
Genomic Insights into Syntrophic Lifestyle of ‘Candidatus Contubernalis alkaliaceticus’ Based on the Reversed Wood–Ljungdahl Pathway and Mechanism of Direct Electron Transfer


{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Genome Sequencing and Assembly
2.2. Genome Annotation
2.3. Growth Experiments
2.4. Microscopy
3. Results and Discussion
3.1. General Genome Properties, Genome Mobility, and Genetic Immunity Systems
3.2. Genomic Determinants of ‘Ca. C. alkaliaceticus’ Metabolism
3.2.1. Syntrophic Acetate Oxidation
3.2.2. Hydrogen Production during Syntrophic Acetate Oxidation
3.2.3. Energy Conservation during Syntrophic Acetate Oxidation
3.2.4. Syntrophic Ethanol Oxidation
3.2.5. Carbon Metabolism
3.2.6. Direct Interspecies Electron Transfer
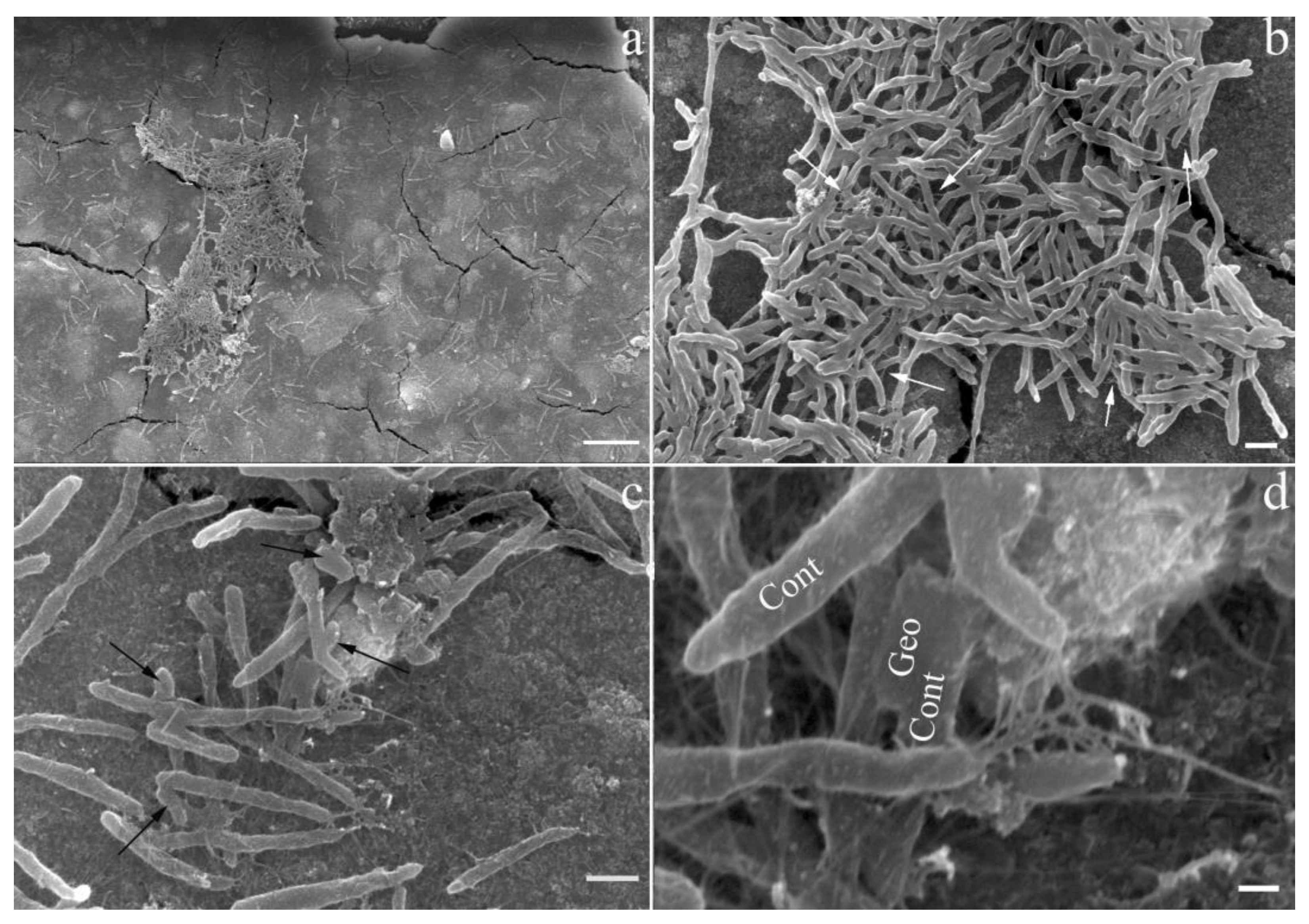
3.3. Biofilm Formation by ‘Ca. C. alkaliaceticus’ and Geoalkalibacter ferrihydriticus in a Binary Culture
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Westerholm, M.; Calusinska, M.; Dolfing, J. Syntrophic propionate-oxidizing bacteria in methanogenic systems. FEMS Microbiol. Rev. 2022, 46, fuab057. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R. Syntrophy goes electric: Direct interspecies electron transfer. Annu. Rev. Microbiol. 2017, 71, 643–664. [Google Scholar] [CrossRef] [PubMed]
- Morris, B.E.; Henneberger, R.; Huber, H.; Moissl-Eichinger, C. Microbial syntrophy: Interaction for the common good. FEMS Microbiol. Rev. 2013, 37, 384–406. [Google Scholar] [CrossRef]
- Stams, A.J.; Plugge, C.M. Electron transfer in syntrophic communities of anaerobic bacteria and archaea. Nat. Rev. Microbiol. 2009, 7, 568–577. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Abbas, B.; Tourova, T.P.; Bumazhkin, B.K.; Kolganova, T.V.; Muyzer, G. Sulfate-dependent acetate oxidation under extremely natron-alkaline conditions by syntrophic associations from hypersaline soda lakes. Microbiology 2014, 160, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Sorokin, D.Y.; Chernyh, N.A.; Poroshina, M.N. Desulfonatronobacter acetoxydans sp. nov.: A first acetate-oxidizing, extremely salt-tolerant alkaliphilic SRB from a hypersaline soda lake. Extremophiles 2015, 19, 899–907. [Google Scholar] [CrossRef]
- Zhilina, T.N.; Zavarzina, D.G.; Kolganova, T.V.; Tourova, T.P.; Zavarzin, G.A. “Candidatus Contubernalis alkaliaceticus”, an obligately syntrophic alkaliphilic bacterium capable of anaerobic acetate oxidation in a coculture with Desulfonatronum cooperativum. Microbiology 2005, 74, 695–703. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Abbas, B.; Geleijnse, M.; Kolganova, T.V.; Kleerebezem, R.; van Loosdrecht, M.C. Syntrophic associations from hypersaline soda lakes converting organic acids and alcohols to methane at extremely haloalkaline conditions. Environ. Microbiol. 2016, 18, 3189–3202. [Google Scholar] [CrossRef]
