
Marine Antimicrobial Peptide TP4 Exerts Anticancer Effects on Human Synovial Sarcoma Cells via Calcium Overload, Reactive Oxygen Species Production and Mitochondrial Hyperpolarization

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
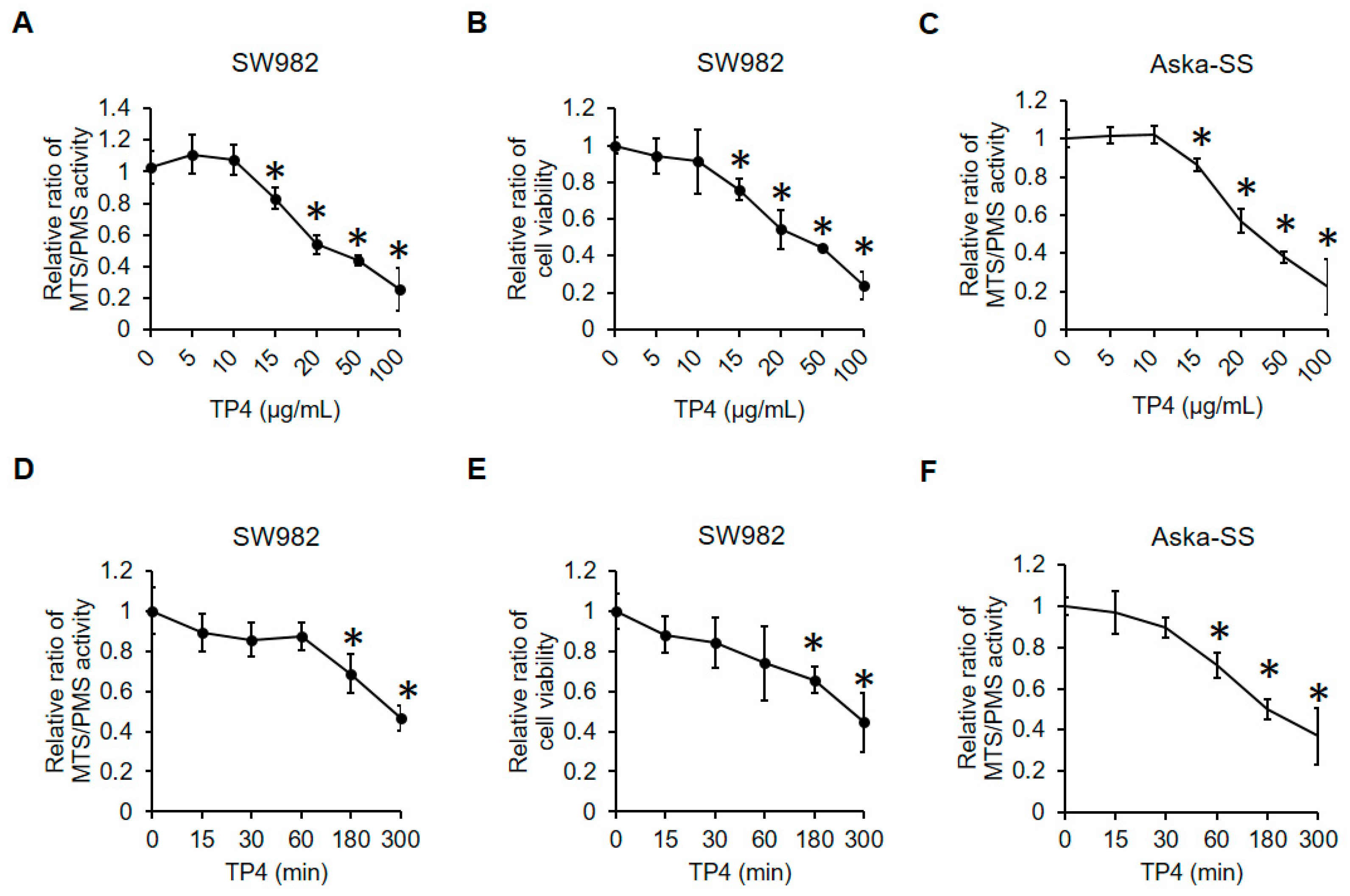
2.1. TP4 Decreases Cell Viability in Human Synovial Sarcoma Cells
2.2. TP4 Induces Necrotic Cell Death in Human Synovial Sarcoma Cells
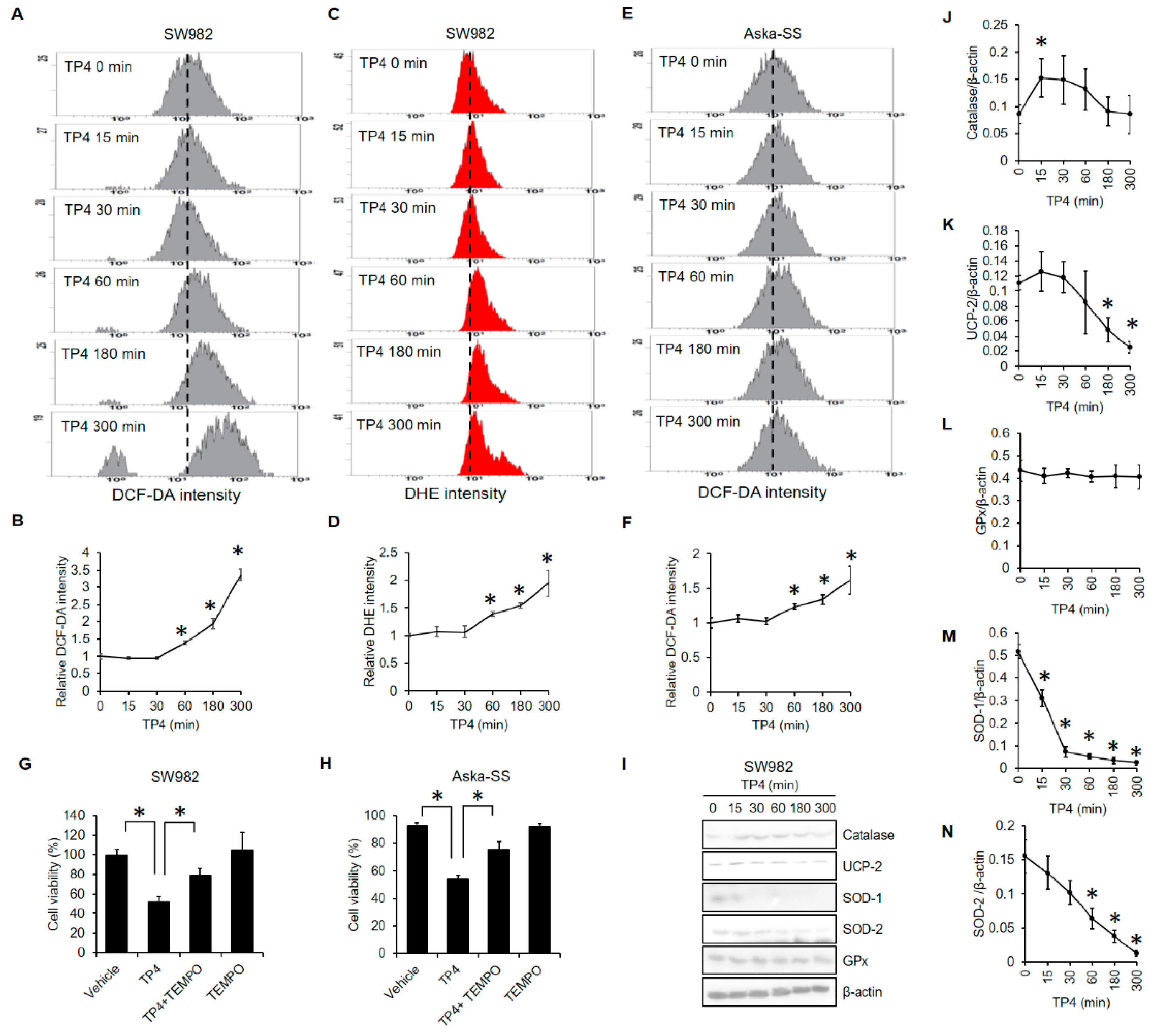
2.3. TP4 Increased ROS Levels and Decreased Antioxidant Proteins
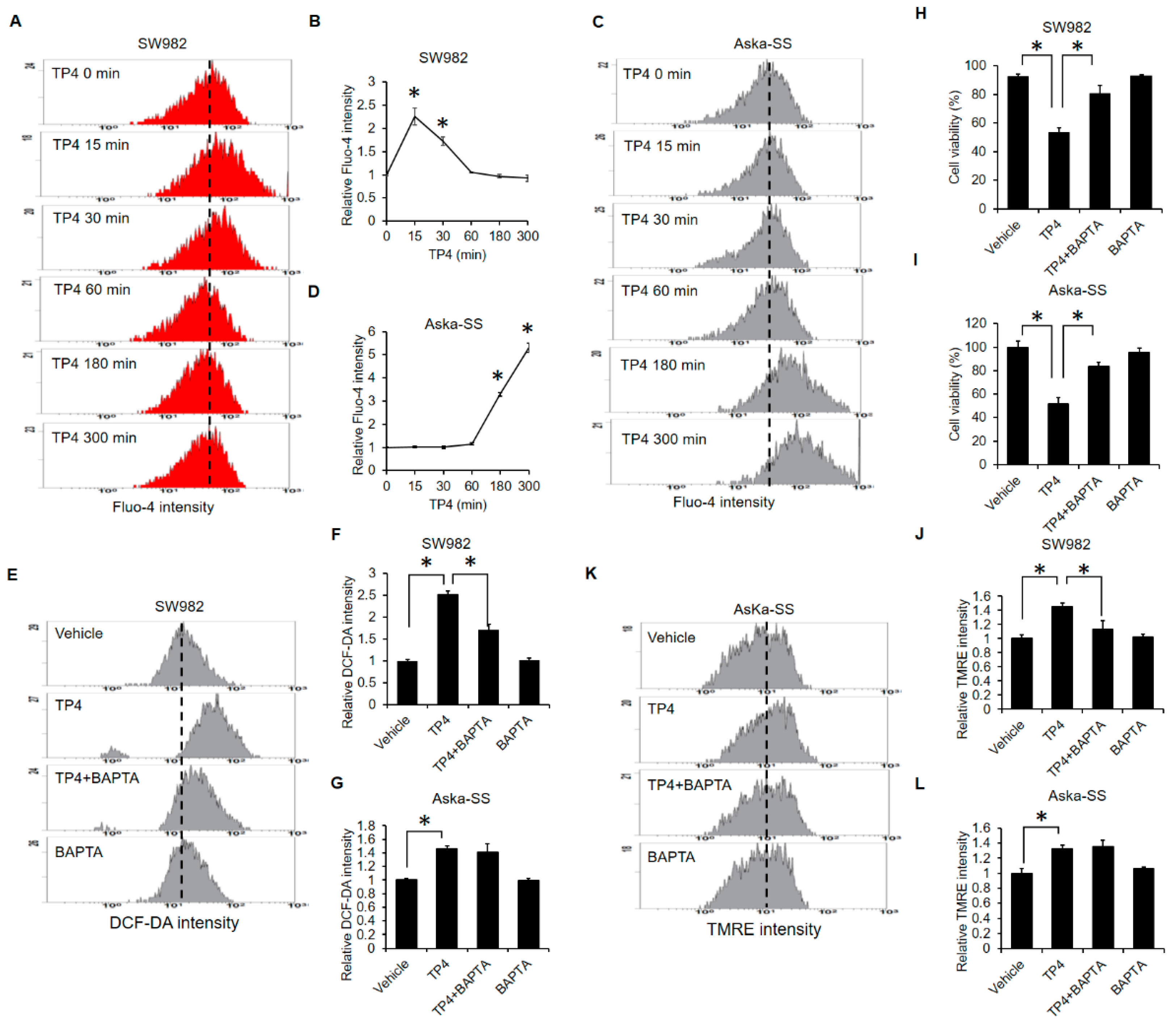
2.4. TP4 Initiates Mitochondrial Hyperpolarization
2.5. Calcium-Overload Is Required for TP4-Induced Cytotoxicity
3. Discussion
4. Materials and Methods
4.1. Reagents
4.2. Cell Culture and Treatments
4.3. Cytotoxicity and PI Exclusion Assay
4.4. Western Blot
4.5. Flow Cytometry
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Joseph, N.; St Laurent, S.; Nelson, J.J.; Zheng, S.; Stirnadel-Farrant, H. Incidence and prevalence of synovial sarcoma in the US: An analysis using SEER(star)Stat. J. Clin. Oncol. 2019, 37. [Google Scholar] [CrossRef]
- Hung, G.Y.; Horng, J.L.; Chen, P.C.H.; Lin, L.Y.; Chen, J.Y.; Chuang, P.H.; Chao, T.C.; Yen, C.C. Incidence of soft tissue sarcoma in Taiwan: A nationwide population-based study (2007–2013). Cancer Epidemiol. 2019, 60, 185–192. [Google Scholar] [CrossRef]
- Ferrari, A.; Sultan, I.; Huang, T.T.; Rodriguez-Galindo, C.; Shehadeh, A.; Meazza, C.; Ness, K.K.; Casanova, M.; Spunt, S.L. Soft Tissue Sarcoma Across the Age Spectrum: A Population-Based Study from the Surveillance Epidemiology and End Results Database. Pediatr. Blood Cancer 2011, 57, 943–949. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, T.O.; Poulin, N.M.; Ladanyi, M. Synovial Sarcoma: Recent Discoveries as a Roadmap to New Avenues for Therapy. Cancer Discov. 2015, 5, 124–134. [Google Scholar] [CrossRef] [Green Version]
