
Effects of Temperature, pH, and NaCl Concentration on Biomass and Bioactive Compound Production by Synechocystis salina
 ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Microorganism
2.2. Experimental Design
2.3. Biomass Production
2.4. Biomass Quantification
2.5. Pigment Extraction and Quantification
2.5.1. Total Carotenoids and Phycobiliprotein Extraction and Quantification
2.5.2. Determination of Total Carotenoids Profile and Quantification
2.6. Determination of Antioxidant Capacity
2.7. Statistical Analysis
3. Results
3.1. Experimental Design
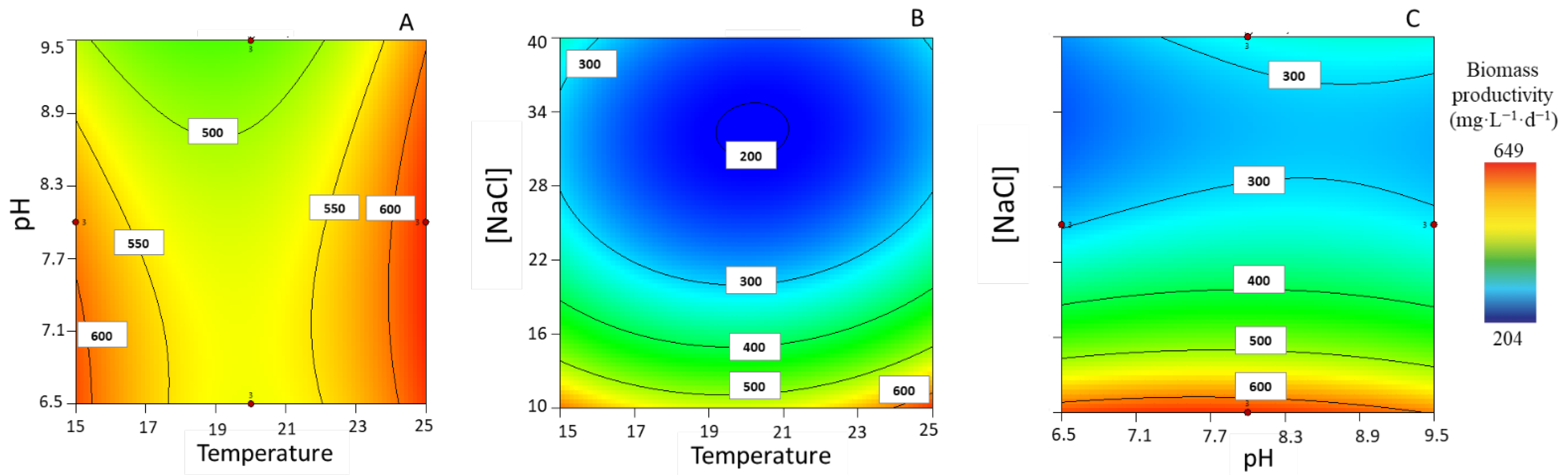
3.1.1. Biomass Productivity
3.1.2. Total Carotenoid Productivity
3.1.3. Total PBPs
3.1.4. PC Productivity
3.1.5. APC Productivity
3.1.6. PE Productivity
3.1.7. Ethanolic Antioxidant Compound Productivity
3.1.8. Aqueous Antioxidant Compounds Productivity
3.2. Model Optimal Conditions
3.3. HPLC Carotenoid Profile
4. Discussion
4.1. Biomass Production
4.2. Total Carotenoids and Profile
4.3. Phycobiliproteins
4.4. Antioxidant Capacity Extracts
4.5. Box-Behnken Design as an Optimization Tool for Cyanobacterial Production
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cardoso, L.G.; Lemos, P.V.F.; de Souza, C.O.; Oliveira, M.B.P.P.; Chinalia, F.A. Current advances in phytoremediation and biochemical composition of Arthrospira (Spirulina) grown in aquaculture wastewater. Aquac. Res. 2022, 53, 4931–4943. [Google Scholar] [CrossRef]
- Ashour, M.; Alprol, A.E.; Heneash, A.M.M.; Saleh, H.; Abualnaja, K.M.; Alhashmialameer, D.; Mansour, A.T. Ammonia bioremediation from aquaculture wastewater effluents using arthrospira platensis niof17/003: Impact of biodiesel residue and potential of ammonia-loaded biomass as rotifer feed. Materials 2021, 14, 5460. [Google Scholar] [CrossRef]
- Saini, D.K.; Pabbi, S.; Shukla, P. Cyanobacterial pigments: Perspectives and biotechnological approaches. Food Chem. Toxicol. 2018, 120, 616–624. [Google Scholar] [CrossRef]
- Mandal, M.K.; Chanu, N.K.; Chaurasia, N. Cyanobacterial pigments and their fluorescence characteristics: Applications in research and industry. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, A.K., Eds.; Academic Press Inc.: New York, NY, USA, 2020; pp. 55–72. [Google Scholar]
- Levasseur, W.; Perré, P.; Pozzobon, V. A review of high value-added molecules production by microalgae in light of the classification. Biotechnol. Adv. 2020, 41, 107545. [Google Scholar] [CrossRef]
- Pagels, F.; Guedes, A.C.; Amaro, H.M.; Kijjoa, A.; Vasconcelos, V. Phycobiliproteins from cyanobacteria: Chemistry and biotechnological applications. Biotechnol. Adv. 2019, 37, 422–443. [Google Scholar] [CrossRef]
- Hu, I.-C. Production of potential coproducts from microalgae. In Biofuels from Algae; Pandey, A., Chang, J.-S., Soccol, C.R., Lee, D.-J., Chisti, Y., Eds.; Elsevier: Amsterdam, The Netherlands, 2019; pp. 345–358. [Google Scholar]
- Assunção, J.; Amaro, H.M.; Malcata, F.X.; Guedes, A.C. Factorial Optimization of Ultrasound-Assisted Extraction of Phycocyanin from Synechocystis salina: Towards a Biorefinery Approach. Life 2022, 12, 1389. [Google Scholar] [CrossRef]
