

IL-18 Signaling Is Essential for Causing Streptococcal Toxic Shock-like Syndrome (STSLS)
by
 and
and
Lei Xu
1,2,†,
Yue Zeng
1,2,†,
Peiying Gao
1,2,
Xi Lu
1,2,
Kunlong Xia
1,2,
Liting Zhou
1,2,
Chengfeng Zhang
1,2,
Chenyang Yi
1,2 and
Anding Zhang
1,2,3,4,* 1
State Key Laboratory of Agricultural Microbiology, Hubei Hongshan Laboratory, College of Veterinary Medicine, Huazhong Agricultural University, Wuhan 430070, China
2
Key Laboratory of Preventive Veterinary Medicine in Hubei Province, The Cooperative Innovation Center for Sustainable Pig Production, Wuhan 430070, China
3
Key Laboratory of Development of Veterinary Diagnostic Products, Ministry of Agriculture of the People’s Republic of China, Wuhan 430070, China
4
International Research Center for Animal Disease, Ministry of Science and Technology of the People’s Republic of China, Wuhan 430070, China
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Life 2022, 12(9), 1324; https://doi.org/10.3390/life12091324
Submission received: 14 July 2022
/
Revised: 9 August 2022
/
Accepted: 24 August 2022
/
Published: 26 August 2022
(This article belongs to the Section Microbiology)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Streptococcus suis (S. suis) is an emerging zoonotic pathogen that can cause multiple diseases, including streptococcal toxic shock-like syndrome (STSLS). The S. suis SC-19 strain could cause NOD-like receptor thermal protein domain-associated protein 3 (NLRP3) inflammasome hyperactivation, then induce a cytokine storm and STSLS. Although IL-18 is the downstream effector of NLRP3 signaling, the role of IL-18 signaling on STSLS remains to be elucidated. Thus, il18r1 gene knockout mice were constructed and challenged with the SC-19 strain. Alleviated clinical signs and tissue damages, as well as improved survival were observed in il18r−/− mice compared with the WT mice post-SC-19 challenge. Meanwhile, an obvious decrease in the inflammatory cytokine levels in blood was observed in the il18r-/- mice infected with SC-19. Therefore, IL-18, the downstream effector of NLRP3 inflammasome activation, was responsible for the cytokine storm and STSLS development caused by S. suis, suggesting that IL-18/IL-18Rα signaling could serve as a new target for STSLS.
1. Introduction
Streptococcus suis (S. suis) is a common and important zoonotic agent that can cause huge economic losses in the swine industry of the world and lead to a significant threat to the public health of humans and immunocompetent patients [1,2]. S. suis infection in humans causes a variety of serious diseases, including meningitis, endocarditis, arthritis, and sepsis [3]. In 1968, human infection with S. suis was first reported and, as of now, more than 1600 people have been infected with S. suis, primarily in Asia, including China, Vietnam, and Thailand [3,4,5,6]. However, in 1998 and 2005, the outbreak of S. suis serotype 2 unusually caused two large-scale human S. suis epidemics, in which 240 humans were infected and 53 died in China [7,8,9]. Moreover, 97.4% of fatal cases were observed with streptococcal toxic-shock-like syndrome (STSLS), including the hallmarks of acute high fever, blood spots, shock, vascular collapse, hypotension, dysfunction of multiple organs, and acute death [8,10], which has attracted widespread attention of the world.
The clinical investigations showed that STSLS patients died with severe inflammation, characterized with cytokine storms and dysfunction of multiple organs [7]. Subsequent studies further confirmed that the production of inflammatory cytokine storms was critical for STSLS [11], and inhibiting excessive inflammatory response could significantly alleviate acute death caused by STSLS [12,13]. Our previous study indicated that high expression of suilysin (SLY) was required for the highly pathogenic S. suis strain SC-19 to cause NOD-like receptor thermal protein domain-associated protein 3 (NLRP3) inflammasome hyperactivation, which in turn causes the induction of cytokine storms and STSLS [14]. This was also confirmed by another group [15]. However, it remains to be elucidated how NLRP3 activation causes severe inflammation.
