

Physiological and Anatomical Responses of Faba Bean Plants Infected with Chocolate Spot Disease to Chemical Inducers
 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Studies on the Causal Pathogens
2.1.1. Isolation of Chocolate Spot Pathogens
2.1.2. Identification of Isolated Fungi
2.1.3. Pathogenicity Test
Inoculum Preparation
Plant Preparation
Pathogenicity Assessment and Development of Choloate Spot Disease
Determination of Chocolate Spot Disease Severity
2.2. Biochemical Analysis
2.2.1. Peroxidase (POX) Assay
2.2.2. Polyphenol Oxidase (PPO) Assay
2.2.3. B-1, 3 Glucanase Assay
2.2.4. Chitinase Assay
2.2.5. Protein Profile
2.3. Anatomical Studies
2.4. Statistical Analysis
3. Results
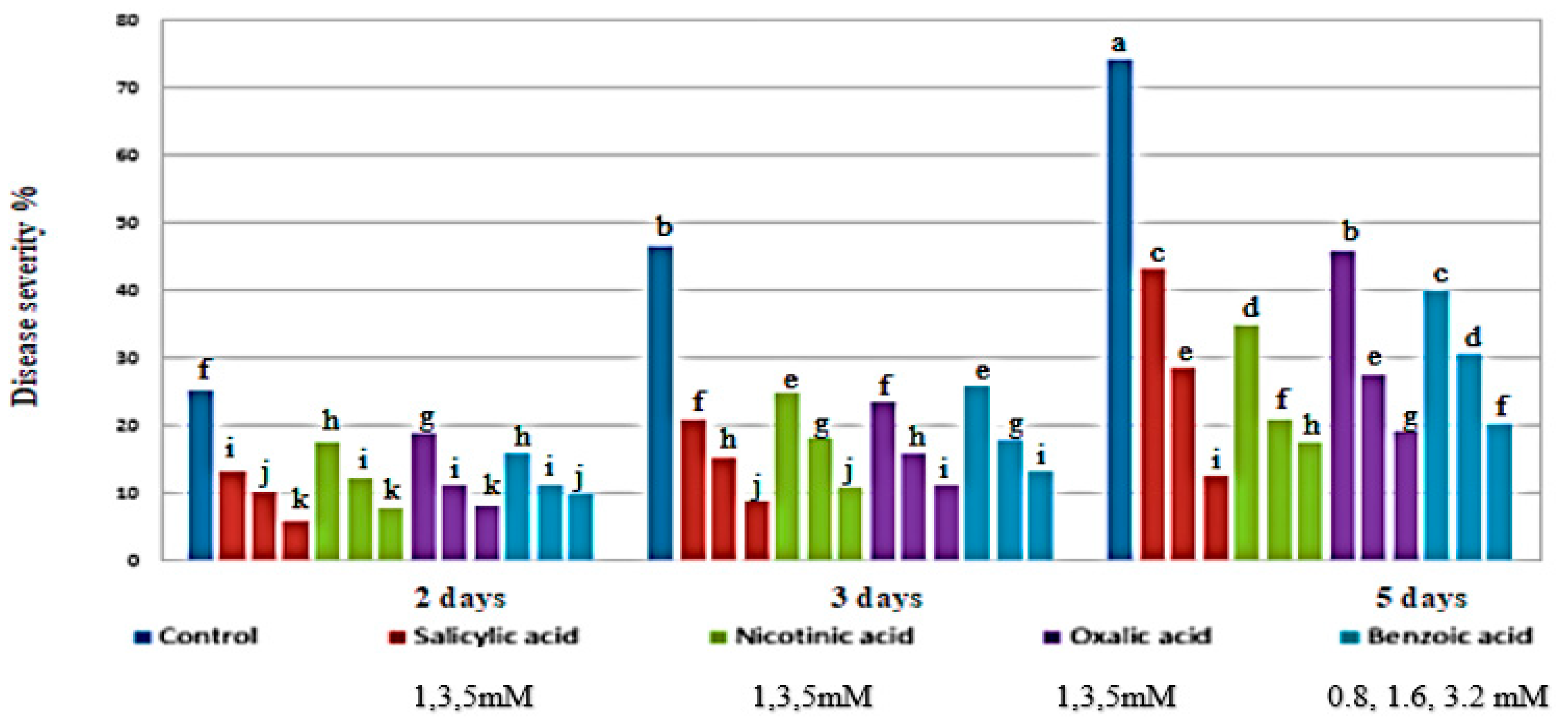
3.1. Severity of Chocolate Spot Disease
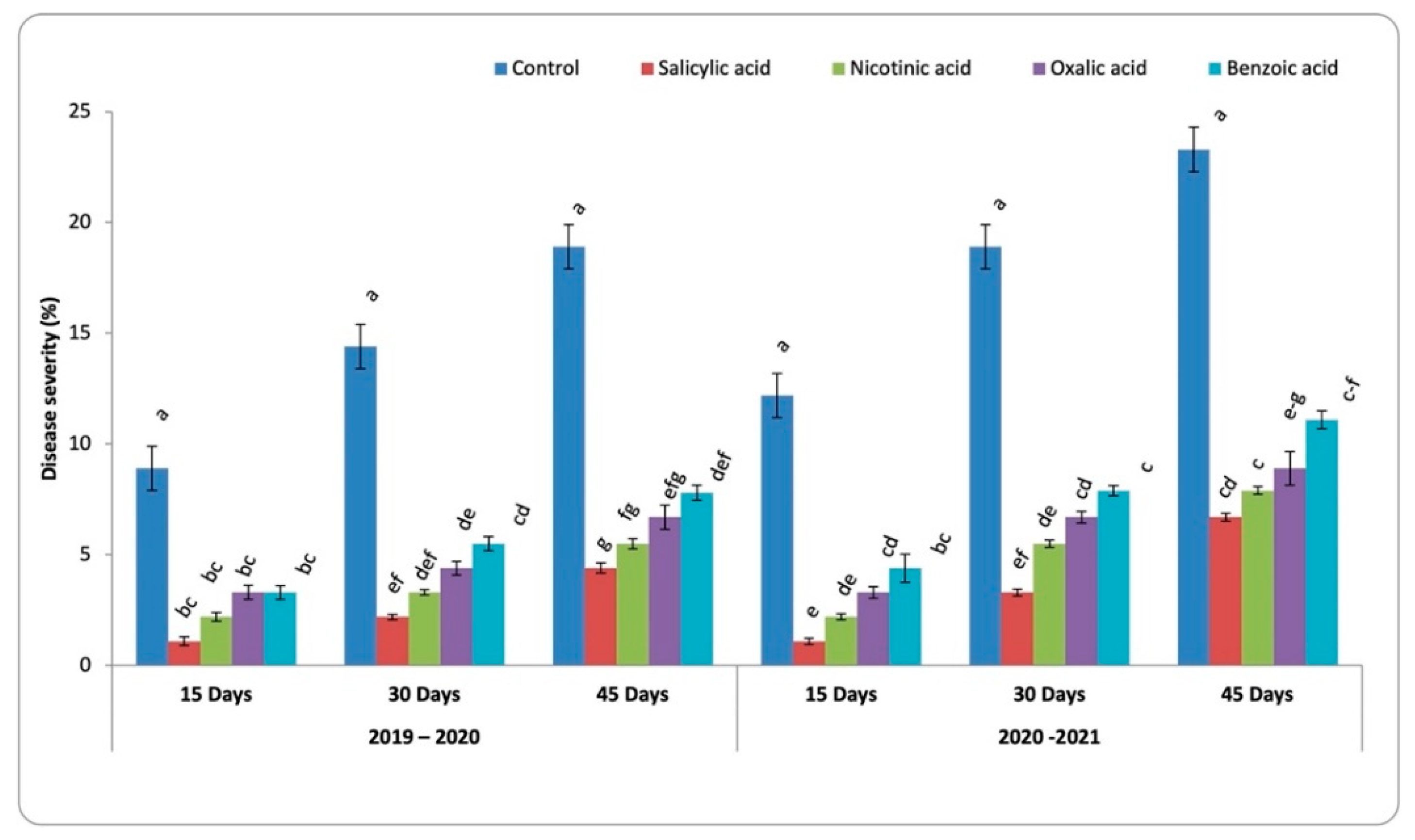
3.2. Development of Chocolate Spot Disease
3.3. Antioxidant Enzymes Activity
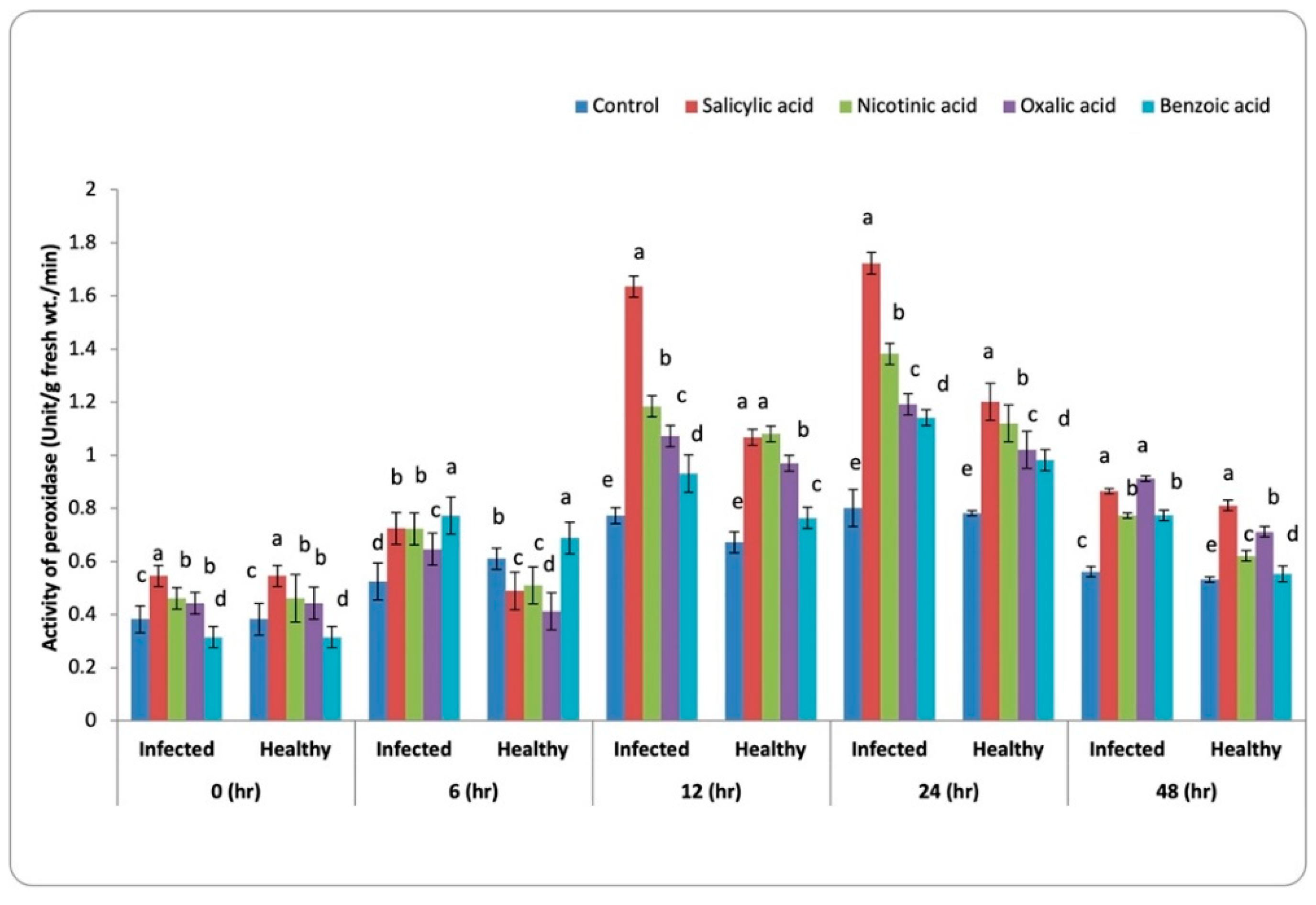
3.3.1. Peroxidase Activity (POX)
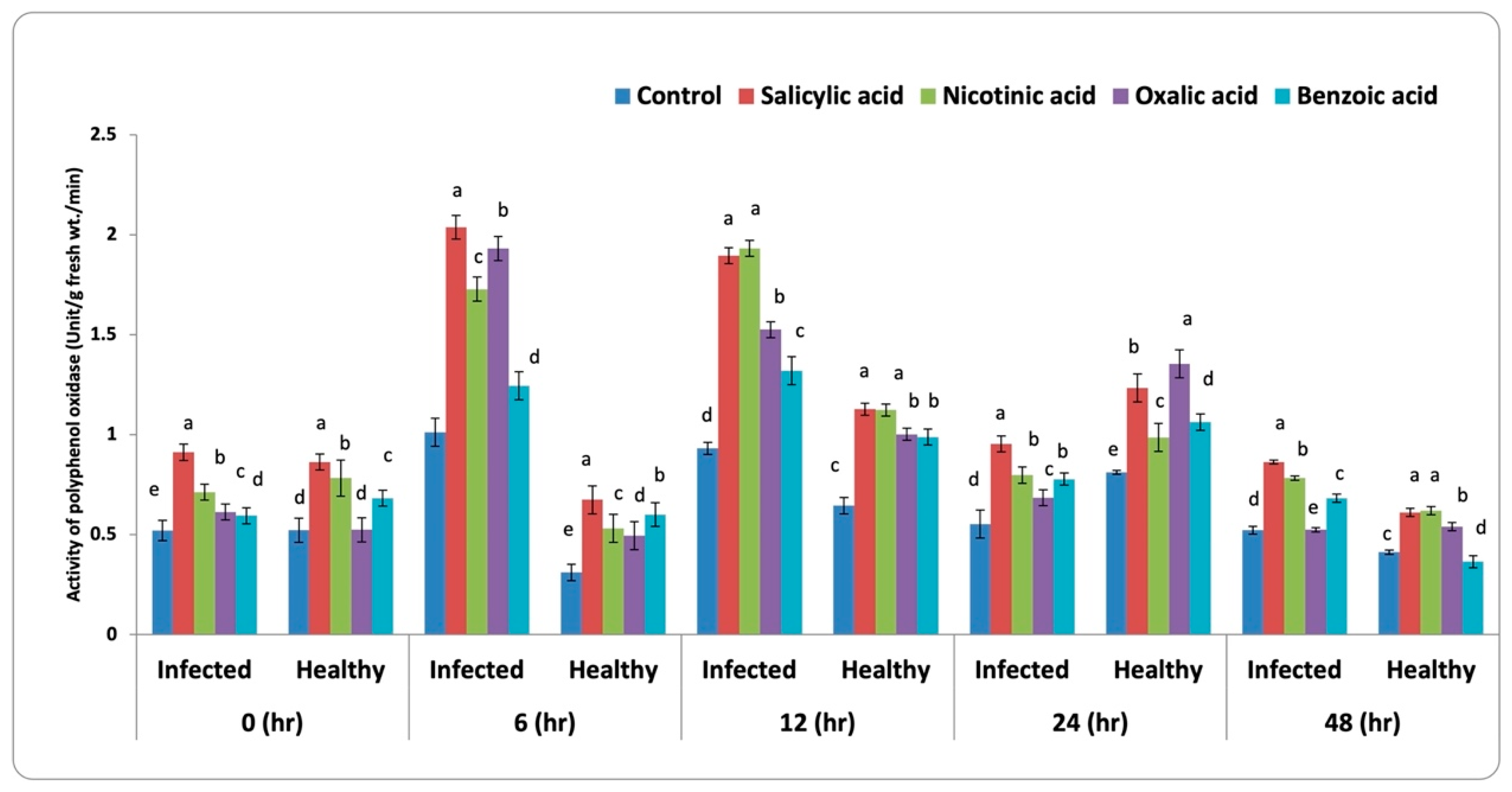
3.3.2. Polyphenol Oxidase (PPO) Activity
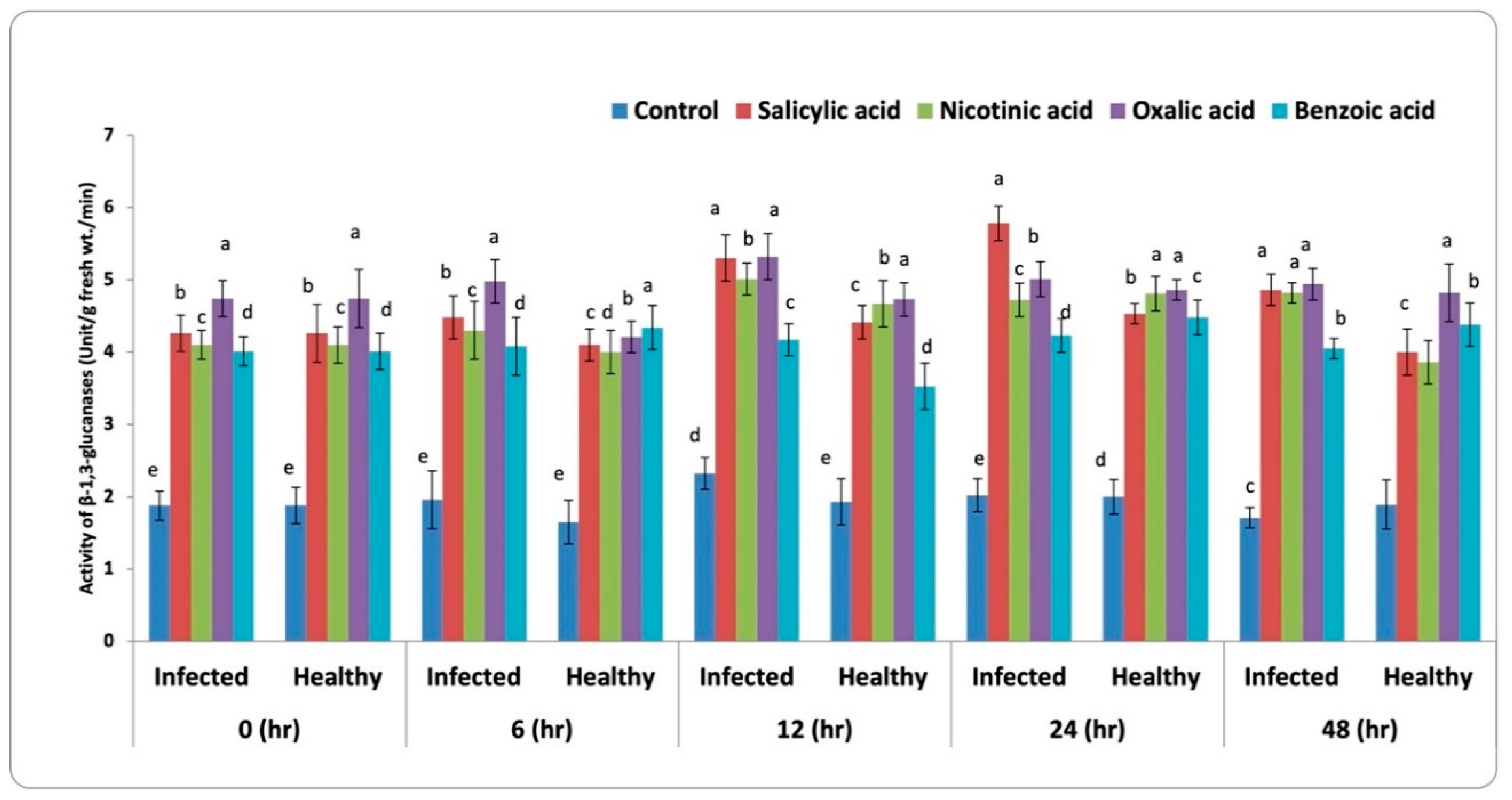
3.3.3. β-1,3-Glucanase Activity
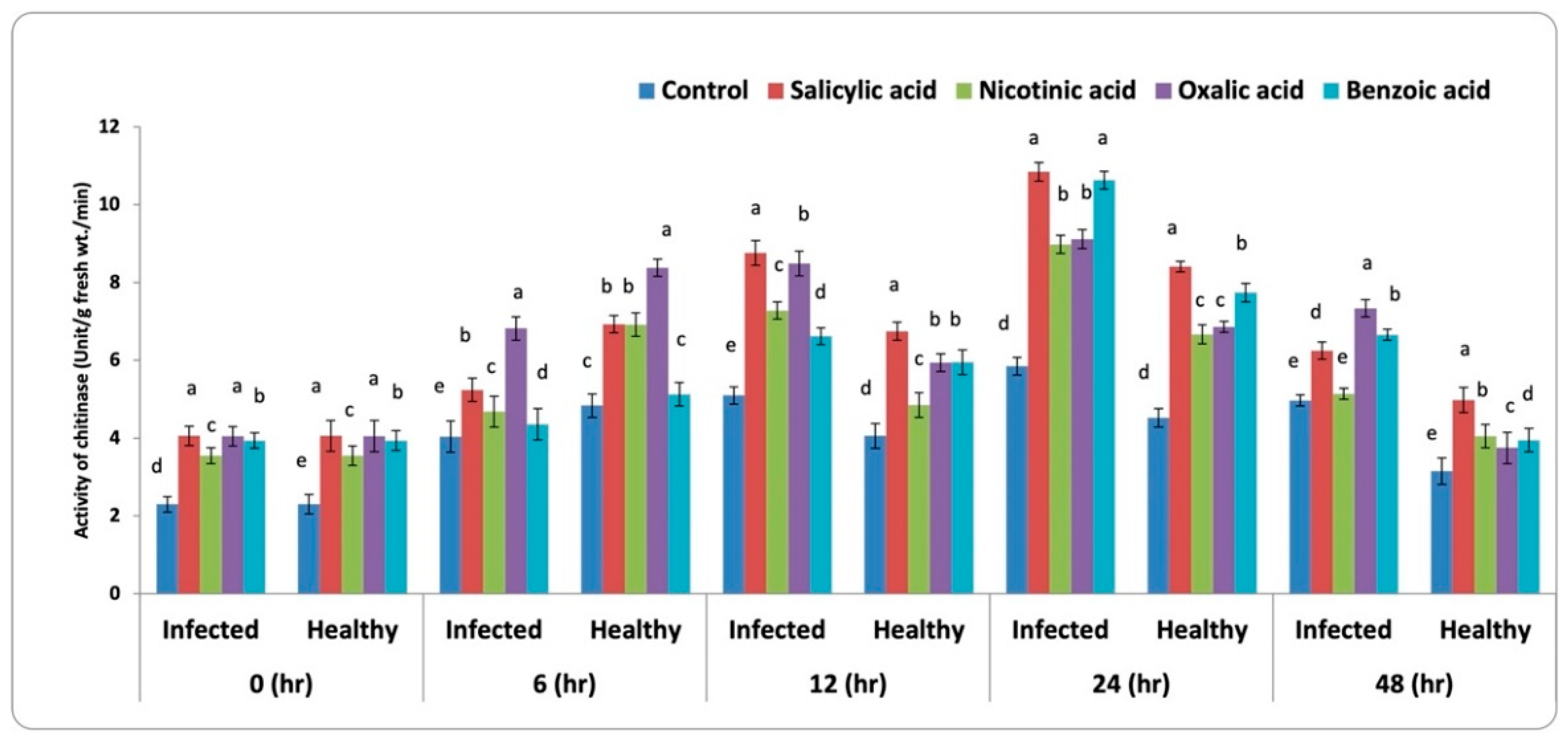
3.3.4. Chitinase Activity
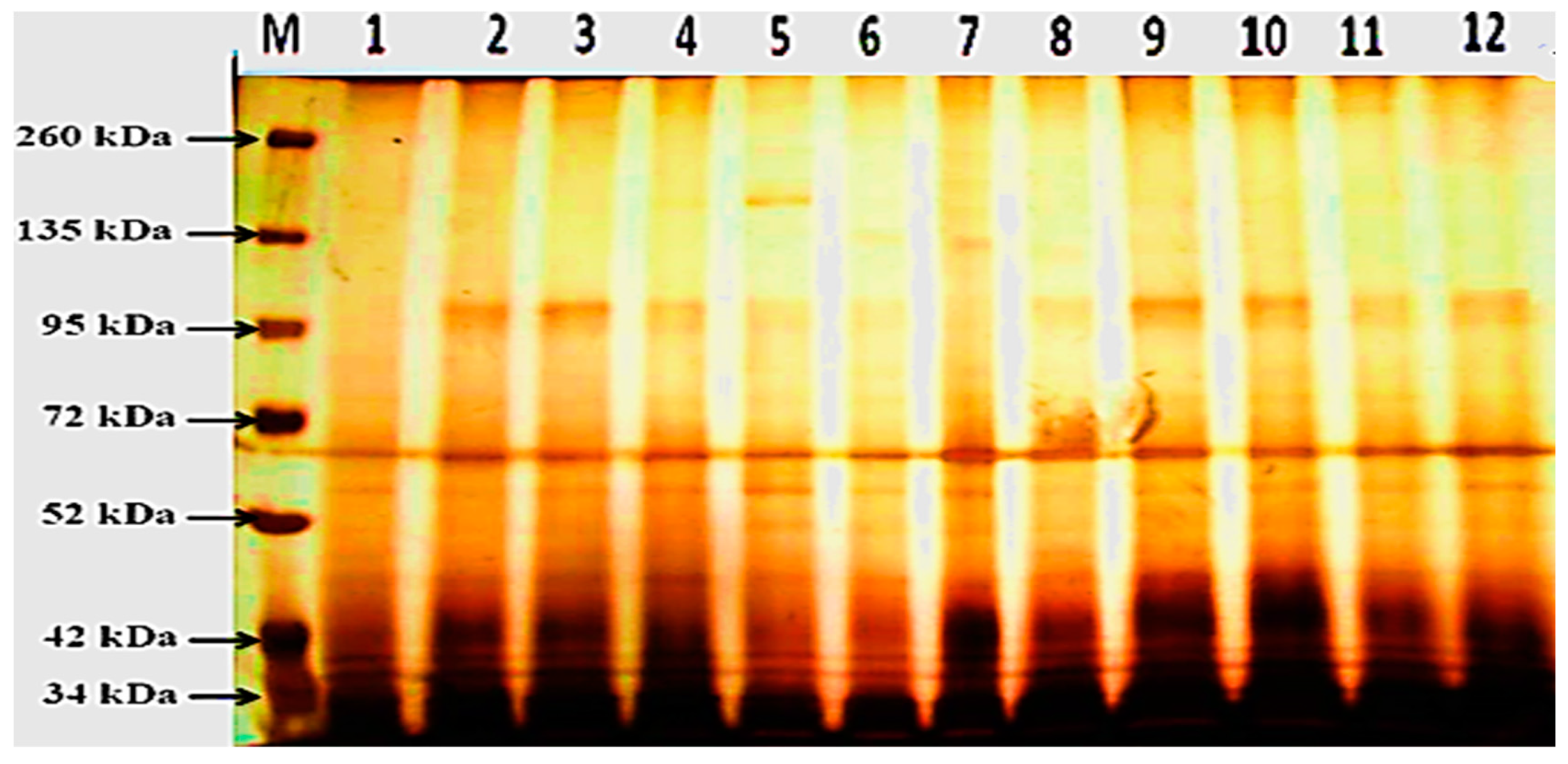
3.4. Protein Electrophoretic Banding Patterns
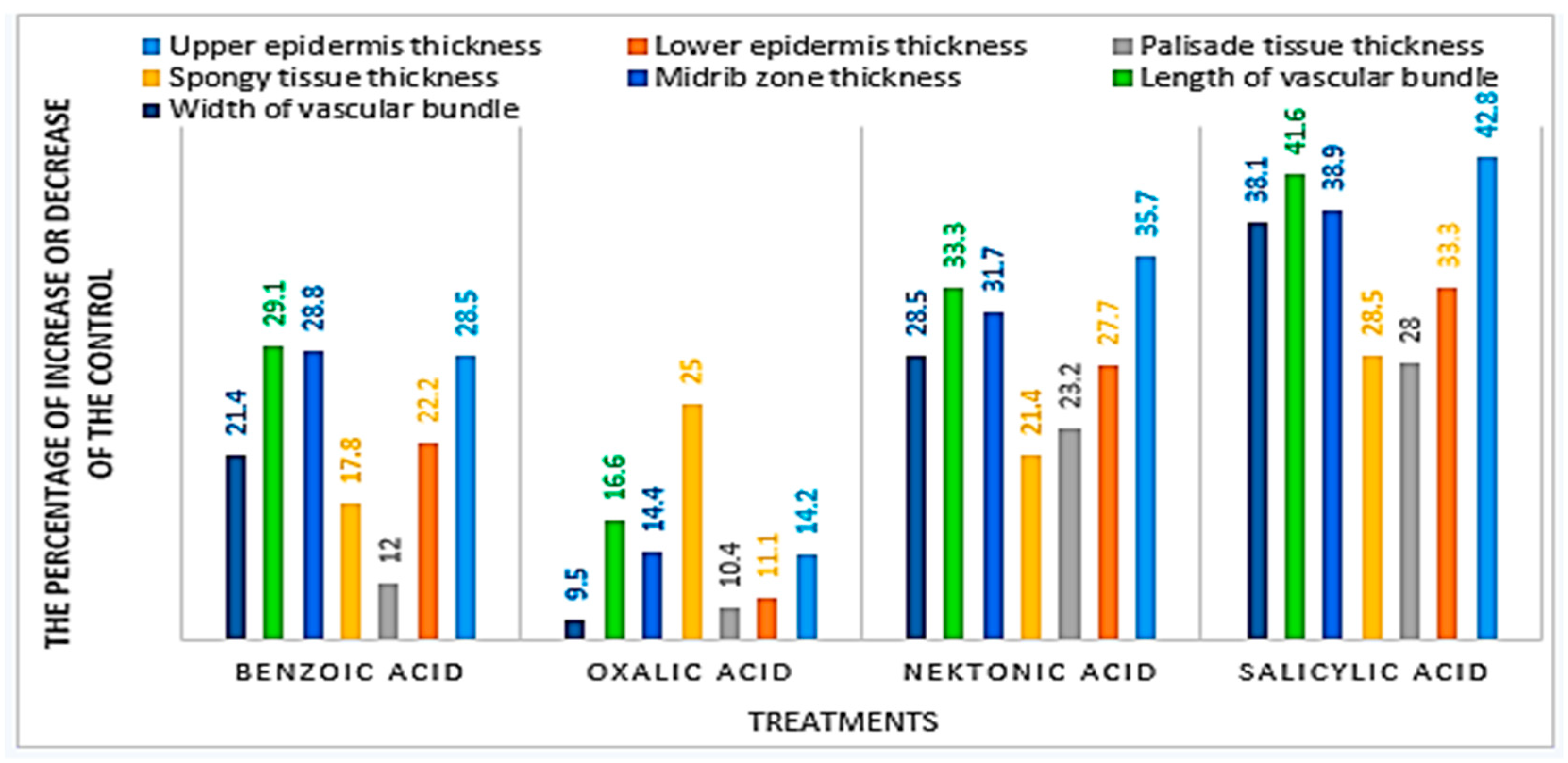
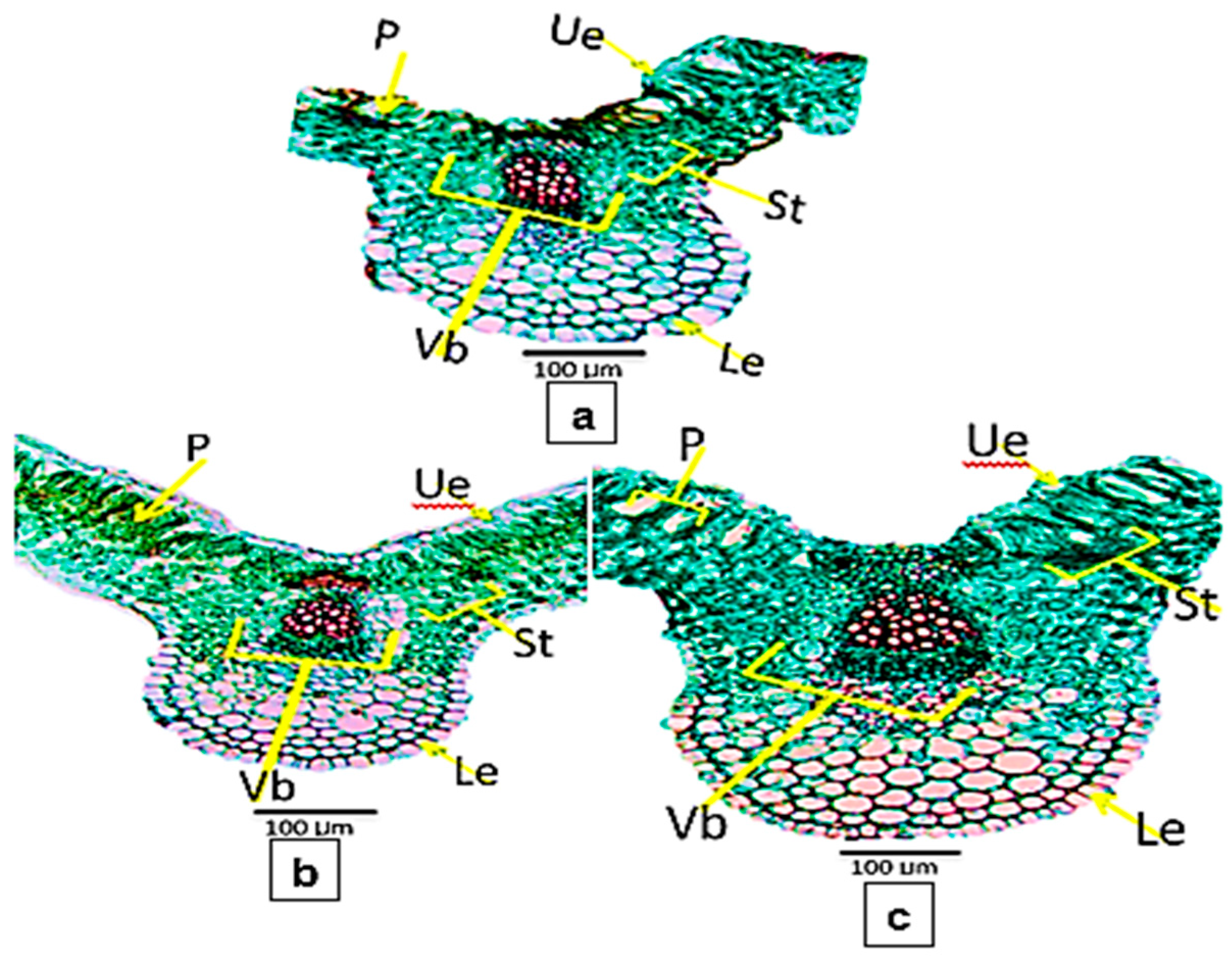
3.5. Anatomical Characteristics
4. Discussion
4.1. Severity of Chocolate Spot Disease
4.2. Antioxidant Enzymes Activity
4.3. Changes in Protein Electrophoretic Patterns
4.4. Anatomical Characteristics
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Crépon, K.; Marget, P.; Peyronnet, C.; Carrouée, B.; Arese, P.; Duc, G. Nutritional value of faba bean (Vicia faba L.) seeds for feed and food. Field Crop. Res. 2010, 115, 329–339. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Hashem, H.A.; Khalil, R.R. Stigmasterol treatment increases salt stress tolerance of faba bean plants by enhancing antioxidant systems. Plant Omics J. 2012, 5, 476–485. [Google Scholar]
- Hungria, M.; Vargas, M.A. Environmental factors affecting nitrogen fixation in grain legumes in the tropics, with an emphasis on Brazil. Field Crop. Res. 2000, 65, 151–164. [Google Scholar] [CrossRef]
- Rhaïem, Z.; Cherif, M.; Kharrat, M.; Mejda Cherif, M.; Moncef Harrabi, M. New faba bean genotypes resistant to chocolate spot caused by Botrytis fabae. Phytopathol. Mediterr. 2002, 41, 99–108. [Google Scholar]
- El-Banoby, F.E.; Abd-AllA, M.A.; Tolba, I.H.; Morsy, A.A.; El-Gamal, N.G.; Khalil, M.S.A. Biological control of chocolate spot disease of faba bean using some bioagents under field conditions. J. Appl. Sci. Res. 2013, 9, 4021–4029. [Google Scholar]