- Assunção, J.; Amaro, H.M.; Malcata, F.X.; Guedes, A.C. Cyanobacterial pigments: Photosynthetic function and biotechnological purposes. In The Pharmacological Potential of Cyanobacteria; Lopes, G., Silva, M., Vasconcelos, V., Eds.; Elsevier Academic Press: Amsterdam, The Netherlands, 2022; ISBN 9780128214916. [Google Scholar]
- Santos-Merino, M.; Singh, A.K.; Ducat, D.C. New Applications of Synthetic Biology Tools for Cyanobacterial Metabolic Engineering. Front. Bioeng. Biotechnol. 2019, 7, 33. [Google Scholar] [CrossRef] [Green Version]
- Heydarizadeh, P.; Poirier, I.; Loizeau, D.; Ulmann, L.; Mimouni, V.; Schoefs, B.; Bertrand, M. Plastids of Marine Phytoplankton Produce Bioactive Pigments and Lipids. Mar. Drugs 2013, 11, 3425–3471. [Google Scholar] [CrossRef]
- Yalcin, D. Growth, lipid content, and fatty acid profile of freshwater cyanobacteria Dolichospermum affine (Lemmermann) Wacklin, Hoffmann, & Komárek by using modified nutrient media. Aquac. Int. 2020, 28, 1371–1388. [Google Scholar] [CrossRef]
- Leão, P.N.; Nakamura, H.; Costa, M.; Pereira, A.R.; Martins, R.; Vasconcelos, V.; Gerwick, W.H.; Balskus, E.P. Biosynthesis-Assisted Structural Elucidation of the Bartolosides, Chlorinated Aromatic Glycolipids from Cyanobacteria. Angew. Chem. Int. Ed. 2015, 54, 11063–11067. [Google Scholar] [CrossRef]
- Afonso, T.B.; Costa, M.S.; de Castro, R.R.; Freitas, S.; Silva, A.; Schneider, M.P.C.; Martins, R.; Leão, P.N. Bartolosides E–K from a Marine Coccoid Cyanobacterium. J. Nat. Prod. 2016, 79, 2504–2513. [Google Scholar] [CrossRef]
- Gonçalves, A.L.; Pires, J.C.; Simões, M. Biotechnological potential of Synechocystis salina co-cultures with selected microalgae and cyanobacteria: Nutrients removal, biomass and lipid production. Bioresour. Technol. 2016, 200, 279–286. [Google Scholar] [CrossRef]
- Plaza, M.; Santoyo, S.; Jaime, L.; Reina, G.G.-B.; Herrero, M.; Señoráns, F.; Ibáñez, E. Screening for bioactive compounds from algae. J. Pharm. Biomed. Anal. 2010, 51, 450–455. [Google Scholar] [CrossRef]
- El Semary, N.A.; El Naby, M.A. Characterization of a Synechocystis sp. from Egypt with the potential of bioactive compounds production. World J. Microbiol. Biotechnol. 2009, 26, 1125–1133. [Google Scholar] [CrossRef]
- Assunção, J.; Amaro, H.M.; Lopes, G.; Tavares, T.; Malcata, F.X.; Guedes, A.C. Synechocystis salina: Potential bioactivity and combined extraction of added-value metabolites. J. Appl. Phycol. 2021, 33, 3731–3746. [Google Scholar] [CrossRef]
- Allen, M.M. Simple conditions for growth of unicellular blue-green algae on plates. J. Phycol. 1968, 4, 1–4. [Google Scholar] [CrossRef]
- Montgomery, D.C. Design and Analysis of Experiments, 10th ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2019; ISBN 9781119492443. [Google Scholar]