- Saito, T. The SYT-SSX fusion protein and histological epithelial differentiation in synovial sarcoma: Relationship with extracellular matrix remodeling. Int. J. Clin. Exp. Patho. 2013, 6, 2272–2279. [Google Scholar]
- Guillou, L.; Benhattar, L.; Bonichon, F.; Gallagher, G.; Terrier, P.; Stauffer, E.; Somerhausen, N.D.A.; Michels, J.J.; Jundt, G.; Vince, D.R.; et al. Histologic grade, but not SYT-SSX fusion type, is an important prognostic factor in patients with synovial sarcoma: A multicenter, retrospective analysis. J. Clin. Oncol. 2004, 22, 4040–4050. [Google Scholar] [CrossRef] [PubMed]
- Ladanyi, M.; Antonescu, C.R.; Leung, D.H.; Woodruff, J.M.; Kawai, A.; Healey, J.H.; Brennan, M.F.; Bridge, J.A.; Neff, J.R.; Barr, F.G.; et al. Impact of SYT-SSX fusion type on the clinical behavior of synovial sarcoma: A multi-institutional retrospective study of 243 patients. Cancer Res. 2002, 62, 135–140. [Google Scholar] [PubMed]
- Corey, R.M.; Swett, K.; Ward, W.G. Epidemiology and survivorship of soft tissue sarcomas in adults: A national cancer database report. Cancer Med. 2014, 3, 1404–1415. [Google Scholar] [CrossRef]
- Naing, K.W.; Monjazeb, A.M.; Li, C.S.; Lee, L.Y.; Yang, A.; Borys, D.; Canter, R.J. Perioperative Radiotherapy is Associated with Improved Survival Among Patients with Synovial Sarcoma: A SEER Analysis. J. Surg. Oncol. 2015, 111, 158–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vlenterie, M.; Ho, V.K.Y.; Kaal, S.E.J.; Vlenterie, R.; Haas, R.; van der Graaf, W.T.A. Age as an independent prognostic factor for survival of localised synovial sarcoma patients. Br. J. Cancer 2015, 113, 1602–1606. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmerini, E.; Staals, E.L.; Alberghini, M.; Zanella, L.; Ferrari, C.; Benassi, M.S.; Picci, P.; Mercuri, M.; Bacci, G.; Ferrari, S. Synovial sarcoma: Retrospective analysis of 250 patients treated at a single institution. Cancer 2009, 115, 2988–2998. [Google Scholar] [CrossRef] [PubMed]
- Stanelle, E.J.; Christison-Lagay, E.R.; Healey, J.H.; Singer, S.; Meyers, P.A.; La Quaglia, M.P. Pediatric and Adolescent Synovial Sarcoma: Multivariate Analysis of Prognostic Factors and Survival Outcomes. Ann. Surg. Oncol. 2013, 20, 73–79. [Google Scholar] [CrossRef]
- Krieg, A.H.; Hefti, F.; Speth, B.M.; Jundt, G.; Guillou, L.; Exner, U.G.; von Hochstetter, A.R.; Cserhati, M.D.; Fuchs, B.; Mouhsine, E.; et al. Synovial sarcomas usually metastasize after >5 years: A multicenter retrospective analysis with minimum follow-up of 10 years for survivors. Ann. Oncol. 2011, 22, 458–467. [Google Scholar] [CrossRef]
- Desar, I.M.E.; Fleuren, E.D.G.; van der Graaf, W.T.A. Systemic Treatment for Adults with Synovial Sarcoma. Curr. Treat. Option Oncol. 2018, 19, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayodele, O.; Razak, A.R.A. Immunotherapy in soft-tissue sarcoma. Curr. Oncol. 2020, 27, 17–23. [Google Scholar] [CrossRef] [PubMed]
- Stacchiotti, S.; Van Tine, B.A. Synovial Sarcoma: Current Concepts and Future Perspectives. J. Clin. Oncol. 2018, 36, 180–187. [Google Scholar] [CrossRef]
- Peng, K.C.; Lee, S.H.; Hour, A.L.; Pan, C.Y.; Lee, L.H.; Chen, J.Y. Five Different Piscidins from Nile Tilapia, Oreochromis niloticus: Analysis of Their Expressions and Biological Functions. PLoS ONE 2012, 7, e50263. [Google Scholar] [CrossRef]