The activation of inflammasomes could lead to pro-caspase-1 into an active protease, which mediates the processing of several targets: it cleaves pro-IL-1β and pro-IL-18 into their biologically active forms and also cleaves gasdermin D (GSDMD), which leads to a particular form of cell death called pyroptosis for secretion of mature IL-1β and IL-18 [16,17,18,19,20]. IL-1β is required for controlling bacterial burdens caused by the ST1 strain but not by the ST7 strain, and il1β gene-deficient mice were more susceptible to S. suis strains in [21]. This was in coincidence with the results of an earlier study indicating the protective role of IL-1 on the resistance to S. suis infection [22]. However, GSDMD-deficient mice were also resistant to STSLS development, indicating that pyroptosis, rather than IL-1β signaling trigged by inflammasome activation, promotes STSLS development [15]. IFN-γ, induced uniquely by IL-18 but not by IL-1β [23,24], also played a broad and important role in severe inflammatory responses and organ injury during shock syndrome [11,25,26]. Furthermore, the highly virulent strain SC-19 caused higher levels of IL-18 and IFN-γ than the meningitic strain P1/7 [27]. These authors suggested that IL-18, as a NLRP3 downstream signaling molecule, may induce high levels of IFN-γ during STSLS. However, the role of IL-18 on STSLS remains to be elucidated.
Therefore, the study aimed to clarify the role of IL-18 signaling during the development of STSLS, and also to delineate signaling cascades for severe inflammatory response during STSLS.
2. Materials and Methods
2.1. S. suis Strain and Culture Conditions
The S. suis serotype 2 epidemic strain SC-19 (ST7) was used in the present study. The epidemic strain SC-19 was originally isolated from the brain of a diseased pig in China and has been widely used in several models of STSLS [28,29]. S. suis was cultured in tryptic soy broth (TSB, Difco Laboratories, Detroit, MI, USA) or on tryptic soy agar (TSA, Difco Laboratories, Detroit, MI, USA) plates with 10% (vol/vol) newborn bovine serum (Sijiqing, Hangzhou, China) at 37 °C.
2.2. Ethics Statement
The mice used in this study were female, 4 to 6 weeks old C57BL/6, and were bred and housed in SPF conditions. All experiments involving infectious S. suis were conducted in strict accordance with the Guide for the Care and Use of Laboratory Animals Monitoring Committee of Hubei Province, China, and approved by the Scientific Ethics Committee of Huazhong Agricultural University. All efforts were made to minimize the suffering of the animals used in the study.
2.3. Construction of il18r1 Gene Knockout Mice
In order to evaluate the role of IL-18 on STSLS, interleukin-18 receptor 1 (il18r1) gene knockout mice were constructed using the CRISPR/Cas9 gene-editing system as previously described [30,31]. In brief, zygotes were collected from sexually immature female C57BL/6 mice. Then, an sgRNA (5′-GCCACCATGAGATGGTTCAA-3′) targeting exon 4 of the il18r1 gene (MGI: 105383) and the Cas9 mRNA were injected into the cytoplasm of pronuclear stage embryos. Finally, the injected embryos were transferred into the oviduct of the recipient mother for KO mouse productions. The newborn mice were genotyped by PCR followed by DNA sequencing analysis. The wild-type, heterozygote, and knockout mice were used for control with each other. Primers for genotyping were as follows: il18r-F, 5′-AGGGGGATCAGGGAAAAATCAC-3′ (forward); il18r-R, 5′-AAATTTAGAGTGACTGGCTTAATA-3′ (reverse); il18r-f1(wt), 5′-CACCATGAGATGGTTCAAAGG-3′ (forward); and il18r-f2(ko), 5′-CACCATGAGATGGTTGTGCTT-3′ (forward). The DNA fragment for WT and HET was 405 bp, and the DNA fragment for KO was 390 bp. All mice were bred and housed in SPF conditions.
2.4. Experimental Infections of Mice
Female, four- to six-week old il18r1 gene knockout mice (il18r1-/-) and C57BL/6 (il18r1+/+) mice (10 mice per group) were challenged with 4 × 108 CFUs of SC-19 by an intraperitoneal (i.p.) injection to directly evaluate the effect of IL-18 on STSLS development [27]. The clinical scores were assigned based on depression, swollen eyes, rough hair coat, and lethargy. The details were described as follows: 0 = normal response to external stimuli; 1 = ruffled coat and slow response to external stimuli; 2 = responds only to repeated stimuli; 3 = no response to external stimuli or walking in circles; and 4 = dead. Mice exhibiting extreme lethargy or neurological signs (score = 3) were considered moribund and were humanely euthanized [32].
To further analyze the role of IL-18 on STSLS, the level of cytokines and bacterial burden were also evaluated during S. suis infection [31]. At 6 h and 12 h post-infection, mice were euthanized, bacterial loads were measured in fifty microliters of blood and the remaining blood was used to analyze the level of cytokines. Half of the liver, lung, tissues, and brain were used for bacterial load analysis. The remaining lung, liver, and spleen tissues were fixed in 10% neutral buffered formalin for histopathology examinations.