- Pagels, F.; Salvaterra, D.; Amaro, H.; Lopes, G.; Sousa-Pinto, I.; Vasconcelos, V.; Guedes, A.C. Factorial optimization of upstream process for Cyanobium sp. pigments production. J. Appl. Phycol. 2020, 32, 3861–3872. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Buschmann, C. Chlorophylls and Carotenoids Measurement and UV-VIS characterization Lichtenthaler 2001. Curr. Protoc. Food Anal. Chem. 2001, F4.3.1–F4.3.8. [Google Scholar] [CrossRef]
- Bennett, A.; Bogorad, L. Complementary chromatic adaptation in a filamentous blue-green alga. J. Cell Biol. 1973, 58, 419–435. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Pereira, R.D.; Malcata, F.X. Effects of temperature and pH on growth and antioxidant content of the microalga Scenedesmus obliquus. Biotechnol. Prog. 2011, 27, 1218–1224. [Google Scholar] [CrossRef]
- Guedes, A.C.; Amaro, H.M.; Gião, M.S.; Malcata, F.X. Optimization of ABTS radical cation assay specifically for determination of antioxidant capacity of intracellular extracts of microalgae and cyanobacteria. Food Chem. 2013, 138, 638–643. [Google Scholar] [CrossRef] [PubMed]
- Chentir, I.; Doumandji, A.; Ammar, J.; Zili, F.; Jridi, M.; Markou, G.; Ben Ouada, H. Induced change in Arthrospira sp. (Spirulina) intracellular and extracellular metabolites using multifactor stress combination approach. J. Appl. Phycol. 2017, 30, 1563–1574. [Google Scholar] [CrossRef]
- Rai, S.; Pandey, S.; Kumar, A.S.; Singh, P.K.; Agrawal, C.; Rai, L.C. Understanding the Mechanisms of Abiotic Stress Management in Cyanobacteria with Special Reference to Proteomics. In Stress Biology of Cyanobacteria; Srivastava, A.K., Rai, A.N., Neilan, B.A., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 112–131. [Google Scholar]
- Yu, Y.; You, L.; Liu, D.; Hollinshead, W.; Tang, Y.J.; Zhang, F. Development of Synechocystis sp. PCC 6803 as a Phototrophic Cell Factory. Mar. Drugs 2013, 11, 2894–2916. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nalley, J.O.; O′Donnell, D.R.; Litchman, E. Temperature effects on growth rates and fatty acid content in freshwater algae and cyanobacteria. Algal Res. 2018, 35, 500–507. [Google Scholar] [CrossRef]
- Li, T.Y.; Winayu, B.N.R.; Hsueh, H.T.; Chu, H. Growth factors arrangement enhances Thermosynechococcus sp. CL-1 carotenoid productivity during CO2 fixation. Food Bioprod. Process. 2020, 124, 258–265. [Google Scholar] [CrossRef]
- Paliwal, C.; Pancha, I.; Ghosh, T.; Maurya, R.; Chokshi, K.; Bharadwaj, S.V.; Ram, S.; Mishra, S. Selective carotenoid accumulation by varying nutrient media and salinity in Synechocystis sp. CCNM 2501. Bioresour. Technol. 2015, 197, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Vonshak, A.; Guy, R.; Guy, M. The response of the filamentous cyanobacterium Spirulina platensis to salt stress. Arch. Microbiol. 1988, 150, 417–420. [Google Scholar] [CrossRef]
- Fal, S.; Aasfar, A.; Rabie, R.; Smouni, A.; Arroussi, H.E. Salt induced oxidative stress alters physiological, biochemical and metabolomic responses of green microalga Chlamydomonas reinhardtii. Heliyon 2022, 8, e08811. [Google Scholar] [CrossRef]
- Pagels, F.; Vasconcelos, V.; Guedes, A. Carotenoids from Cyanobacteria: Biotechnological Potential and Optimization Strategies. Biomolecules 2021, 11, 735. [Google Scholar] [CrossRef]