- Hazam, P.K.; Chen, J.Y. Therapeutic utility of the antimicrobial peptide Tilapia Piscidin 4 (TP4). Aquac. Rep. 2020, 17, 100409. [Google Scholar] [CrossRef]
- Narayana, J.L.; Huang, H.N.; Wu, C.J.; Chen, J.Y. Efficacy of the antimicrobial peptide TP4 against Helicobacter pylori infection: In vitro membrane perturbation via micellization and in vivo suppression of host immune responses in a mouse model. Oncotarget 2015, 6, 12936–12954. [Google Scholar] [CrossRef] [Green Version]
- Pan, C.Y.; Tsai, T.Y.; Su, B.C.; Hui, C.F.; Chen, J.Y. Study of the Antimicrobial Activity of Tilapia Piscidin 3 (TP3) and TP4 and Their Effects on Immune Functions in Hybrid Tilapia (Oreochromis spp.). PLoS ONE 2017, 12, e0169678. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.N.; Chan, Y.L.; Wu, C.J.; Chen, J.Y. Tilapia Piscidin 4 (TP4) Stimulates Cell Proliferation and Wound Closure in MRSA-Infected Wounds in Mice. Mar. Drugs 2015, 13, 2813–2833. [Google Scholar] [CrossRef]
- Su, B.C.; Pan, C.Y.; Chen, J.Y. Antimicrobial Peptide TP4 Induces ROS-Mediated Necrosis by Triggering Mitochondrial Dysfunction in Wild-Type and Mutant p53 Glioblastoma Cells. Cancers 2019, 11, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ting, C.H.; Chen, J.Y. Nile Tilapia Derived TP4 Shows Broad Cytotoxicity toward to Non-Small-Cell Lung Cancer Cells. Mar. Drugs 2018, 16, 506. [Google Scholar] [CrossRef] [Green Version]
- Ting, C.H.; Liu, Y.C.; Lyu, P.C.; Chen, J.Y. Nile Tilapia Derived Antimicrobial Peptide TP4 Exerts Antineoplastic Activity Through Microtubule Disruption. Mar. Drugs 2018, 16, 462. [Google Scholar] [CrossRef] [Green Version]