- Timmers, P.H.A.; Vavourakis, C.D.; Kleerebezem, R.; Damsté, J.S.S.; Muyzer, G.; Stams, A.J.M.; Sorokin, D.Y.; Plugge, C.M. Metabolism and occurrence of methanogenic and sulfate-reducing syntrophic acetate oxidizing communities in haloalkaline environments. Front. Microbiol. 2018, 9, 3039. [Google Scholar] [CrossRef]
- Parks, D.H.; Chuvochina, M.; Waite, D.W.; Rinke, C.; Skarshewski, A.; Chaumeil, P.A.; Hugenholtz, P. A standardized bacterial taxonomy based on genome phylogeny substantially revises the tree of life. Nat. Biotechnol. 2018, 36, 996–1004. [Google Scholar] [CrossRef]
- Sorokin, D.Y.; Merkel, A.Y. Dethiobacteria class. nov. In Bergey’s Manual of Systematics of Archaea and Bacteria; John Wiley & Sons: Hoboken, NJ, USA, 2022; pp. 1–2. [Google Scholar]
- Zavarzina, D.G.; Gavrilov, S.N.; Chistyakova, N.I.; Antonova, A.V.; Gracheva, M.A.; Merkel, A.Y.; Perevalova, A.A.; Chernov, M.S.; Zhilina, T.N.; Bychkov, A.Y.; et al. Syntrophic growth of alkaliphilic anaerobes controlled by ferric and ferrous minerals transformation coupled to acetogenesis. ISME J. 2020, 14, 425–436. [Google Scholar] [CrossRef] [PubMed]
- Zavarzina, D.G.; Kolganova, T.V.; Bulygina, E.S.; Kostrikina, N.A.; Turova, T.P.; Zavarzin, G.A. Geoalkalibacter ferrihydriticus gen. nov. sp. nov., the first alkaliphilic representative of the family Geobacteraceae, isolated from a soda lake. Microbiology 2006, 75, 673–682. [Google Scholar] [CrossRef]
- Wick, R.R.; Judd, L.M.; Gorrie, C.L.; Holt, K.E. Unicycler: Resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 2017, 13, e1005595. [Google Scholar] [CrossRef] [PubMed]
- Parks, D.H.; Imelfort, M.; Skennerton, C.T.; Hugenholtz, P.; Tyson, G.W. CheckM: Assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 2015, 25, 1043–1055. [Google Scholar] [CrossRef] [PubMed]
- Menzel, P.; Ng, K.L.; Krogh, A. Fast and sensitive taxonomic classification for metagenomics with Kaiju. Nat. Commun. 2016, 7, 11257. [Google Scholar] [CrossRef] [PubMed]
- Tatusova, T.; DiCuccio, M.; Badretdin, A.; Chetvernin, V.; Nawrocki, E.P.; Zaslavsky, L.; Lomsadze, A.; Pruitt, K.D.; Borodovsky, M.; Ostell, J. NCBI prokaryotic genome annotation pipeline. Nucleic Acids Res. 2016, 44, 6614–6624. [Google Scholar] [CrossRef]
- Couvin, D.; Bernheim, A.; Toffano-Nioche, C.; Touchon, M.; Michalik, J.; Néron, B.; Rocha, E.P.C.; Vergnaud, G.; Gautheret, D.; Pourcel, C. CRISPRCasFinder, an update of CRISRFinder, includes a portable version, enhanced performance and integrates search for Cas proteins. Nucleic Acids Res. 2018, 46, W246–W251. [Google Scholar] [CrossRef]
- Bertelli, C.; Laird, M.R.; Williams, K.P.; Simon Fraser University Research Computing Group; Lau, B.Y.; Hoad, G.; Winsor, G.L.; Brinkman, F.S.L. IslandViewer 4: Expanded prediction of genomic islands for larger-scale datasets. Nucleic Acids Res. 2017, 45, W30–W35. [Google Scholar] [CrossRef]