- Prasanna, R.; Sood, A.; Jaiswal, P.; Nayak, S.; Gupta, V.; Chaudhary, V.; Joshi, M.; Natarajan, C. Rediscovering cyanobacteria as valuable sources of bioactive compounds (Review). Appl. Biochem. Microbiol. 2010, 46, 119–134. [Google Scholar] [CrossRef]
- Torregrosa-Crespo, J.; Montero, Z.; Fuentes, J.L.; García-Galbis, M.R.; Garbayo, I.; Vílchez, C.; Martínez-Espinosa, R.M. Exploring the Valuable Carotenoids for the Large-Scale Production by Marine Microorganisms. Mar. Drugs 2018, 16, 203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschberg, J.; Chamovitz, D. Carotenoids in Cyanobacteria. In The Molecular Biology of Cyanobacteria; Springer: Dordrecht, The Netherlands, 1994; pp. 559–579. [Google Scholar]
- Markou, G.; Nerantzis, E. Microalgae for high-value compounds and biofuels production: A review with focus on cultivation under stress conditions. Biotechnol. Adv. 2013, 31, 1532–1542. [Google Scholar] [CrossRef] [PubMed]
- Guedes, A.C.; Amaro, H.M.; Malcata, F.X. Microalgae as Sources of Carotenoids. Mar. Drugs 2011, 9, 625–644. [Google Scholar] [CrossRef] [PubMed]
- Kłodawska, K.; Bujas, A.; Turos-Cabal, M.; Żbik, P.; Fu, P.; Malec, P. Effect of growth temperature on biosynthesis and accumulation of carotenoids in cyanobacterium Anabaena sp. PCC 7120 under diazotrophic conditions. Microbiol. Res. 2019, 226, 34–40. [Google Scholar] [CrossRef]
- Ismaiel, M.M.S.; El-Ayouty, Y.M.; Piercey-Normore, M. Role of pH on antioxidants production by Spirulina (Arthrospira) platensis. Braz. J. Microbiol. 2016, 47, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Mehnert, G.; Rücker, J.; Nicklisch, A.; Leunert, F.; Wiedner, C. Effects of thermal acclimation and photoacclimation on lipophilic pigments in an invasive and a native cyanobacterium of temperate regions. Eur. J. Phycol. 2012, 47, 182–192. [Google Scholar] [CrossRef]
- Singh, N.; Sonani, R.R.; Rastogi, R.P.; Madamwar, D. The phycobilisomes: An early requisite for efficient photosynthesis in cyanobacteria. EXCLI J. 2015, 14, 268–289. [Google Scholar] [CrossRef]
- Hemlata; Fatma, T. Screening of Cyanobacteria for Phycobiliproteins and Effect of Different Environmental Stress on Its Yield. Bull. Environ. Contam. Toxicol. 2009, 83, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.M.; Kumar, K.; Das, D. Physicochemical parameters optimization, and purification of phycobiliproteins from the isolated Nostoc sp. Bioresour. Technol. 2014, 166, 541–547. [Google Scholar] [CrossRef]
- Chaneva, G.; Furnadzhieva, S.; Minkova, K.; Lukavsky, J. Effect of light and temperature on the cyanobacterium Arthronema africanum—A prospective phycobiliprotein-producing strain. J. Appl. Phycol. 2007, 19, 537–544. [Google Scholar] [CrossRef]
- Keithellakpam, O.; Nath, T.; Oinam, A.; Thingujam, I.; Oinam, G.; Dutt, S.G. Effect of external pH on cyanobacterial phycobiliproteins production and ammonium excretion. J. Appl. Biol. Biotechnol. 2015, 3, 38–42. [Google Scholar] [CrossRef] [Green Version]