- Ting, C.H.; Chen, Y.C.; Wu, C.J.; Chen, J.Y. Targeting FOSB with a cationic antimicrobial peptide, TP4, for treatment of triple-negative breast cancer. Oncotarget 2016, 7, 40329–40347. [Google Scholar] [CrossRef]
- Su, B.C.; Li, C.C.; Horng, J.L.; Chen, J.Y. Calcium-Dependent Calpain Activation-Mediated Mitochondrial Dysfunction and Oxidative Stress Are Required for Cytotoxicity of Epinecidin-1 in Human Synovial Sarcoma SW982 Cells. Int. J. Mol. Sci. 2020, 21, 2109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Festjens, N.; Vanden Berghe, T.; Vandenabeele, P. Necrosis, a well-orchestrated form of cell demise: Signalling cascades, important mediators and concomitant immune response. Bba-Bioenerg. 2006, 1757, 1371–1387. [Google Scholar] [CrossRef]
- Choi, K.; Kim, J.; Kim, G.W.; Choi, C. Oxidative Stress-Induced Necrotic Cell Death via Mitochondira-Dependent Burst of Reactive Oxygen Species. Curr. Neurovasc. Res. 2009, 6, 213–222. [Google Scholar] [CrossRef] [PubMed]
- Zhivotovsky, B.; Orrenius, S. Calcium and cell death mechanisms: A perspective from the cell death community. Cell Calcium 2011, 50, 211–221. [Google Scholar] [CrossRef]
- Perillo, B.; Di Donato, M.; Pezone, A.; Di Zazzo, E.; Giovannelli, P.; Galasso, G.; Castoria, G.; Migliaccio, A. ROS in cancer therapy: The bright side of the moon. Exp. Mol. Med. 2020, 52, 192–203. [Google Scholar] [CrossRef] [PubMed]
- Cockfield, J.A.; Schafer, Z.T. Antioxidant Defenses: A Context-Specific Vulnerability of Cancer Cells. Cancers 2019, 11, 1208. [Google Scholar] [CrossRef] [Green Version]