2.5. Multiplex Cytokine Assays
Serum was obtained by centrifugation at 500 g for 30 min at 4 °C, and stored at −80 °C until analysis. Serum levels of cytokines TNF-α, IFN-γ, IL-6, IL-1β, IL-17A, and IL-12p70 were measured in all mice samples, and quantified using U-PLEX electrochemiluminescence ELISA (Meso Scale Discovery, MD, USA). Data acquisition was achieved by MESOTM QuickPlex SQ120 (Meso Scale Discovery, MD, USA) and analyzed in the MSD Discovery Workbench Desktop 4.0 software (Meso Scale Discovery, MD, USA), where values were expressed in pg/mL for each cytokine.
2.6. Histopathology Examinations
Tissues were fixed in 10% neutral buffered formalin for over 24 h. After embedding in paraffin, tissues were cut into 2–3 μm sections. Subsequently, dewaxing was performed through a xylene and ethanol series to deionized water. Finally, the sections were stained with hematoxylin and eosin (H&E) following standard procedures [13], and examined under light microscopy (Olympus, Tokyo, Japan).
2.7. Bacterial Load in The Blood and Tissues
The colonization capabilities of the SC-19 strain were detected in il18r1-/- mice and il18r1+/+ mice as described previously [33]. Briefly, bacterial counts in blood were determined by plating serial dilutions on TSA plates. The tissues were weighed, homogenized, serially diluted, and then plated on TSA plates to evaluate the bacterial counts [34].
2.8. Statistical Analysis
GraphPad Prism 6 software was used in data analysis by two-tailed, unpaired t-tests. A log-rank test and two-way RM ANOVA were used to compare survival rates and clinical scores, respectively. All assays were repeated ≥3 times. For all tests, a value of p < 0.05 was considered to be of significance.
3. Results and Discussion
3.1. Construction of il18r1 Gene Knockout Mice
In order to analyze the effects of IL-18 on STSLS, il18r1 knockout mice were constructed with the CRISPR/Cas9 system (Figure 1). Sanger sequencing indicated that there is 8 bp deletion (CAAAGGCA) in the fourth exon of the il18r1 gene in the il18r1–/– mouse. It indicated the frameshift of the il18r1 gene, suggesting successfully constructed il18r1 gene knockout mice.
3.2. Knockout of il18r1 Could Significantly Decrease Mortality during STSLS
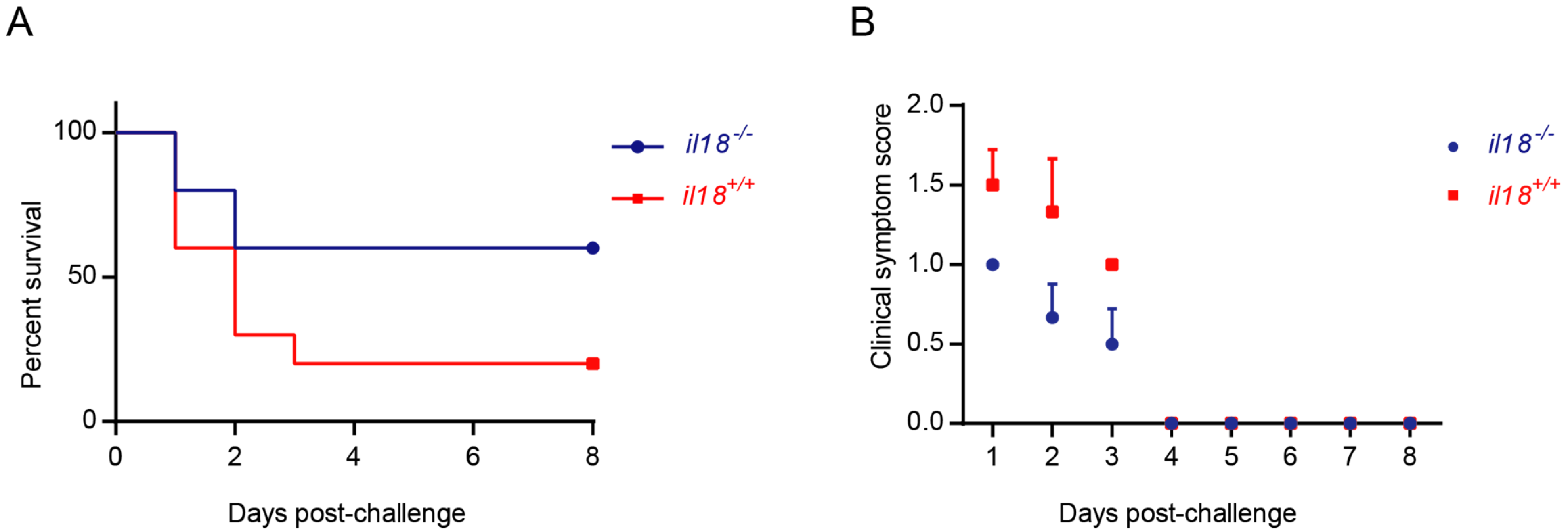
Highly virulent S. suis infection induces STSLS, which is characterized by inflammatory cytokine storms, multi-organ damages, and ultimately, acute death [7,8]. In order to further analyze the function of IL-18 on STSLS, il18r1-/- and il18r1+/+ mice were challenged with the highly virulent S. suis strain SC-19. As described before, the SC-19 strain caused severe death in il18r1+/+ mice [34]. However, morbidity and mortality caused by S. suis infection were significantly decreased in il18r1-/- mice, and acute death was also reduced (Figure 2). This indicated that IL-18 might play a vital role on STSLS caused by the highly virulent strain SC-19.