- Maurya, S.S.; Maurya, J.N.; Pandey, V.D. Factors regulating phycobiliprotein production in cyanobacteria. Int. J. Curr. Microbiol. Appl. Sci. 2014, 3, 764–771. [Google Scholar]
- Hong, S.-J.; Lee, C.-G. Statistical optimization of culture media for production of phycobiliprotein by Synechocystis sp. PCC 6701. Biotechnol. Bioprocess Eng. 2008, 13, 491–498. [Google Scholar] [CrossRef]
- Lee, H.; Noh, Y.; Hong, S.-J.; Lee, H.; Kim, D.-M.; Cho, B.-K.; Lee, C.-G.; Choi, H.-K. Photosynthetic pigment production and metabolic and lipidomic alterations in the marine cyanobacteria Synechocystis sp. PCC 7338 under various salinity conditions. J. Appl. Phycol. 2020, 33, 197–209. [Google Scholar] [CrossRef]
- López-Pacheco, I.Y.; Fuentes-Tristan, S.; Rodas-Zuluaga, L.I.; Castillo-Zacarías, C.; Pedro-Carrillo, I.; Martínez-Prado, M.A.; Iqbal, H.M.N.; Parra-Saldívar, R. Influence of Low Salt Concentration on Growth Behavior and General Biomass Composition in Lyngbya purpurem (Cyanobacteria). Mar. Drugs 2020, 18, 621. [Google Scholar] [CrossRef] [PubMed]
- Sharma, G.; Kumar, M.; Ali, M.I.; Jasuja, N.D. Effect of carbon content, salinity and pH on Spirulina platensis for phycocyanin, allophycocyanin and phycoerythrin accumulation. J. Microb. Biochem. Technol. 2014, 6, 202–206. [Google Scholar] [CrossRef] [Green Version]
- Lemus, N.; Guevara, J.; Lodeiros, C.; Vásquez-Suárez, A.; Freites, L.; Licet, B. Growth and biochemical composition of Limnothrix sp. at different salinities and concentrations of nitrate. Colomb. J. Biotechnol. 2013, 15, 159–166. [Google Scholar]
- Babele, P.K.; Kumar, J.; Chaturvedi, V. Proteomic De-Regulation in Cyanobacteria in Response to Abiotic Stresses. Front. Microbiol. 2019, 10, 1315. [Google Scholar] [CrossRef] [Green Version]
- Guedes, A.C.; Gião, M.S.; Seabra, R.; Ferreira, A.C.S.; Tamagnini, P.; Moradas-Ferreira, P.; Malcata, F.X. Evaluation of the Antioxidant Activity of Cell Extracts from Microalgae. Mar. Drugs 2013, 11, 1256–1270. [Google Scholar] [CrossRef] [Green Version]
- Singh, D.P.; Prabha, R.; Meena, K.K.; Sharma, L.; Sharma, A.K. Induced Accumulation of Polyphenolics and Flavonoids in Cyanobacteria under Salt Stress Protects Organisms through Enhanced Antioxidant Activity. Am. J. Plant Sci. 2014, 5, 726–735. [Google Scholar] [CrossRef]
- Azim, N.H.; Subki, A.; Yusof, Z.N.B. Abiotic stresses induce total phenolic, total flavonoid and antioxidant properties in Malaysian indigenous microalgae and cyanobacterium. Malays. J. Microbiol. 2018, 14, 25–33. [Google Scholar] [CrossRef]
- Patipong, T.; Hibino, T.; Waditee-Sirisattha, R.; Kageyama, H. Induction of antioxidative activity and antioxidant molecules in the halotolerant cyanobacterium Halothece sp. PCC7418 by temperature shift. Nat. Prod. Commun. 2019, 14, 1–6. [Google Scholar]
- Singh, E.V.S.; Mishra, N.A.K. Salinity-induced oxidative stress-mediated change in fatty acids composition of cyanobacterium Synechococcus sp. PCC7942. Int. J. Environ. Sci. Technol. 2018, 16, 875–886. [Google Scholar]