- Arndt, D.; Grant, J.R.; Marcu, A.; Sajed, T.; Pon, A.; Liang, Y.; Wishart, D.S. PHASTER: A better, faster version of the PHAST phage search tool. Nucleic Acids Res. 2016, 44, W16–W21. [Google Scholar] [CrossRef]
- Robert, X.; Gouet, P. Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 2014, 42, W320–W324. [Google Scholar] [CrossRef]
- Garber, A.I.; Nealson, K.H.; Okamoto, A.; McAllister, S.M.; Chan, C.S.; Barco, R.A.; Merino, N. FeGenie: A Comprehensive tool for the identification of iron genes and iron gene neighborhoods in genome and metagenome assemblies. Front. Microbiol. 2020, 11, 37. [Google Scholar] [CrossRef] [PubMed]
- Zavarzina, D.G.; Merkel, A.Y.; Klyukina, A.A.; Elizarov, I.M.; Pikhtereva, V.A.; Rusakov, V.S.; Chistyakova, N.I.; Ziganshin, R.H.; Maslov, A.A.; Gavrilov, S.N. Iron or sulfur respiration-an adaptive choice determining the fitness of a natronophilic bacterium Dethiobacter alkaliphilus in geochemically contrasting environments. Front. Microbiol. 2023, 14, 1108245. [Google Scholar] [CrossRef] [PubMed]
- Kevbrin, V.V.; Zavarzin, G.A. The effect of sulfur compounds on growth of halophilic homoacetic bacterium Acetohalobium arabaticum. Microbiology 1992, 61, 812–817. [Google Scholar]
- Osawa, T.; Inanaga, H.; Sato, C.; Numata, T. Crystal structure of the CRISPR-Cas RNA silencing Cmr complex bound to a target analog. Mol. Cell. 2015, 58, 418–430. [Google Scholar] [CrossRef] [PubMed]
- Varble, A.; Meaden, S.; Barrangou, R.; Westra, E.R.; Marraffini, L.A. Recombination between phages and CRISPR-cas loci facilitates horizontal gene transfer in staphylococci. Nat. Microbiol. 2019, 4, 956–963. [Google Scholar] [CrossRef] [PubMed]
- Nozhevnikova, A.N.; Russkova, Y.I.; Litti, Y.V.; Parshina, S.N.; Zhuravleva, E.A.; Nikitina, A.A. Syntrophy and interspecies electron transfer in methanogenic microbial communities. Microbiology 2020, 89, 129–147. [Google Scholar] [CrossRef]
- Manzoor, S.; Schnüre, A.; Bongcam-Rudloff, E.; Müller, B. Genome-guided analysis of Clostridium ultunense and comparative genomics reveal different strategies for acetate oxidation and energy conservation in syntrophic acetate-oxidising bacteria. Genes 2018, 9, 225. [Google Scholar] [CrossRef]
- Müller, B.; Sun, L.; Schnürer, A. First insights into the syntrophic acetate-oxidising bacteria—A genetic study. MicrobiologyOpen 2013, 2, 35–53. [Google Scholar] [CrossRef]
- Mock, J.; Wang, S.; Huang, H.; Kahnt, J.; Thauer, R.K. Evidence for a hexaheteromeric methylenetetrahydrofolate reductase in Moorella thermoacetica. J. Bacteriol. 2014, 196, 3303–3314. [Google Scholar] [CrossRef]
- Keller, A.; Schink, B.; Müller, N. Alternative pathways of acetogenic ethanol and methanol degradation in the thermophilic anaerobe Thermacetogenium phaeum. Front. Microbiol. 2019, 10, 423. [Google Scholar] [CrossRef]