- Mbazia, A.; Ben Youssef, N.O.; Kharrat, M. Effect of some chemical inducers on chocolate spot disease of faba bean in Tunisia. J. Crop. Prot. 2016, 5, 541–552. [Google Scholar] [CrossRef] [Green Version]
- Aldesuquy, H.; Baka, Z.; Alazab, N. Shikimic and salicylic acids induced resistance in faba bean plants against chocolate spot disease. J. Plant Pathol. Microbiol. 2015, 6, 257. [Google Scholar] [CrossRef] [Green Version]
- Latef, A.A.H.A.; Chaoxing, H. Does Inoculation with Glomus mosseae improve salt tolerance in pepper plants? J. Plant Growth Regul. 2014, 33, 644–653. [Google Scholar] [CrossRef]
- Mittova, V.; Tal, M.; Volokita, M.; Guy, M. Up-regulation of the leaf mitochondrial and peroxisomal antioxidative systems in response to salt-induced oxidative stress in the wild salt-tolerant tomato species Lycopersicon pennellii. Plant Cell Environ. 2003, 26, 845–856. [Google Scholar] [CrossRef]
- Lu, T.; Finkel, T. Free radicals and senescence. Exp. Cell Res. 2008, 314, 1918–1922. [Google Scholar] [CrossRef] [PubMed]
- Makoi, J.H.J.R.; Ndakidemi, P.A. Biological, ecological and agronomic significance of plant phenolic compounds in rhizosphere of the symbiotic legumes. Afr. J. Biotechnol. 2007, 6, 1358–1368. [Google Scholar]
- Abd El-Gawad, H.G.; Bondok, A.M. Response of tomato plants to salicylic acid and chitosan under infection with tomato mosaic virus. Am.-Eurasian J. Agric. Environ. Sci. 2015, 15, 1520–1529. [Google Scholar] [CrossRef]
- Jayakannan, M.; Bose, J.; Babourina, O.; Rengel, Z.; Shabala, S. Salicylic acid in plant salinity stress signalling and tolerance. Plant Growth Regul. 2015, 76, 25–40. [Google Scholar] [CrossRef]
- Aldesuquy, H.S.; Baka, Z.A.; Abass, M.A.; Nahla Alazab, T. Faba bean can adapt to chocolate spot disease by pretreatment with shikimic and salicylic acids through osmotic adjustment, solutes allocation and leaf turgidity. J. Stress Physiol. Biochem. 2014, 10, 230–243. [Google Scholar]
- El-Sayed, S.A. Efficiency of some antioxidants, biocontrol agents and fungicides in induction of faba bean resistance to chocolate spot disease. SF J. Mycol. 2017, 1, 10–13. [Google Scholar]
- Cárcamo, H.; Herle, C.; Hervet, V. Greenhouse studies of thiamethoxam effects on pea leaf weevil, Sitona lineatus. J. Insect Sci. 2012, 12, 151. [Google Scholar] [CrossRef] [PubMed]
- Nour, K.A.M.; Mansour, N.T.S.; Eisa, G.S.A. Effect of Some Antioxidants on Some Physiological and Anatomical Characters of Snap Bean Plants. N. Y. Sci. J. 2012, 5. Available online: https://www.sciencepub.net/newyork (accessed on 12 October 2022).