- Tanaka, K.; Shimakawa, G.; Nakanishi, S. Time-of-day-dependent responses of cyanobacterial cellular viability against oxidative stress. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Chi, X.; Yang, Q.; Zhao, F.; Qin, S.; Yang, Y.; Shen, J.; Lin, H. Comparative Analysis of Fatty Acid Desaturases in Cyanobacterial Genomes. Comp. Funct. Genom. 2008, 2008, 1–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tasaka, Y.; Gombos, Z.; Nishiyama, Y.; Mohanty, P.; Ohba, T.; Ohki, K.; Murata, N. Targeted mutagenesis of acyl-lipid desaturases in Synechocystis: Evidence for the important roles of polyunsaturated membrane lipids in growth, respiration and photosynthesis. EMBO J. 1996, 15, 6416–6425. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Long, L.; Sun, X.; Wu, H.; Li, T.; Xiang, W. Optimization of Medium Using Response Surface Methodology for Lipid Production by Scenedesmus sp. Mar. Drugs 2014, 12, 1245–1257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bajwa, K.; Bishnoi, N.R.; Kirrolia, A.; Gupta, S.; Selvan, S.T. Response surface methodology as a statistical tool for optimization of physio-biochemical cellular components of microalgae Chlorella pyrenoidosa for biodiesel production. Appl. Water Sci. 2019, 9, 128. [Google Scholar] [CrossRef] [Green Version]
- Henrard, A.; de Morais, M.; Costa, J. Vertical tubular photobioreactor for semicontinuous culture of Cyanobium sp. Bioresour. Technol. 2011, 102, 4897–4900. [Google Scholar] [CrossRef] [PubMed]
- Belhaj, D.; Frikha, D.; Athmouni, K.; Jerbi, B.; Ahmed, M.B.; Bouallagui, Z.; Kallel, M.; Maalej, S.; Zhou, J.; Ayadi, H. Box-Behnken design for extraction optimization of crude polysaccharides from Tunisian Phormidium versicolor cyanobacteria (NCC 466): Partial characterization, in vitro antioxidant and antimicrobial activities. Int. J. Biol. Macromol. 2017, 105, 1501–1510. [Google Scholar] [CrossRef]
- Gündoğdu, T.K.; Deniz, İ.; Çalişkan, G.; Şahin, E.S.; Azbar, N. Experimental design methods for bioengineering applications. Crit. Rev. Biotechnol. 2016, 36, 368–388. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Factors | |||
|---|---|---|---|
| Runs | A: Temperature (°C) | B: pH | C: [NaCl] (g·L−1) |
| 1 | 15 | 6.5 | 25 |
| 2 | 15 | 8 | 10 |
| 3 | 15 | 8 | 40 |
| 4 | 15 | 9.5 | 25 |
| 5 | 20 | 6.5 | 10 |
| 6 | 20 | 6.5 | 40 |
| 7 | 20 | 8 | 25 |
| 8 | 20 | 9.5 | 10 |
| 9 | 20 | 9.5 | 40 |
| 10 | 25 | 6.5 | 25 |
| 11 | 25 | 8 | 10 |
| 12 | 25 | 8 | 40 |
| 13 | 25 | 9.5 | 25 |
| Parameters (Objective Function) | p-Value | R2 | Equation |
|---|---|---|---|
| Px (mg·L−1·d−1) | <0.0001 | 0.98 | 232.67 + 8.17 T − 10.75 pH − 141.75 [NaCl] + 18.75 T × pH − 16.08 T × [NaCl] + 22.42 pH × [NaCl] + 87.21 T2 − 23.79 pH2 + 149.37 [NaCl]2 |
| Total carotenoids (mg·L−1·d−1) | <0.0001 | 0.72 | 0.1690 + 0.0386 T + 0.0159 pH − 0.0219 [NaCl] − 0.0165 T × pH − 0.0045 T × [NaCl] − 0.0020 pH × [NaCl] + 0.0288 T2 − 0.0423 pH2 + 0.0647 [NaCl] 2 |
| TPBP (mg·L−1·d−1) | <0.0001 | 0.81 | 2.22 + 0.2687 T− 0.0358 pH − 0.2182[NaCl] + 0.4686 T × pH − 0.0596 T × [NaCl] − 0.3596 pH × [NaCl] − 0.6547 T2 + 0.1737 pH2 − 0.0484 [NaCl] 2 |