3.3. Knockout of il18r1 Could Decrease the Tissue Damages during STSLS
As described before [14], infection with the SC-19 strain can cause acute multi-organ dysfunctions in the il18r1+/+ mice, such as severe congestion and infiltration of inflammatory cells in the lung, necrosis and vacuolated degeneration in the liver, and congestion in the spleen (Figure 3). However, the infection on the il18r1-/- mice did not show severe tissue injury. The results indicated that IL-18 might play an essential role in multi-organ dysfunctions and acute death during STSLS.
3.4. Knockout of il18r1 Decreases Serum Inflammatory Cytokine Storms but Does Not Significantly Decrease Bacterial Load
The SC-19 strain can cause high levels of inflammatory cytokine storms and high bacterial burden in mice [14]. Although IL-18 signaling was reported to play a role in bacterial control and protection against Streptococcus pneumoniae and Streptococcus agalactiae infection [35,36,37,38], there was no significant difference in bacterial load of various tissues in il18r-/- mice and il18r1+/+ mice (Figure 4). This indicates different roles of IL-18 in different bacterial infections.
However, the levels of blood cytokines such as IL-17A and IFN-γ were significantly reduced in il18r-/- mice compared with il18r1+/+ mice (Figure 4). This indicates that IL-18, one of the downstream effectors for NLRP3 inflammasome activation, was responsible for inflammatory cytokine storms and STSLS. In addition, the Chinese epidemic S. suis strain induced a strong and fast IFN-γ response by NK cells. Furthermore, IFN-γ-deficient mice infected with the epidemic S. suis showed significantly better survival rates than wild-type mice [11]. The decreased level of IFN-γ in il18r-/- demonstrates that IL-18 was responsible for IFN-γ induction during STSLS development. This suggests that NLRP3/IL-18/IFN-γ signaling was responsible for the STSLS development, which could serve as a new target for STSLS.
3.5. Limitation
The CRISPR/Cas9 system has become the mainstream technology for gene knockout, even if it has off-target potential. To illustrate the function of IL-18, the il18r1 gene knockout mice were constructed based on this technology, so we could not completely rule out the side effects due to off-target potential.
4. Conclusions
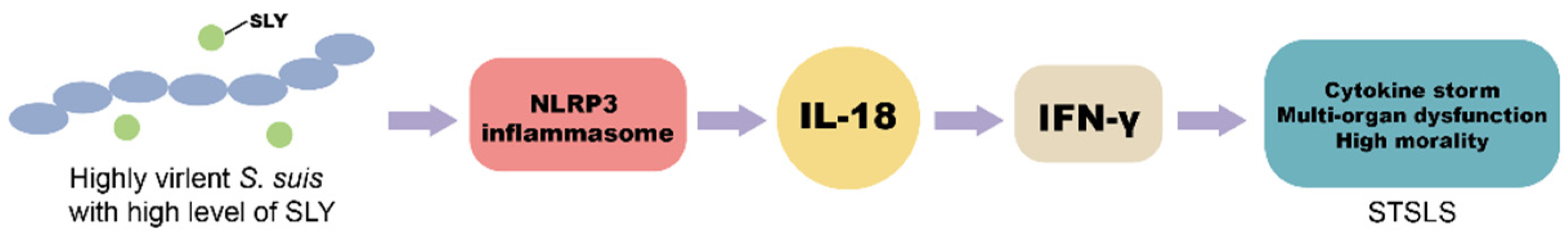
A previous study has demonstrated that SLY was the essential and sufficient condition for NLRP3 inflammasome hyperactivation, causing cytokine storms and STSLS [14]. The present study indicated that IL-18, the downstream of NLPR3 signaling, was responsible for IFN-γ induction and STSLS development (Figure 5).
Author Contributions
Conceptualization, A.Z. and L.X.; methodology and formal analysis, L.X., Y.Z., P.G. and K.X.; investigation, L.X., K.X., L.Z. and C.Z.; writing—original draft preparation, L.X. and X.L.; writing—review and editing, L.X., C.Y. and A.Z.; data curation, L.X. and X.L.; visualization, L.X.; resources, supervision, project administration, and funding acquisition, A.Z. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China, grant number 2021YFD1800402, and the National Natural Science Foundation of China, grant number 31972649.
