 | 54.3 | 78.9 | 79.1 | 74.6 | 70.5 | 177.7 | 25.3 | |
| rI | 5.22 | 4.46 | 4.21 | 4.74 | 4.59 | 1.33 | 101.3 | 78.0 | 69.3 | 83.8 | 69.5 | 18.7 | ||||
| rU | 3.83 | 4.05 | 4.05 | 4.17 | 4.32 | 65.1 | 73.3 | 80.6 | 83.0 | 75.0 | 180.0 | |||||
| 11 | dI | 5.50 | 4.42 | 4.14 | 4.69 | 4.35 | 1.30 | 99.0 | 77.7 | 69.1 | 83.4 | 69.0 | 18.4 | |||
| dU | 4.90 | 3.88 | 4.50 | 5.75 | 105.9 | 73.1 | 79.1 | 109.1 | 149.8 | 171.6 | ||||||
| dA | 4.60 | 4.11 | 4.16 | 4.96 | 4.04 | 4.31/4.18 | 2.05 | 102.5 | 54.1 | 78.7 | 79.1 | 74.6 | 70.5 | 177.7 | 25.3 | |
| III | 5.40 | 3.80 | 4.04 | 4.77 | 4.88 | 1.37 | 101.3 | 71.1 | 71.5 | 83.9 | 69.0 | 18.7 | ||||
| U | 4.48 | 3.62 | 3.69 | 4.00 | 3.69 | 106.3 | 76.6 | 80.2 | 78.3 | 79.8 | 177.7 | |||||
| rA | 4.79 | 4.04 | 4.04 | 4.81 | 4.04 | 4.28/4.18 | 2.05 | 103.4 | 54.3 | 78.2 | 79.1 | 74.7 | 70.5 | 177.7 | 25.3 | |
| rI | 5.22 | 4.44 | 4.20 | 4.73 | 4.58 | 1.32 | 101.2 | 78.0 | 69.2 | 83.8 | 69.5 | 18.7 | ||||
| rU | 3.80 | 4.03 | 4.03 | 4.15 | 4.32 | 65.1 | 73.4 | 80.7 | 82.8 | 74.9 | 180.1 | |||||
| 12 | dIII | 5.20 | 3.80 | 4.01 | 4.62 | 4.33 | 1.29 | 101.4 | 71.1 | 71.2 | 83.4 | 69.1 | 18.6 | |||
| dU | 4.94 | 3.88 | 4.41 | 5.75 | 105.5 | 72.9 | 78.8 | 109.5 | 149.3 | 171.7 | ||||||
| dA | 4.61 | 4.11 | 4.16 | 4.95 | 4.06 | 4.30/4.20 | 2.06 | 102.5 | 54.2 | 78.7 | 79.0 | 75.0 | 70.0 | 177.4 | 25.3 | |
| I | 5.69 | 4.49 | 4.16 | 4.86 | 4.93 | 1.37 | 99.3 | 77.8 | 69.1 | 83.9 | 69.0 | 18.6 | ||||
| U | 4.47 | 3.66 | 3.75 | 3.95 | 3.69 | 106.6 | 76.5 | 80.0 | 78.2 | 79.7 | 177.7 | |||||
| rA | 4.77 | 4.02 | 4.02 | 4.82 | 4.03 | 4.29/4.18 | 2.06 | 103.5 | 54.3 | 78.9 | 79.0 | 74.6 | 70.5 | 177.7 | 25.3 | |
| rI | 5.22 | 4.45 | 4.21 | 4.74 | 4.58 | 1.32 | 101.3 | 78.0 | 69.3 | 83.8 | 69.5 | 18.7 | ||||
| rU | 3.79 | 4.03 | 4.04 | 4.17 | 4.31 | 65.1 | 73.3 | 80.7 | 83.0 | 74.9 | 180.0 | |||||
| 13a | dI | 5.51 | 4.43 | 4.10 | 4.69 | 4.35 | 1.30 | 98.9 | 77.7 | 69.1 | 83.5 | 69.1 | 18.4 | |||
| dU | 4.90 | 3.89 | 4.52 | 5.76 | 106.0 | 73.1 | 79.0 | 109.0 | 149.9 | 171.6 | ||||||
| dA | 4.61 | 4.17 | 4.16 | 4.97 | 4.06 | 4.38/4.30 | 2.06 | 102.6 | 54.2 | 78.7 | 79.0 | 74.5 | 70.5 | 177.8 | 25.4 | |
| A’ | 5.28 | 4.25 | 3.88 | 4.11 | 4.18 | 3.78/3.74 | 2.09 | 100.5 | 52.1 | 70.9 | 71.4 | 74.5 | 64.3 | 177.8 | 25.4 | |
| II | 5.45 | 4.15 | 4.69 | 5.15 | 4.98 | 1.43 | 101.2 | 74.7 | 78.1 | 82.2 | 69.2 | 18.7 | ||||
| U | 4.50 | 3.62 | 3.80 | 3.96 | 3.71 | 105.6 | 76.3 | 80.4 | 78.0 | 80.1 | 177.8 | |||||
| rA | 4.80 | 4.05 | 4.13 | 4.76 | 4.06 | 4.29/4.20 | 2.05 | 103.3 | 54.3 | 76.2 | 79.1 | 74.8 | 70.5 | 178.1 | 25.3 | |
| rI | 5.22 | 4.46 | 4.21 | 4.75 | 4.64 | 1.33 | 101.0 | 78.0 | 69.2 | 83.8 | 69.5 | 18.7 | ||||
| rU | 3.80 | 4.02 | 4.05 | 4.19 | 4.33 | 65.1 | 73.5 | 80.5 | 82.8 | 75.1 | 180.1 | |||||
| Comp. | Molecular Ions | m/z | Molecular Formula | Mw | |
|---|---|---|---|---|---|
| Observed | Calculated | ||||