- Epperly, M.W.; Epstein, C.J.; Travis, E.L.; Greenberger, J.S. Decreased pulmonary radiation resistance of manganese superoxide dismutase (MnSOD)-deficient mice is corrected by human manganese superoxide dismutase-plasmid/liposome (SOD2-PL) intratracheal gene therapy. Radiat. Res. 2000, 154, 365–374. [Google Scholar] [CrossRef]
- Qu, Y.M.; Zhao, S.P.; Hong, J.D.; Tang, S. Radiosensitive gene therapy through imRNA expression for silencing manganese superoxide dismutase. J. Cancer Res. Clin. 2010, 136, 953–959. [Google Scholar] [CrossRef] [PubMed]
- Samuels, B.L.; Murray, J.L.; Cohen, M.B.; Safa, A.R.; Sinha, B.K.; Townsend, A.J.; Beckett, M.A.; Weichselbaum, R.R. Increased Glutathione-Peroxidase Activity in a Human Sarcoma Cell-Line with Inherent Doxorubicin Resistance. Cancer Res. 1991, 51, 521–527. [Google Scholar]
- Hochwald, S.N.; Rose, D.M.; Brennan, M.F.; Burt, M.E. Elevation of glutathione and related enzyme activities in high-grade and metastatic extremity soft tissue sarcoma. Ann. Surg. Oncol. 1997, 4, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Nathan, F.M.; Singh, V.A.; Dhanoa, A.; Palanisamy, U.D. Oxidative stress and antioxidant status in primary bone and soft tissue sarcoma. BMC Cancer 2011, 11, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Toda, C.; Diano, S. Mitochondrial UCP2 in the central regulation of metabolism. Best Pract. Res. Clin. Endocrinol. 2014, 28, 757–764. [Google Scholar] [CrossRef]
- Pak, O.; Sommer, N.; Hoeres, T.; Bakr, A.; Waisbrod, S.; Sydykov, A.; Haag, D.; Esfandiary, A.; Kojonazarov, B.; Veit, F.; et al. Mitochondrial Hyperpolarization in Pulmonary Vascular Remodeling Mitochondrial Uncoupling Protein Deficiency as Disease Model. Am. J. Respir. Cell Mol. 2013, 49, 358–367. [Google Scholar] [CrossRef]