- Gomaa, E.F.; Nassar, R.M.A.; Madkour, M.A. Effect of foliar spray with salicylic acid on vegetative growth, stem and leaf anatomy, photosynthetic pigments and productivity of Egyptian lupine plant (Lupinus termis forssk.). Int. J. Adv. Res. 2015, 3, 803–813. [Google Scholar]
- Abdallah, M.M.; El-Bassiouny, H.M.S.; Elewa, T.A.; El-Sebai, T.N. Effect of salicylic acid and benzoic acid on growth, yield and some biochemical aspects of quinoa plant grown in sandy soil. Int. J. Chem. Tech. Res. 2015, 8, 216–225. [Google Scholar]
- Dawood, M.G.; Sadak, M.S.; Bakry, B.A.; El Karamany, M.F. Comparative studies on the role of benzoic, t-cinnamic, and salicylic acids on growth, some biochemical aspects, and yield of three flax cultivars grown under sandy soil conditions. Bull. Natl. Res. Cent. 2019, 43, 112. [Google Scholar] [CrossRef] [Green Version]
- Senaratna, T.; Merritt, D.; Dixon, K.; Bunn, E.; Touchell, D.; Sivasithamparam, K. Benzoic acid may act as the functional group in salicylic acid and derivatives in the induction of multiple stress tolerance in plants. Plant Growth Regul. 2003, 39, 77–81. [Google Scholar] [CrossRef]
- Cessna, S.G.; Sears, V.E.; Dickman, M.B.; Low, P.S. Oxalic acid, a pathogenicity factor for Sclerotinia sclerotiorum, suppresses the oxidative burst of the host plant. Plant Cell 2000, 12, 2191–2199. [Google Scholar] [CrossRef] [Green Version]
- Godoy, G.; Steadman, J.; Dickman, M.; Dam, R. Use of mutants to demonstrate the role of oxalic acid in pathogenicity of Sclerotinia sclerotiorum on Phaseolus vulgaris. Physiol. Mol. Plant Pathol. 1990, 37, 179–191. [Google Scholar] [CrossRef]
- Fagundes-Nacarath, I.; Debona, D.; Rodrigues, F. Oxalic acid-mediated biochemical and physiological changes in the common bean-Sclerotinia sclerotiorum interaction. Plant Physiol. Biochem. 2018, 129, 109–121. [Google Scholar] [CrossRef]
- Williams, B.; Kabbage, M.; Kim, H.J.; Britt, R.; Dickman, M.B. Tipping the balance: Sclerotinia sclerotiorum secreted oxalic acid suppresses host defenses by manipulating the host redox environment. PLoS Pathog. 2011, 7, e1002107. [Google Scholar] [CrossRef] [Green Version]
- Malenčić, D.; Kiprovski, B.; Popović, M.; Prvulović, D.; Miladinović, J.; Djordjević, V. Changes in antioxidant systems in soybean as affected by Sclerotinia sclerotiorum (Lib.) de Bary. Plant Physiol. Biochem. 2010, 48, 903–908. [Google Scholar] [CrossRef]
- Sadak, M.S.; Rady, M.; Badr, N.M.; Gaballah, M.S. Increasing sunflower salt tolerance using nicotinamide and α-tocopherol. Int. J. Acad. Res. 2010, 2, 263–270. [Google Scholar]