| PC (mg·L−1·d−1) | <0.0001 | 0.86 | 1.66 + 0.340 T + 0.1933 pH − 0.2354 [NaCl] + 0.1055 T × pH − 0.1265 T × [NaCl] − 0.0334 pH × [NaCl] − 0.4497 T2 − 0.2247 pH2 − 0.1265 [NaCl]2 |
| APC (mg·L−1·d−1) | <0.0001 | 0.85 | 0.7785 + 0.0121 T − 0.0330 pH − 0.2216 [NaCl] + 0.2486 T × pH − 0.0439 T × [NaCl] − 0.0526 pH × [NaCl] − 0.1351 T2 − 0.0409 pH2 − 0.1403 [NaCl]2 |
| PE (mg·L−1·d−1) | <0.0001 | 0.91 | 0.4306 − 0.0047 T − 0.0512 pH—0.0317 [NaCl] + 0.3337 T × pH + 0.0126 T × [NaCl] − 0.0783 pH × [NaCl] − 0.0222 T2 + 0.1471 pH2 − 0.1927 [NaCl]2 |
| AOX-EtOH (mgTE·L−1·d−1) | <0.0001 | 0.92 | 1.53 − 0.0445 T + 0.4679 pH − 0.1503 [NaCl] − 0.3758 T × pH − 0.0756 T × [NaCl] + 0.1469 pH × [NaCl] + 0.1510 T2 − 0.5047 pH2 + 0.1395 [NaCl]2 |
| AOX-Water (mgTE·L−1·d−1) | <0.0001 | 0.90 | 1.26 − 0.1356 T + 0.1022 pH − 0.0120 [NaCl] − 0.0403 T × pH − 0.0800 T × [NaCl] − 0.0392 pH × [NaCl] − 0.3917 T2 − 0.2588 pH2 − 0.1617 [NaCl]2 |
| Model Prediction | |||||||
|---|---|---|---|---|---|---|---|
| Objective Function | Experimental Value of the Central Point (Control) | T (°C) | pH | [NaCl] (g·L−1) | Maximum Productivity | Desirability | Extent of Increase (%) |
| Px (mg·L−1·d−1) | 233.10 ± 4.01 | 25 | 7.50 | 10 | 637.43 ± 20.56 | 0.974 | 175% |
| Total carotenoids mg·L−1·d−1 | 0.172 ± 0.00 | 25 | 8.00 | 10 | 0.33 ± 0.04 | 0.992 | 91% |
| TPBP (mg·L−1·d−1) | 2.22 ± 0.06 | 22.9 | 9.48 | 10.9 | 3.09 ± 0.32 | 1 | 39% |
| PC (mg·L−1·d−1) | 1.66 ± 0.04 | 22.9 | 8.96 | 10 | 1.97 ± 0.20 | 0.974 | 19% |
| APC (mg·L−1·d−1) | 0.78 ± 0.02 | 25 | 9.50 | 10 | 1.17 ± 0.12 | 0.940 | 50% |
| PE (mg·L−1·d−1) | 0.43 ± 0.03 | 15 | 6.50 | 26.3 | 0.99 ± 0.09 | 0.968 | 130% |
| AOX-EtOH (mgTE·L−1·d−1) | 2.10 ± 0.01 | 15 | 9.45 | 16.3 | 2.24 ± 0.18 | 1 | 6% |
| AOX-Water (mgTE·L−1·d−1) | 1.45 ± 0.02 | 19.2 | 8.20 | 25.5 | 1.47 ± 0.09 | 0.988 | 1.38% |
| Peak | Retention Time (min) | Identified Pigment | Concentration of Pigment (µg·gDW−1) |
|---|---|---|---|
| 1 | 10.2 | Unidentified carotenoid * | 2.18 ± 0.18 a |
| 2 | 13.2 | Zeaxanthin | 3.26 ± 0.12 a |
| 4 | 24.6 | Chl a | 63.25 ± 1.16 b |
| 5 | 25.2 | Echinenone | 0.35 ± 0.01 c |
| 6 | 25.8 | Unidentified carotenoid * | 0.68 ± 0.04 c |
| 7 | 32.8 | β-carotene | 3.41 ± 0.07 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Assunção, J.; Amaro, H.M.; Tavares, T.; Malcata, F.X.; Guedes, A.C. Effects of Temperature, pH, and NaCl Concentration on Biomass and Bioactive Compound Production by Synechocystis salina. Life 2023, 13, 187. https://doi.org/10.3390/life13010187
Assunção J, Amaro HM, Tavares T, Malcata FX, Guedes AC. Effects of Temperature, pH, and NaCl Concentration on Biomass and Bioactive Compound Production by Synechocystis salina. Life. 2023; 13(1):187. https://doi.org/10.3390/life13010187
Chicago/Turabian StyleAssunção, Joana, Helena M. Amaro, Tânia Tavares, F. Xavier Malcata, and A. Catarina Guedes. 2023. "Effects of Temperature, pH, and NaCl Concentration on Biomass and Bioactive Compound Production by Synechocystis salina" Life 13, no. 1: 187. https://doi.org/10.3390/life13010187