Institutional Review Board Statement
The experimental infections protocol was approved by the Scientific Ethics Committee of Huazhong Agricultural University (protocol code HZAUMO-2019-048 and 04/20/2019 of approval).
Informed Consent Statement
Not applicable.
Data Availability Statement
All data are available within the article or from the corresponding author upon reasonable request.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
References
- Segura, M. Streptococcus suis research: Progress and challenges. Pathogens 2020, 9, 707. [Google Scholar] [CrossRef]
- Xing, X.; Bi, S.; Fan, X.; Jin, M.; Liu, W.; Wang, B. Intranasal Vaccination With Multiple Virulence Factors Promotes Mucosal Clearance of Streptococcus suis Across Serotypes and Protects Against Meningitis in Mice. J. Infect. Dis. 2019, 220, 1679–1687. [Google Scholar] [CrossRef] [PubMed]
- Huong, V.T.; Ha, N.; Huy, N.T.; Horby, P.; Nghia, H.D.; Thiem, V.D.; Zhu, X.; Hoa, N.T.; Hien, T.T.; Zamora, J.; et al. Epidemiology, clinical manifestations, and outcomes of Streptococcus suis infection in humans. Emerg. Infect. Dis. 2014, 20, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Staats, J.J.; Feder, I.; Okwumabua, O.; Chengappa, M.M. Streptococcus suis: Past and present. Vet. Res. Commun. 1997, 21, 381–407. [Google Scholar] [CrossRef]
- Segura, M.; Aragon, V.; Brockmeier, S.L.; Gebhart, C.; Greeff, A.; Kerdsin, A.; O’Dea, M.A.; Okura, M.; Salery, M.; Schultsz, C.; et al. Update on Streptococcus suis research and prevention in the era of antimicrobial restriction: 4th international workshop on S. suis. Pathogens 2020, 9, 374. [Google Scholar] [CrossRef]
- Kerdsin, A.; Segura, M.; Fittipaldi, N.; Gottschalk, M. Sociocultural Factors Influencing Human Streptococcus suis Disease in Southeast Asia. Foods 2022, 11, 1190. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Zheng, H.; Zhang, J.; Jing, H.; Wang, L.; Xiong, Y.; Wang, W.; Zhou, Z.; Sun, Q.; Luo, X.; et al. Clinical, experimental, and genomic differences between intermediately pathogenic, highly pathogenic, and epidemic Streptococcus suis. J. Infect. Dis. 2009, 199, 97–107. [Google Scholar] [CrossRef]
- Tang, J.; Wang, C.; Feng, Y.; Yang, W.; Song, H.; Chen, Z.; Yu, H.; Pan, X.; Zhou, X.; Wang, H.; et al. Streptococcal toxic shock syndrome caused by Streptococcus suis serotype 2. PLoS Med. 2006, 3, e151. [Google Scholar]
- Segura, M. Streptococcus suis: An emerging human threat. J. Infect. Dis. 2009, 199, 4–6. [Google Scholar] [CrossRef]
- Lun, Z.R.; Wang, Q.P.; Chen, X.G.; Li, A.X.; Zhu, X.Q. Streptococcus suis: An emerging zoonotic pathogen. Lancet Infect. Dis. 2007, 7, 201–209. [Google Scholar] [CrossRef]
- Lachance, C.; Gottschalk, M.; Gerber, P.P.; Lemire, P.; Xu, J.; Segura, M. Exacerbated type II interferon response drives hypervirulence and toxic shock by an emergent epidemic strain of Streptococcus suis. Infect. Immun. 2013, 81, 1928–1939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lachance, C.; Segura, M.; Dominguez-Punaro, M.C.; Wojewodka, G.; De Sanctis, J.B.; Radzioch, D.; Gottschalk, M. Deregulated balance of omega-6 and omega-3 polyunsaturated fatty acids following infection by the zoonotic pathogen Streptococcus suis. Infect. Immun. 2014, 82, 1778–1785. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Zhao, J.; Lin, L.; Pan, S.; Fu, L.; Han, L.; Jin, M.; Zhou, R.; Zhang, A. Targeting TREM-1 signaling in the presence of antibiotics is effective against Streptococcal Toxic-Shock-Like Syndrome (STSLS) caused by Streptococcus suis. Front. Cell Infect. Microbiol. 2015, 5, 79. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Xu, L.; Lv, W.; Han, L.; Xiang, Y.; Fu, L.; Jin, M.; Zhou, R.; Chen, H.; Zhang, A. An NLRP3 inflammasome-triggered cytokine storm contributes to Streptococcal toxic shock-like syndrome (STSLS). PLoS Pathog. 2019, 15, e1007795. [Google Scholar] [CrossRef]