- Bearder, I.R. Plant hormones and other background, structure and occurrence, 9–112. In Hormonal Regulation and Development; Encyclopedia of Plant, Physiology; Millan, J.M., Ed.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 1980. [Google Scholar]
- Berglund, T. Nicotinamide, a missing link in the early stress response in eukaryotic cells: A hypothesis with special reference to oxidative stress in plants. FEPS Lett. 1994, 351, 375–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhu, X.; Pan, L.; Xiao, T.; Ren, X.; Liu, Z. Exogenous niacin treatment increases NADPH oxidase in kiwifruit. Braz. J. Biol. 2018, 78, 686–690. [Google Scholar] [CrossRef] [Green Version]
- Sahile, S.; Ahmed, S.; Fininsa, C.; Abang, M.M.; Sakhuja, P.K. Survey of chocolate spot (Botrytis fabae) disease of faba bean (Vicia faba L.) and assessment of factors influencing disease epidemics in northern Ethiopia. Crop. Prot. 2008, 27, 1457–1463. [Google Scholar] [CrossRef]
- Shifa, H.; Temam, H.; Sakhuja, P.K. Association of faba bean rust (Uromyces viciae-fabae) with environmental factors and cultural practices in the Hararghe Highlands, Eastern Ethiopia. East Afr. J. Sci. 2011, 5, 55–58. [Google Scholar]
- Riker, A.J.; Riker, R.S. Introduction to Research on Plant Diseases; John S. Swift Co., Inc.: New York, NY, USA, 1936; p. 117. [Google Scholar]
- Moussa, H.E.; El-Shatoury, S.A.; Abdul, O.A.; Dewedar, W.; Dewedar, A. Characterization of endophytic actinomycetes from wild Egyptian plants as antagonists to some phytopathogenic fungi. Egypt. J. Nat. Toxins 2011, 8, 32–48. [Google Scholar]
- Last, F.T.; Hamley, R.E. A local-lesion technique for measuring the infectivity of conidia of Botrytis fabae Sard. Ann. Appl. Biol. 1956, 44, 410–418. [Google Scholar] [CrossRef]
- Bernier, C.C.; Hanounik, S.B.; Hussein, M.M.; Mohamed, H.A. Field Manual of Common Faba Bean Diseases in the Nile Valley; International Center for Agricultural Research in the Dry Areas (ICARDA): Aleppo, Syria, 1993; Volume 3. [Google Scholar]
- Chen, J.X.; Wang, X.F. Plant Physiology Experimental Guide; Higher Education Press: Beijing, China, 2006; pp. 55–56. [Google Scholar] [CrossRef] [Green Version]
- Kumar, K.B.; Khan, P.A. Peroxidase and polyphenol oxidase in excised ragi (Eleusine coracana cv. PR 202) leaves during senescence. Indian J. Exp. Bot. 1982, 20, 412–416. [Google Scholar]
- Atrooz, O.M. Some Properties of the polyphenol oxidase from Cyclamen persicum. Egypt. J. Agric. Sci. 2009, 60, 408–414. [Google Scholar] [CrossRef]