| 8 | [M−3Na]3− [M−4Na + H]3− | 614.2876 606.9607 | 614.2869 606.9596 | C40H54O54N2S8Na10 | 1913.2097 |
| 9 | [M−3Na + H]2− [M−4Na + H]3−[M−3Na]3− | 1023.4730 674.6529 681.9800 | 1023.4737 674.6527 681.9800 | C48H67O59N3S8Na10 | 2116.4037 |
| 10 | [M−4Na + H]3− [M−5Na + H]4− [M−6Na + H]5− | 789.6083 586.4597 464.5697 | 789.6094 586.4597 464.5700 | C52H69O70N2S10Na13 | 2461.5350 |
| 11 | [M−3Na]3− [M−4Na]4− | 762.9568 566.4712 | 762.9571 566.4705 | C52H70O67N2S9Na12 | 2359.4962 |
| 12 | [M−4Na + H]3− [M−5Na + H]4− [M−6Na + H]5− | 755.6310 560.9767 444.1845 | 755.6298 560.9751 444.1822 | C52H70O67N2S9Na12 | 2359.4962 |
| 13a | [M−4Na + H]3− | 857.3014 | 857.3025 | C60H82O75N3S10Na13 | 2664.7290 |
| [M−5Na + H]4− [M−4Na]4− | 637.2300 642.7250 | 637.2296 642.7251 | |||
| 13b | [M−3Na]3− | 830.6498 | 830.6503 | C60H83O72N3S9Na12 | 2562.6902 |
| [M−4Na + H]3− [M−4Na]4− | 823.3222 617.2408 | 823.3229 617.2404 | |||
| Treatment | Mw (kDa) | Conc. (μg/mL) | Neurite Length (% of Control) |
|---|---|---|---|
| F2 | ~1.5 a | 50 | 152.10 ± 21.18 |
| F3 | ~2.5a | 50 | 179.70 ± 6.83 ** |
| F4 | ~3.5 a | 50 | 188.60 ± 9.83 ** |
| F5 | ~4.4a | 50 | 218.50 ± 13.24 ** |
| dFCSAm | 4.13 b | 50 | 150.40 ± 13.07 |
| FCSAm | 51.7b | 50 | 148.90 ± 9.13 * |
| CS-E | 72c | 50 | 112.00 ± 27.06 |
| Control | / | - | 100 ± 3.10 |
| Conc. (μM) | 1 | 3 | 4 | 5 | 6 | 8 |
| 4.4 | 116.30 ± 18.76 | 130.80 ± 12.35 | 138.80 ± 21.85 | 129.40 ± 14.98 | 144.70 ± 13.40 | 146.10 ± 5.16 |
| 13.3 | 140.20 ± 13.45 | 146.00 ± 16.81 | 141.20 ± 16.85 | 147.10 ± 15.19 | 175.60 ± 9.89 *** | 177.90 ± 16.96 *** |
| 40 | 149.10 ± 16.68 | 159.80 ± 4.43 ** | 164.30 ± 10.43 ** | 171.80 ± 23.78 *** | 186.40 ± 4.10 *** | 189.70 ± 18.06 *** |
| Conc. (μM) | 9 | 10 | 11 | 12 | 13 | |
| 4.4 | 146.00 ± 12.51 | 167.80 ± 25.99 | 133.60 ± 14.12 | 160.30 ± 20.22 ** | 160.80 ± 7.96 ** | |
| 13.3 | 206.00 ± 6.56 *** | 202.90 ± 10.72 *** | 157.10 ± 11.42 * | 187.60 ± 12.16 *** | 196.20 ± 15.59 *** | |
| 40 | 193.40 ± 18.15 *** | 206.60 ± 25.99 *** | 203.20 ± 11.42 *** | 217.40 ± 22.29 *** | 216.60 ± 31.12 *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; Mao, H.; Li, S.; Zhang, L.; Yang, L.; Yin, R.; Zhao, J. Branched Chondroitin Sulfate Oligosaccharides Derived from the Sea Cucumber Acaudina molpadioides Stimulate Neurite Outgrowth. Mar. Drugs 2022, 20, 653. https://doi.org/10.3390/md20100653
Wang W, Mao H, Li S, Zhang L, Yang L, Yin R, Zhao J. Branched Chondroitin Sulfate Oligosaccharides Derived from the Sea Cucumber Acaudina molpadioides Stimulate Neurite Outgrowth. Marine Drugs. 2022; 20(10):653. https://doi.org/10.3390/md20100653
Chicago/Turabian StyleWang, Weili, Hui Mao, Sujuan Li, Longlong Zhang, Lian Yang, Ronghua Yin, and Jinhua Zhao. 2022. "Branched Chondroitin Sulfate Oligosaccharides Derived from the Sea Cucumber Acaudina molpadioides Stimulate Neurite Outgrowth" Marine Drugs 20, no. 10: 653. https://doi.org/10.3390/md20100653