- Song, L.; Li, X.; Xiao, Y.; Huang, Y.; Jiang, Y.; Meng, G.; Ren, Z. Contribution of nlrp3 inflammasome activation mediated by suilysin to streptococcal toxic shock-like syndrome. Front. Microbiol. 2020, 11, 1788. [Google Scholar] [CrossRef]
- Shi, J.J.; Zhao, Y.; Wang, K.; Shi, X.Y.; Wang, Y.; Huang, H.W.; Zhuang, Y.H.; Cai, T.; Wang, F.C.; Shao, F. Cleavage of GSDMD by inflammatory caspases determines pyroptotic cell death. Nature 2015, 526, 660–665. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Z.B.; Ruan, J.B.; Pan, Y.D.; Magupalli, V.G.; Wu, H.; Lieberman, J. Inflammasome-activated gasdermin D causes pyroptosis by forming membrane pores. Nature 2016, 535, 153–158. [Google Scholar] [CrossRef]
- Ding, J.J.; Wang, K.; Liu, W.; She, Y.; Sun, Q.; Shi, J.J.; Sun, H.Z.; Wang, D.C.; Shao, F. Pore-forming activity and structural autoinhibition of the gasdermin family. Nature 2016, 535, 111–116. [Google Scholar] [CrossRef]
- Mulvihill, E.; Sborgi, L.; Mari, S.A.; Pfreundschuh, M.; Hiller, S.; Muller, D.J. Mechanism of membrane pore formation by human gasdermin-D. EMBO J. 2018, 37, e98321. [Google Scholar] [CrossRef]
- Chan, A.H.; Schroder, K. Inflammasome signaling and regulation of interleukin-1 family cytokines. J. Exp. Med. 2020, 217, e20190314. [Google Scholar] [CrossRef]
- Lavagna, A.; Auger, J.P.; Dumesnil, A.; Roy, D.; Girardin, S.E.; Gisch, N.; Segura, M.; Gottschalk, M. Interleukin-1 signaling induced by Streptococcus suis serotype 2 is strain-dependent and contributes to bacterial clearance and inflammation during systemic disease in a mouse model of infection. Vet. Res. 2019, 50, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, J.; Goodband, R.D.; Chengappa, M.M.; Nelssen, J.L.; Tokach, M.D.; McVey, D.S.; Blecha, F. Influence of interleukin-1 on neutrophil function and resistance to Streptococcus suis in neonatal pigs. J. Leukoc. Biol. 1994, 56, 88–94. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.K.; Kim, S.H.; Lewis, E.C.; Azam, T.; Reznikov, L.L.; Dinarello, C.A. Differences in signaling pathways by IL-1beta and IL-18. Proc. Natl. Acad. Sci. USA 2004, 101, 8815–8820. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.L.; Chae, J.J.; Park, Y.H.; De Nardo, D.; Stirzaker, R.A.; Ko, H.J.; Tye, H.; Cengia, L.; DiRago, L.; Metcalf, D.; et al. Aberrant actin depolymerization triggers the pyrin inflammasome and autoinflammatory disease that is dependent on IL-18, not IL-1 beta. J. Exp. Med. 2015, 212, 927–938. [Google Scholar] [CrossRef] [PubMed]
- Shultz, D.B.; Rani, M.R.; Fuller, J.D.; Ransohoff, R.M.; Stark, G.R. Roles of IKK-beta, IRF1, and p65 in the activation of chemokine genes by interferon-gamma. J. Interferon Cytokine Res. 2009, 29, 817–824. [Google Scholar] [CrossRef]
- Tilahun, A.Y.; Holz, M.; Wu, T.T.; David, C.S.; Rajagopalan, G. Interferon gamma-dependent intestinal pathology contributes to the lethality in bacterial superantigen-induced toxic shock syndrome. PLoS ONE 2011, 6, e16764. [Google Scholar] [CrossRef]
- Xu, L.; Lin, L.; Lu, X.; Xiao, P.; Liu, R.; Wu, M.; Jin, M.; Zhang, A. Acquiring high expression of suilysin enable non-epidemic Streptococcus suis to cause streptococcal toxic shock-like syndrome (STSLS) through NLRP3 inflammasome hyperactivation. Emerg. Microbes Infect. 2021, 10, 1309–1319. [Google Scholar] [CrossRef]
- Ye, C.; Zhu, X.; Jing, H.; Du, H.; Segura, M.; Zheng, H.; Kan, B.; Wang, L.; Bai, X.; Zhou, Y.; et al. Streptococcus suis sequence type 7 outbreak, Sichuan, China. Emerg. Infect. Dis. 2006, 12, 1203–1208. [Google Scholar] [CrossRef]