- Abeles, F.B.; Forrence, L.E. Temporal and hormonal control of β-1,3-glucanase in Phaseolus vulgaris L. Plant Physiol. 1970, 45, 395–400. [Google Scholar] [CrossRef] [Green Version]
- Reid, J.D.; Ogrydziak, D.M. Chitinase-overproducing mutant of Serratia marcescens. Appl. Environ. Microbiol. 1981, 41, 664–669. [Google Scholar] [CrossRef] [Green Version]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef]
- Sammons, D.W.; Adams, L.D.; Nishizawa, E.E. Ultrasensitive silver-based color staining of polypeptides in polyacrylamide gels. Electrophoresis 1981, 2, 135–141. [Google Scholar] [CrossRef]
- Nassar, M.A.; El-Sahhar, K.F. Botanical Preparations and Microscopy (microtechnique); Academic Bookshop: Giza, Egypt, 1998; Volume 219, pp. 35–40. [Google Scholar] [CrossRef] [Green Version]
- Snedecor, G.W.; Cochran, W.G. Statistical Methods, 8th ed.; Iowa State Univ., Press: Ames, IA, USA, 1990. [Google Scholar] [CrossRef]
- Firdu, Z.; Maia, L.; Teodoro, J.; Alemu, T.; Assefa, F. Characterization of faba bean (Vicia faba L.) rhizosphere associating rhi-zobacteria against Botrytis fabae AAUBF-12 and their plant growth-promoting properties. Heliyon 2022, 8, e08861. [Google Scholar] [CrossRef]
- Metwaly, H. Control of chocolate spot disease by non traditional methods on faba bean plants. Egypt. J. Phytopathol. 2014, 42, 143–158. [Google Scholar] [CrossRef]
- Thakur, M.; Sohal, B.S. Role of elicitors in inducing resistance in plants against pathogen infection: A Review. ISRN Biochem. 2013, 2013, 762412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zian, A.H.; El-Gendy, H.M.R.; Shehata, H.S. Enhancing biocontrol agents by hydroquinone and salicylic acid for controlling root-rot and wilt diseases of lupine Egypt. J. Phytopathol. 2019, 47, 97–120. [Google Scholar] [CrossRef] [Green Version]
- El-Rahman, A.; Saieda, S.; Mohamed, H.I. Application of benzothiadiazole and Trichoderma harzianum to control faba bean chocolate spot disease and their effect on some physiological and biochemical traits. Acta Physiol. Plantarum. 2014, 36, 343–354. [Google Scholar] [CrossRef]
- Gholami, M.; Rahemi, M.; Kholdebarin, B. Effect of drought stress induced by polyethylene glycol on seed germination of four wild almond species. Aust. J. Basic Appl. Sci. 2010, 4, 785–791. [Google Scholar]
- Dat, J.F.; Lopez-Delgado, H.; Foyer, C.H.; Scott, I.M. Effects of salicylic acid on oxidative stress and thermotolerance in tobacco. J. Plant Physiol. 2000, 156, 659–665. [Google Scholar] [CrossRef]
- Sairam, R.; Srivastava, G. Changes in antioxidant activity in sub-cellular fractions of tolerant and susceptible wheat genotypes in response to long term salt stress. Plant Sci. 2002, 162, 897–904. [Google Scholar] [CrossRef]