- Wang, L.Y.; Nevin, K.P.; Woodard, T.L.; Mu, B.Z.; Lovley, D.R. Expanding the Diet for DIET: Electron Donors Supporting Direct Interspecies Electron Transfer (DIET) in Defined Co-Cultures. Front. Microbiol. 2016, 7, 236. [Google Scholar] [CrossRef] [PubMed]
- Lovley, D.R. Happy together: Microbial communities that hook up to swap electrons. ISME J. 2017, 11, 327–336. [Google Scholar] [CrossRef] [PubMed]
- Light, S.H.; Su, L.; Rivera-Lugo, R.; Cornejo, J.A.; Louie, A.; Iavarone, A.T.; Ajo-Franklin, C.M.; Portnoy, D.A. A flavin-based extracellular electron transfer mechanism in diverse Gram-positive bacteria. Nature 2018, 562, 140–144. [Google Scholar] [CrossRef]
- Melville, S.; Craig, L. Type IV pili in Gram-positive bacteria. Microbiol. Mol. Biol. Rev. 2013, 77, 323–341. [Google Scholar] [CrossRef]
- Gavrilov, S.N.; Zavarzina, D.G.; Elizarov, I.M.; Tikhonova, T.V.; Dergousova, N.I.; Popov, V.O.; Lloyd, J.R.; Knight, D.; El-Naggar, M.Y.; Pirbadian, S.; et al. Novel extracellular electron transfer channels in a gram-positive thermophilic bacterium. Front. Microbiol. 2021, 11, 597818. [Google Scholar] [CrossRef] [PubMed]
- Jeuken, L.J.C.; Hards, K.; Nakatani, Y. Extracellular Electron Transfer: Respiratory or Nutrient Homeostasis? J. Bacteriol. 2020, 202, e00029-20. [Google Scholar] [CrossRef]
- Holmes, D.E.; Dang, Y.; Walker, D.J.F.; Lovley, D.R. The electrically conductive pili of Geobacter species are a recently evolved feature for extracellular electron transfer. Microb. Genom. 2016, 2, e000072. [Google Scholar] [CrossRef] [PubMed]
- Morita, M.; Malvankar, N.S.; Franks, A.E.; Summers, Z.M.; Giloteaux, L.; Rotaru, A.E.; Rotaru, C.; Lovley, D.R. Potential for direct interspecies electron transfer in methanogenic wastewater digester aggregates. mBio 2011, 2, e00159-11. [Google Scholar] [CrossRef]
- Rotaru, A.-E.; Shrestha, P.M.; Liu, F.; Shrestha, M.; Shrestha, D.; Embree, M.; Zengler, K.; Wardman, C.; Nevin, K.P.; Lovley, D.R. A new model for electron flow during anaerobic digestion: Direct interspecies electron transfer to Methanosaeta for the reduction of carbon dioxide to methane. Energy Environ. Sci. 2014, 7, 408–415. [Google Scholar] [CrossRef]
- Holmes, D.E.; Shrestha, P.M.; Walker, D.J.F.; Dang, Y.; Nevin, K.P.; Woodard, T.L.; Lovley, D.R. Metatranscriptomic evidence for direct interspecies electron transfer between Geobacter and Methanothrix species in methanogenic rice paddy soils. Appl. Environ. Microbiol. 2017, 83, e00223-17. [Google Scholar] [CrossRef]
- Shrestha, P.M.; Rotaru, A.E.; Summers, Z.M.; Shrestha, M.; Liu, F.; Lovley, D.R. Transcriptomic and genetic analysis of direct interspecies electron transfer. Appl. Environ. Microbiol. 2013, 79, 2397–2404. [Google Scholar] [CrossRef] [PubMed]