- Zhang, A.; Chen, B.; Yuan, Z.; Li, R.; Liu, C.; Zhou, H.; Chen, H.; Jin, M. HP0197 contributes to CPS synthesis and the virulence of Streptococcus suis via CcpA. PLoS ONE 2012, 7, e50987. [Google Scholar] [CrossRef]
- Ren, H.; Hua, Z.; Meng, J.; Molenaar, A.; Bi, Y.; Cheng, N.; Zheng, X. Generation of Acsl4 Gene Knockout Mouse Model by CRISPR/Cas9-Mediated Genome Engineering. Crit. Rev. Biomed. Eng. 2019, 47, 419–426. [Google Scholar] [CrossRef]
- Xu, L.; Lu, X.; Xiao, P.; Liu, R.; Xia, K.; Wu, M.; Jin, M.; Zhang, A. Interleukin-17A Contributed to the Damage of Blood-CNS Barriers During Streptococcus suis Meningitis. Mol. Neurobiol. 2022, 59, 2116–2128. [Google Scholar] [CrossRef] [PubMed]
- Dominguez-Punaro, M.C.; Segura, M.; Plante, M.M.; Lacouture, S.; Rivest, S.; Gottschalk, M. Streptococcus suis serotype 2, an important swine and human pathogen, induces strong systemic and cerebral inflammatory responses in a mouse model of infection. J. Immunol. 2007, 179, 1842–1854. [Google Scholar] [CrossRef]
- Zhu, H.; Huang, D.; Zhang, W.; Wu, Z.; Lu, Y.; Jia, H.; Wang, M.; Lu, C. The novel virulence-related gene stp of Streptococcus suis serotype 9 strain contributes to a significant reduction in mouse mortality. Microb. Pathog. 2011, 51, 442–453. [Google Scholar] [CrossRef] [PubMed]
- Xu, L.; Lu, X.; Xiao, P.; Liu, R.; Xia, K.L.; Wu, M.Z.; Jin, M.L.; Zhang, A.D. Interleukin-17A Contributes to Bacterial Clearance in a Mouse Model of Streptococcal Toxic Shock-Like Syndrome. Pathogens 2021, 10, 766. [Google Scholar] [CrossRef]
- Kuranaga, N.; Kinoshita, M.; Kawabata, T.; Habu, Y.; Shinomiya, N.; Seki, S. Interleukin-18 protects splenectomized mice from lethal Streptococcus pneumoniae sepsis independent of interferon-gamma by inducing IgM production. J. Infect. Dis. 2006, 194, 993–1002. [Google Scholar] [CrossRef]
- Paterson, G.K.; Blue, C.E.; Mitchell, T.J. Role of interleukin-18 in experimental infections with Streptococcus pneumoniae. J. Med. Microbiol. 2005, 54, 323–326. [Google Scholar] [CrossRef]
- Mancuso, G.; Midiri, A.; Beninati, C.; Zummo, S.; Biondo, C. Protective role of IL-18 in host defenses against group B Streptococcus. Eur. J. Clin. Microbiol. Infect. Dis. 2021, 40, 2657–2663. [Google Scholar] [CrossRef]
- Cusumano, V.; Midiri, A.; Cusumano, V.V.; Bellantoni, A.; De Sossi, G.; Teti, G.; Beninati, C.; Mancuso, G. Interleukin-18 is an essential element in host resistance to experimental group B streptococcal disease in neonates. Infect. Immun. 2004, 72, 295–300. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Confirmation of il18r1-/- mice. (A) DNA sequencing of the il18r1 gene in the il18r1-/- and il18r1+/+ mice. An 8bp DNA sequence was deleted in the il18r1-/- mice. (B) Genotyping by PCR. A 390 bp DNA fragment for KO and HET. A 405 bp DNA fragment for WT and HET. WT: wild-type mouse; HET: heterozygote mouse; KO: knockout mouse. Ctrl: negative control (H2O).
Figure 1.
Confirmation of il18r1-/- mice. (A) DNA sequencing of the il18r1 gene in the il18r1-/- and il18r1+/+ mice. An 8bp DNA sequence was deleted in the il18r1-/- mice. (B) Genotyping by PCR. A 390 bp DNA fragment for KO and HET. A 405 bp DNA fragment for WT and HET. WT: wild-type mouse; HET: heterozygote mouse; KO: knockout mouse. Ctrl: negative control (H2O).

Figure 2.
Knockout of il18r1 could decrease mortality during STSLS. The wild-type mice (il18r1+/+) and the il18r1-deficient mice (il18r1-/-) were infected (i.p.) with SC-19. (A) Survival of infected mice (n = 10). (B) Clinical symptom scores of infected mice; deceased mice were excluded (n = 10). il18r1+/+: wild type mouse. il18r1-/-: il18r1 knockout mouse.