- Haddoudi, I.; Cabrefiga, J.; Mora, I.; Mhadhbi, H.; Montesinos, E.; Mrabet, M. Biological control of Fusarium wilt caused by Fusarium equiseti in Vicia faba with broad spectrum antifungal plant-associated Bacillus spp. Biol. Control. 2021, 160, 104671. [Google Scholar] [CrossRef]
- Sharma, P.; Dubey, R.S. Involvement of oxidative stress and role of antioxidative defense system in growing rice seedlings exposed to toxic concentrations of aluminum. Plant Cell Rep. 2007, 26, 2027–2038. [Google Scholar] [CrossRef]
- Mohammadi, M.; Karr, A.L. β-1,3-glucanase and chitinase activities in soybean root nodules. J. Plant Physiol. 2002, 159, 245–256. [Google Scholar] [CrossRef]
- Manochehrifar, P. Effect of salisylic acid in plants. In Proceedings of the 1st symposium of New Findings in Chemistry and Engineering Chemistry, Iran Corpus. Kyoto, Japan, 1–3 August 2010; p. 43790350. [Google Scholar]
- Zhang, E.P.; Zhang, S.H.; Zhang, W.B.; Li, L.L. Effects of exogenic benzoic acid and cinnamic acid on the root oxidative dam-age of tomato seedlings. J. Hortic. For. 2010, 2, 22–29. [Google Scholar] [CrossRef]
- Amist, N.; Singh, N.B. Comparative effects of benzoic acid and water stress on wheat seedlings. Russ. J. Plant Physiol. 2018, 65, 709–716. [Google Scholar] [CrossRef]
- Hassanein, R.A.; Bassuony, F.M.; Baraka, D.M.; Khalil, R.R. Physiological effects of nicotinamide and ascorbic acid on Zea mays plant grown under salinity stress I-changes in growth, some relevant metabolic activities and oxidative defense systems. Res. J. Agric. Biol. Sci. 2009, 5, 72–81. [Google Scholar]
- Muller, H.P.; Gottschelk, W. Quantitative and qualitative situation of Pisum sativum. In Nuclear Techniques for Seed Protein Improvement; I.A.E.A.: Vienna, Austria, 1973; pp. 235–253. [Google Scholar]
- Spreitzer, R.J.; Salvucci, M.E. Rubisco: Structure, regulatory interactions and possibilities for a better enzyme. Annu. Rew. Plant Biol. 2002, 53, 449–475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whitney, S.M.; Houtz, R.L.; Alonso, H. Advancing our understanding and capacity to engineer nature’s CO2-sequestering enzyme, Rubisco. Plant Physiol. 2011, 155, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Bassiouny, H.M.S.; Sadak, M.S.S. Impact of foliar application of ascorbic acid and α-tocopherol on antioxidant activity and some biochemical aspects of flax cultivars under salinity stress. Acta Biol. Colomb. 2015, 20, 209–222. [Google Scholar] [CrossRef]