- Su, B.C.; Liu, Y.C.; Ting, C.H.; Lyu, P.C.; Chen, J.Y. Antimicrobial Peptide TP4 Targets Mitochondrial Adenine Nucleotide Translocator 2. Mar. Drugs 2020, 18, 417. [Google Scholar] [CrossRef]
- Pfeffer, C.M.; Singh, A.T.K. Apoptosis: A Target for Anticancer Therapy. Int. J. Mol. Sci. 2018, 19, 448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, H.L.; Yuen, K.L.; Tang, H.M.; Fung, M.C. Reversibility of apoptosis in cancer cells. Br. J. Cancer 2009, 100, 118–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaravadi, R.K.; Thompson, C.B. The roles of therapy-induced autophagy and necrosis in cancer treatment. Clin. Cancer Res. 2007, 13, 7271–7279. [Google Scholar] [CrossRef] [PubMed] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, B.-C.; Hung, G.-Y.; Tu, Y.-C.; Yeh, W.-C.; Lin, M.-C.; Chen, J.-Y. Marine Antimicrobial Peptide TP4 Exerts Anticancer Effects on Human Synovial Sarcoma Cells via Calcium Overload, Reactive Oxygen Species Production and Mitochondrial Hyperpolarization. Mar. Drugs 2021, 19, 93. https://doi.org/10.3390/md19020093
Su B-C, Hung G-Y, Tu Y-C, Yeh W-C, Lin M-C, Chen J-Y. Marine Antimicrobial Peptide TP4 Exerts Anticancer Effects on Human Synovial Sarcoma Cells via Calcium Overload, Reactive Oxygen Species Production and Mitochondrial Hyperpolarization. Marine Drugs. 2021; 19(2):93. https://doi.org/10.3390/md19020093
Chicago/Turabian StyleSu, Bor-Chyuan, Giun-Yi Hung, Yun-Chieh Tu, Wei-Chen Yeh, Meng-Chieh Lin, and Jyh-Yih Chen. 2021. "Marine Antimicrobial Peptide TP4 Exerts Anticancer Effects on Human Synovial Sarcoma Cells via Calcium Overload, Reactive Oxygen Species Production and Mitochondrial Hyperpolarization" Marine Drugs 19, no. 2: 93. https://doi.org/10.3390/md19020093