Figure 2.
Knockout of il18r1 could decrease mortality during STSLS. The wild-type mice (il18r1+/+) and the il18r1-deficient mice (il18r1-/-) were infected (i.p.) with SC-19. (A) Survival of infected mice (n = 10). (B) Clinical symptom scores of infected mice; deceased mice were excluded (n = 10). il18r1+/+: wild type mouse. il18r1-/-: il18r1 knockout mouse.

Figure 3.
Knockout of il18r1 could alleviate the tissue damages during STSLS. H&E staining of tissue sections of mice at 6 h post-infection infected with S. suis. Necrosis in the liver: yellow arrow; congestion in the lung and spleen: red arrow; infiltration of inflammatory cells in the lung: green arrow. Scale bar indicates 50 μM. il18r1+/+: wild-type mouse group. il18r1–/–: il18r1 knockout mouse group.
Figure 3.
Knockout of il18r1 could alleviate the tissue damages during STSLS. H&E staining of tissue sections of mice at 6 h post-infection infected with S. suis. Necrosis in the liver: yellow arrow; congestion in the lung and spleen: red arrow; infiltration of inflammatory cells in the lung: green arrow. Scale bar indicates 50 μM. il18r1+/+: wild-type mouse group. il18r1–/–: il18r1 knockout mouse group.

Figure 4.
Knockout of il18r1 decreases serum inflammatory cytokine storm. The wild-type mice (il18r1+/+) and the il18r1-deficient mice (il18r1-/-) were infected (i.p.) with S. suis SC-19. (A) Cytokine levels in the blood at 6 h and 12 h post-infection were determined (n = 5). (B) The bacterial burdens in the liver, lung, blood, and brain at 6 h and 12 h post-infection were determined (n = 5). Each symbol represents the bacteria recovered from 1 mouse. Error bars represent the mean ± standard deviations. il18r1+/+: wild-type mouse group. il18r1-/-: il18r1 knockout mouse group.
Figure 4.
Knockout of il18r1 decreases serum inflammatory cytokine storm. The wild-type mice (il18r1+/+) and the il18r1-deficient mice (il18r1-/-) were infected (i.p.) with S. suis SC-19. (A) Cytokine levels in the blood at 6 h and 12 h post-infection were determined (n = 5). (B) The bacterial burdens in the liver, lung, blood, and brain at 6 h and 12 h post-infection were determined (n = 5). Each symbol represents the bacteria recovered from 1 mouse. Error bars represent the mean ± standard deviations. il18r1+/+: wild-type mouse group. il18r1-/-: il18r1 knockout mouse group.

Figure 5.
Scheme of the relationship of STSLS and IL-18. Previous study has illustrated that SLY was the essential and sufficient condition for NLRP3 inflammasome hyperactivation, causing cytokine storms and STSLS [14]. IFN-γ has been demonstrated to activate cytokine storms and STSLS [11]. Our study showed that IL-18, the downstream of NLPR3 signaling, was responsible for IFN-γ induction and STSLS development.
Figure 5.
Scheme of the relationship of STSLS and IL-18. Previous study has illustrated that SLY was the essential and sufficient condition for NLRP3 inflammasome hyperactivation, causing cytokine storms and STSLS [14]. IFN-γ has been demonstrated to activate cytokine storms and STSLS [11]. Our study showed that IL-18, the downstream of NLPR3 signaling, was responsible for IFN-γ induction and STSLS development.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xu, L.; Zeng, Y.; Gao, P.; Lu, X.; Xia, K.; Zhou, L.; Zhang, C.; Yi, C.; Zhang, A. IL-18 Signaling Is Essential for Causing Streptococcal Toxic Shock-like Syndrome (STSLS). Life 2022, 12, 1324. https://doi.org/10.3390/life12091324
AMA Style
Xu L, Zeng Y, Gao P, Lu X, Xia K, Zhou L, Zhang C, Yi C, Zhang A. IL-18 Signaling Is Essential for Causing Streptococcal Toxic Shock-like Syndrome (STSLS). Life. 2022; 12(9):1324. https://doi.org/10.3390/life12091324
Chicago/Turabian StyleXu, Lei, Yue Zeng, Peiying Gao, Xi Lu, Kunlong Xia, Liting Zhou, Chengfeng Zhang, Chenyang Yi, and Anding Zhang. 2022. "IL-18 Signaling Is Essential for Causing Streptococcal Toxic Shock-like Syndrome (STSLS)" Life 12, no. 9: 1324. https://doi.org/10.3390/life12091324
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.