- Tegeder, M. Transporters involved in source to sink partitioning of amino acids and ureides: Opportunities for crop improvement. J. Exp. Bot. 2014, 65, 1865–1878. [Google Scholar] [CrossRef]
- Hildebrandt, T.M.; Nesi, A.N.; Araújo, W.L.; Braun, H.-P. Amino acid catabolism in plants. Mol. Plant 2015, 8, 1563–1579. [Google Scholar] [CrossRef] [Green Version]
- Ramadan, A.A.; Younis, A.S.M.; Bakry, B.A.; El-Bassiouny, H.M.S. Biochemical and yield of flax in responses to some natural antioxidant substances under sandy soil conditions. Asian J. Plant Sci. 2020, 19, 261–272. [Google Scholar] [CrossRef]
- Mahgoob, M.; Talaat, E. Physiological response of rose geranium (Pelargonium gravelones L.) to phenylalanine and nicotinic acid. Ann. Agric. Sci. Moshtohor 2005, 43, 807–822. [Google Scholar]
- Goñi, O.; Sanchez-Ballesta, M.T.; Merodio, C.; Escribano, M.I. Potent cryoprotective activity of cold and CO2-regulated cherimoya (Annona cherimola) endochitinase. J. Plant Physiol. 2010, 167, 1119–1129. [Google Scholar] [CrossRef]
- Shukry, W.M. Effect of soil type on growth vigour, water relations, mineral uptake and contents of fatty acids and protein of yielded seeds of Linum usitatissimum. Pak. J. Biol. Sci. 2001, 4, 1470–1478. [Google Scholar] [CrossRef] [Green Version]
- Ali, Z.A.; Hussein, M.M.; El-Taher, A.M. Effect of antioxidants on some morphological and anatomical features of maize grown under salinity conditions. Int. J. Chem. Tech. Res. 2015, 8, 389–400. [Google Scholar]
- Khalil, M.; Ramadan, A.; El-Sayed, S.; El-Taher, A. Effectiveness of natural antioxidants on physiological, anatomical changes and controlling downy, powdery mildew and rust diseases in pea plants. Asian J. Plant Sci. 2023, 22, 25–36. [Google Scholar] [CrossRef]
- Maddah, S.M.; Falahian, F.A.; Sabaghpour, S.H.; Chalabian, F. Effect of salicylic acid on yield, yield components and anatomical structures of chickpea (Cicer arietinum L.). J. Sci. Islam. Azad Univ. 2007, 16, 61–70. [Google Scholar]
- El-Taher, A.M.; El-Raouf, H.S.A.; Osman, N.A.; Azoz, S.N.; Omar, M.A.; Elkelish, A.; El-Hady, A.; Mahmoudy, A.M. Effect of salt stress and foliar application of salicylic acid on morphological, biochemical, anatomical, and productivity characteristics of cowpea (Vigna unguiculata L.) Plants. Plants 2021, 11, 115. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Band No. | Marker | After 24 h from Application | After 48 h from Application | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Molecular Weight (kDa) | Control | SA | NA | OA | BA | Control | SA | NA | OA | BA | |||
| Healthy | Infected | Healthy | Infected | ||||||||||