- Rotaru, A.E.; Shrestha, P.M.; Liu, F.; Markovaite, B.; Chen, S.; Nevin, K.P.; Lovley, D.R. Direct interspecies electron transfer between Geobacter metallireducens and Methanosarcina barkeri. Appl. Environ. Microbiol. 2014, 80, 4599–4605. [Google Scholar] [CrossRef] [PubMed]
- Ueki, T.; Nevin, K.P.; Rotaru, A.E.; Wang, L.Y.; Ward, J.E.; Woodard, T.L.; Lovley, D.R. Geobacter strains expressing poorly conductive pili reveal constraints on direct interspecies electron transfer mechanisms. mBio 2018, 9, e01273-18. [Google Scholar] [CrossRef] [PubMed]
- Summers, Z.M.; Fogarty, H.E.; Leang, C.; Franks, A.E.; Malvankar, N.S.; Lovley, D.R. Direct exchange of electrons within aggregates of an evolved syntrophic coculture of anaerobic bacteria. Science 2010, 30, 1413–1415. [Google Scholar] [CrossRef]
- Kato, S.; Hashimoto, K.; Watanabe, K. Iron-oxide minerals affect extracellular electron-transfer paths of Geobacter spp. Microbes Environ. 2013, 28, 141–148. [Google Scholar] [CrossRef]
- Cheng, J.; Li, H.; Ding, L.; Zhou, J.; Song, W.; Li, Y.-Y.; Lin, R. Improving hydrogen and methane co-generation in cascading dark fermentation and anaerobic digestion: The effect of magnetite nanoparticles on microbial electron transfer and syntrophism. Chem. Eng. J. 2020, 397, 125394. [Google Scholar] [CrossRef]
- Chen, S.; Rotaru, A.E.; Liu, F.; Philips, J.; Woodard, T.L.; Nevin, K.P.; Lovley, D.R. Carbon cloth stimulates direct interspecies electron transfer in syntrophic co-cultures. Bioresour. Technol. 2014, 173, 82–86. [Google Scholar] [CrossRef]
- Chen, L.; Fang, W.; Chang, J.; Liang, J.; Zhang, P.; Zhang, G. Improvement of direct interspecies electron transfer via adding conductive materials in anaerobic digestion: Mechanisms, performances, and challenges. Front. Microbiol. 2022, 13, 860749. [Google Scholar] [CrossRef]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Frolov, E.N.; Gavrilov, S.N.; Toshchakov, S.V.; Zavarzina, D.G. Genomic Insights into Syntrophic Lifestyle of ‘Candidatus Contubernalis alkaliaceticus’ Based on the Reversed Wood–Ljungdahl Pathway and Mechanism of Direct Electron Transfer. Life 2023, 13, 2084. https://doi.org/10.3390/life13102084
Frolov EN, Gavrilov SN, Toshchakov SV, Zavarzina DG. Genomic Insights into Syntrophic Lifestyle of ‘Candidatus Contubernalis alkaliaceticus’ Based on the Reversed Wood–Ljungdahl Pathway and Mechanism of Direct Electron Transfer. Life. 2023; 13(10):2084. https://doi.org/10.3390/life13102084
Chicago/Turabian StyleFrolov, Evgenii N., Sergey N. Gavrilov, Stepan V. Toshchakov, and Daria G. Zavarzina. 2023. "Genomic Insights into Syntrophic Lifestyle of ‘Candidatus Contubernalis alkaliaceticus’ Based on the Reversed Wood–Ljungdahl Pathway and Mechanism of Direct Electron Transfer" Life 13, no. 10: 2084. https://doi.org/10.3390/life13102084