| 1 | 320 | + | + | + | + | + | + | + | + | + | + | + | + |
| 2 | 301 | − | − | + | − | − | − | − | − | − | − | − | − |
| 3 | 399 | − | − | − | − | + | − | − | − | − | − | − | − |
| 4 | 297 | − | + | − | − | − | − | − | − | − | − | − | − |
| 5 | 176 | − | − | − | + | + | + | − | − | − | − | − | − |
| 6 | 172 | − | − | + | − | + | + | − | − | + | + | + | + |
| 7 | 137 | − | − | − | − | − | − | + | − | + | + | − | + |
| 8 | 130 | − | − | − | − | + | + | + | + | − | − | + | − |
| 9 | 117 | + | + | − | − | + | + | − | − | − | − | − | − |
| 10 | 116 | − | − | − | − | − | − | − | − | − | − | − | + |
| 11 | 104 | − | + | + | + | − | − | − | + | + | + | + | + |
| 12 | 98 | + | − | − | − | + | − | − | − | − | − | − | + |
| 13 | 85 | − | − | − | + | + | + | − | − | − | − | + | − |
| 14 | 84 | − | − | − | − | − | + | + | + | + | − | − | − |
| 15 | 78 | + | − | − | − | − | − | − | − | − | + | + | − |
| 16 | 76 | − | − | − | − | + | + | − | − | − | − | − | − |
| 17 | 66 | + | − | − | − | − | − | + | + | − | + | + | + |
| 18 | 65 | − | + | + | + | + | + | − | − | + | − | − | − |
| 19 | 62 | − | − | − | − | + | − | − | − | − | + | − | − |
| 20 | 59 | − | − | − | + | + | + | − | + | + | + | + | + |
| 21 | 51 | − | − | + | + | + | + | + | + | + | + | + | + |
| 22 | 44 | − | − | − | − | − | − | − | − | + | + | + | − |
| 23 | 43 | − | + | + | − | + | + | + | + | − | − | − | − |
| 24 | 42 | − | − | − | − | − | − | − | − | + | − | + | − |
| 25 | 41 | − | − | − | + | + | + | + | + | − | − | − | + |
| 26 | 40 | − | + | + | + | + | + | − | − | − | − | + | − |
| 27 | 35 | − | − | − | − | − | − | − | + | + | + | + | + |
| Total bands | 5 | 7 | 8 | 9 | 16 | 14 | 8 | 10 | 11 | 11 | 13 | 11 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alnefaie, R.M.; EL-Sayed, S.A.; Ramadan, A.A.; Elmezien, A.I.; El-Taher, A.M.; Randhir, T.O.; Bondok, A. Physiological and Anatomical Responses of Faba Bean Plants Infected with Chocolate Spot Disease to Chemical Inducers. Life 2023, 13, 392. https://doi.org/10.3390/life13020392
Alnefaie RM, EL-Sayed SA, Ramadan AA, Elmezien AI, El-Taher AM, Randhir TO, Bondok A. Physiological and Anatomical Responses of Faba Bean Plants Infected with Chocolate Spot Disease to Chemical Inducers. Life. 2023; 13(2):392. https://doi.org/10.3390/life13020392
Chicago/Turabian StyleAlnefaie, Rasha M., Sahar A. EL-Sayed, Amany A. Ramadan, Ahmed I. Elmezien, Ahmed M. El-Taher, Timothy O. Randhir, and Ahmed Bondok. 2023. "Physiological and Anatomical Responses of Faba Bean Plants Infected with Chocolate Spot Disease to Chemical Inducers" Life 13, no. 2: 392. https://doi.org/10.3390/life13020392